
Effect of In Situ Bioremediation of Soil Contaminated with DDT and DDE by Stenotrophomonas sp. Strain DXZ9 and Ryegrass on Soil Microorganism

Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganism Strain DXZ9
2.2. Soil Sample
2.3. Experimental Design
2.4. Collection of Soil Samples
2.5. Microbial Enumeration and Community Structure and Enzymatic Activity of Tested Soil
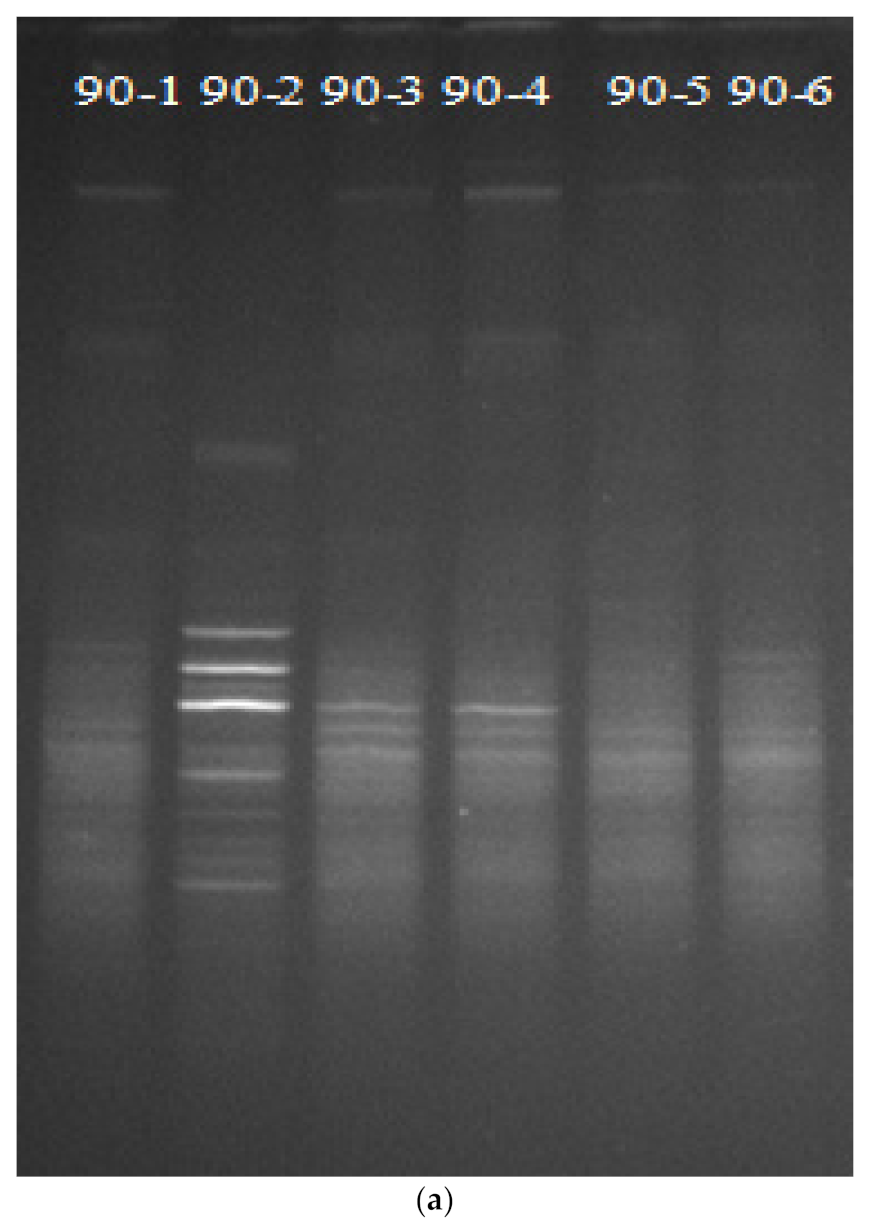
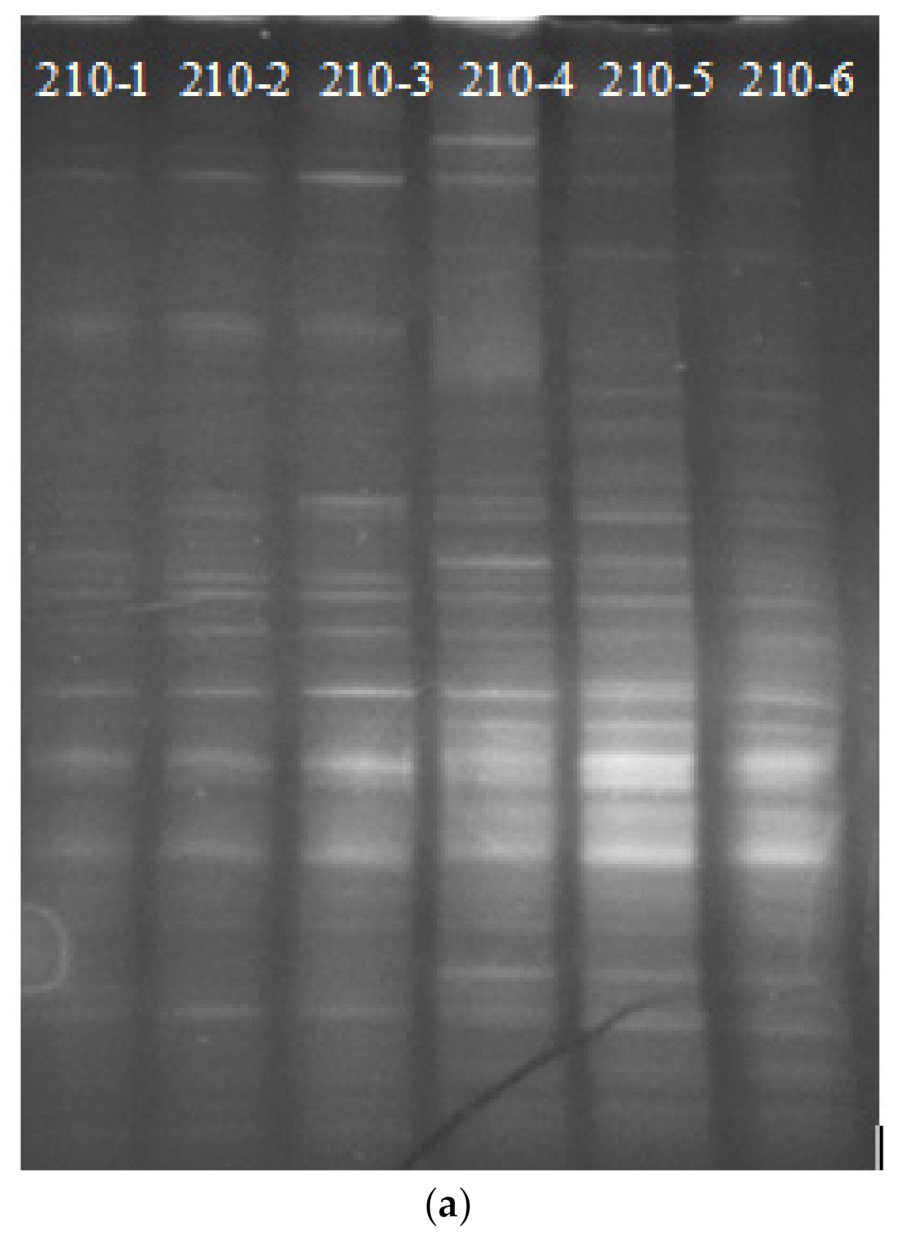
2.6. DGGE Analysis
2.7. Data Analysis and Statistical Methods
3. Results
3.1. Combined Bioremediation of DDT and DDE with Ryegrass and Microorganisms in Soil
3.2. Effect of Bioremediation on Soil Microorganism Quantity
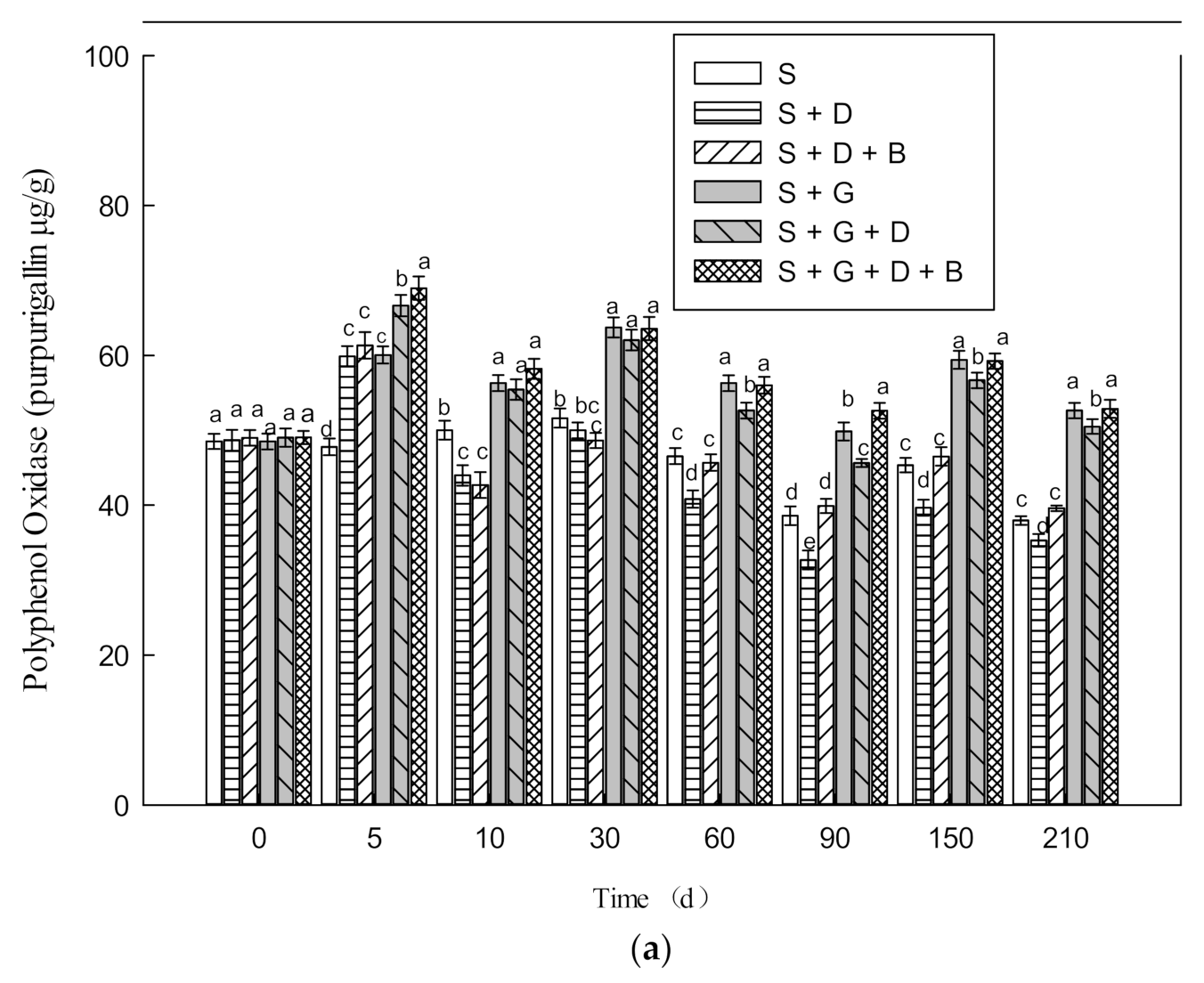
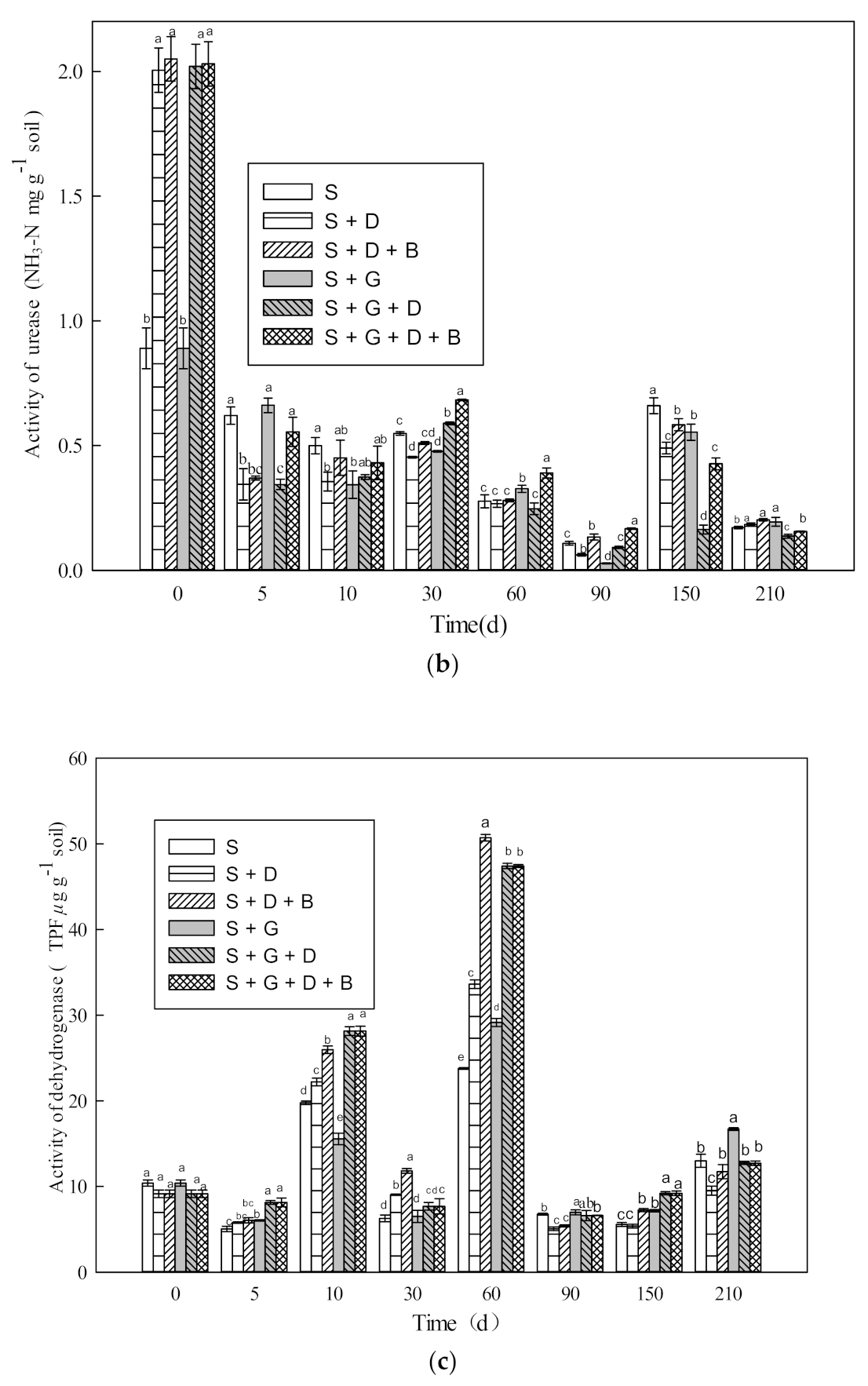
3.3. Effect of Bioremediation on the Activity of Soil Enzyme
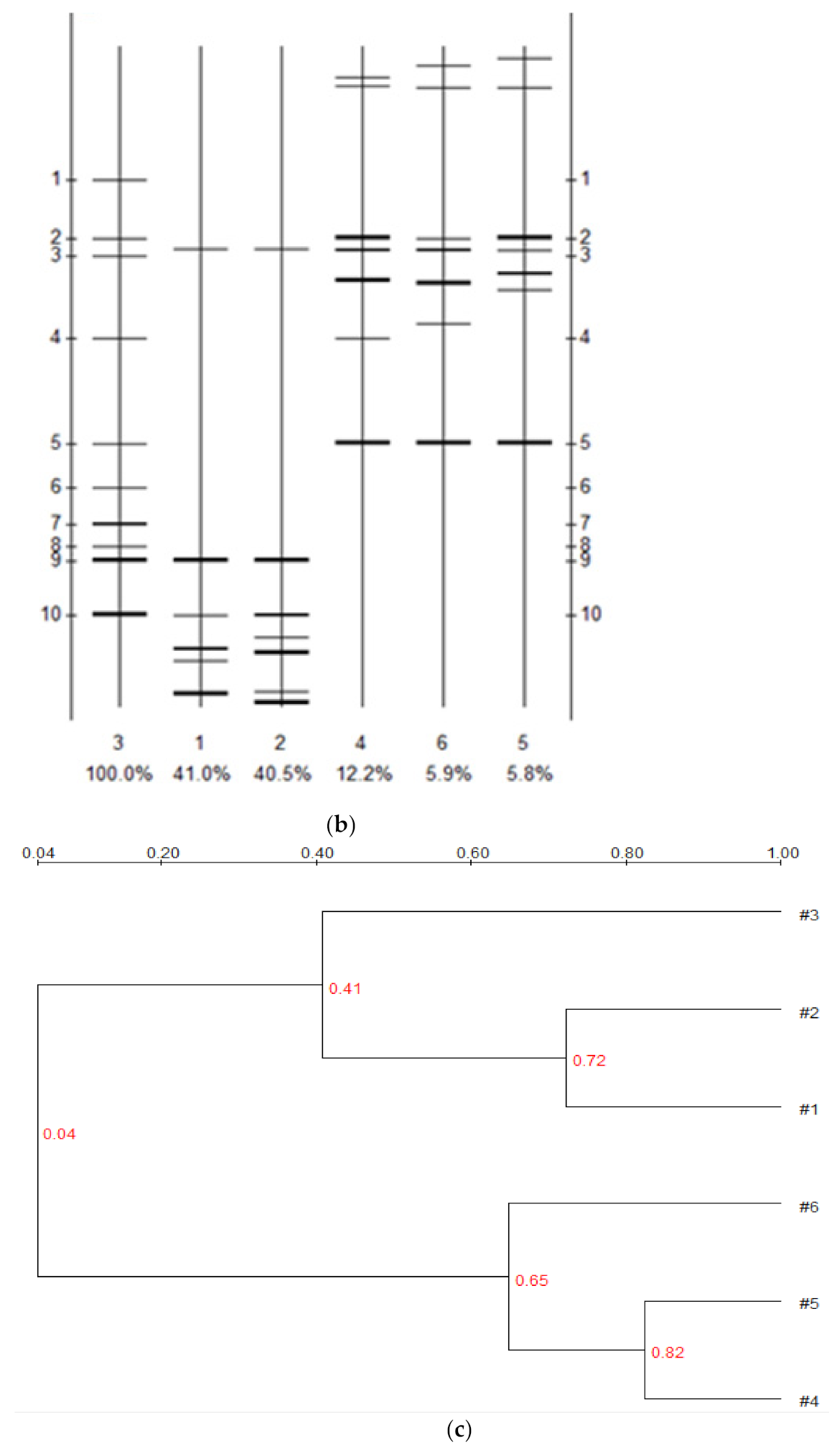
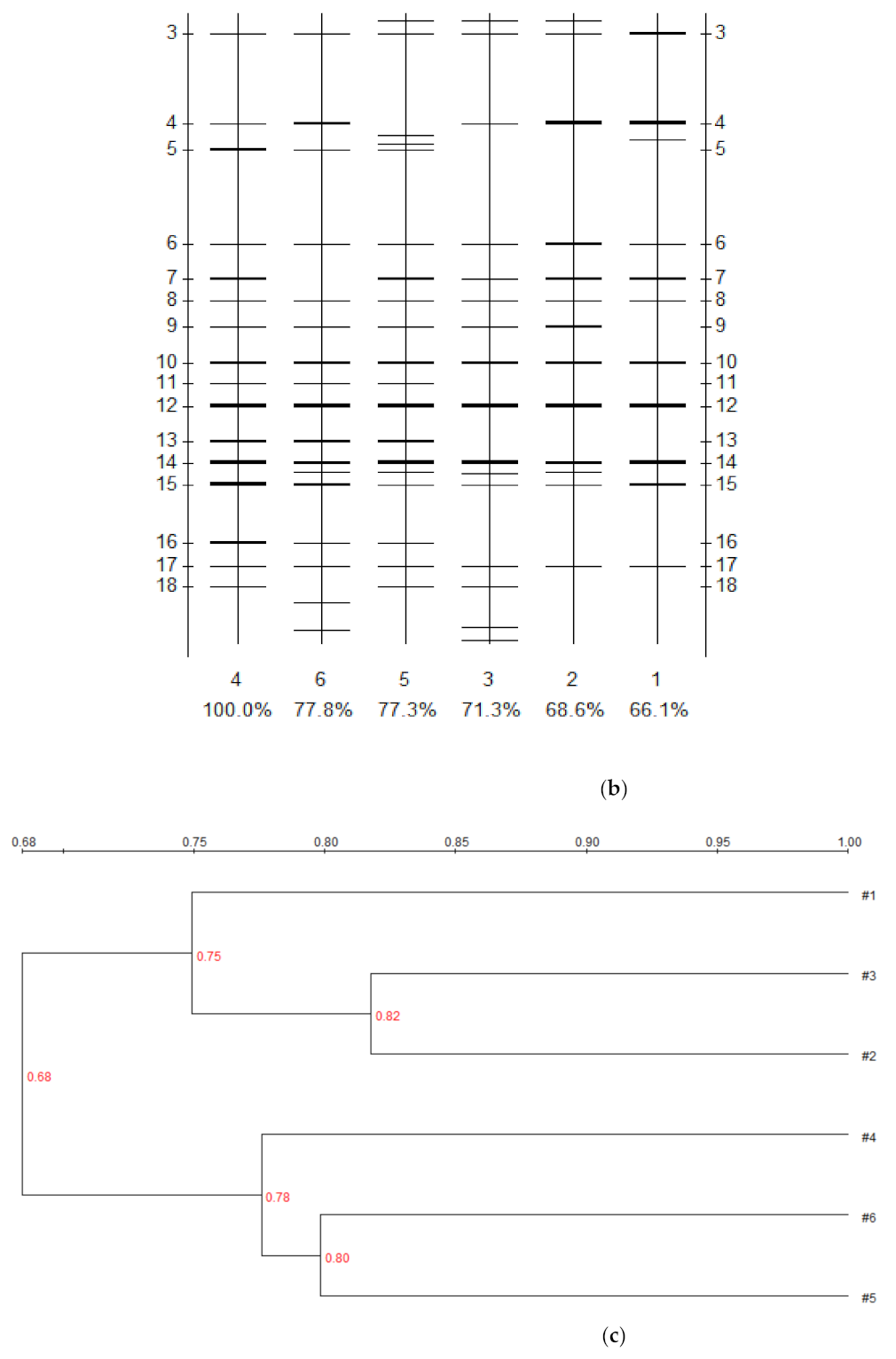
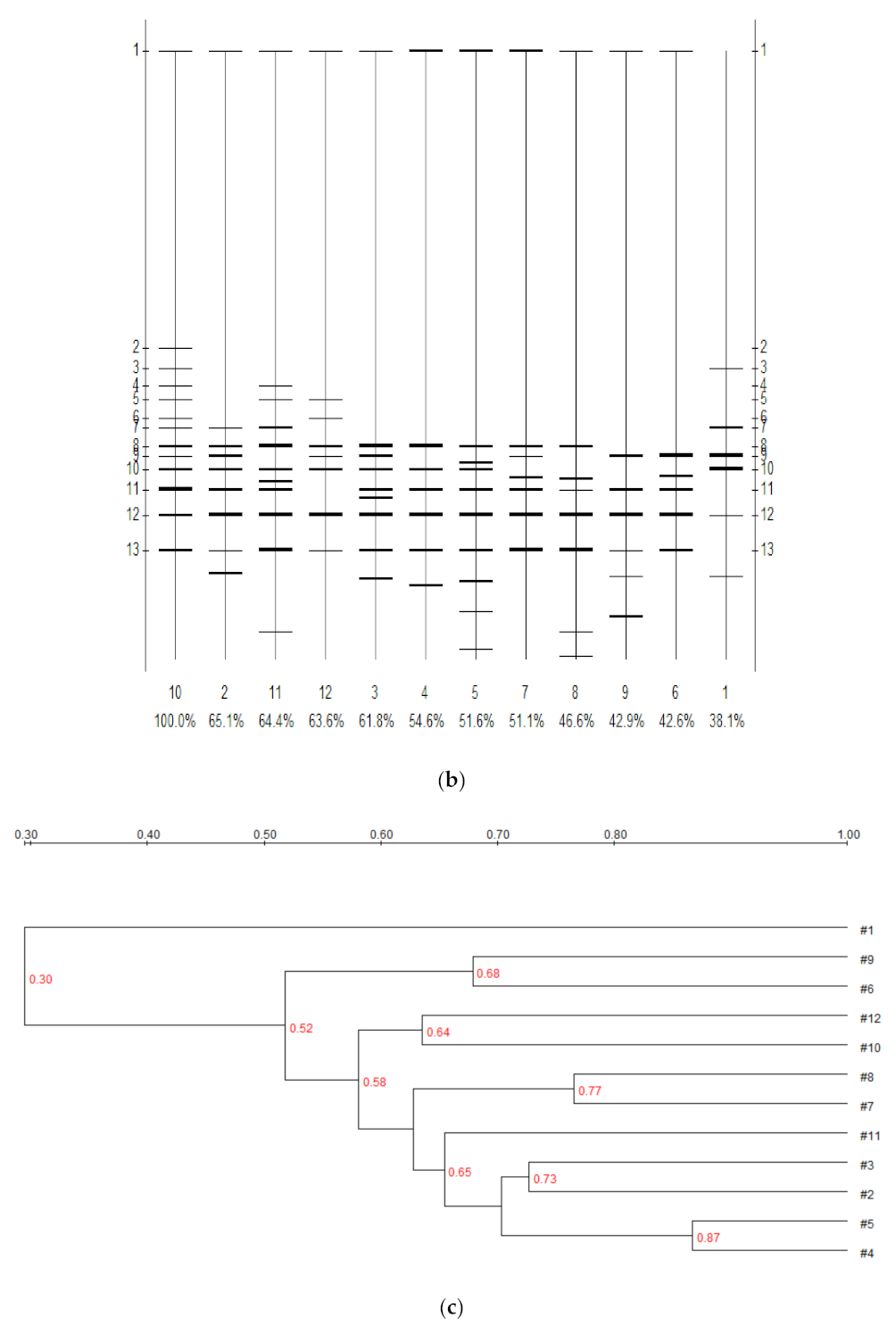
3.4. Effect of Bioremediation on Soil Bacterial Community Structure Diversity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Umulisa, V.; Kalisa, D.; Skutlarek, D.; Reichert, B. First evaluation of DDT (dichlorodiphenyltrichloroethane) residues and other Persistence Organic Pollutants in soils of Rwanda: Nyabarongo urban versus rural wetlands. Ecotoxicol. Environ. Saf. 2020, 197, 110574. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.C.; Xie, H.J.; Zhang, J.; Xu, J.T.; Liang, S. Spatial distribution of organochlorine pesticides (OCPs) and effect of soil characters: A case study of a pesticide producing factory. Chemosphere 2013, 90, 2381–2387. [Google Scholar] [CrossRef]
- Koureas, M.; Rousou, X.; Haftiki, H.; Mouchtouri, V.A.; Rachiotis, G.; Rakitski, V.; Tsakalof, A.; Hadjichristodoulou, C. Spatial and temporal distribution of p,p′-DDE (1-dichloro-2,2bis-(pchlorophenyl) ethylene) blood levels across the globe. A systematic review and meta-analysis. Sci. Total Environ. 2019, 686, 440–451. [Google Scholar] [CrossRef] [PubMed]
- Łozowicka, B.; Kaczyński, P.; Wolejko, E.; Piekutin, J.; Sagitov, A.; Toleubayev, K.; Isenova, G.; Abzeitova, E. Evaluation of organochlorine pesticide residues in soil and plants from East Europe and Central Asia. Desalinat. Water Treat. 2015, 57, 1310–1321. [Google Scholar] [CrossRef]
- Hu, W.Y.; Lu, Y.L.; Wang, T.Y.; Luo, W.; Shi, Y.J.; Giesy, J.P.; Geng, J.; Jiao, W.T.; Wang, G.; Chen, C.L. Spatial variability and temporal trends of HCH and DDT in soils around Beijing Guanting Reservoir, China. Environ. Geochem. Health 2010, 32, 441–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, J.G.; Shi, R.G.; Cai, Y.M.; Liu, Y.; Wang, Z.H.; Feng, J.M.; Zhao, M. Assessment of 20 Organochlorine Pesticides (OCPs) Pollution in Suburban Soil in Tianjin, China. Bull. Environ. Contam. Toxicol. 2010, 85, 137–141. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.N.; Hu, W.L.; Li, X.; Hu, F.W.; Liu, Y.F.; Xie, T.H.; Liu, B.; Xi, Y.N.; Su, Z.; Zhang, C. Effects of perfluoroalkyl substances on root and rhizosphere bacteria: Phytotoxicity, phyto-microbial remediation, risk assessment. Chemosphere 2021, 289, 133137. [Google Scholar] [CrossRef]
- Miao, R.H.; Guo, M.X.; Zhao, X.Y.; Gong, Z.Q.; Jia, C.Y.; Li, X.J.; Zhuang, J. Response of soil bacterial communities to polycyclic aromatic hydrocarbons during the phyto-microbial remediation of a contaminated soil. Chemosphere 2020, 261, 127779. [Google Scholar] [CrossRef]
- Ni, G.; Shi, G.Y.; Hu, C.X.; Wang, X.; Nie, M.; Cai, M.M.; Cheng, Q.; Zhao, X.H. Selenium improved the combined remediation efficiency of Pseudomonas aeruginosa and ryegrass on cadmium-nonylphenol co-contaminated soil. Environ. Pollut. 2021, 287, 117552. [Google Scholar] [CrossRef]
- Zhang, X.L.; Li, R.X.; Song, J.T.; Ren, Y.Y.; Luo, X.; Li, Y.; Li, X.J.; Li, T.; Wang, X.; Zhou, Q.X. Combined phyto-microbial-electrochemical system enhanced the removal of petroleum hydrocarbons from soil: A profundity remediation strategy. J. Hazard. Mater. 2021, 420, 126592. [Google Scholar] [CrossRef]
- Zhu, Z.-Q.; Yang, X.-E.; Wang, K.; Huang, H.-G.; Zhang, X.; Fang, H.; Li, T.-Q.; Alva, A.; He, Z.-L. Bioremediation of Cd-DDT co-contaminated soil using the Cd-hyperaccumulator Sedum alfredii and DDT-degrading microbes. J. Hazard. Mater. 2012, 235–236, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Koner, S.; Chen, J.S.; Hsu, B.M.; Rathod, J.; Huang, S.W.; Chien, H.Y.; Hussain, B.; Chan, M.W.Y. Depth-resolved microbial diversity and functional profiles of trichloroethylene-contaminated soils for Biolog EcoPlate-based biostimulation strategy. J. Hazard. Mater. 2022, 424, 127266. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Leoz, B.; Garbisu, C.; Charcosset, J.-Y.; Sánchez-Pérez, J.M.; Antigüedad, I.; Ruiz-Romera, E. Non-target effects of three formulated pesticides on microbially-mediated processes in a clay-loam soil. Sci. Total. Environ. 2013, 449, 345–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samarajeewa, A.D.; Velicogna, J.R.; Princz, J.I.; Subasinghe, R.M.; Scroggins, R.P.; Beaudette, L.A. Effect of silver nano-particles on soil microbial growth, activity and community diversity in a sandy loam soil. Environ. Pollut. 2017, 220, 504–513. [Google Scholar] [CrossRef]
- Fragoeiro, S.; Magan, N. Impact of Trametes versicolor and Phanerochaete chrysosporium on differential breakdown of pesticide mixtures in soil microcosms at two water potentials and associated respiration and enzyme activity. Int. Biodeterior. Biodegrad. 2008, 62, 376–383. [Google Scholar] [CrossRef] [Green Version]
- Tejada, M.; Go´mez, I.; Toro, M.D. Use of organic amendments as a bioremediation strategy to reduce the bioavailability of chlorpyrifos insecticide in soils. Effects on soil biology. Ecotox. Environ. Safe. 2011, 74, 2075–2081. [Google Scholar] [CrossRef]
- Megharaj, M.; Kantachote, D.; Singleton, I.; Naidu, R. Effects of long-term contamination of DDT on soil microform with special reference to soil algae and algal transformation of DDT. Environ. Pollut. 2000, 109, 35–42. [Google Scholar] [CrossRef]
- Xie, H.; Zhu, L.S.; Wang, J. Combined treatment of contaminated soil with a bacterial Stenotrophomonas strain DXZ9 and ryegrass (Lolium perenne) enhances DDT and DDE remediation. Environ. Sci. Pollut. Res. 2018, 25, 31895–31905. [Google Scholar] [CrossRef]
- Xie, H.; Zhu, L.; Xu, Q.; Wang, J.; Liu, W.; Jiang, J.; Meng, Y. Isolation and degradation ability of the DDT-degrading bacterial strain KK. Environ. Earth Sci. 2010, 62, 93–99. [Google Scholar] [CrossRef]
- Guo, P.P.; Zhu, L.S.; Wang, J.H.; Wang, J.; Xie, H.; Lv, D.D. Enzymatic activities and microbial biomass in black soil as affected by azoxystrobin. Environ. Earth. Sci. 2015, 74, 1353–1361. [Google Scholar] [CrossRef]
- Wei, J.; Zhang, X.Y.; Liu, X.Y.; Liang, X.; Chen, X.P. Influence of root components of celery on pyrene bioaccessibility, soil enzymes and microbial communities in pyrene and pyrene-diesel spiked soils. Sci. Total. Environ. 2017, 599–600, 50–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stpniewska, Z.; Wolińska, A.; Ziomek, J. Response of soil catalase activity to chromium contamination. J. Environ. Sci. 2009, 21, 1142–1147. [Google Scholar] [CrossRef]
- Chen, T.R.; Liu, X.Y.; Zhang, X.Y.; Chen, X.; Tao, K.Y.; Hu, X.X. Effect of alkyl polyglucoside and nitrilotriacetic acid combined application on lead/pyrene bioavailability and dehydrogenase activity in co-contaminated soils. Chemosphere 2016, 154, 515–520. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.M.; Zhu, L.S.; Wang, J.; Xie, H.; Wang, J.H.; Wang, F.H.; Sun, F.X. Effects of fomesafen on soil enzyme activity, microbial population, and bacterial community composition. Environ. Monit. Assess. 2014, 186, 2801–2812. [Google Scholar] [CrossRef] [PubMed]
- Yousaf, S.; Andria, V.; Reichenauer, T.G.; Smalla, K.; Sessitsch, A. Phylogenetic and functional diversity of alkane degrading bacteria associated with Italian ryegrass (Lolium multiflorum) and Birdsfoot trefoil (Lotus corniculatus) in a petroleum oil-contaminated environment. J. Hazard. Mater. 2010, 184, 523–532. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.L.; Dong, Y.M.; Zhang, Z.; Song, Z.G. Effect of dibutyl phthalate on microbial function diversity and enzyme activity in wheat rhizosphere and non-rhizosphere soils. Environ. Pollut. 2020, 265, 114800. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Li, W.; Dick, W.A.; Ye, X.; Chen, K.; Kost, D.; Chen, L. Bioremediation of hydrocarbon degradation in a petroleum-contaminated soil and microbial population and activity determination. Chemosphere 2017, 169, 124–130. [Google Scholar] [CrossRef]
- Wang, B.B.; Wang, Q.L.; Liu, W.X.; Liu, X.Y.; Hou, J.Y.; Teng, Y.; Luo, Y.M.; Christie, P. Biosurfactant-producing microorganism Pseudomonas sp. SB assists the phytoremediation of DDT-contaminated soil by two grass species. Chemosphere 2017, 182, 137–142. [Google Scholar] [CrossRef]
- Siciliano, S.D.; Fortin, N.; Mihoc, A.; Wisse, G.; Labelle, S.; Beaumier, D.; Ouellette, D.; Roy, R.; Whyte, L.G.; Banks, M.K.; et al. Selection of Specific Endophytic Bacterial Genotypes by Plants in Response to Soil Contamination. Appl. Environ. Microbiol. 2001, 67, 2469–2475. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Zhang, S.; Huang, H.; Christie, P. Behavior of decabromodiphenyl ether (BDE-209) in soil: Effects of rhizosphere and mycorrhizal colonization of ryegrass roots. Environ. Pollut. 2011, 159, 749–753. [Google Scholar] [CrossRef] [Green Version]
- Xie, X.; Liao, M.; Fang, S.; Peng, Y.; Yang, J.; Chai, J. Spacial characteristics of pyrene degradation and soil microbial activity with the distance from the ryegrass (Lolium perenne L.) root surface in a multi-interlayer rhizobox. J. Hazard. Mater. 2012, 213–214, 156–160. [Google Scholar] [CrossRef] [PubMed]
- Anderson, T.A.; Guthrie, E.A.; Walton, B.T. Bioremediation in the rhizosphere. Environ. Sci. Technol. 1993, 27, 2630–2636. [Google Scholar] [CrossRef]
- Rentz, J.A.; Alvarez, P.J.J.; Schnoor, J.L. Benzo[a]pyrene co-metabolism in the presence of plant root extracts and exudates: Implications for phytoremediation. Environ. Pollut. 2005, 136, 477–484. [Google Scholar] [CrossRef]
- Korade, D.L.; Fulekar, M. Rhizosphere remediation of chlorpyrifos in mycor rhizospheric soil using ryegrass. J. Hazard. Mater. 2009, 172, 1344–1350. [Google Scholar] [CrossRef]
- Kidd, P.S.; Prieto-Fernandez, A.; Monterroso, C.; Acea, M.J. Rhizosphere microbial community and hexachlorocyclohexane degradative potential in contrasting plant species. Plant Soil 2007, 302, 233–247. [Google Scholar] [CrossRef]
- Tang, J.C.; Wang, R.G.; Niu, X.W.; Zhou, Q.X. Enhancement of soil petroleum remediation by using a combination of ryegrass (Lolium perenne) and different microorganisms. Soil Tillage Res. 2010, 110, 87–93. [Google Scholar] [CrossRef]
- Wang, J.J.; Hu, S.B.; Wei, L.Q.; Chai, Q.Q.; Liu, J.B.; Wang, J.Y.; Yang, W.X. Phytoremediation of polycyclic aromatic hydrocarbons (PAHs)-contaminated soil by intercropped beet-grasses. J. Agro-Environ. Sci. 2016, 35, 1090–1096. (In Chinese) [Google Scholar]
- Ye, J.; Yin, H.; Peng, H.; Bai, J.; Li, Y. Pyrene removal and transformation by joint application of alfalfa and exogenous microorganisms and their influence on soil microbial community. Ecotoxicol. Environ. Saf. 2014, 110, 129–135. [Google Scholar] [CrossRef]
- Wu, M.L.; Dick, W.A.; Li, W.; Wang, Q.Y.X.; Yang, Q.; Wang, T.T.; Xu, L.M.; Zhang, M.H.; Chen, L.M. Bioaugmentation and biostimulation of hydrocarbon degradation and the microbial community in a petroleum-contaminated soil. Int. Biodeterior. Biodegrad. 2016, 107, 158–164. [Google Scholar] [CrossRef]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time (Day) | Treatment | Mean Concentration (mg·kg−1) | |||
|---|---|---|---|---|---|
| p,p′-DDE | p,p′-DDD | p,p′-DDT | Total Concentration | ||
| 0 | 1.26 ± 0.18 | 0.16 ± 0.06 | 2.23 ± 0.25 | 3.65 | |
| 5 | S + G + D + B | 0.75 ± 0.12 c | 0.14 ± 0.04 a | 1.51 ± 0.07 c | 2.4 |
| S + D + B | 0.79 ± 0.09 c | 0.14 ± 0.02 a | 1.58 ± 0.05 c | 2.51 | |
| S + G + D | 0.87 ± 0.05 b | 0.16 ± 0.03 a | 1.70 ± 0.08 b | 2.73 | |
| S + D | 1.15 ± 0.07 a | 0.16 ± 0.02 a | 2.11 ± 0.11 a | 3.42 | |
| 10 | S + G + D + B | 0.69 ± 0.03 c | 0.13 ± 0.01 a | 1.32 ± 0.10 c | 2.14 |
| S + D + B | 0.74 ± 0.04 c | 0.13 ± 0.01 a | 1.41 ± 0.12 c | 2.28 | |
| S + G + D | 0.84 ± 0.08 b | 0.16 ± 0.03 a | 1.53 ± 0.15 b | 2.53 | |
| S + D | 1.06 ± 0.09 a | 0.16 ± 0.02 a | 2.08 ± 0.17 a | 3.30 | |
| 30 | S + G + D + B | 0.70 ± 0.03 c | 0.13 ± 0.03 a | 1.27 ± 0.11 c | 2.10 |
| S + D + B | 0.70 ± 0.02 c | 0.13 ± 0.02 a | 1.31 ± 0.10 c | 2.14 | |
| S + G + D | 0.83 ± 0.05 b | 0.15 ± 0.01 a | 1.45 ± 0.13 b | 2.43 | |
| S + D | 0.99 ± 0.07 a | 0.15 ± 0.02 a | 1.85 ± 0.17 a | 2.99 | |
| 60 | S + G + D + B | 0.71 ± 0.02 b | 0.12 ± 0.03 a | 1.06 ± 0.09 c | 1.89 |
| S + D + B | 0.75 ± 0.04 b | 0.13 ± 0.02 a | 1.11 ± 0.08 b,c | 1.99 | |
| S + G + D | 0.79 ± 0.03 b | 0.15 ± 0.02 a | 1.16 ± 0.07 b | 2.1 | |
| S + D | 0.95 ± 0.07 a | 0.15 ± 0.04 a | 1.72 ± 0.10 a | 2.82 | |
| 90 | S + G + D + B | 0.69 ± 0.02 b | 0.12 ± 0.03 a | 0.85 ± 0.06 | 1.66 |
| S + D + B | 0.73 ± 0.01 b | 0.12 ± 0.02 a | 0.90 ± 0.03 | 1.70 | |
| S + G + D | 0.75 ± 0.03 b | 0.13 ± 0.04 a | 0.99 ± 0.05 | 1.87 | |
| S + D | 0.92 ± 0.04 a | 0.13 ± 0.01 a | 1.66 ± 0.09 | 2.71 | |
| 150 | S + G + D + B | 0.68 ± 0.02 b | 0.17 ± 0.03 a | 0.75 ± 0.08 b | 1.60 |
| S + D + B | 0.69 ± 0.04 b | 0.16 ± 0.01 a | 0.78 ± 0.06 b | 1.63 | |
| S + G + D | 0.70 ± 0.06 b | 0.15 ± 0.02 a | 0.80 ± 0.05 b | 1.65 | |
| S + D | 0.90 ± 0.03 a | 0.14 ± 0.01 a | 1.60 ± 0.12 a | 2.64 | |
| 210 | S + G + D + B | 0.56 ± 0.01 c | 0.14 ± 0.02 a | 0.43 ± 0.01 d | 1.13 |
| S + D + B | 0.60 ± 0.02b c | 0.15 ± 0.01 a | 0.51 ± 0.03 c | 1.26 | |
| S + G + D | 0.65 ± 0.02 b | 0.16 ± 0.03 a | 0.62 ± 0.06 b | 1.43 | |
| S + D | 0.87 ± 0.03 a | 0.17 ± 0.02 a | 1.56 ± 0.09 a | 2.60 | |
| Time (Day) | Lane | 1 | 2 | 3 | 4 | 5 | 6 |
|---|---|---|---|---|---|---|---|
| 10 | 1 | 100 | 72.2 | 41 | 1.4 | 1.6 | 1.7 |
| 2 | 100 | 40.5 | 2.2 | 2.5 | 2.5 | ||
| 3 | 100 | 12.2 | 5.8 | 5.9 | |||
| 4 | |||||||
| 5 | 100 | 59.7 | |||||
| 90 | 1 | 100 | 38.2 | 54.8 | 78.2 | 33.2 | 79.1 |
| 2 | 100 | 35.0 | 35.0 | 31.9 | 27.3 | ||
| 3 | 100 | 52.5 | 35.2 | 53.9 | |||
| 4 | 100 | 40.8 | 76.9 | ||||
| 5 | 100 | 38.1 | |||||
| 210 | 1 | 100 | 75.5 | 74.4 | 66.1 | 61.6 | 62.6 |
| 2 | 100 | 81.8 | 68.6 | 71.2 | 70.6 | ||
| 3 | 100 | 71.3 | 73.1 | 71.0 | |||
| 4 | 100 | 77.3 | 77.8 | ||||
| 5 | 79.8 |
| Lane | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 100 | 43.5 | 37.9 | 20.7 | 24.5 | 34.1 | 16.4 | 10.0 | 36.3 | 38.1 | 32.8 | 30.2 |
| 2 | 100 | 72.7 | 72.5 | 71.2 | 57.0 | 66.5 | 51.0 | 65.2 | 65.1 | 63.0 | 66.0 | |
| 3 | 100 | 70.8 | 66.8 | 53.9 | 61.1 | 55.5 | 56.1 | 61.8 | 65.9 | 57.9 | ||
| 4 | 100 | 86.7 | 50.8 | 71.8 | 59.0 | 50.8 | 54.6 | 66.0 | 64.3 | |||
| 5 | 100 | 52.7 | 71.6 | 62.6 | 53.6 | 51.6 | 66.9 | 60.9 | ||||
| 6 | 100 | 67.9 | 57.2 | 67.9 | 42.6 | 43.1 | 45.9 | |||||
| 7 | 100 | 76.5 | 54.6 | 51.1 | 66.4 | 61.9 | ||||||
| 8 | 100 | 46.2 | 46.6 | 62.5 | 50.3 | |||||||
| 9 | 100 | 42.9 | 38.8 | 53.8 | ||||||||
| 10 | 100 | 64.4 | 63.6 | |||||||||
| 11 | 100 | 56.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, H.; Liu, R.; Xu, Y.; Liu, X.; Sun, F.; Ma, Y.; Wang, Y. Effect of In Situ Bioremediation of Soil Contaminated with DDT and DDE by Stenotrophomonas sp. Strain DXZ9 and Ryegrass on Soil Microorganism. Microbiol. Res. 2022, 13, 64-86. https://doi.org/10.3390/microbiolres13010005
Xie H, Liu R, Xu Y, Liu X, Sun F, Ma Y, Wang Y. Effect of In Situ Bioremediation of Soil Contaminated with DDT and DDE by Stenotrophomonas sp. Strain DXZ9 and Ryegrass on Soil Microorganism. Microbiology Research. 2022; 13(1):64-86. https://doi.org/10.3390/microbiolres13010005
Chicago/Turabian StyleXie, Hui, Ruiyuan Liu, Yuxin Xu, Xin Liu, Fengxia Sun, Yuhan Ma, and Yuying Wang. 2022. "Effect of In Situ Bioremediation of Soil Contaminated with DDT and DDE by Stenotrophomonas sp. Strain DXZ9 and Ryegrass on Soil Microorganism" Microbiology Research 13, no. 1: 64-86. https://doi.org/10.3390/microbiolres13010005
APA StyleXie, H., Liu, R., Xu, Y., Liu, X., Sun, F., Ma, Y., & Wang, Y. (2022). Effect of In Situ Bioremediation of Soil Contaminated with DDT and DDE by Stenotrophomonas sp. Strain DXZ9 and Ryegrass on Soil Microorganism. Microbiology Research, 13(1), 64-86. https://doi.org/10.3390/microbiolres13010005