
Non-Antibiotic Compounds Synergistically Kill Chronic Wound-Associated Bacteria and Disrupt Their Biofilms
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
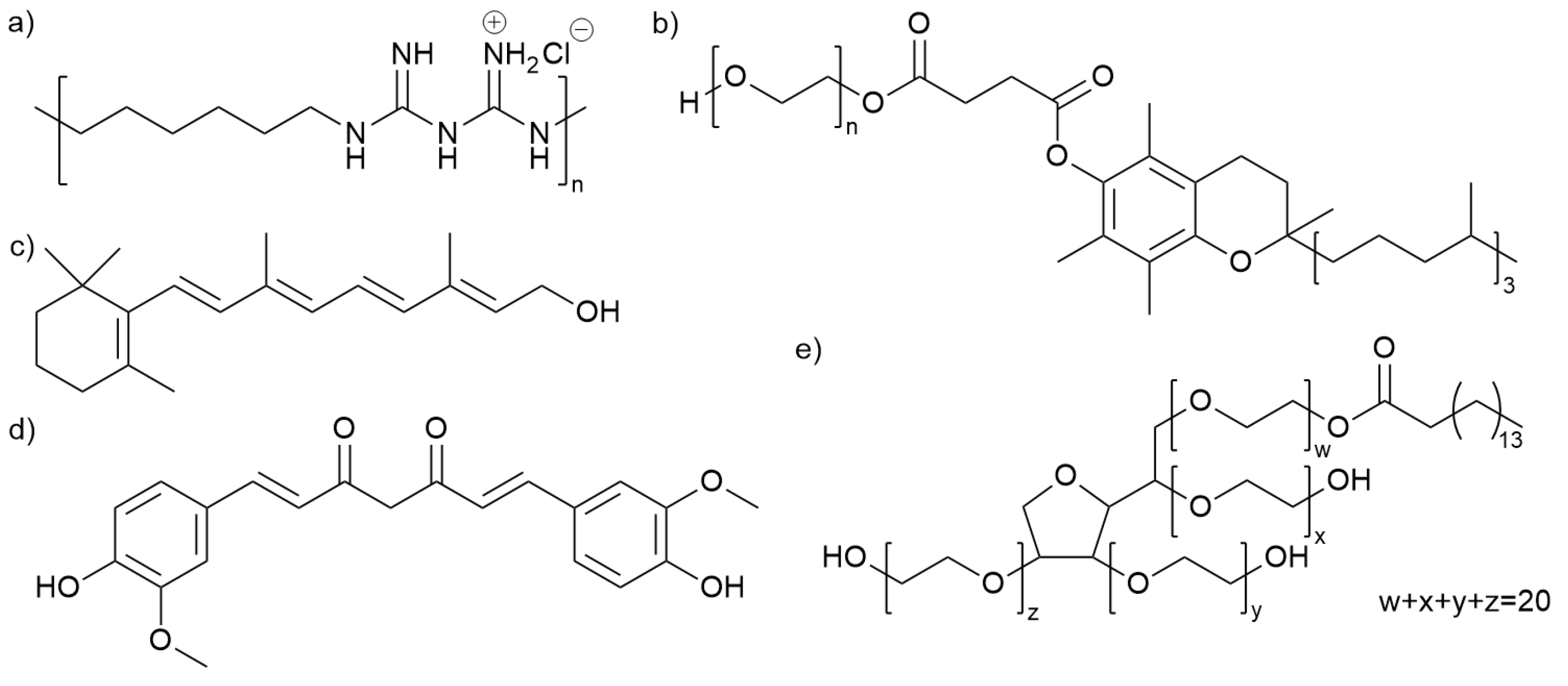
2.1. Chemicals
2.2. Bacterial Isolates and Growth Conditions
2.3. Minimum Inhibitory Concentration (MIC)
2.4. Minimum Bactericidal Concentration (MBC)
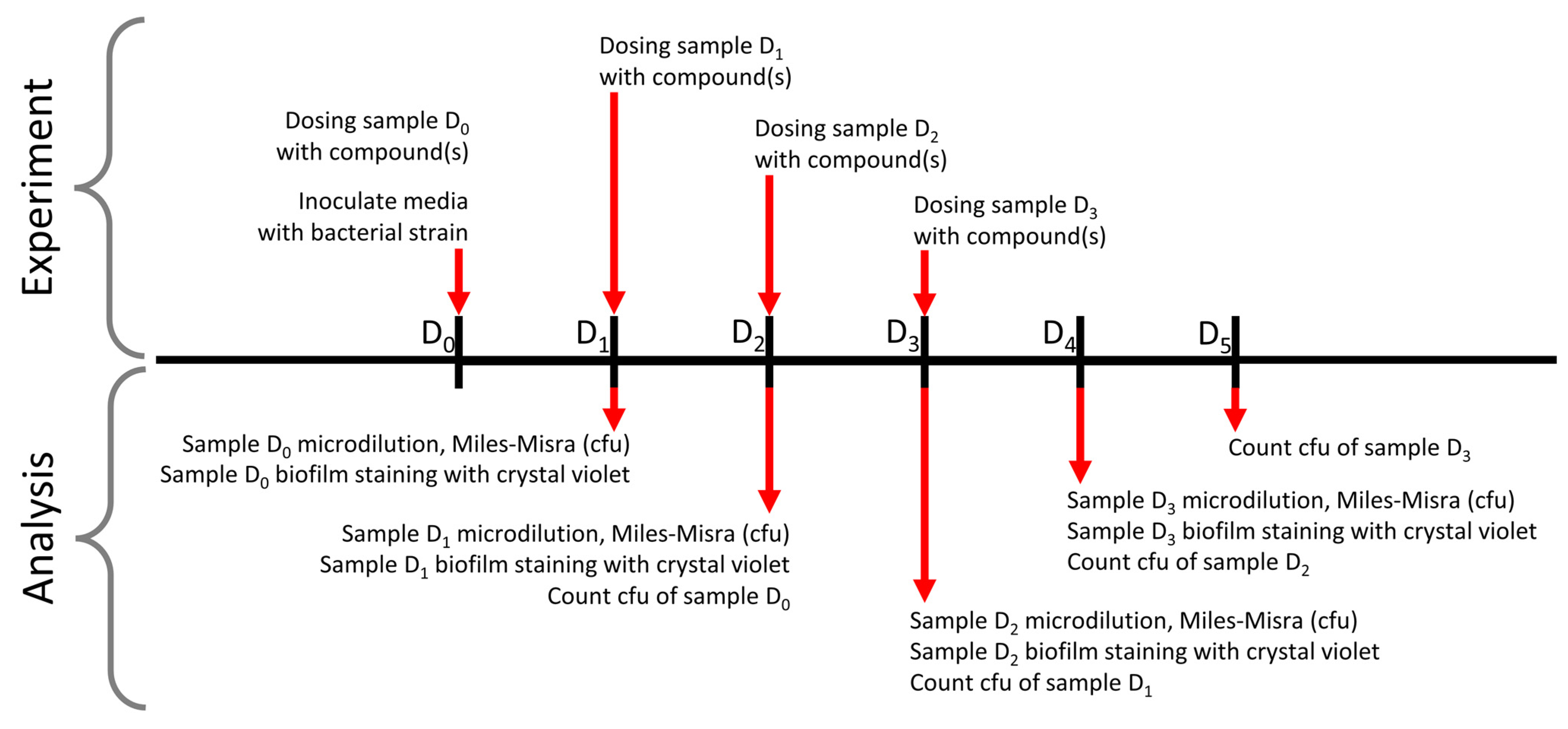
2.5. Bacterial Kill Curves
2.6. Crystal Violet Biofilm Clearance Assays
2.7. Biofilm Bacterial Survival Assays
3. Results
3.1. Antibacterial Activity of Individual Compounds
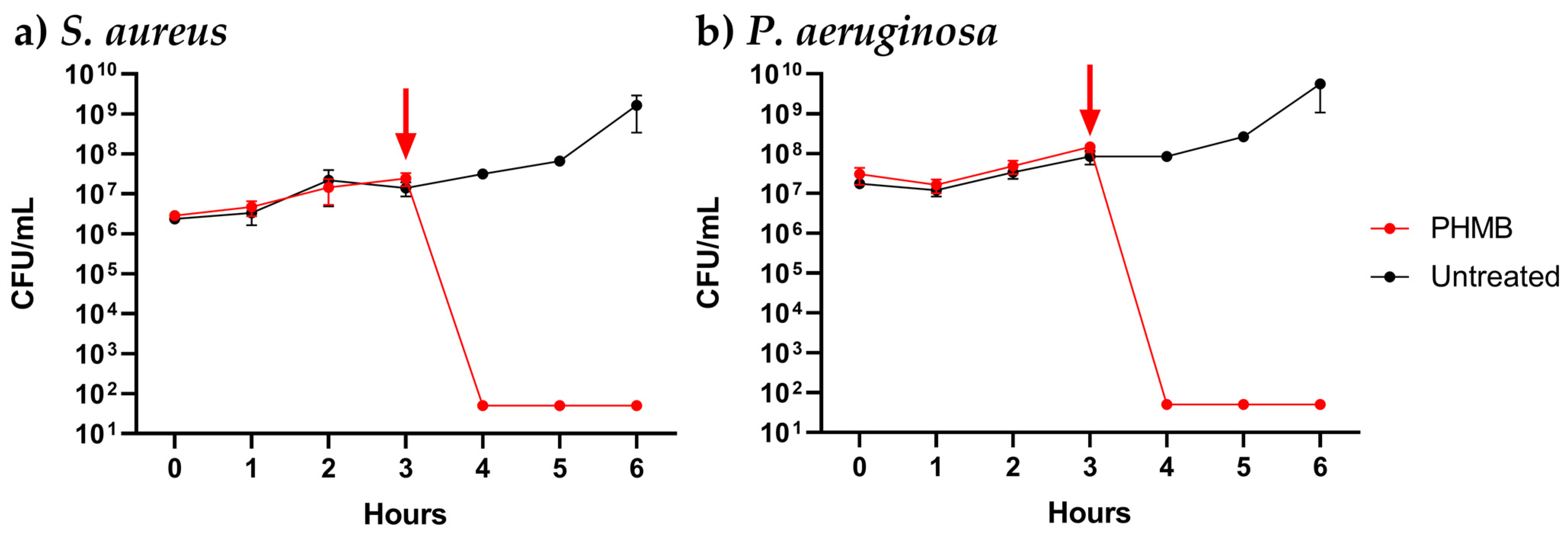
3.2. Dynamics of PHMB Antimicrobial Activity
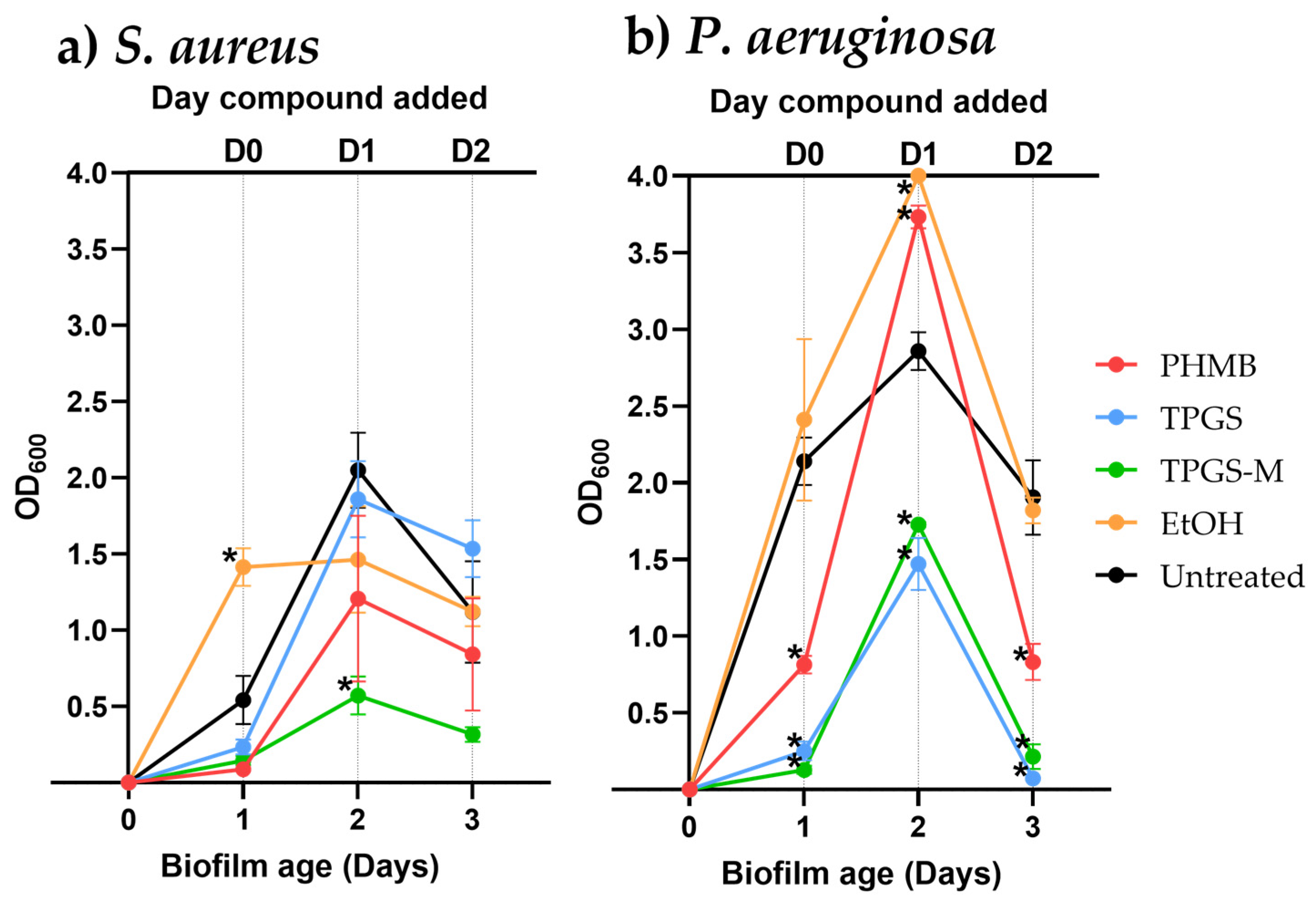
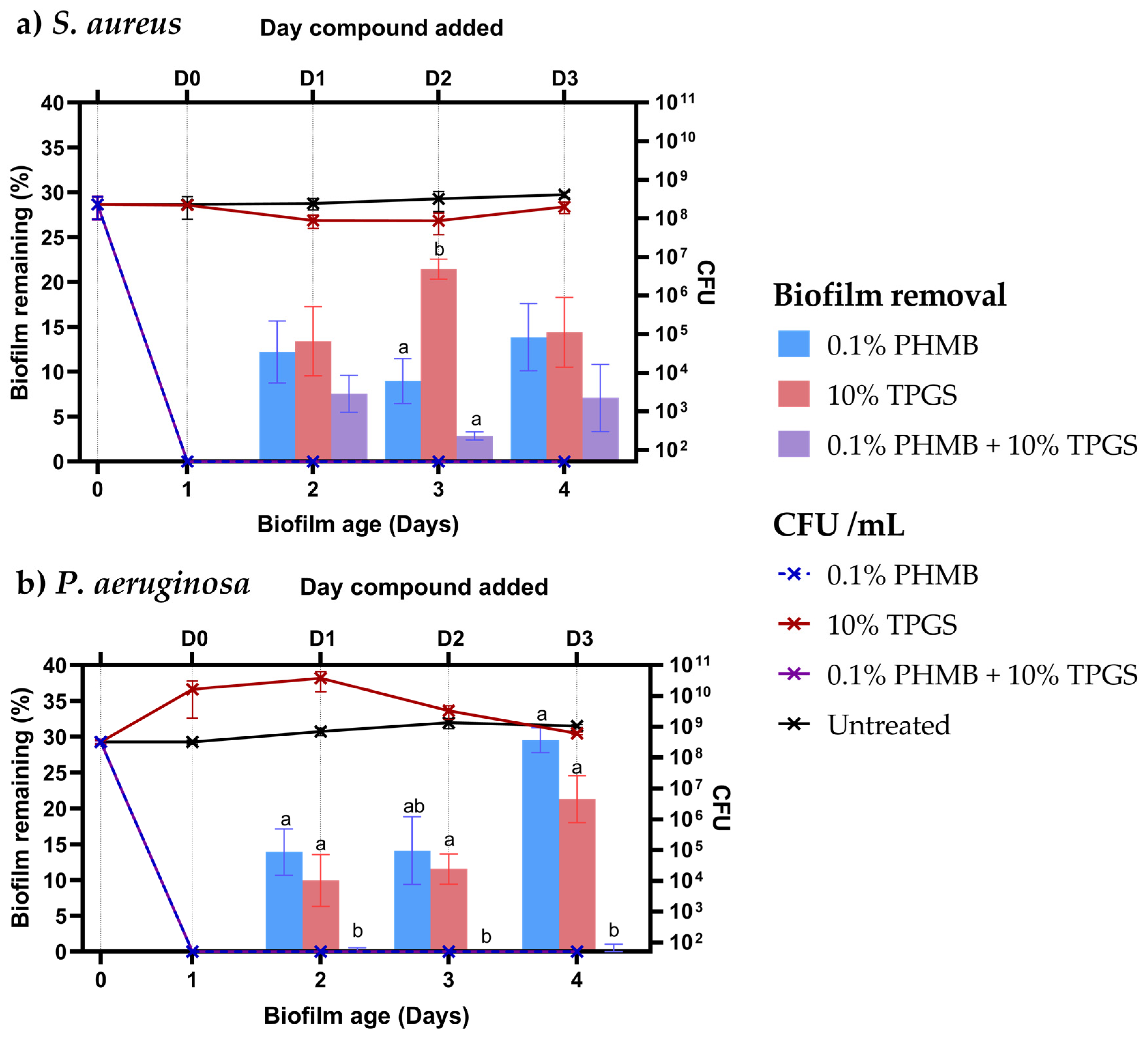
3.3. Biofilm Removal
3.4. Compounds in Combination
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jarbrink, K.; Ni, G.; Sonnergren, H.; Schmidtchen, A.; Pang, C.; Bajpai, R.; Car, J. Prevalence and incidence of chronic wounds and related complications: A protocol for a systematic review. Syst. Rev. 2016, 5, 152. [Google Scholar] [CrossRef] [PubMed]
- Sen, C.K. Human Wounds and Its Burden: An Updated Compendium of Estimates. Adv. Wound Care 2019, 8, 39–48. [Google Scholar] [CrossRef]
- Frykberg, R.G.; Banks, J. Challenges in the Treatment of Chronic Wounds. Adv. Wound Care 2015, 4, 560–582. [Google Scholar] [CrossRef]
- United Nations, Department of Economic and Social Affairs, Population Division. World Population Ageing 2019: Highlights (ST/ESA/SER.A/430); United Nations: New York, NY, USA, 2019. [Google Scholar]
- Percival, S.L.; Hill, K.E.; Williams, D.W.; Hooper, S.J.; Thomas, D.W.; Costerton, J.W. A review of the scientific evidence for biofilms in wounds. Wound Repair Regen. 2012, 20, 647–657. [Google Scholar] [CrossRef]
- Serra, R.; Grande, R.; Butrico, L.; Rossi, A.; Settimio, U.F.; Caroleo, B.; Amato, B.; Gallelli, L.; de Franciscis, S. Chronic wound infections: The role of Pseudomonas aeruginosa and Staphylococcus aureus. Expert Rev. Anti-Infect. Ther. 2015, 13, 605–613. [Google Scholar] [CrossRef]
- Flemming, H.C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef]
- Attinger, C.; Wolcott, R. Clinically Addressing Biofilm in Chronic Wounds. Adv. Wound Care 2012, 1, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Stoodley, P.; Sauer, K.; Davies, D.G.; Costerton, J.W. Biofilms as Complex Differentiated Communities. Annu. Rev. Microbiol. 2002, 56, 187–209. [Google Scholar] [CrossRef] [PubMed]
- Sauer, K.; Stoodley, P.; Goeres, D.M.; Hall-Stoodley, L.; Burmølle, M.; Stewart, P.S.; Bjarnsholt, T. The biofilm life cycle: Expanding the conceptual model of biofilm formation. Nat. Rev. Microbiol. 2022, 20, 608–620. [Google Scholar] [CrossRef]
- Hu, Y.; Ruan, X.; Lv, X.; Xu, Y.; Wang, W.; Cai, Y.; Ding, M.; Dong, H.; Shao, J.; Yang, D.; et al. Biofilm microenvironment-responsive nanoparticles for the treatment of bacterial infection. Nano Today 2022, 46, 101602. [Google Scholar] [CrossRef]
- Zhao, G.; Usui, M.L.; Lippman, S.I.; James, G.A.; Stewart, P.S.; Fleckman, P.; Olerud, J.E. Biofilms and Inflammation in Chronic Wounds. Adv. Wound Care 2013, 2, 389–399. [Google Scholar] [CrossRef]
- Costerton, J.W.; Stewart, P.S.; Greenberg, E.P. Bacterial Biofilms: A Common Cause of Persistent Infections. Science 1999, 284, 1318–1322. [Google Scholar] [CrossRef] [PubMed]
- Ammons, M.C. Anti-biofilm strategies and the need for innovations in wound care. Recent Pat. Antiinfect. Drug Discov. 2010, 5, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Lipsky, B.A.; Dryden, M.; Gottrup, F.; Nathwani, D.; Seaton, R.A.; Stryja, J. Antimicrobial stewardship in wound care: A Position Paper from the British Society for Antimicrobial Chemotherapy and European Wound Management Association. J. Antimicrob. Chemother. 2016, 71, 3026–3035. [Google Scholar] [CrossRef] [PubMed]
- Wolcott, R.D.; Rumbaugh, K.P.; James, G.; Schultz, G.; Phillips, P.; Yang, Q.; Watters, C.; Stewart, P.S.; Dowd, S.E. Biofilm maturity studies indicate sharp debridement opens a time- dependent therapeutic window. J. Wound Care 2010, 19, 320–328. [Google Scholar] [CrossRef]
- Wise, R.; Hart, T.; Cars, O.; Streulens, M.; Helmuth, R.; Huovinen, P.; Sprenger, M. Antimicrobial resistance. BMJ 1998, 317, 609–610. [Google Scholar] [CrossRef]
- Hoang, T.p.n.; Ghori, M.U.; Ousey, K.J.; Conway, B.R. Current and advanced therapies for chronic wound infection: A review. Pharm. J. 2022, 309, 1–17. [Google Scholar]
- Chindera, K.; Mahato, M.; Sharma, A.K.; Horsley, H.; Kloc-Muniak, K.; Kamaruzzaman, N.F.; Kumar, S.; McFarlane, A.; Stach, J.; Bentin, T.; et al. The antimicrobial polymer PHMB enters cells and selectively condenses bacterial chromosomes. Sci. Rep. 2016, 6, 23121. [Google Scholar] [CrossRef]
- Gray, D.; Barrett, S.; Battacharyya, M.; Butcher, M.; Enoch, S.; Fumerola, S.; Stephen-Haynes, J.; Edwards-Jones, V.; Leaper, D.; Strohal, R.; et al. PHMB and its potential contribution to wound management. Wounds UK 2010, 6, 40–46. [Google Scholar]
- Moore, K.; Gray, D. Using PHMB antimicrobial to prevent wound infection. Wound UK 2007, 3, 96. [Google Scholar]
- Phillips, P.L.; Yang, Q.; Schultz, G.S. The effect of negative pressure wound therapy with periodic instillation using antimicrobial solutions on Pseudomonas aeruginosa biofilm on porcine skin explants. Int. Wound J. 2013, 10 (Suppl. S1), 48–55. [Google Scholar] [CrossRef]
- Kamaruzzaman, N.F.; Chong, S.Q.Y.; Edmondson-Brown, K.M.; Ntow-Boahene, W.; Bardiau, M.; Good, L. Bactericidal and Anti-biofilm Effects of Polyhexamethylene Biguanide in Models of Intracellular and Biofilm of Staphylococcus aureus Isolated from Bovine Mastitis. Front. Microbiol. 2017, 8, 1518. [Google Scholar] [CrossRef] [PubMed]
- Percival, S.L.; Mayer, D.; Kirsner, R.S.; Schultz, G.; Weir, D.; Roy, S.; Alavi, A.; Romanelli, M. Surfactants: Role in biofilm management and cellular behaviour. Int. Wound J. 2019, 16, 753–760. [Google Scholar] [CrossRef] [PubMed]
- Toutain-Kidd, C.M.; Kadivar, S.C.; Bramante, C.T.; Bobin, S.A.; Zegans, M.E. Polysorbate 80 inhibition of Pseudomonas aeruginosa biofilm formation and its cleavage by the secreted lipase LipA. Antimicrob. Agents Chemother. 2009, 53, 136–145. [Google Scholar] [CrossRef] [PubMed]
- Babickaite, L.; Grigonis, A.; Ramanauskiene, K.; Matusevicius, A.P.; Zamokas, G.; Daunoras, G.; Ivaskiene, M. Therapeutic activity of chlorhexidine-poloxamer antiseptic gel on wound healing in rats: A preclinical study. Pol. J. Vet. Sci. 2018, 21, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Luo, J.; Tan, S.; Otieno, B.O.; Zhang, Z. The applications of Vitamin E TPGS in drug delivery. Eur. J. Pharm. Sci. 2013, 49, 175–186. [Google Scholar] [CrossRef]
- Tavares Luiz, M.; Delello Di Filippo, L.; Carolina Alves, R.; Sousa Araújo, V.H.; Lobato Duarte, J.; Maldonado Marchetti, J.; Chorilli, M. The use of TPGS in drug delivery systems to overcome biological barriers. Eur. Polym. J. 2021, 142, 110129. [Google Scholar] [CrossRef]
- Zhang, Z.; Tan, S.; Feng, S.S. Vitamin E TPGS as a molecular biomaterial for drug delivery. Biomaterials 2012, 33, 4889–4906. [Google Scholar] [CrossRef]
- Pechère, M.; Germanier, L.; Siegenthaler, G.; Pechère, J.-C.; Saurat, J.-H. The Antibacterial Activity of Topical Retinoids: The Case of Retinaldehyde. Dermatology 2002, 205, 153–158. [Google Scholar] [CrossRef]
- Oluwole, D.O.; Coleman, L.; Buchanan, W.; Chen, T.; La Ragione, R.M.; Liu, L.X. Antibiotics-Free Compounds for Chronic Wound Healing. Pharmaceutics 2022, 14, 1021. [Google Scholar] [CrossRef]
- Moghadamtousi, S.Z.; Kadir, H.A.; Hassandarvish, P.; Tajik, H.; Abubakar, S.; Zandi, K. A review on antibacterial, antiviral, and antifungal activity of curcumin. Biomed. Res. Int. 2014, 2014, 186864. [Google Scholar] [CrossRef]
- Adamczak, A.; Ozarowski, M.; Karpinski, T.M. Curcumin, a Natural Antimicrobial Agent with Strain-Specific Activity. Pharmaceuticals 2020, 13, 153. [Google Scholar] [CrossRef] [PubMed]
- Mun, S.H.; Joung, D.K.; Kim, Y.S.; Kang, O.H.; Kim, S.B.; Seo, Y.S.; Kim, Y.C.; Lee, D.S.; Shin, D.W.; Kweon, K.T.; et al. Synergistic antibacterial effect of curcumin against methicillin-resistant Staphylococcus aureus. Phytomedicine 2013, 20, 714–718. [Google Scholar] [CrossRef] [PubMed]
- European Committee for Antimicrobial Susceptibility Testing (EUCAST) of the European Society of Clinical Microbiology and Infectious Diseases (ESCMID). Determination of minimum inhibitory concentrations (MICs) of antibacterial agents by broth dilution. Clin. Microbiol. Infect. 2003, 9, ix–xv. [Google Scholar] [CrossRef]
- Miles, A.A.; Misra, S.S.; Irwin, J.O. The estimation of the bactericidal power of the blood. J. Hyg. 1938, 38, 732–749. [Google Scholar] [CrossRef]
- Merritt, J.H.; Kadouri, D.E.; O’Toole, G.A. Growing and analyzing static biofilms. In Current Protocol in Microbiology; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2005; Chapter 1, Unit 1B 1. [Google Scholar] [CrossRef]
- Reynolds, R.; Shackcloth, J.; Felmingham, D.; MacGowan, A. Comparison of BSAC agar dilution and NCCLS broth microdilution MIC methods for in vitro susceptibility testing of Streptococcus pneumoniae, Haemophilus influenzae and Moraxella catarrhalis: The BSAC Respiratory Resistance Surveillance Programme. J. Antimicrob. Chemother. 2003, 52, 925–930. [Google Scholar] [CrossRef]
- Pankey, G.A.; Sabath, L.D. Clinical Relevance of Bacteriostatic versus Bactericidal Mechanisms of Action in the Treatment of Gram-Positive Bacterial Infections. Clin. Infect. Dis. 2004, 38, 864–870. [Google Scholar] [CrossRef]
- Bjarnsholt, T.; Kirketerp-Møller, K.; Jensen, P.Ø.; Madsen, K.G.; Phipps, R.; Krogfelt, K.; Høiby, N.; Givskov, M. Why chronic wounds will not heal: A novel hypothesis. Wound Repair Regen. 2008, 16, 2–10. [Google Scholar] [CrossRef]
- Bernauer, U.; Bodin, L.; Celleno, L.; Chaudhry, Q.; Coenraads, P.; Dusinska, M.; Ezendam, J.; Gaffet, E.; Galli, C.; Granum, B.; et al. SCCS Final Opinion on Polyaminopropyl Biguanide (PHMB)—Submission III"—SCCS/1581/16—Final Version; Scientific Committee on Consumer Safety: Brussels, Belgium, 2018. [Google Scholar]
- Kim, W.; Zhu, W.; Hendricks, G.L.; Van Tyne, D.; Steele, A.D.; Keohane, C.E.; Fricke, N.; Conery, A.L.; Shen, S.; Pan, W.; et al. A new class of synthetic retinoid antibiotics effective against bacterial persisters. Nature 2018, 556, 103–107. [Google Scholar] [CrossRef]
- Tan, F.; She, P.; Zhou, L.; Liu, Y.; Chen, L.; Luo, Z.; Wu, Y. Bactericidal and Anti-biofilm Activity of the Retinoid Compound CD437 Against Enterococcus faecalis. Front. Microbiol. 2019, 10, 2301. [Google Scholar] [CrossRef]
- Zheng, D.; Huang, C.; Huang, H.; Zhao, Y.; Khan, M.R.U.; Zhao, H.; Huang, L. Antibacterial Mechanism of Curcumin: A Review. Chem. Biodivers. 2020, 17, e2000171. [Google Scholar] [CrossRef] [PubMed]
- Monserrat-Martinez, A.; Gambin, Y.; Sierecki, E. Thinking Outside the Bug: Molecular Targets and Strategies to Overcome Antibiotic Resistance. Int. J. Mol. Sci. 2019, 20, 1255. [Google Scholar] [CrossRef] [PubMed]
- Barrigah-Benissan, K.; Ory, J.; Dunyach-Remy, C.; Pouget, C.; Lavigne, J.P.; Sotto, A. Antibiofilm Properties of Antiseptic Agents Used on Pseudomonas aeruginosa Isolated from Diabetic Foot Ulcers. Int. J. Mol. Sci. 2022, 23, 1270. [Google Scholar] [CrossRef]
- Zheng, Y.; Wang, D.; Ma, L.Z. Effect of Polyhexamethylene Biguanide in Combination with Undecylenamidopropyl Betaine or PslG on Biofilm Clearance. Int. J. Mol. Sci. 2021, 22, 768. [Google Scholar] [CrossRef] [PubMed]
- Rippon, M.G.; Rogers, A.A.; Ousey, K. Polyhexamethylene biguanide and its antimicrobial role in wound healing: A narrative review. J. Wound Care 2023, 32, 5–20. [Google Scholar] [CrossRef]
- Allen, M.J.; Morby, A.P.; White, G.F. Cooperativity in the binding of the cationic biocide polyhexamethylene biguanide to nucleic acids. Biochem. Biophys. Res. Commun. 2004, 318, 397–404. [Google Scholar] [CrossRef]
- Yanai, R.; Ueda, K.; Nishida, T.; Toyohara, M.; Mori, O. Effects of ionic and surfactant agents on the antimicrobial activity of polyhexamethylene biguanide. Eye Contact Lens 2011, 37, 85–89. [Google Scholar] [CrossRef]
- He, S.; Zhan, Z.; Shi, C.; Wang, S.; Shi, X. Ethanol at Subinhibitory Concentrations Enhances Biofilm Formation in Salmonella Enteritidis. Foods 2022, 11, 2237. [Google Scholar] [CrossRef]
- Solano, C.; Echeverz, M.; Lasa, I. Biofilm dispersion and quorum sensing. Curr. Opin. Microbiol. 2014, 18, 96–104. [Google Scholar] [CrossRef]
- Wan, F.; Bohr, S.S.-R.; Kłodzińska, S.N.; Jumaa, H.; Huang, Z.; Nylander, T.; Thygesen, M.B.; Sørensen, K.K.; Jensen, K.J.; Sternberg, C.; et al. Ultrasmall TPGS–PLGA Hybrid Nanoparticles for Site-Specific Delivery of Antibiotics into Pseudomonas aeruginosa Biofilms in Lungs. ACS Appl. Mater. Interfaces 2020, 12, 380–389. [Google Scholar] [CrossRef]
- Vergalito, F.; Pietrangelo, L.; Petronio Petronio, G.; Colitto, F.; Alfio Cutuli, M.; Magnifico, I.; Venditti, N.; Guerra, G.; Di Marco, R. Vitamin E for prevention of biofilm-caused Healthcare-associated infections. Open Med. 2019, 15, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Brindhadevi, K.; LewisOscar, F.; Mylonakis, E.; Shanmugam, S.; Verma, T.N.; Pugazhendhi, A. Biofilm and Quorum sensing mediated pathogenicity in Pseudomonas aeruginosa. Process Biochem. 2020, 96, 49–57. [Google Scholar] [CrossRef]
- Soltani, S.; Fazly Bazzaz, B.S.; Hadizadeh, F.; Roodbari, F.; Soheili, V. New Insight into Vitamins E and K(1) as Anti-Quorum-Sensing Agents against Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2021, 65, e01342-20. [Google Scholar] [CrossRef] [PubMed]
- Barchitta, M.; Maugeri, A.; Favara, G.; Magnano San Lio, R.; Evola, G.; Agodi, A.; Basile, G. Nutrition and Wound Healing: An Overview Focusing on the Beneficial Effects of Curcumin. Int. J. Mol. Sci. 2019, 20, 1119. [Google Scholar] [CrossRef] [PubMed]
- Polcz, M.E.; Barbul, A. The Role of Vitamin A in Wound Healing. Nutr. Clin. Pract. 2019, 34, 695–700. [Google Scholar] [CrossRef] [PubMed]
- Kai, K.; Bi, W.; Bo, Y.; Ye, Y.; Zhang, D. Curcumin-A Review of Its Antibacterial Effect. Biomed. J. Sci. Tech. Res. 2020, 26, 19585–19587. [Google Scholar] [CrossRef]
- Cendra, M.D.M.; Blanco-Cabra, N.; Pedraz, L.; Torrents, E. Optimal environmental and culture conditions allow the in vitro coexistence of Pseudomonas aeruginosa and Staphylococcus aureus in stable biofilms. Sci. Rep. 2019, 9, 16284. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Molecular Weight | Purity/ Concentration | Solvent |
|---|---|---|---|
| Retinol | 286.45 | 100 mg/mL; 10% | Ethanol |
| Curcumin | 368.38 | 4 mg/mL; 0.4% | Ethanol |
| Curcumin | 368.38 | 4 mg/mL; 0.4% | Polysorbate 40 |
| Polyhexamethylene biguanide (PHMB) | - 1 | 208 mg/mL; 20% | water |
| D-α-Tocopherol polyethylene glycol 1000 succinate (TPGS) 3 | - 1 | 200 mg/mL; 20% | Milli-Q water |
| D-α-Tocopherol polyethylene glycol 1000 succinate micelles (TPGS-M) 3 | - 1 | 200 mg/mL; 20% | Milli-Q water |
| Polysorbate 40 | 1277 2 | 100% | - |
| Ethanol | 46.07 | 100% (>99.7%) | - |
| Compound (Concentration before Inoculation) | Vehicle | Bacterial Strain | |
|---|---|---|---|
| S. aureus | P. aeruginosa | ||
| MIC (mg/mL) | |||
| Retinol | Ethanol | 50 | 50 |
| Curcumin | Ethanol | 2 | 2 |
| Curcumin | Polysorbate 40 | 0.125 | 1 |
| PHMB | Water | 0.00649 | 0.0519 |
| TPGS | Water | 200 | 200 |
| Ethanol | - | 49.3125 | 49.3125 |
| Polysorbate 40 | - | >270.75 | >270.75 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coleman, L.; Adams, J.R.G.; Buchanan, W.; Chen, T.; La Ragione, R.M.; Liu, L.X. Non-Antibiotic Compounds Synergistically Kill Chronic Wound-Associated Bacteria and Disrupt Their Biofilms. Pharmaceutics 2023, 15, 1633. https://doi.org/10.3390/pharmaceutics15061633
Coleman L, Adams JRG, Buchanan W, Chen T, La Ragione RM, Liu LX. Non-Antibiotic Compounds Synergistically Kill Chronic Wound-Associated Bacteria and Disrupt Their Biofilms. Pharmaceutics. 2023; 15(6):1633. https://doi.org/10.3390/pharmaceutics15061633
Chicago/Turabian StyleColeman, Lucy, James R. G. Adams, Will Buchanan, Tao Chen, Roberto M. La Ragione, and Lian X. Liu. 2023. "Non-Antibiotic Compounds Synergistically Kill Chronic Wound-Associated Bacteria and Disrupt Their Biofilms" Pharmaceutics 15, no. 6: 1633. https://doi.org/10.3390/pharmaceutics15061633
APA StyleColeman, L., Adams, J. R. G., Buchanan, W., Chen, T., La Ragione, R. M., & Liu, L. X. (2023). Non-Antibiotic Compounds Synergistically Kill Chronic Wound-Associated Bacteria and Disrupt Their Biofilms. Pharmaceutics, 15(6), 1633. https://doi.org/10.3390/pharmaceutics15061633