

Herb Robert’s Gift against Human Diseases: Anticancer and Antimicrobial Activity of Geranium robertianum L.

 , , , ,
, , , ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Preparation of Extracts for Screening
2.3. Compound Identification
2.4. Preparation of Hexane and Ethyl Acetate Fractions
2.5. Cytotoxicity and Antiviral Activity
2.5.1. Evaluation of Cytotoxicity
2.5.2. Antiviral Activity
2.6. Antibacterial and Antifungal Activity
3. Results
3.1. Phytochemical Profile of G. robertianum Extracts
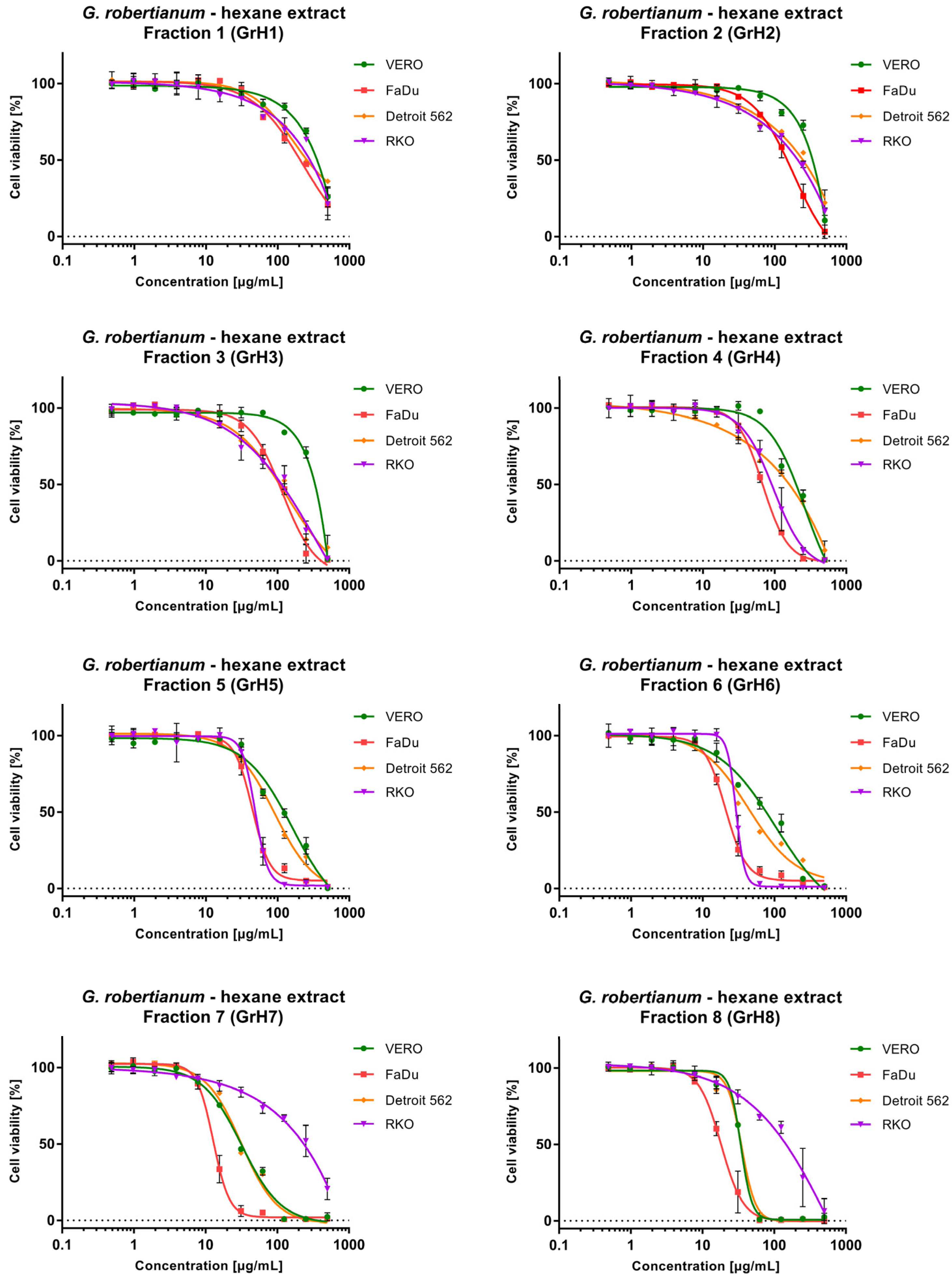
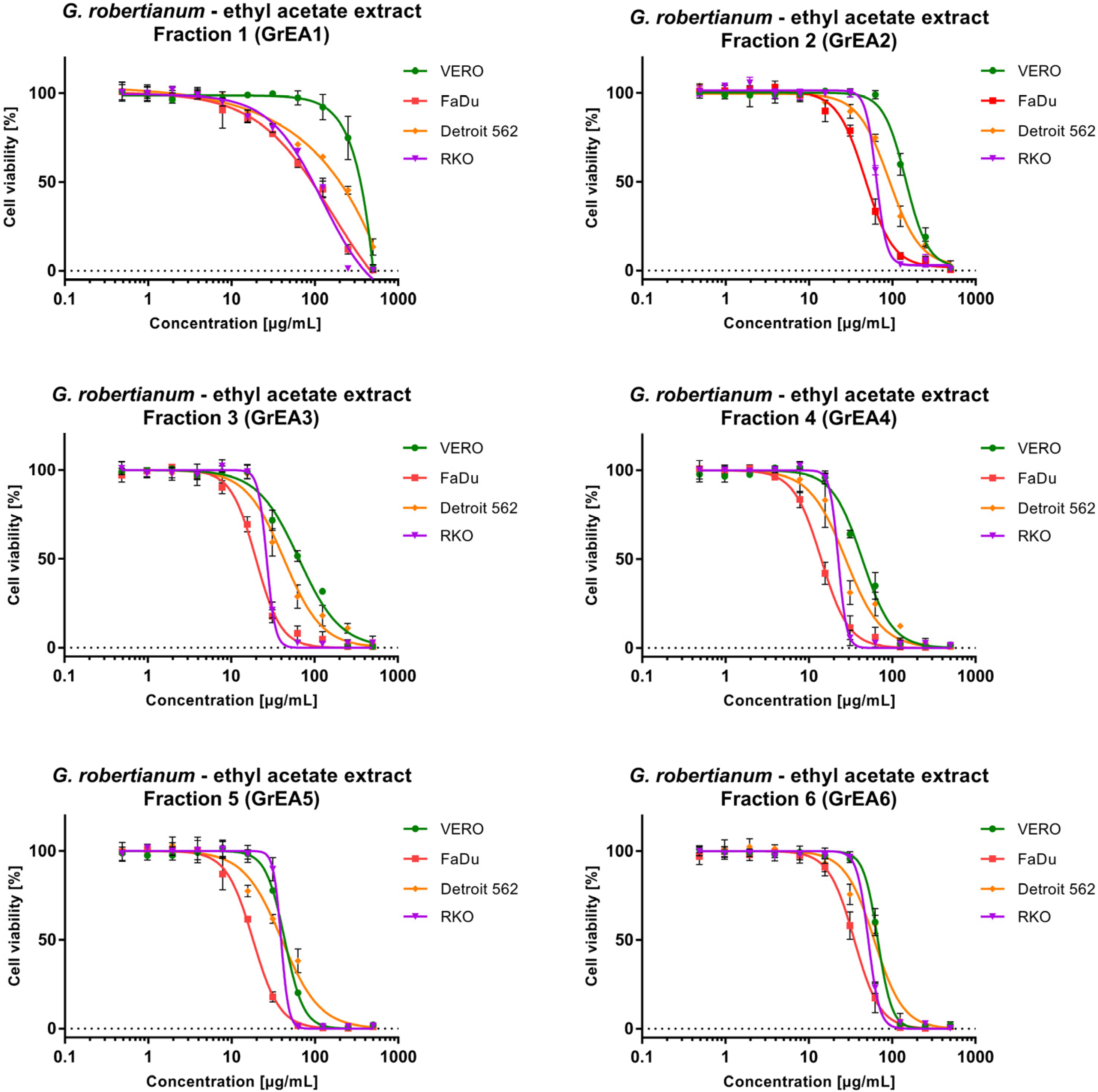
3.2. The Evaluation of Cytotoxicity towards Non-Cancerous and Cancer Cells
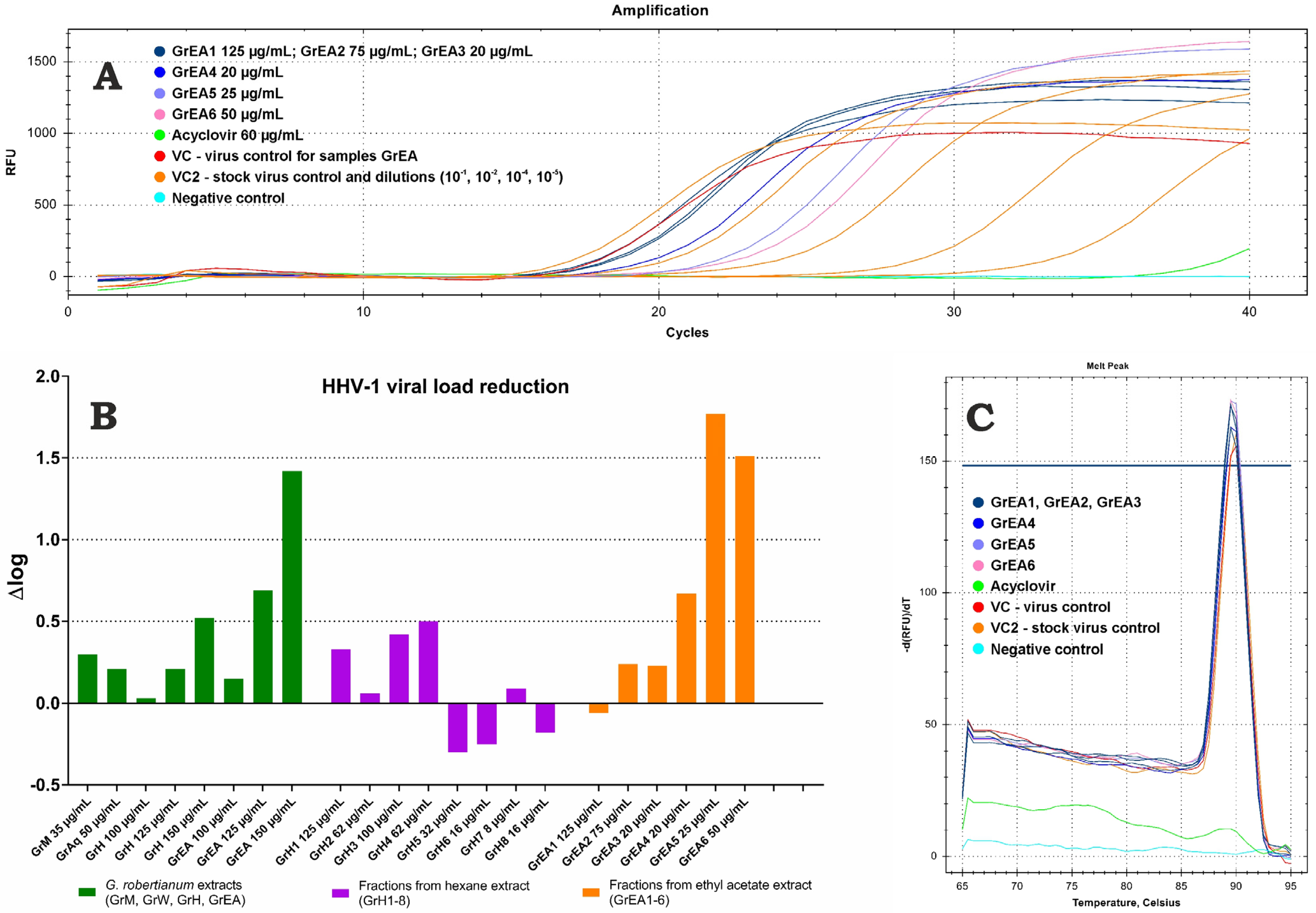
3.3. Assessment of Antiherpesviral Activity
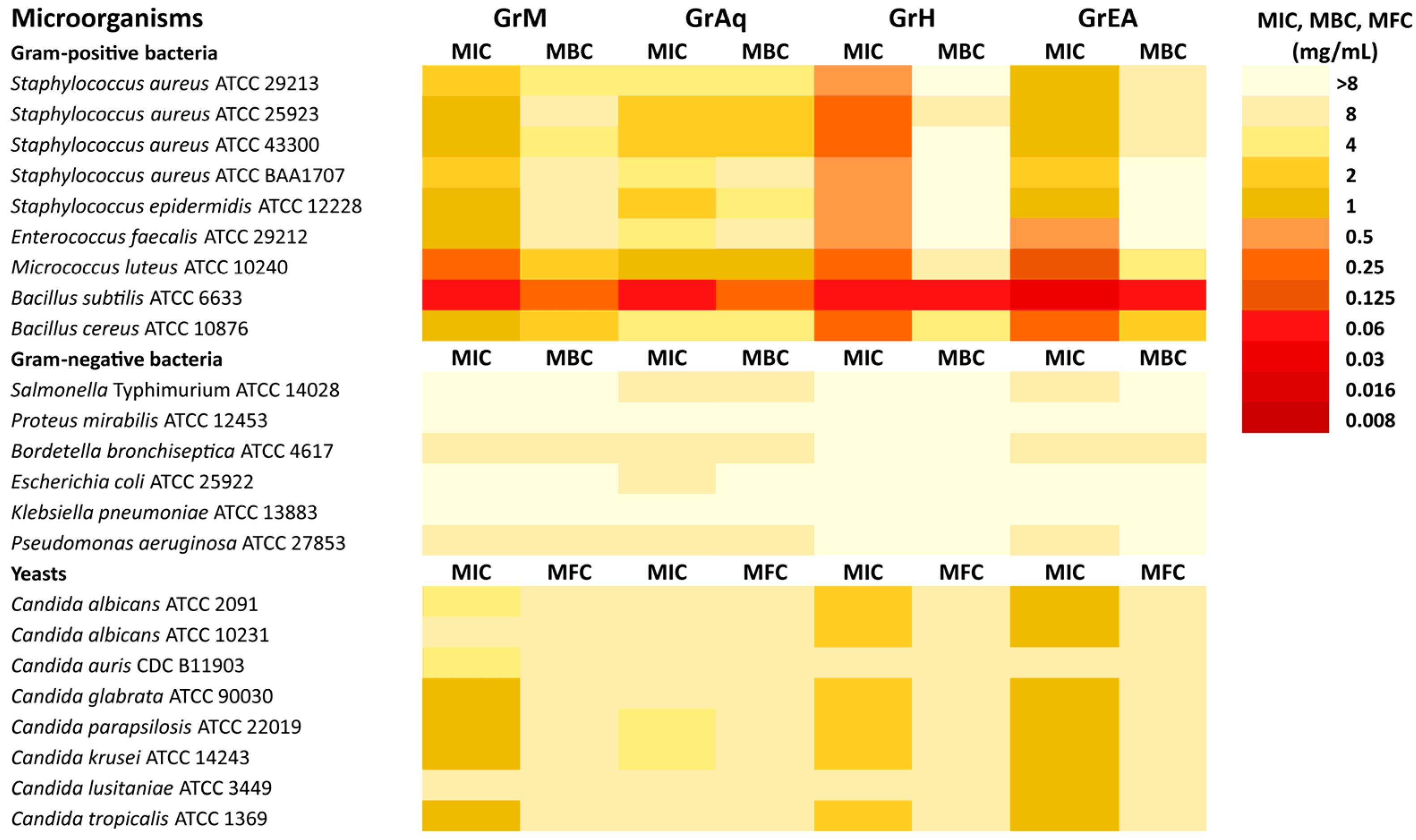
3.4. Antimicrobial Activity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Gruenwald, J.; Brendler, T.; Jaenicke, C. PDR for Herbal Medicines: Physicians’ Desk Reference for Herbal Medicines, 3rd ed.; Thomson PDR: Montvale, NJ, USA, 2004. [Google Scholar]
- Graça, V.C.; Ferreira, I.C.F.R.; Santos, P.F. Phytochemical composition and biological activities of Geranium robertianum L.: A review. Ind. Crops Prod. 2016, 87, 363–378. [Google Scholar] [CrossRef]
- Menković, N.; Šavikin, K.; Tasić, S.; Zdunić, G.; Stešević, D.; Milosavljević, S.; Vincek, D. Ethnobotanical study on traditional uses of wild medicinal plants in Prokletije Mountains (Montenegro). J. Ethnopharmacol. 2011, 133, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Graça, V.C.; Barros, L.; Calhelha, R.C.; Dias, M.I.; Ferreira, I.C.F.R.; Santos, P.F. Bio-guided fractionation of extracts of Geranium robertianum L.: Relationship between phenolic profile and biological activity. Ind. Crops Prod. 2017, 108, 543–552. [Google Scholar] [CrossRef]
- Ahmad, I.; Wilson, D.W. HSV-1 Cytoplasmic Envelopment and Egress. Int. J. Mol. Sci. 2020, 21, 5969. [Google Scholar] [CrossRef] [PubMed]
- van de Sand, L.; Bormann, M.; Schmitz, Y.; Heilingloh, C.S.; Witzke, O.; Krawczyk, A. Antiviral Active Compounds Derived from Natural Sources against Herpes Simplex Viruses. Viruses 2021, 13, 1386. [Google Scholar] [CrossRef] [PubMed]
- Serkedjieva, J.; Ivancheva, S. Antiherpes virus activity of extracts from the medicinal plant Geranium sanguineum L. J. Ethnopharmacol. 1998, 64, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Serkedjieva, J.; Hay, A.J. In vitro anti-influenza virus activity of a plant preparation from Geranium sanguineum L. Antivir. Res. 1998, 37, 121–130. [Google Scholar] [CrossRef]
- Li, J.; Huang, H.; Zhou, W.; Feng, M.; Zhou, P. Anti-hepatitis B Virus Activities of Geranium carolinianum L. Extracts and Identification of the Active Components. Biol. Pharm. Bull. 2008, 31, 743–747. [Google Scholar] [CrossRef]
- Świątek, Ł.; Sieniawska, E.; Sinan, K.I.; Maciejewska-Turska, M.; Boguszewska, A.; Polz-Dacewicz, M.; Senkardes, I.; Guler, G.O.; Bibi Sadeer, N.; Mahomoodally, M.F.; et al. LC-ESI-QTOF-MS/MS Analysis, Cytotoxic, Antiviral, Antioxidant, and Enzyme Inhibitory Properties of Four Extracts of Geranium pyrenaicum Burm. f.: A Good Gift from the Natural Treasure. Int. J. Mol. Sci. 2021, 22, 7621. [Google Scholar] [CrossRef]
- Graça, V.C.; Barros, L.; Calhelha, R.C.; Dias, M.I.; Carvalho, A.M.; Santos-Buelga, C.; Santos, P.F.; Ferreira, I.C. Chemical characterization and bioactive properties of aqueous and organic extracts of Geranium robertianum L. Food Funct. 2016, 7, 3807–3814. [Google Scholar] [CrossRef]
- Radulović, N.; Dekić, M.; Stojanović-Radić, Z. Chemical composition and antimicrobial activity of the volatile oils of Geranium sanguineum L. and G. robertianum L. (Geraniaceae). Med. Chem. Res. 2011, 21, 601–615. [Google Scholar] [CrossRef]
- EUCAST, European Committee for Antimicrobial Susceptibility Testing (EUCAST) of the European Society of Clinical Microbiology and Infectious Diseases (ESCMID). Determination of minimum inhibitory concentrations (MICs) of antibacterial agents by broth dilution. Clin. Microbiol. Infect. 2003, 6, 509–515. [Google Scholar] [CrossRef]
- Malm, A.; Grzegorczyk, A.; Biernasiuk, A.; Baj, T.; Rój, E.; Tyśkiewicz, K.; Dębczak, A.; Stolarski, M.J.; Krzyżaniak, M.; Olba-Zięty, E. Could Supercritical Extracts from the Aerial Parts of Helianthus salicifolius A. Dietr. and Helianthus tuberosus L. Be Regarded as Potential Raw Materials for Biocidal Purposes? Agriculture 2020, 11, 10. [Google Scholar] [CrossRef]
- Catarino, M.D.; Silva, A.M.S.; Cruz, M.T.; Cardoso, S.M. Antioxidant and anti-inflammatory activities of Geranium robertianum L. decoctions. Food Funct. 2017, 8, 3355–3365. [Google Scholar] [CrossRef]
- Sieniawska, E.; Świątek, Ł.; Sinan, K.I.; Zengin, G.; Boguszewska, A.; Polz-Dacewicz, M.; Bibi Sadeer, N.; Etienne, O.K.; Mahomoodally, M.F. Phytochemical Insights into Ficus sur Extracts and Their Biological Activity. Molecules 2022, 27, 1863. [Google Scholar] [CrossRef]
- Singh, A.; Bajpai, V.; Kumar, S.; Sharma, K.R.; Kumar, B. Profiling of Gallic and Ellagic Acid Derivatives in Different Plant Parts of Terminalia arjuna by HPLC-ESI-QTOF-MS/MS. Nat. Prod. Commun. 2016, 11, 239–244. [Google Scholar] [CrossRef]
- Geran, R.S.; Greenberg, N.H.; Macdonald, M.M.; Schumacher, A.M.; Abbott, B.J. Protocols for Screening Chemical Agents and Natural Products against Animal Tumors and Other Biological Systems. Cancer Chemother. Rep. 1972, 13, 17–27. [Google Scholar]
- Łaska, G.; Sieniawska, E.; Świątek, Ł.; Zjawiony, J.; Khan, S.; Boguszewska, A.; Stocki, M.; Angielczyk, M.; Polz-Dacewicz, M. Phytochemistry and biological activities of Polemonium caeruleum L. Phytochem. Lett. 2019, 30, 314–323. [Google Scholar] [CrossRef]
- Mirandeli Bautista, Á.; de Lúcio, J.A.G.; Nancy Vargas, M.; Claudia Velázquez, G.; De la O Arciniega, M.; Georgina Almaguer, V. Geranium Species as Antioxidants. In Oxidative Stress and Chronic Degenerative Diseases; José, A.M.-G., Ed.; IntechOpen: Rijeka, Croatia, 2013; Chapter 5. [Google Scholar]
- Bate-Smith, E.C. The phenolic constituents of plants and their taxonomic significance. I. Dicotyledons. J. Linn. Soc. Lond. Bot. 1962, 58, 95–173. [Google Scholar] [CrossRef]
- Verma, S.; Singh, A.; Mishra, A. Gallic acid: Molecular rival of cancer. Environ. Toxicol. Pharmacol. 2013, 35, 473–485. [Google Scholar] [CrossRef]
- Zhang, H.M.; Zhao, L.; Li, H.; Xu, H.; Chen, W.W.; Tao, L. Research progress on the anticarcinogenic actions and mechanisms of ellagic acid. Cancer Biol. Med. 2014, 11, 92–100. [Google Scholar] [PubMed]
- Chen, Y.; Gong, S.; Liu, Y.; Cao, X.; Zhao, M.; Xiao, J.; Feng, C. Geraniin inhibits cell growth and promoted autophagy-mediated cell death in the nasopharyngeal cancer C666-1 cells. Saudi J. Biol. Sci. 2022, 29, 168–174. [Google Scholar] [CrossRef] [PubMed]
- Kopustinskiene, D.M.; Jakstas, V.; Savickas, A.; Bernatoniene, J. Flavonoids as Anticancer Agents. Nutrients 2020, 12, 457. [Google Scholar] [CrossRef] [PubMed]
- Paun, G.; Neagu, E.; Litescu, C.; Rotinberg, P.; Radu, L. Membrane processes application on the Symphytum officinale and Geranium robertianum extracts concentration to obtain high antioxidative activity compounds. J. Serb. Chem. Soc. 2012, 77, 1191–1203. [Google Scholar] [CrossRef]
- Gorphe, P. A comprehensive review of Hep-2 cell line in translational research for laryngeal cancer. Am. J. Cancer Res. 2019, 9, 644–649. [Google Scholar]
- Yang, C.-M.; Cheng, H.-Y.; Lin, T.-C.; Chiang, L.-C.; Lin, C.-C. The in vitro activity of geraniin and 1,3,4,6-tetra-O-galloyl-β-d-glucose isolated from Phyllanthus urinaria against herpes simplex virus type 1 and type 2 infection. J. Ethnopharmacol. 2007, 110, 555–558. [Google Scholar] [CrossRef]
- Park, S.W.; Kwon, M.J.; Yoo, J.Y.; Choi, H.-J.; Ahn, Y.-J. Antiviral activity and possible mode of action of ellagic acid identified in Lagerstroemia speciosa leaves toward human rhinoviruses. BMC Complement. Altern. Med. 2014, 14, 171. [Google Scholar] [CrossRef]
- Kullappan, M.; Benedict, B.A.; Rajajagadeesan, A.; Baskaran, P.; Periadurai, N.D.; Ambrose, J.M.; Gandhamaneni, S.H.; Nakkella, A.K.; Agarwal, A.; Veeraraghavan, V.P.; et al. Ellagic Acid as a Potential Inhibitor against the Nonstructural Protein NS3 Helicase of Zika Virus: A Molecular Modelling Study. BioMed Res. Int. 2022, 2022, 2044577. [Google Scholar] [CrossRef]
- Civra, A.; Acquadro, S.; Cagliero, C.; Marengo, A.; Rittà, M.; Francese, R.; Sanna, C.; Bertea, C.; Sgorbini, B.; Lembo, D.; et al. Punica granatum Leaf Ethanolic Extract and Ellagic Acid as Inhibitors of Zika Virus Infection. Planta Med. 2020, 86, 1363–1374. [Google Scholar]
- Cui, Q.; Du, R.; Anantpadma, M.; Schafer, A.; Hou, L.; Tian, J.; Davey, R.; Cheng, H.; Rong, L. Identification of Ellagic Acid from Plant Rhodiola rosea L. as an Anti-Ebola Virus Entry Inhibitor. Viruses 2018, 10, 152. [Google Scholar] [CrossRef]
- El-Aguel, A.; Pennisi, R.; Smeriglio, A.; Kallel, I.; Tamburello, M.P.; D’Arrigo, M.; Barreca, D.; Gargouri, A.; Trombetta, D.; Mandalari, G.; et al. Punica granatum Peel and Leaf Extracts as Promising Strategies for HSV-1 Treatment. Viruses 2022, 14, 2639. [Google Scholar] [CrossRef] [PubMed]
- Di Petrillo, A.; Orrù, G.; Fais, A.; Fantini, M.C. Quercetin and its derivates as antiviral potentials: A comprehensive review. Phytother. Res. 2021, 36, 266–278. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Kim, N.-E.; Park, B.J.; Kwon, H.C.; Song, Y.-J. Kaempferol Interferes with Varicella-Zoster Virus Replication in Human Foreskin Fibroblasts. Pharmaceuticals 2022, 15, 1582. [Google Scholar] [CrossRef] [PubMed]
- Mitrocotsa, D.; Mitaku, S.; Axarlis, S.; Harvala, C.; Malamas, M. Evaluation of the Antiviral Activity of Kaempferol and its Glycosides Against Human Cytomegalovirus. Planta Med. 2000, 66, 377–379. [Google Scholar] [CrossRef]
- Zhu, L.; Wang, P.; Yuan, W.; Zhu, G. Kaempferol inhibited bovine herpesvirus 1 replication and LPS-induced inflammatory response. Acta Virol. 2018, 62, 220–225. [Google Scholar] [CrossRef]
- Poehlmann, S.; Zhang, T.; Wu, Z.; Du, J.; Hu, Y.; Liu, L.; Yang, F.; Jin, Q. Anti- Japanese-Encephalitis-Viral Effects of Kaempferol and Daidzin and Their RNA-Binding Characteristics. PLoS ONE 2012, 7, e30259. [Google Scholar]
- Behbahani, M.; Sayedipour, S.; Pourazar, A.; Shanehsazzadeh, M. In vitro anti-HIV-1 activities of kaempferol and kaempferol-7-O-glucoside isolated from Securigera securidaca. Res. Pharm. Sci. 2014, 9, 463–469. [Google Scholar]
- Walasek-Janusz, M.; Grzegorczyk, A.; Zalewski, D.; Malm, A.; Gajcy, S.; Gruszecki, R. Variation in the Antimicrobial Activity of Essential Oils from Cultivars of Lavandula angustifolia and L. × intermedia. Agronomy 2022, 12, 2955. [Google Scholar] [CrossRef]
- Lima, I.d.S. Estudos de Metabolismo In Vitro de Extractos Aquosos de São Roberto, Geranium robertianum: Aplicações Terapêuticas na Doença de Alzheimer. Master’s Thesis, Universidade de Lisboa, Lisboa, Portugal, 2009. [Google Scholar]
- Tosun, F.; Akyüz Kızılay, Ç.; Şener, B.; Vural, M. The evaluation of plants from Turkey for in vitro antimycobacterial activity. Pharm. Biol. 2005, 43, 58–63. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extract | VERO | FaDu | Detroit 562 | RKO | |||
|---|---|---|---|---|---|---|---|
| CC50 | CC50 | SI | CC50 | SI | CC50 | SI | |
| Methanolic | 187.17 ± 28.32 | 115.9 ± 11.33 | 1.61 | 188.65 ± 10.68 | 0.99 | 180.57 ± 16.80 | 1.04 |
| Aqueous | 313.17 ± 35.3 | 145.83 ± 13.46 | 2.15 | 381.1 ± 8.49 | 0.82 | 284.93 ± 10.15 | 1.1 |
| Hexane | 286.97 ± 29.39 | 109.23 ± 8.62 | 2.63 | 117.57 ± 9.43 | 2.44 | 98.91 ± 3.61 | 2.9 |
| Ethyl acetate | 278.13 ± 19.08 | 63.36 ± 8.13 | 4.39 | 137.51 ± 9.88 | 2.02 | 63.63 ± 8.13 | 4.37 |
| Fractions | VERO | FaDu | Detroit 562 | RKO | |||
|---|---|---|---|---|---|---|---|
| CC50 | CC50 | SI | CC50 | SI | CC50 | SI | |
| GrH1 | 330.05 ± 26.23 | 203.95 ± 23.41 | 1.62 | 273.65 ± 20.58 | 1.21 | 238.5 ± 22.97 | 1.38 |
| GrH2 | 302.5 ± 3.25 | 142.55 ± 22.27 | 2.12 | 224.85 ± 20.44 | 1.35 | 168.62 ± 11.39 | 1.79 |
| GrH3 | 288.2 ± 5.94 | 101.9 ± 1.13 | 2.83 | 102.69 ± 4.55 | 2.81 | 99.52 ± 6.68 | 2.90 |
| GrH4 | 180.15 ± 6.72 | 67.62 ± 2.84 | 2.66 | 129.85 ± 6.86 | 1.39 | 89.68 ± 4.52 | 2.01 |
| GrH5 | 113.3 ± 8.49 | 46.25 ± 3.92 | 2.45 | 88.71 ± 4.74 | 1.28 | 49.16 ± 1.46 | 2.30 |
| GrH6 | 70.88 ± 3.77 | 22.04 ± 1.82 | 3.22 | 48.71 ± 1.36 | 1.46 | 29.59 ± 0.99 | 2.40 |
| GrH7 | 30.82 ± 0.31 | 13.35 ± 1.11 | 2.31 | 31.87 ± 0.25 | 0.97 | 187.0 ± 14.9 | 0.16 |
| GrH8 | 34.15 ± 0.14 | 18.51 ± 2.52 | 1.85 | 34.61 ± 0.32 | 0.99 | 125.10 ± 9.43 | 0.27 |
| GrEA1 | 289.07 ± 16.05 | 82.69 ± 1.94 | 3.50 | 168.80 ± 7.5 | 1.71 | 89.67 ± 5.42 | 3.22 |
| GrEA2 | 147.87 ± 4.66 | 49.02 ± 4.54 | 3.02 | 93.78 ± 5.02 | 1.58 | 65.6 ± 1.07 | 2.25 |
| GrEA3 | 65.01 ± 0.62 | 19.86 ± 0.73 | 3.27 | 45.06 ± 6.05 | 1.44 | 26.73 ± 1.28 | 2.43 |
| GrEA4 | 43.83 ± 3.23 | 14.22 ± 1.43 | 3.08 | 27.98 ± 6.50 | 1.57 | 22.55 ± 0.61 | 1.94 |
| GrEA5 | 43.46 ± 0.29 | 17.89 ± 0.68 | 2.43 | 40.23 ± 3.44 | 1.08 | 39.28 ± 2.09 | 1.11 |
| GrEA6 | 67.29 ± 3.54 | 35.48 ± 4.89 | 1.90 | 61.55 ± 6.33 | 1.09 | 51.86 ± 1.15 | 1.30 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Świątek, Ł.; Wasilewska, I.; Boguszewska, A.; Grzegorczyk, A.; Rezmer, J.; Rajtar, B.; Polz-Dacewicz, M.; Sieniawska, E. Herb Robert’s Gift against Human Diseases: Anticancer and Antimicrobial Activity of Geranium robertianum L. Pharmaceutics 2023, 15, 1561. https://doi.org/10.3390/pharmaceutics15051561
Świątek Ł, Wasilewska I, Boguszewska A, Grzegorczyk A, Rezmer J, Rajtar B, Polz-Dacewicz M, Sieniawska E. Herb Robert’s Gift against Human Diseases: Anticancer and Antimicrobial Activity of Geranium robertianum L. Pharmaceutics. 2023; 15(5):1561. https://doi.org/10.3390/pharmaceutics15051561
Chicago/Turabian StyleŚwiątek, Łukasz, Inga Wasilewska, Anastazja Boguszewska, Agnieszka Grzegorczyk, Jakub Rezmer, Barbara Rajtar, Małgorzata Polz-Dacewicz, and Elwira Sieniawska. 2023. "Herb Robert’s Gift against Human Diseases: Anticancer and Antimicrobial Activity of Geranium robertianum L." Pharmaceutics 15, no. 5: 1561. https://doi.org/10.3390/pharmaceutics15051561
APA StyleŚwiątek, Ł., Wasilewska, I., Boguszewska, A., Grzegorczyk, A., Rezmer, J., Rajtar, B., Polz-Dacewicz, M., & Sieniawska, E. (2023). Herb Robert’s Gift against Human Diseases: Anticancer and Antimicrobial Activity of Geranium robertianum L. Pharmaceutics, 15(5), 1561. https://doi.org/10.3390/pharmaceutics15051561