
Antibacterial Effect and Mode of Action of Secondary Metabolites from Fungal Endophyte Associated with Aloe ferox Mill

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of Plant Samples
2.2. Plant Treatment
2.3. Fungal Isolation
2.4. Screening for Production of Antimicrobial Compounds
2.4.1. Primary Screening
2.4.2. Secondary Screening
2.5. Secondary Metabolite Production Phase
2.6. Molecular Identification of the Fungus
2.7. Extraction of the Secondary Metabolites
2.8. Antibacterial Activity of the Extract
2.8.1. Minimum Inhibitory Concentration (MIC) of the Extract
2.8.2. Minimum Bactericidal Concentration (MBC) of the Extract
2.9. Mode of Action
2.9.1. Determination of the Respiratory Chain Dehydrogenase Activity
2.9.2. Effect of the Extract on the Release of 260 nm Absorbing Materials
2.10. Characterisation of the Bacterial Extract
2.10.1. Chemical Composition
2.10.2. Assessment of Functional Groups
2.11. Statistical Analysis
3. Results and Discussion
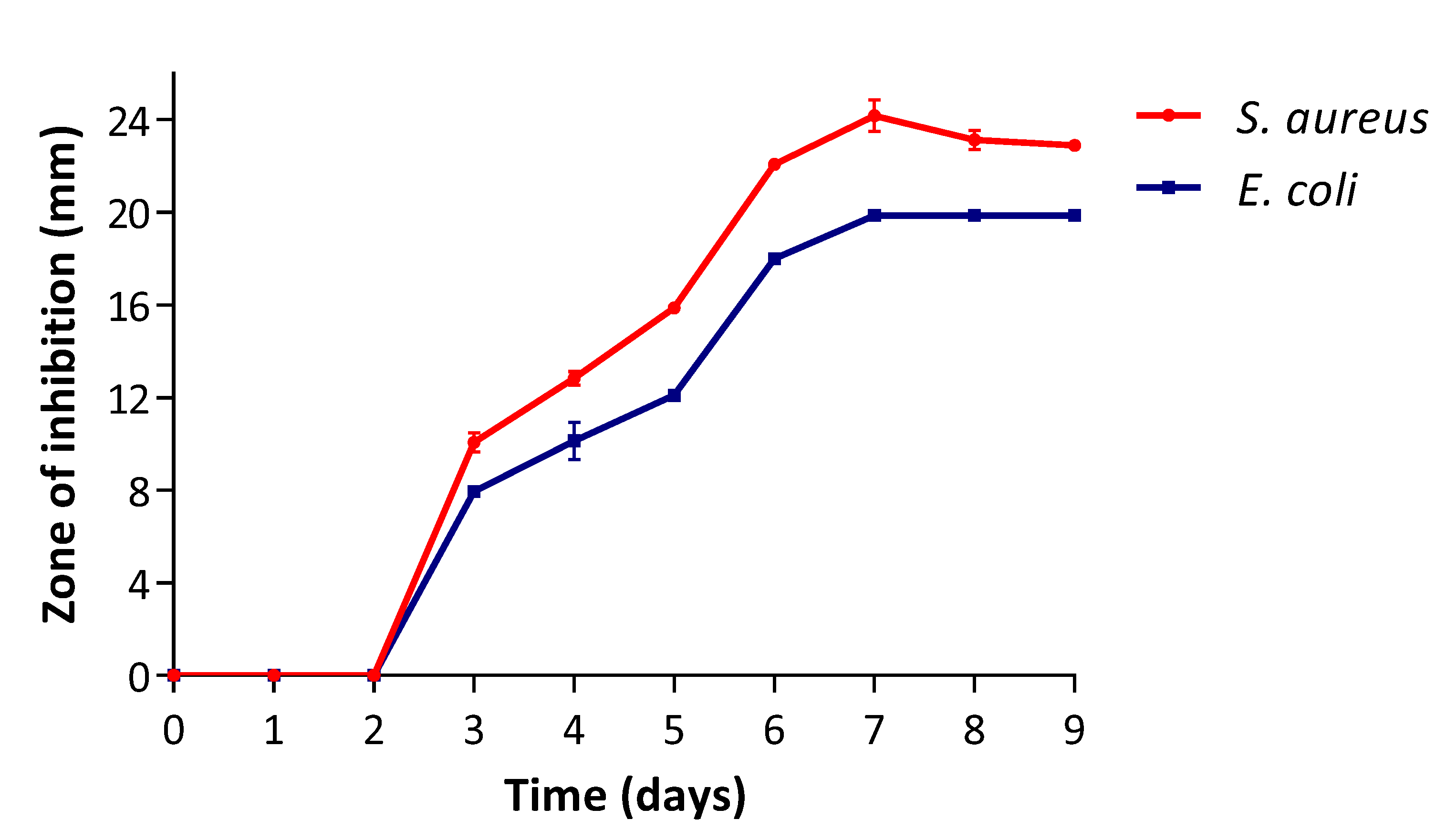
3.1. Secondary Metabolite Production Stage
3.2. Antibacterial Activity of the Fungal Extract
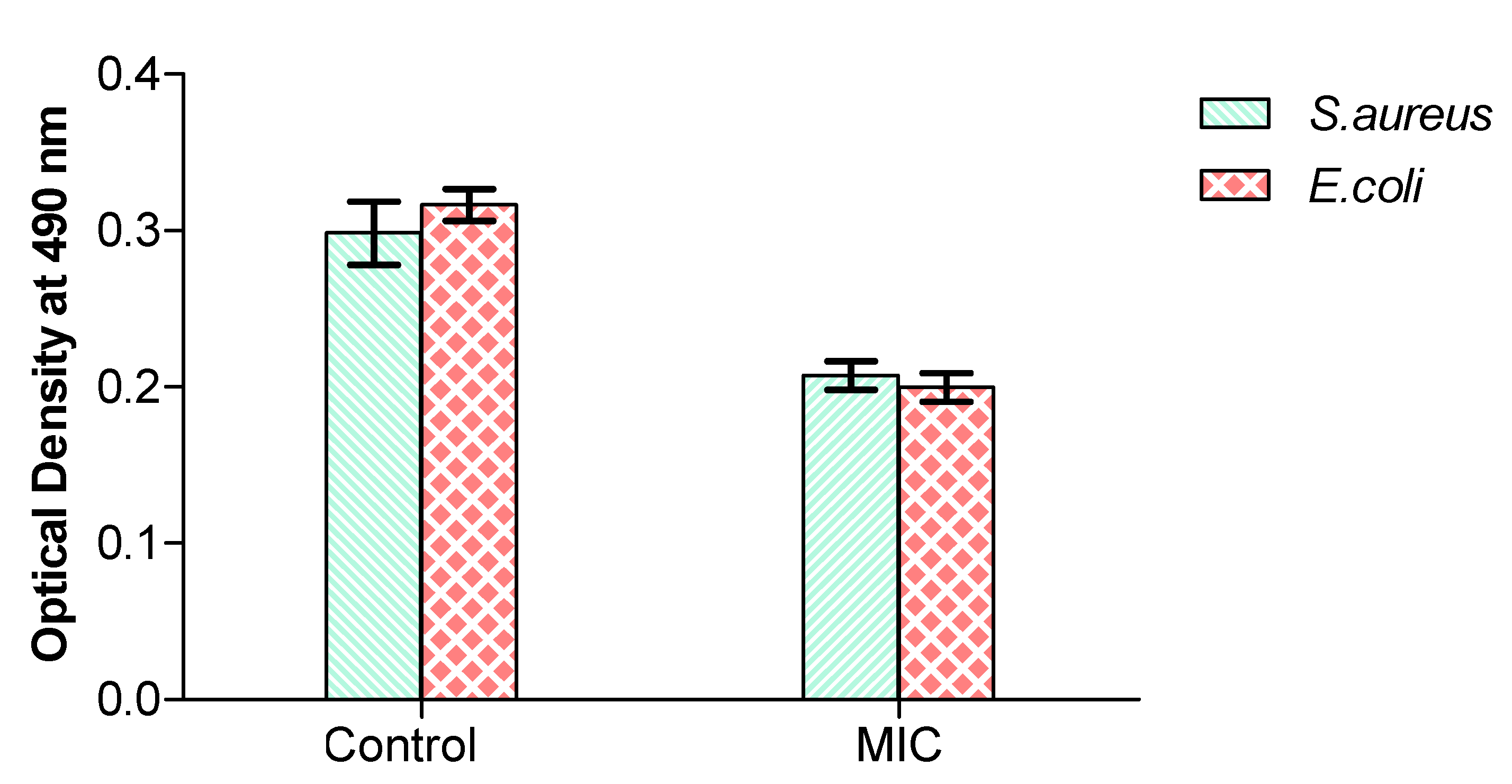
3.3. RCD Activity
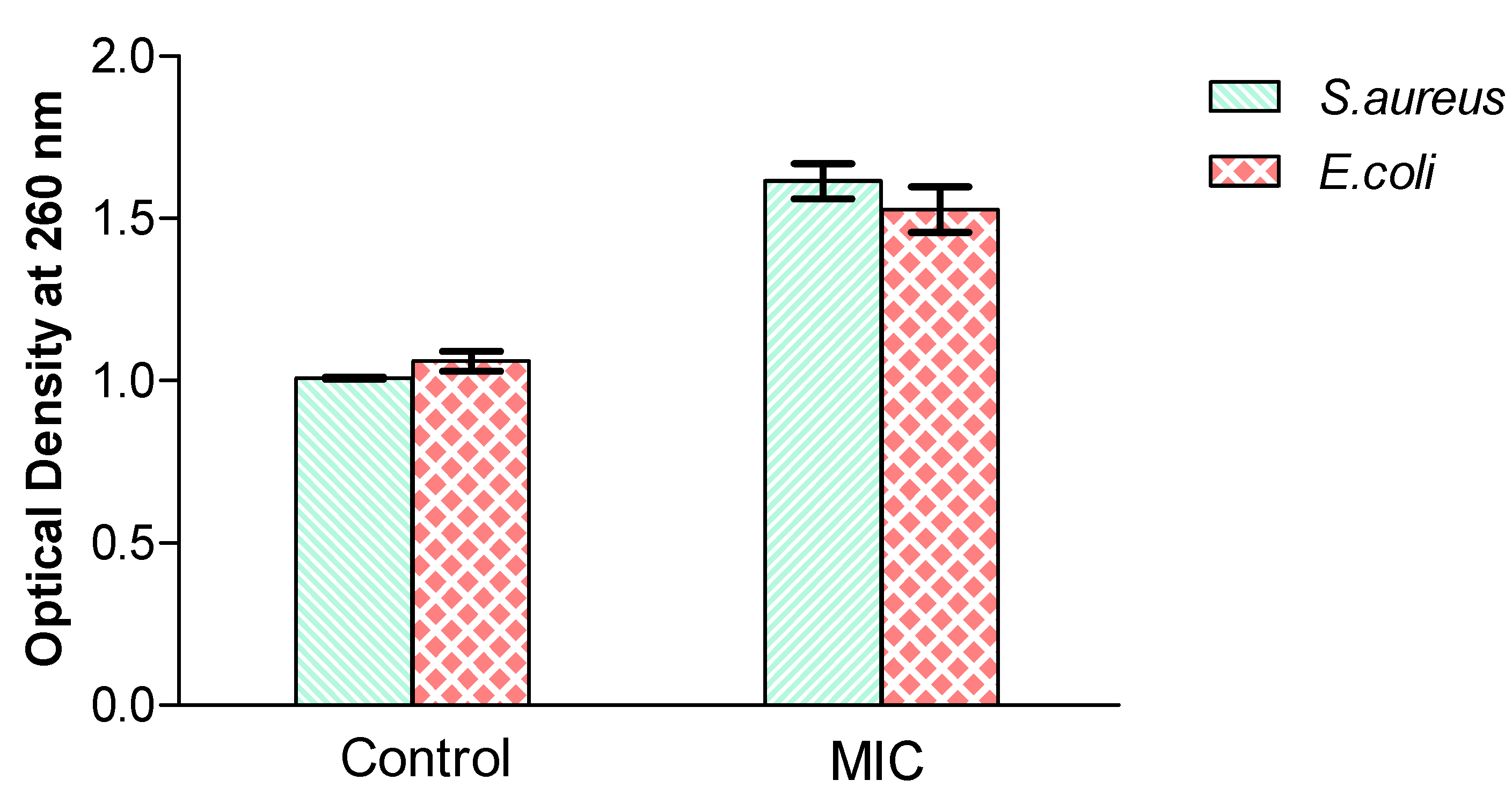
3.4. Effect of the Extract on the Release of 260 nm Absorbing Materials
3.5. Chemical Constituents of the Extract
3.6. Functional Groups of the Extract
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Orsi, R.H.; den Bakker, H.C.; Wiedmann, M. Listeria monocytogenes lineages: Genomics, evolution, ecology, and phenotypic characteristics. Int. J. Med. Microbiol. 2011, 301, 79–96. [Google Scholar] [CrossRef] [PubMed]
- Mujaddidi, N.; Nisa, S.; Al Ayoubi, S.; Bibi, Y.; Khan, S.; Sabir, M.; Zia, M.; Ahmad, S.; Qayyum, A. Pharmacological properties of biogenically synthesized silver nanoparticles using endophyte Bacillus cereus extract of Berberis lyceum against oxidative stress and pathogenic multidrug-resistant bacteria. Saudi J. Biol. Sci. 2021, 28, 6432–6440. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Global Action Plan for the Prevention and Control of Noncommunicable Diseases 2013–2020; World Health Organization: Geneva Switzerland, 2013. [Google Scholar]
- Kebede, Y.; Hayder, A.; Girma, K.; Abamecha, F.; Alemayehu, G.; Abebe, L.; Sudhakar, M.; Birhanu, Z. Primary school students’ poetic malaria messages from Jimma zone, Oromia, Ethiopia: A qualitative content analysis. BMC Public Health 2021, 21, 1688. [Google Scholar] [CrossRef] [PubMed]
- Mitra, I.; Huang, B.; Mousavi, N.; Ma, N.; Lamkin, M.; Yanicky, R.; Shleizer-Burko, S.; Lohmueller, K.E.; Gymrek, M. Patterns of de novo tandem repeat mutations and their role in autism. Nature 2021, 589, 246–250. [Google Scholar] [CrossRef]
- Pál, S.; Tóth, R.; Nosanchuk, J.; Vágvölgyi, C.; Németh, T.; Gácser, A. A Candida parapsilosis overexpression collection reveals genes required for pathogenesis. J. Fungi 2021, 7, 97. [Google Scholar] [CrossRef]
- Jain, R.; Bhardwaj, P.; Pandey, S.S.; Kumar, S. Arnebia euchroma, a plant species of cold desert in the Himalayas, harbors beneficial cultivable endophytes in roots and leaves. Front. Microbiol. 2021, 12, 696667. [Google Scholar] [CrossRef]
- Khalmuratova, R.; Shin, H.-W. Crosstalk between mucosal inflammation and bone metabolism in chronic rhinosinusitis. Clin. Exp. Otorhinolaryngol. 2021, 14, 43. [Google Scholar] [CrossRef]
- Martinez-Klimova, E.; Rodríguez-Peña, K.; Sánchez, S. Endophytes as sources of antibiotics. Biochem. Pharmacol. 2017, 134, 1–17. [Google Scholar] [CrossRef]
- Khare, E.; Mishra, J.; Arora, N.K. Multifaceted interactions between endophytes and plant: Developments and prospects. Front. Microbiol. 2018, 9, 2732. [Google Scholar] [CrossRef]
- Moghaddam, N.; Dabard, C.; Dufour, M.; Po, H.; Xu, X.; Pons, T.; Lhuillier, E.; Ithurria, S. Surface modification of CdE (E: S, Se, and Te) nanoplatelets to reach thicker nanoplatelets and homostructures with confinement-induced intraparticle type I energy level alignment. J. Am. Chem. Soc. 2021, 143, 1863–1872. [Google Scholar] [CrossRef]
- Mulla, K.P.; Dandu, C.; Thirumalanadhuni, V.; Shaik, S.; Shaik, I.A.; Nimmanapalli, Y. Phytochemical, elemental, physico-chemical, HPTLC and anticancer investigations of Ceropegia spiralis Wight. Tuber extracts. Pharma Innov. J. 2021, 10, 95–105. [Google Scholar] [CrossRef]
- Song, E.; Zhang, C.; Israelow, B.; Lu-Culligan, A.; Prado, A.V.; Skriabine, S.; Lu, P.; Weizman, O.-E.; Liu, F.; Dai, Y. Neuroinvasion of SARS-CoV-2 in human and mouse brain. J. Exp. Med. 2021, 218, e20202135. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Chen, X.; Cai, Y.; Zhou, X.; Xu, S.; Huang, H.; Zhang, L.; Zhou, X.; Du, C.; Zhang, Y. Risk factors associated with acute respiratory distress syndrome and death in patients with coronavirus disease 2019 pneumonia in Wuhan, China. JAMA Intern. Med. 2020, 180, 934–943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Brien, C.; Van Wyk, B.-E.; Van Heerden, F. Physical and chemical characteristics of Aloe ferox leaf gel. South Afr. J. Bot. 2011, 77, 988–995. [Google Scholar] [CrossRef] [Green Version]
- Šírová, K.; Vaculík, M. Toxic effects of cadmium on growth of Aloe ferox Mill. South Afr. J. Bot. 2021. [Google Scholar] [CrossRef]
- Galli, C.L.; Cinelli, S.; Ciliutti, P.; Melzi, G.; Marinovich, M. Aloe-emodin, a hydroxyanthracene derivative, is not genotoxic in an in vivo comet test. Regul. Toxicol. Pharmacol. 2021, 124, 104967. [Google Scholar] [CrossRef]
- Nxumalo, C.I.; Ngidi, L.S.; Shandu, J.S.E.; Maliehe, T.S. Isolation of endophytic bacteria from the leaves of Anredera cordifolia CIX1 for metabolites and their biological activities. BMC Complementary Med. Ther. 2020, 20, 300. [Google Scholar] [CrossRef]
- Lu, Y.; Chen, C.; Chen, H.; Zhang, J.; Chen, W. Isolation and identification of endophytic fungi from Actinidia macrosperma and investigation of their bioactivities. Evid. Based Complementary Altern. Med. 2012, 2012, 382742. [Google Scholar] [CrossRef] [Green Version]
- Dudeja, S.; Chhokar, V.; Badgujjar, H.; Chauhan, R.; Soni, S.; Beniwal, V.; Kumar, A. Isolation and screening of antibiotic producing fungi from solid-state waste. Polymorphism 2020, 4, 59–71. [Google Scholar]
- Darshit, R.; Pandya, D.D. Screening and characteristic study of antimicrobial actinomycetes from near-by soil of medicinal plants. Int. J. Pharm. Pharma. Sci. 2018, 10, 66. [Google Scholar] [CrossRef]
- Pryce, T.; Palladino, S.; Kay, I.; Coombs, G. Rapid identification of fungi by sequencing the ITS1 and ITS2 regions using an automated capillary electrophoresis system. Med Mycol. 2003, 41, 369–381. [Google Scholar] [CrossRef] [PubMed]
- Sharma, D.; Pramanik, A.; Agrawal, P.K. Evaluation of bioactive secondary metabolites from endophytic fungus Pestalotiopsis neglecta BAB-5510 isolated from leaves of Cupressus torulosa D. Don. 3 Biotech 2016, 6, 210. [Google Scholar] [CrossRef] [Green Version]
- Eloff, J.N. A sensitive and quick microplate method to determine the minimal inhibitory concentration of plant extracts for bacteria. Planta medica 1998, 64, 711–713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nkala, B.A. The Cytotoxic Effects, Anti-Iflammatory, Antioxidant, Antibacterial, and Antidiabetic Properties of Eight Selected South African Plants for Medicinal Purposes. Ph.D. Thesis, University of KwaZulu Natal, Durban, South Africa, 2020. [Google Scholar]
- Guo, F.; Chen, Q.; Liang, Q.; Zhang, M.; Chen, W.; Chen, H.; Yun, Y.; Zhong, Q.; Chen, W. Antimicrobial activity and proposed action mechanism of linalool against pseudomonas fluorescens. Front. Microbiol. 2021, 12, 49. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Wang, J.-Q.; Song, H.-Y.; Zhang, Q.; Liu, G.-F. Chemical analysis and in vitro antimicrobial effects and mechanism of action of Trachyspermum copticum essential oil against Escherichia coli. Asian Pac. J. Trop. Med. 2017, 10, 663–669. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Khan, M.R.; Sajid, M.; Zahra, Z. Phytochemical investigation and antimicrobial appraisal of Parrotiopsis jacquemontiana (Decne) Rehder. BMC Complementary Altern. Med. 2018, 18, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ngema, S.; Basson, A.; Maliehe, T. Synthesis, characterization and application of polyacrylamide grafted bioflocculant. Phys. Chem. Earth Parts A/B/C 2020, 115, 102821. [Google Scholar] [CrossRef]
- Yadav, G.; Meena, M. Bioprospecting of endophytes in medicinal plants of Thar Desert: An attractive resource for biopharmaceuticals. Biotechnol. Rep. 2021, 30, e00629. [Google Scholar] [CrossRef]
- Vesth, T.C.; Nybo, J.L.; Theobald, S.; Frisvad, J.C.; Larsen, T.O.; Nielsen, K.F.; Hoof, J.B.; Brandl, J.; Salamov, A.; Riley, R. Investigation of inter-and intraspecies variation through genome sequencing of Aspergillus section Nigri. Nat. Genet. 2018, 50, 1688–1695. [Google Scholar] [CrossRef] [Green Version]
- Ravindran, C.; Naveenan, T.; Varatharajan, G.R.; Rajasabapathy, R.; Meena, R.M. Antioxidants in mangrove plants and endophytic fungal associations. Bot. Mar. 2012, 55, 269–279. [Google Scholar] [CrossRef]
- Da Silva, I.P.; Brissow, E.; Kellner Filho, L.C.; Senabio, J.; de Siqueira, K.A.; Vandresen Filho, S.; Damasceno, J.L.; Mendes, S.A.; Tavares, D.C.; Magalhães, L.G. Bioactive compounds of Aspergillus terreus—F7, an endophytic fungus from Hyptis suaveolens (L.) Poit. World J. Microbiol. Biotechnol. 2017, 33, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Sornakili, A.; Thankappan, S.; Sridharan, A.; Nithya, P.; Uthandi, S. Antagonistic fungal endophytes and their metabolite-mediated interactions against phytopathogens in rice. Physiol. Mol. Plant Pathol. 2020, 112, 101525. [Google Scholar] [CrossRef]
- Liu, J.; Song, Y.; Zhang, Z.; Wang, L.; Guo, Z.; Zou, W.; Tan, R. Aspergillus fumigatus CY018, an endophytic fungus in Cynodon dactylon as a versatile producer of new and bioactive metabolites. J. Biotechnol. 2004, 114, 279–287. [Google Scholar] [CrossRef]
- Almaary, K.S.; Alharbi, N.S.; Kadaikunnan, S.; Khaled, J.M.; Rajivgandhi, G.; Ramachandran, G.; Kanisha, C.C.; Murugan, M.; Alanzi, K.F.; Manoharan, N. Anti-bacterial effect of marine sea grasses mediated endophytic actinomycetes against K. pneumoniae. J. King Saud Univ. Sci. 2021, 33, 101528. [Google Scholar] [CrossRef]
- Quintanilha-Peixoto, G.; Torres, R.O.; Reis, I.M.A.; Oliveira, T.A.S.d.; Bortolini, D.E.; Duarte, E.A.A.; Ariston de Carvalho Azevedo, V.; Brenig, B.; Aguiar, E.R.G.R.; Soares, A.C.F. Calm before the storm: A glimpse into the secondary metabolism of Aspergillus welwitschiae, the etiologic agent of the sisal bole rot. Toxins 2019, 11, 631. [Google Scholar] [CrossRef] [Green Version]
- Han, J.; Yang, N.; Wei, S.; Jia, J.; Lin, R.; Li, J.; Bi, H.; Song, F.; Xu, X. Dimeric hexylitaconic acids from the marine-derived fungus Aspergillus welwitschiae CUGBMF180262. Nat. Prod. Res. 2020, 36, 1–8. [Google Scholar] [CrossRef]
- Loges, L.A.; Silva, D.B.; Paulino, G.V.; Landell, M.F.; Macedo, A.J. Polyketides from marine-derived Aspergillus welwitschiae inhibit Staphylococcus aureus virulence factors and potentiate vancomycin antibacterial activity in vivo. Microb. Pathog. 2020, 143, 104066. [Google Scholar] [CrossRef]
- Ying, L.; Zhong, D.; Peng, D.-L.; LIU, S.-M.; Kong, L.-A.; Huan, P.; Xiang, C.; LI, Z.-C.; Huang, W.-K. Evaluation of the biocontrol potential of Aspergillus welwitschiae against the root-knot nematode Meloidogyne graminicola in rice (Oryza sativa L.). J. Integr. Agric. 2019, 18, 2561–2570. [Google Scholar]
- El-Naggar, N.E.-A.; El-Shweihy, N.M. Bioprocess development for L-asparaginase production by Streptomyces rochei, purification and in-vitro efficacy against various human carcinoma cell lines. Sci. Rep. 2020, 10, 1–21. [Google Scholar] [CrossRef]
- Vanzela, D.O.A.; Massi, F.P.; de Oliveira, A.L.M.; Fungaro, M.H.P.; Sartori, D. Isolation and identification of aspergillus section Nigri, and genotype associated with ochratoxin A and fumonisin B 2 production in garlic marketed in Brazil. Curr. Microbiol. 2020, 77, 1–9. [Google Scholar] [CrossRef]
- Almeida, C.; Bills, G.; González-Menéndez, V.; Martin, J.; Tormo, J.R.; Genilloud, O. Time-dependent production of the bioactive peptides endolides A and B and the polyketide mariline A from the sponge-derived fungus Stachylidium bicolor 293K04. Fermentation 2017, 3, 45. [Google Scholar] [CrossRef] [Green Version]
- Khalil, Z.G.; Kalansuriya, P.; Capon, R.J. Lipopolysaccharide (lps) stimulation of fungal secondary metabolism. Mycology 2014, 5, 168–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdalla, M.A.; Aro, A.O.; Gado, D.; Passari, A.K.; Mishra, V.K.; Singh, B.P.; McGaw, L.J. Isolation of endophytic fungi from South African plants, and screening for their antimicrobial and extracellular enzymatic activities and presence of type I polyketide synthases. South Afr. J. Bot. 2020, 134, 336–342. [Google Scholar] [CrossRef]
- Asong, J.A.; Amoo, S.O.; McGaw, L.J.; Nkadimeng, S.M.; Aremu, A.O.; Otang-Mbeng, W. Antimicrobial activity, antioxidant potential, cytotoxicity and phytochemical profiling of four plants locally used against skin diseases. Plants 2019, 8, 350. [Google Scholar] [CrossRef] [Green Version]
- Elisha, I.L.; Botha, F.S.; McGaw, L.J.; Eloff, J.N. The antibacterial activity of extracts of nine plant species with good activity against Escherichia coli against five other bacteria and cytotoxicity of extracts. BMC Complementary Altern. Med. 2017, 17, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Dejoies, L.; Le Neindre, K.; Reissier, S.; Felden, B.; Cattoir, V. Distinct expression profiles of regulatory RNAs in the response to biocides in Staphylococcus aureus and Enterococcus faecium. Sci. Rep. 2021, 11, 6892. [Google Scholar] [CrossRef]
- Liu, C.H.; Zhang, E.; Wong, G.T.F.; Hyun, S. Factors associated with depression, anxiety, and PTSD symptomatology during the COVID-19 pandemic: Clinical implications for US young adult mental health. Psychiatry Res. 2020, 290, 113172. [Google Scholar] [CrossRef]
- Gomaa, E.Z. Silver nanoparticles as an antimicrobial agent: A case study on Staphylococcus aureus and Escherichia coli as models for Gram-positive and Gram-negative bacteria. J. Gen. Appl. Microbiol. 2017, 63, 36–43. [Google Scholar] [CrossRef] [Green Version]
- Dong, S.; Yang, X.; Zhao, L.; Zhang, F.; Hou, Z.; Xue, P. Antibacterial activity and mechanism of action saponins from Chenopodium quinoa Willd. husks against foodborne pathogenic bacteria. Ind. Crop. Prod. 2020, 149, 112350. [Google Scholar] [CrossRef]
- Tang, H.; Chen, W.; Dou, Z.-M.; Chen, R.; Hu, Y.; Chen, W.; Chen, H. Antimicrobial effect of black pepper petroleum ether extract for the morphology of Listeria monocytoge nes and Salmonella typhimurium. J. Food Sci. Technol. 2017, 54, 2067–2076. [Google Scholar] [CrossRef]
- Jayaram, A.; Deer, E.; Amaral, L.M.; Campbell, N.; Vaka, V.R.; Cunningham, M.; Ibrahim, T.; Cornelius, D.C.; LaMarca, B.B. The role of tumor necrosis factor in triggering activation of natural killer cell, multi-organ mitochondrial dysfunction and hypertension during pregnancy. Pregnancy Hypertens. 2021, 24, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Alibi, S.; Ramos-Vivas, J.; Selma, W.B.; Mansour, H.B.; Boukadida, J.; Navas, J. Virulence of clinically relevant multidrug resistant Corynebacterium striatum strains and their ability to adhere to human epithelial cells and inert surfaces. Microb. Pathog. 2021, 155, 104887. [Google Scholar] [CrossRef] [PubMed]
- Pandey, D.; Pandey, V.C. Sacred plants from ancient to modern era: Traditional worshipping towards plants conservation. Trop. Plant Res. 2016, 3, 136–141. [Google Scholar]
- Mahale, S.; Goswami-Giri, A. Composition and characterization of refined oil compared with its crude oil from waste obtained from Mangifera indica. Asian J. Research Chem. 2011, 4, 1415–1419. [Google Scholar]
- Al-Majedy, Y.K.; Kadhum, A.A.H.; Al-Amiery, A.A.; Mohamad, A.B. Coumarins: The antimicrobial agents. Syst. Rev. Pharm. 2017, 8, 62. [Google Scholar] [CrossRef]
- Abd Hamid, H.; Mutazah, R.; Yusoff, M.M.; Abd Karim, N.A.; Razis, A.F.A. Comparative analysis of antioxidant and antiproliferative activities of Rhodomyrtus tomentosa extracts prepared with various solvents. Food Chem. Toxicol. 2017, 108, 451–457. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.-L.; Zhang, L.-F.; Xu, J.-G. Chemical composition, antibacterial activity and action mechanism of different extracts from hawthorn (Crataegus pinnatifida Bge.). Sci. Rep. 2020, 10, 1–13. [Google Scholar]
- Burman, S.; Bhattacharya, K.; Mukherjee, D.; Chandra, G. Antibacterial efficacy of leaf extracts of Combretum album Pers. against some pathogenic bacteria. BMC Complementary Altern. Med. 2018, 18, 213. [Google Scholar] [CrossRef] [Green Version]
- Dhivya, S.; Kalaichelvi, K. Phytochemical studies and gas chromatography-mass spectrometry analysis of sarcostemma brevistigma, wight & arn. Asian J. Pharm. Clin. Res. 2017, 10, 462–466. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primary Screening | Secondary Screening | |||||
|---|---|---|---|---|---|---|
| Isolates | S. aureus | E. coli | S. aureus | E. coli | Fungal Name | GenBank Accession Number |
| FUN01 | +++ | +++ | +++ | ++ | Aspergillus welwitschiae | MH545928.1 |
| FUN02 | +++ | ++ | ++ | − | Un-identified | Un-identified |
| FUN03 | +++ | ++ | + | − | Un-identified | Un-identified |
| FUN04 | ++ | + | + | − | Un-identified | Un-identified |
| FUN05 | ++ | + | + | − | Un-identified | Un-identified |
| FUN06 | ++ | − | + | − | Un-identified | Un-identified |
| FUN07 | + | − | − | − | Un-identified | Un-identified |
| FUN08 | − | − | Un-identified | Un-identified | ||
| FUN09 | − | − | Un-identified | Un-identified | ||
| FUN010 | − | − | Un-identified | Un-identified | ||
| FUN011 | − | − | Un-identified | Un-identified | ||
| FUN012 | − | − | Un-identified | Un-identified | ||
| FUN013 | − | − | Un-identified | Un-identified | ||
| FUN014 | − | − | Un-identified | Un-identified | ||
| FUN015 | − | − | Un-identified | Un-identified | ||
| Bacteria | Extract | Ciprofloxacin | ||
|---|---|---|---|---|
| MIC (mg/mL) | MBC (mg/mL) | MIC (µg/mL) | MBC (µg/mL) | |
| S. aureus (ATCC 25925) | 1 | >2 | 0.015 | 0.031 |
| E. coli (ATCC 25922) | 1 | >2 | 0.015 | 0.031 |
| Compounds | Presence/Absence of Compounds |
|---|---|
| Terpenoids | + |
| Flavonoids | − |
| Cardiac glycosides | + |
| Coumarins | + |
| Saponins | − |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maliehe, T.S.; Mbambo, M.; Nqotheni, M.I.; Senzo, N.S.; Shandu, J.S.E. Antibacterial Effect and Mode of Action of Secondary Metabolites from Fungal Endophyte Associated with Aloe ferox Mill. Microbiol. Res. 2022, 13, 90-101. https://doi.org/10.3390/microbiolres13010007
Maliehe TS, Mbambo M, Nqotheni MI, Senzo NS, Shandu JSE. Antibacterial Effect and Mode of Action of Secondary Metabolites from Fungal Endophyte Associated with Aloe ferox Mill. Microbiology Research. 2022; 13(1):90-101. https://doi.org/10.3390/microbiolres13010007
Chicago/Turabian StyleMaliehe, Tsolanku Sidney, Melusi Mbambo, Mduduzi Innocent Nqotheni, Ngema Siyanda Senzo, and Jabulani Siyabonga Emmanuel Shandu. 2022. "Antibacterial Effect and Mode of Action of Secondary Metabolites from Fungal Endophyte Associated with Aloe ferox Mill" Microbiology Research 13, no. 1: 90-101. https://doi.org/10.3390/microbiolres13010007
APA StyleMaliehe, T. S., Mbambo, M., Nqotheni, M. I., Senzo, N. S., & Shandu, J. S. E. (2022). Antibacterial Effect and Mode of Action of Secondary Metabolites from Fungal Endophyte Associated with Aloe ferox Mill. Microbiology Research, 13(1), 90-101. https://doi.org/10.3390/microbiolres13010007