
Promotion of Peanut (Arachis hypogaea L.) Growth by Plant Growth-Promoting Microorganisms


 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of the Pots in the Greenhouse
2.2. Experimental Design in the Field
2.3. Inoculum Preparation and Applications
2.4. Experimental Design in the Greenhouse
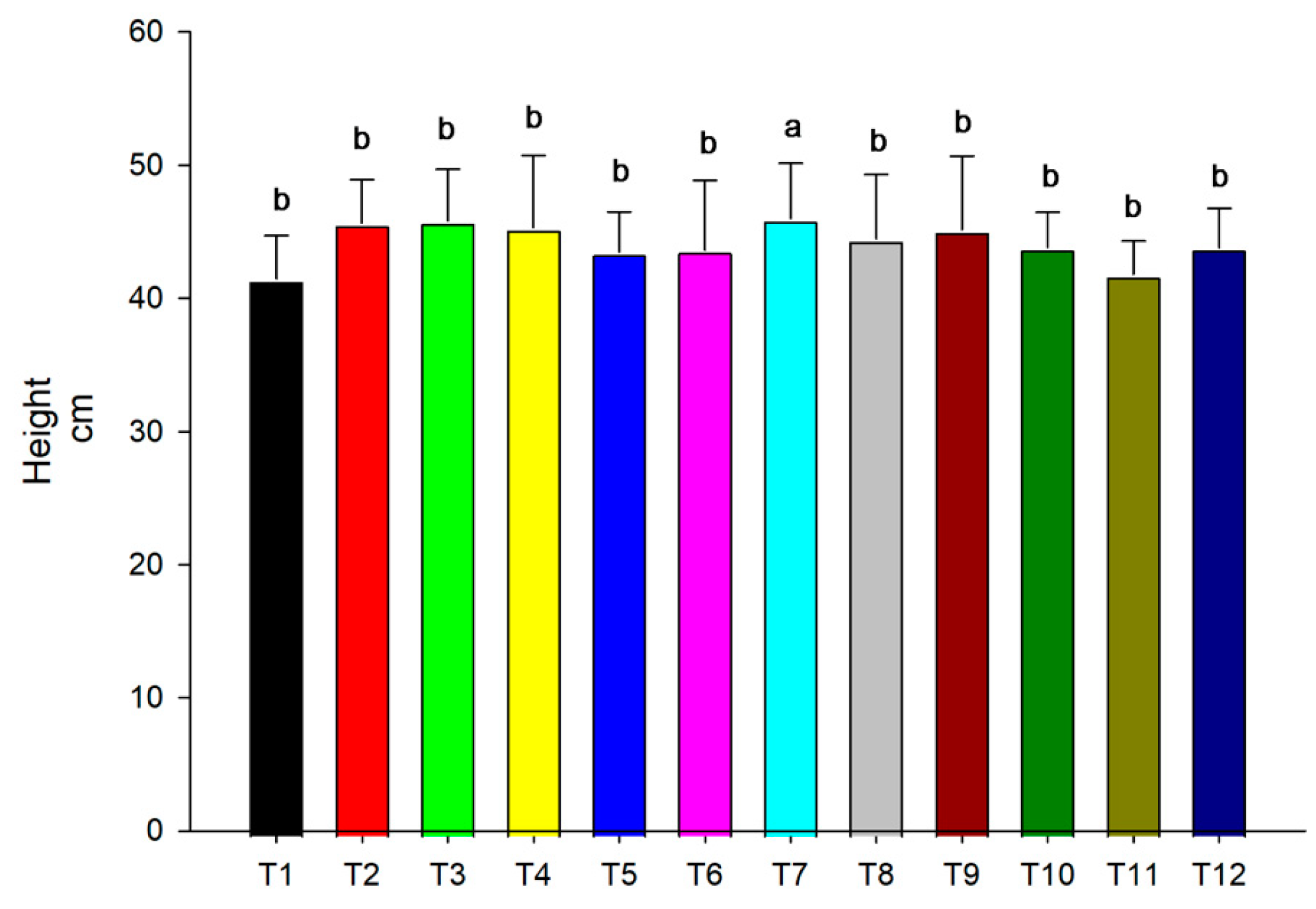
2.5. Plant Height
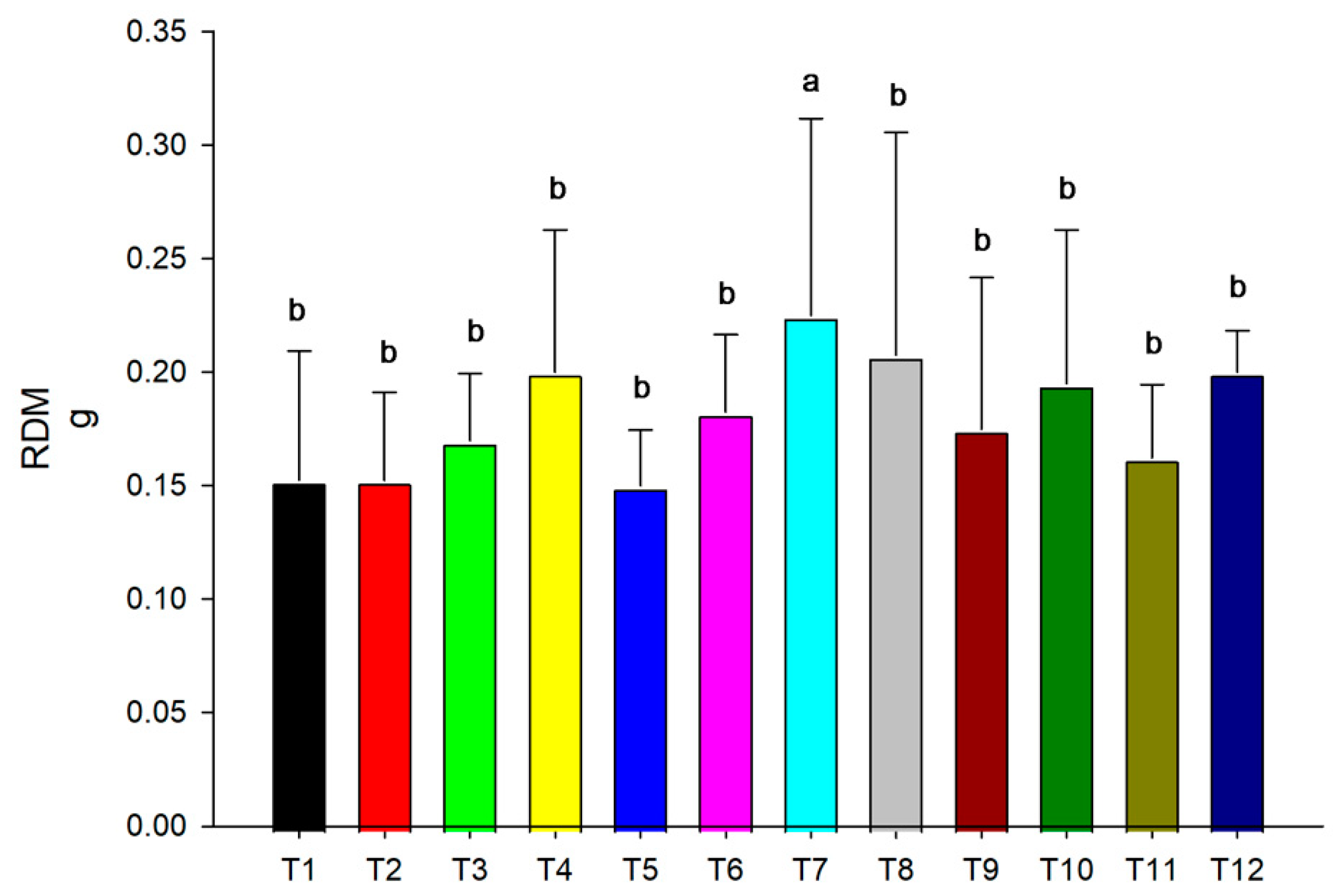
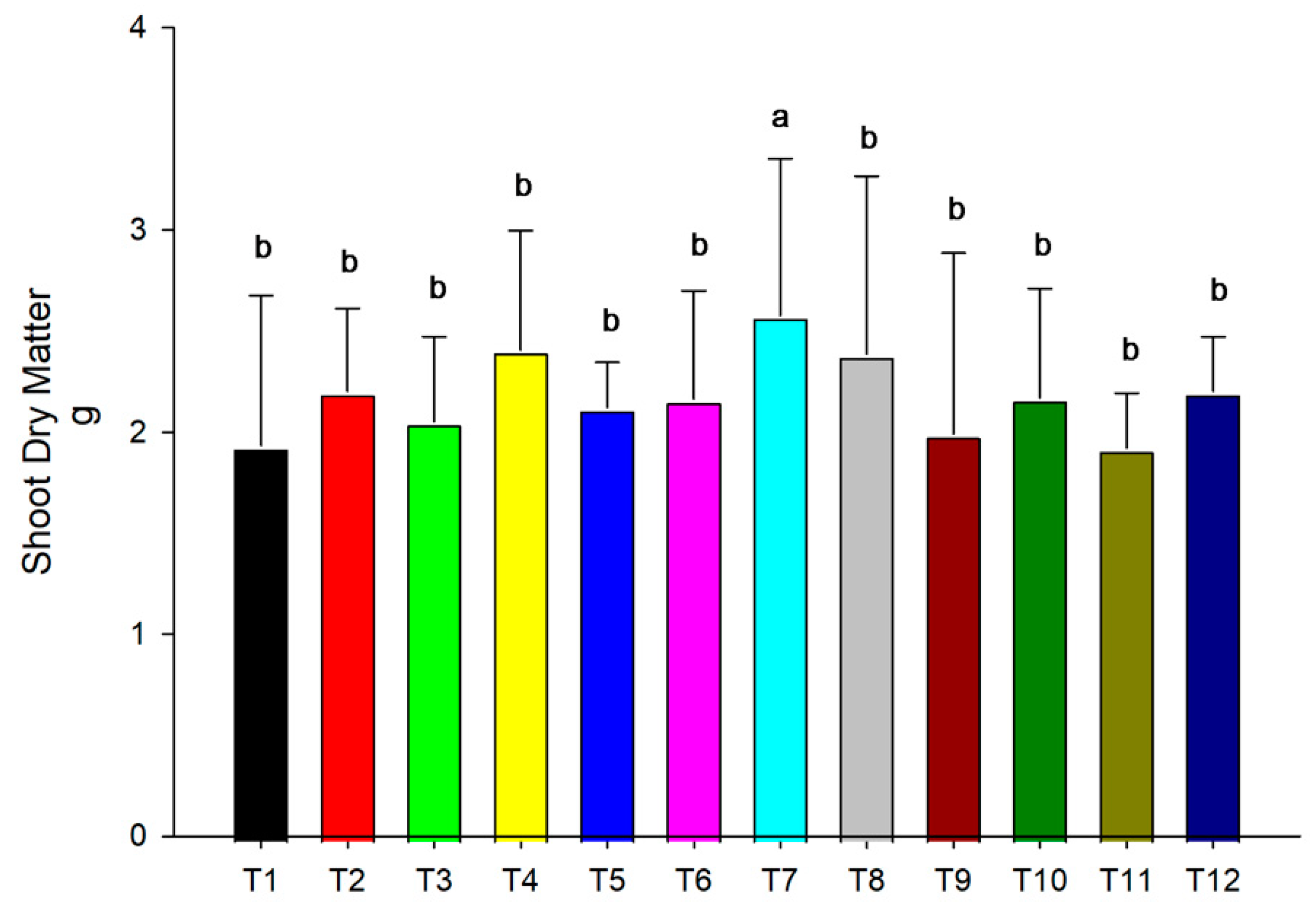
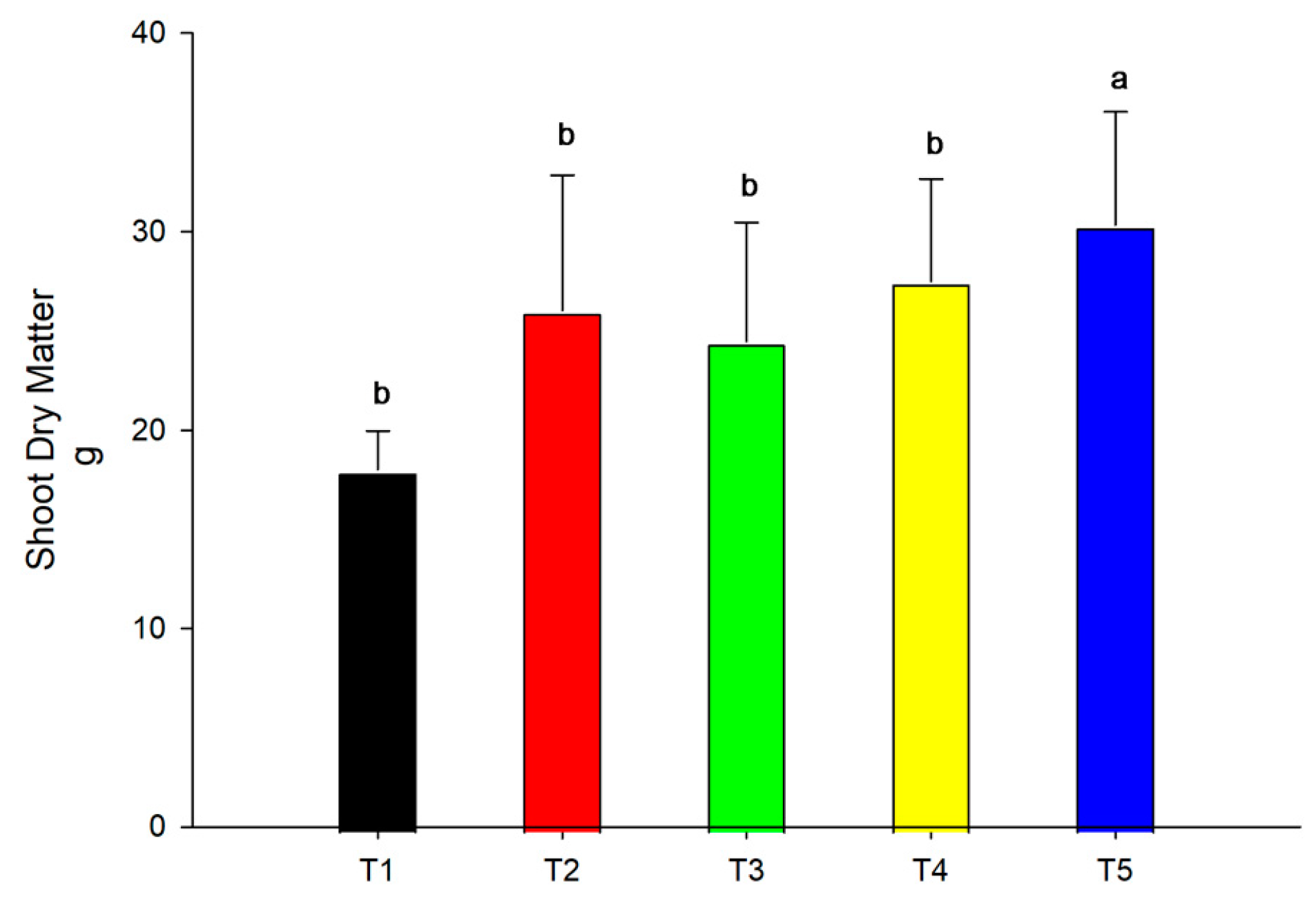
2.6. Dry Matter
2.7. Nitrogen and Phosphorus Concentration
2.8. Urease and Acid Phosphatase Enzymatic Activity
2.9. Productivity
2.10. Analysis of Results
3. Results
3.1. Dry Matter of Plants
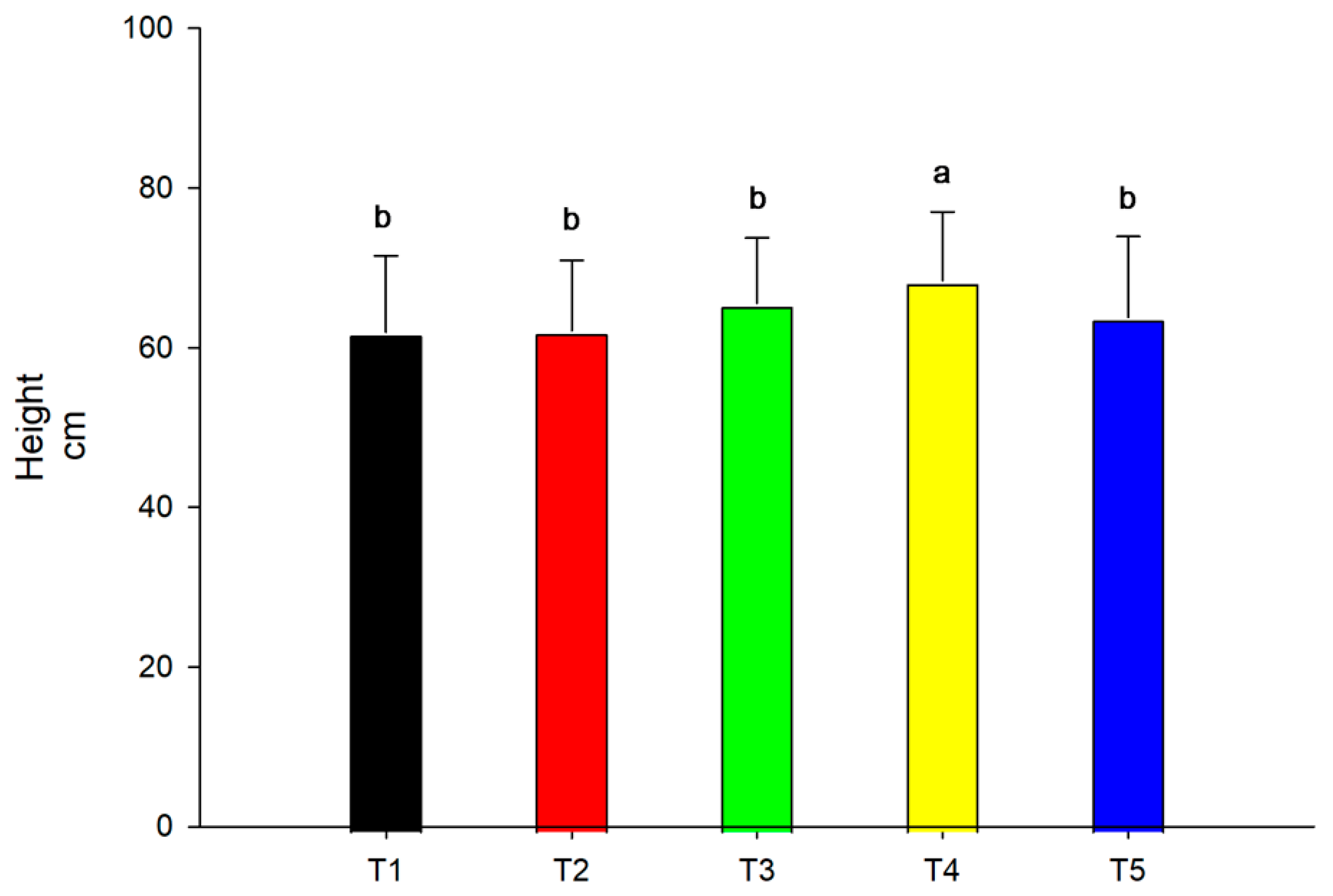
3.2. Height
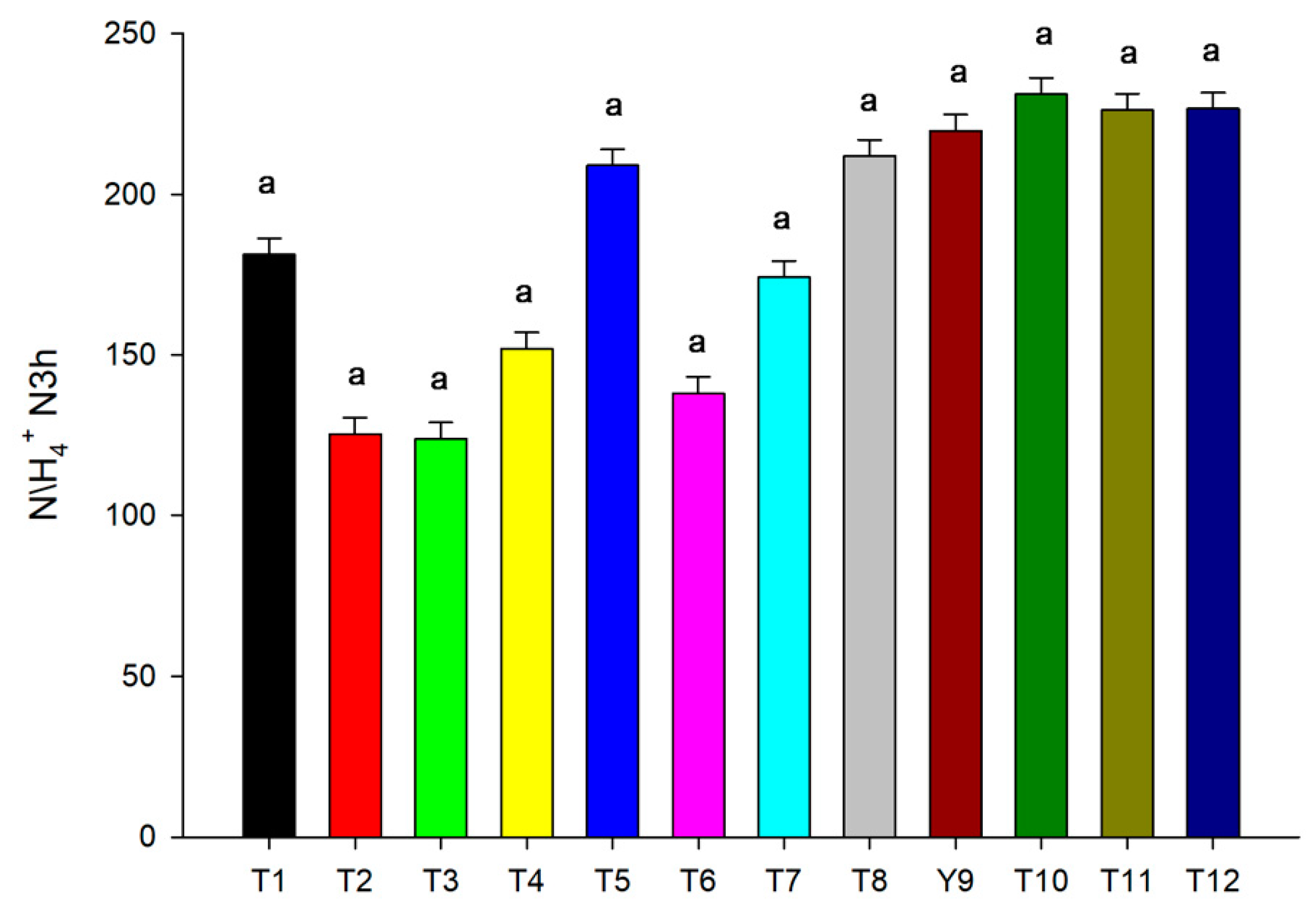
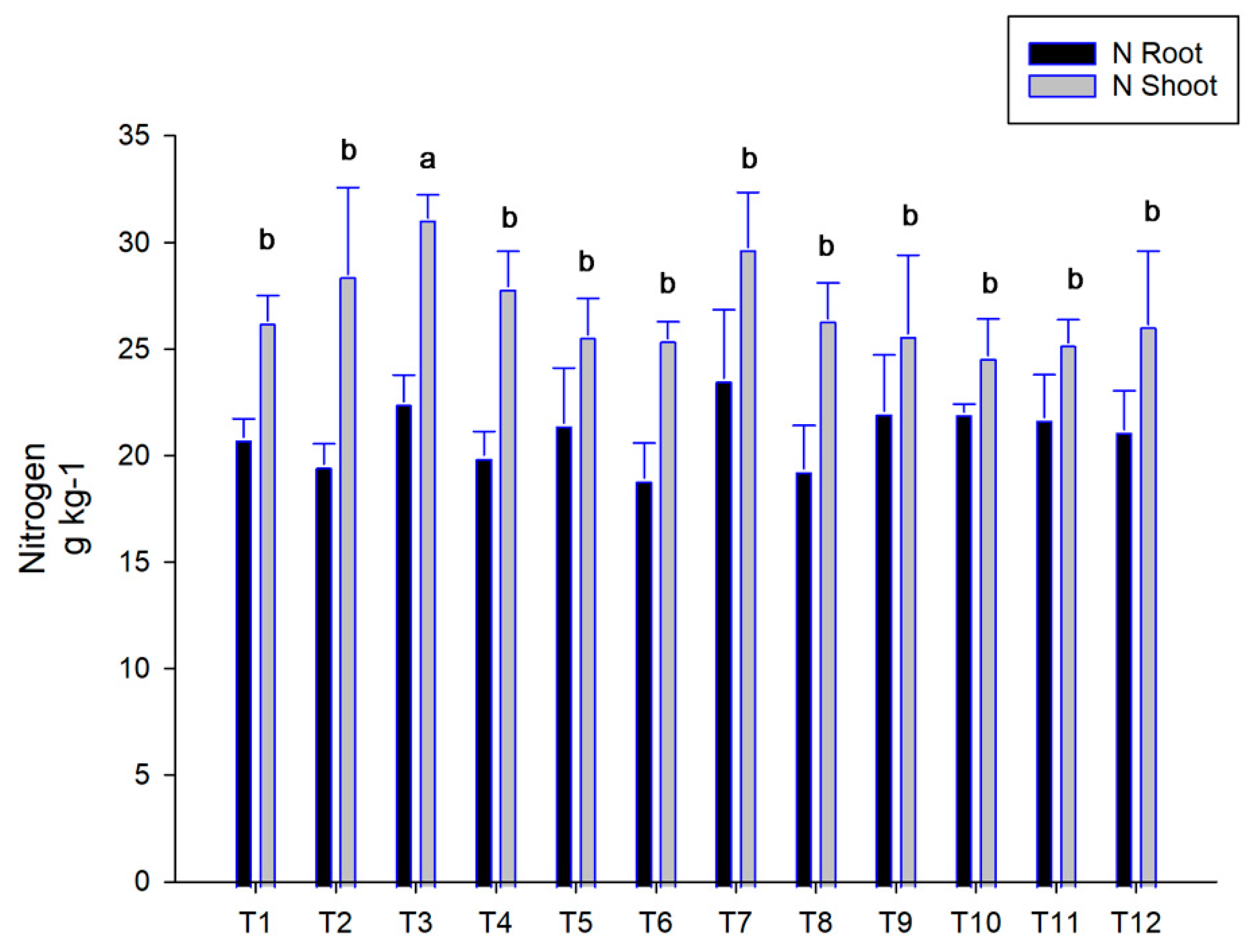
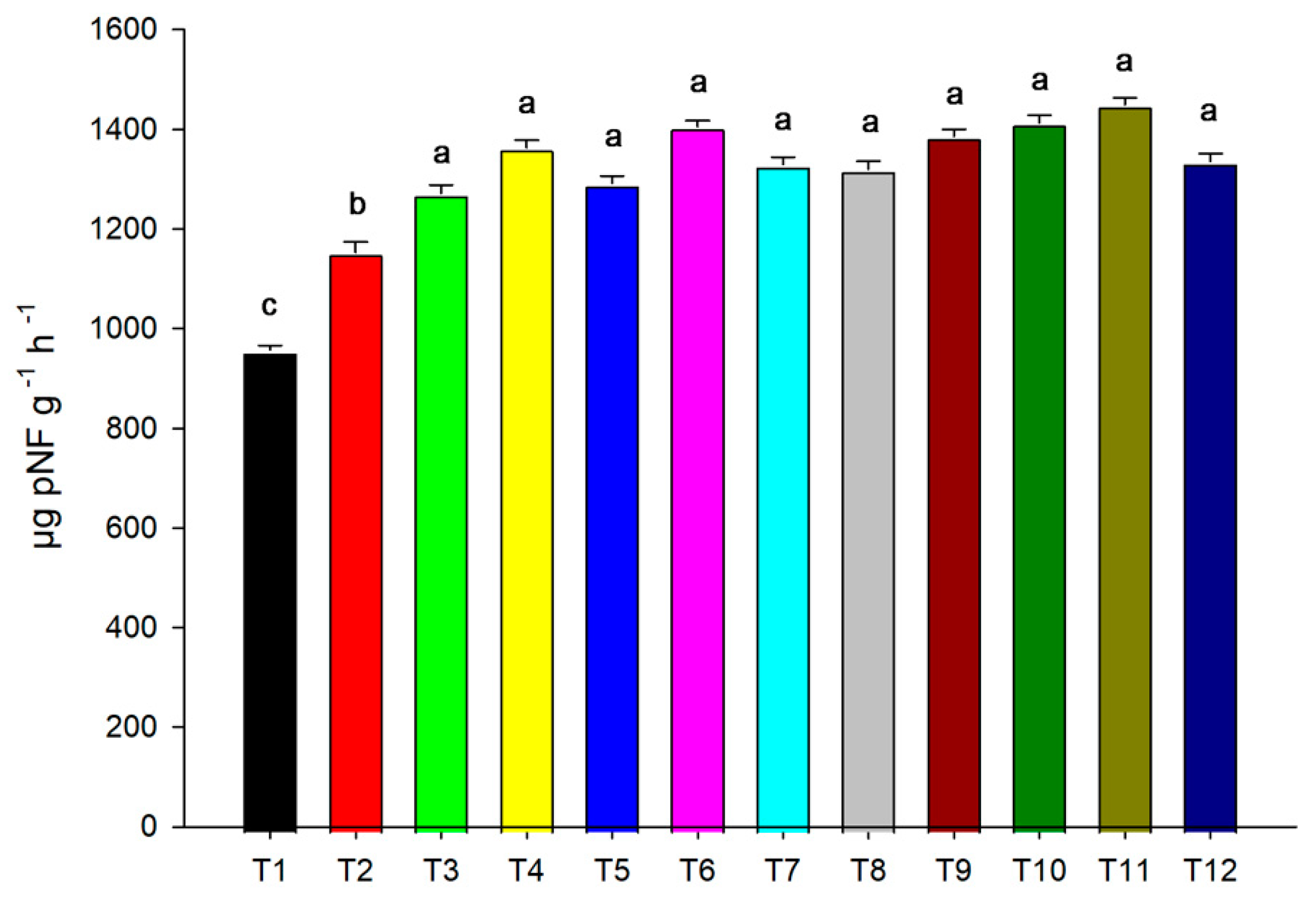
3.3. Urease and Nitrogen Concentration
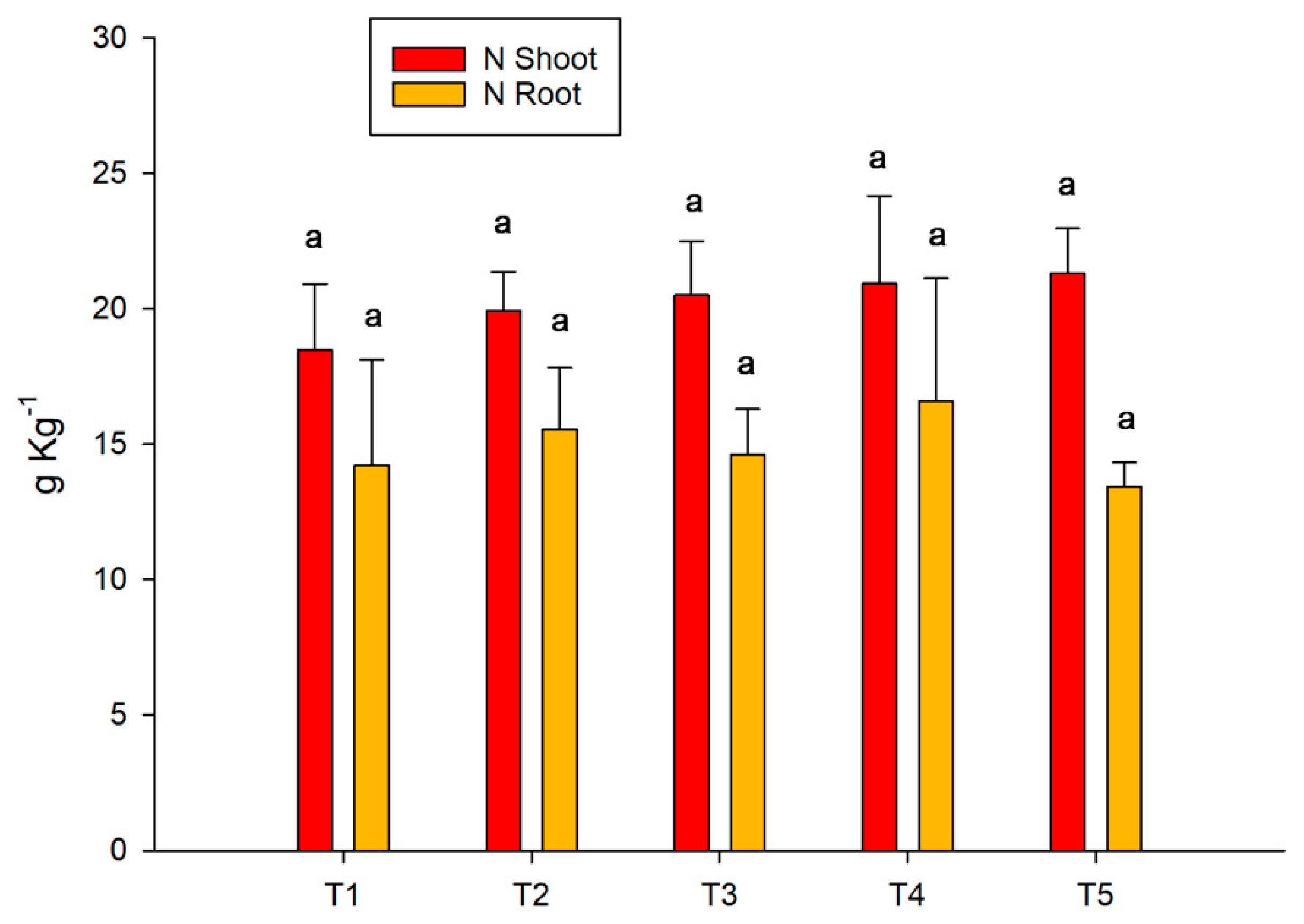
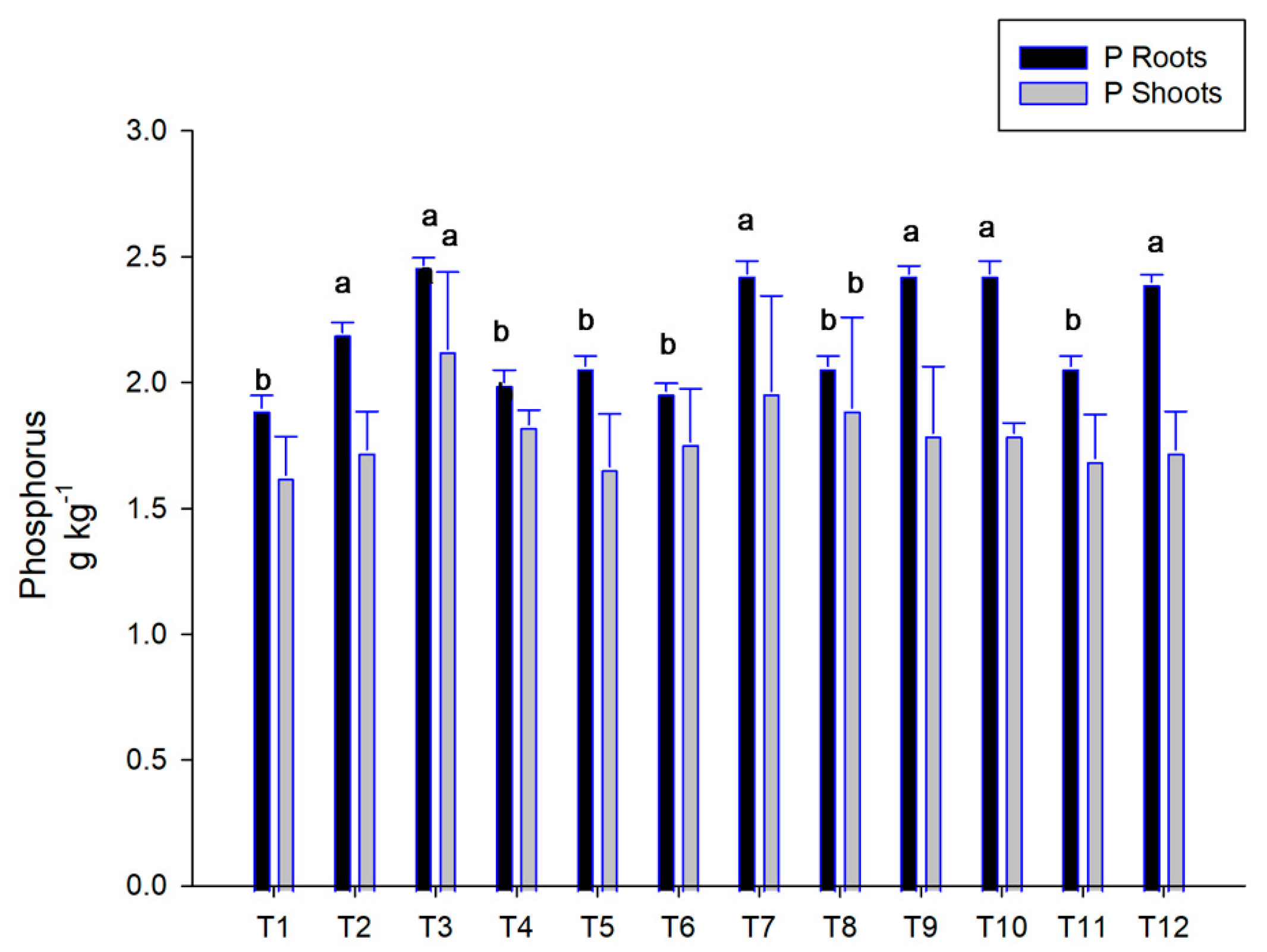
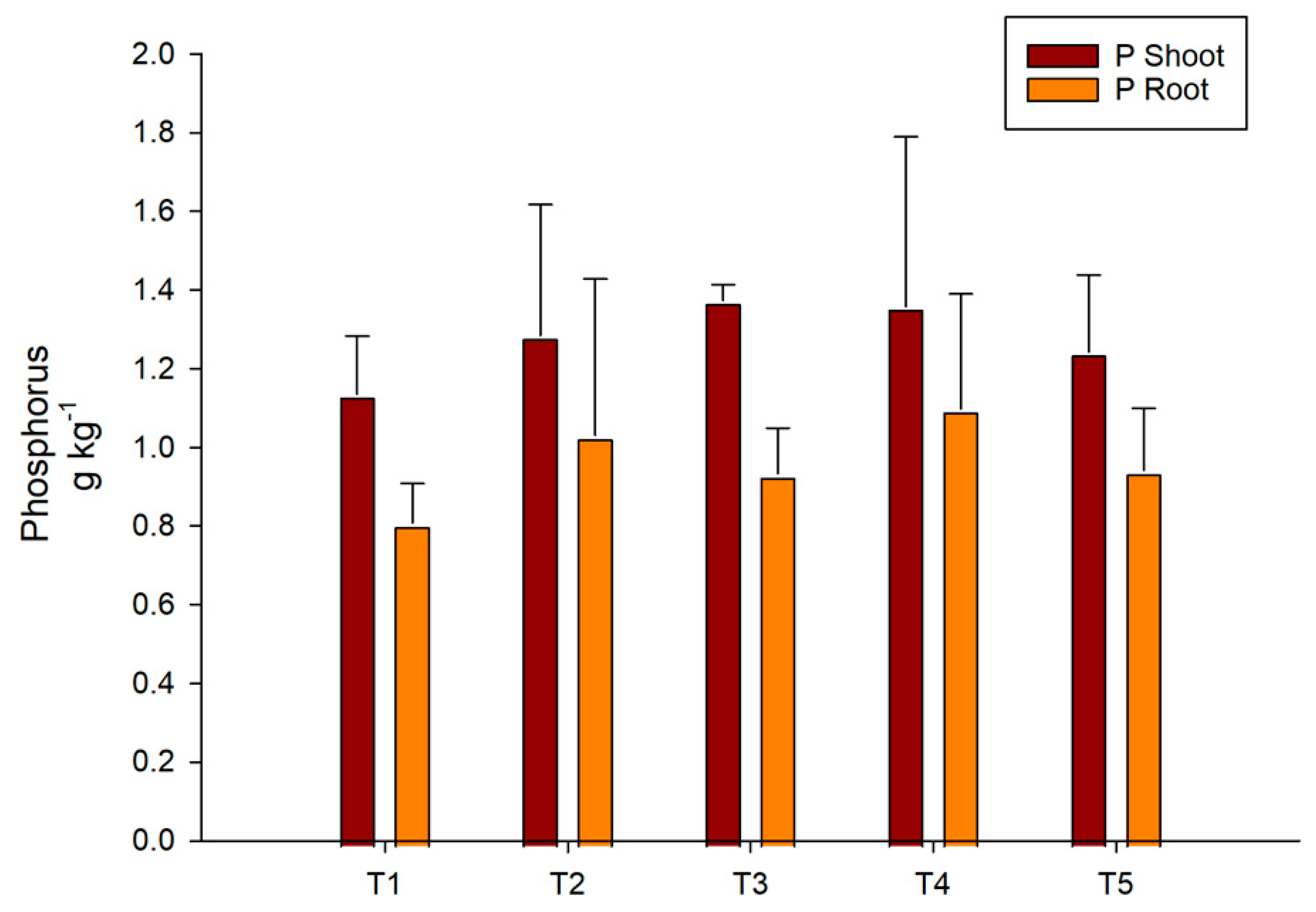
3.4. Acid Phosphatase and Phosphorus Concentration
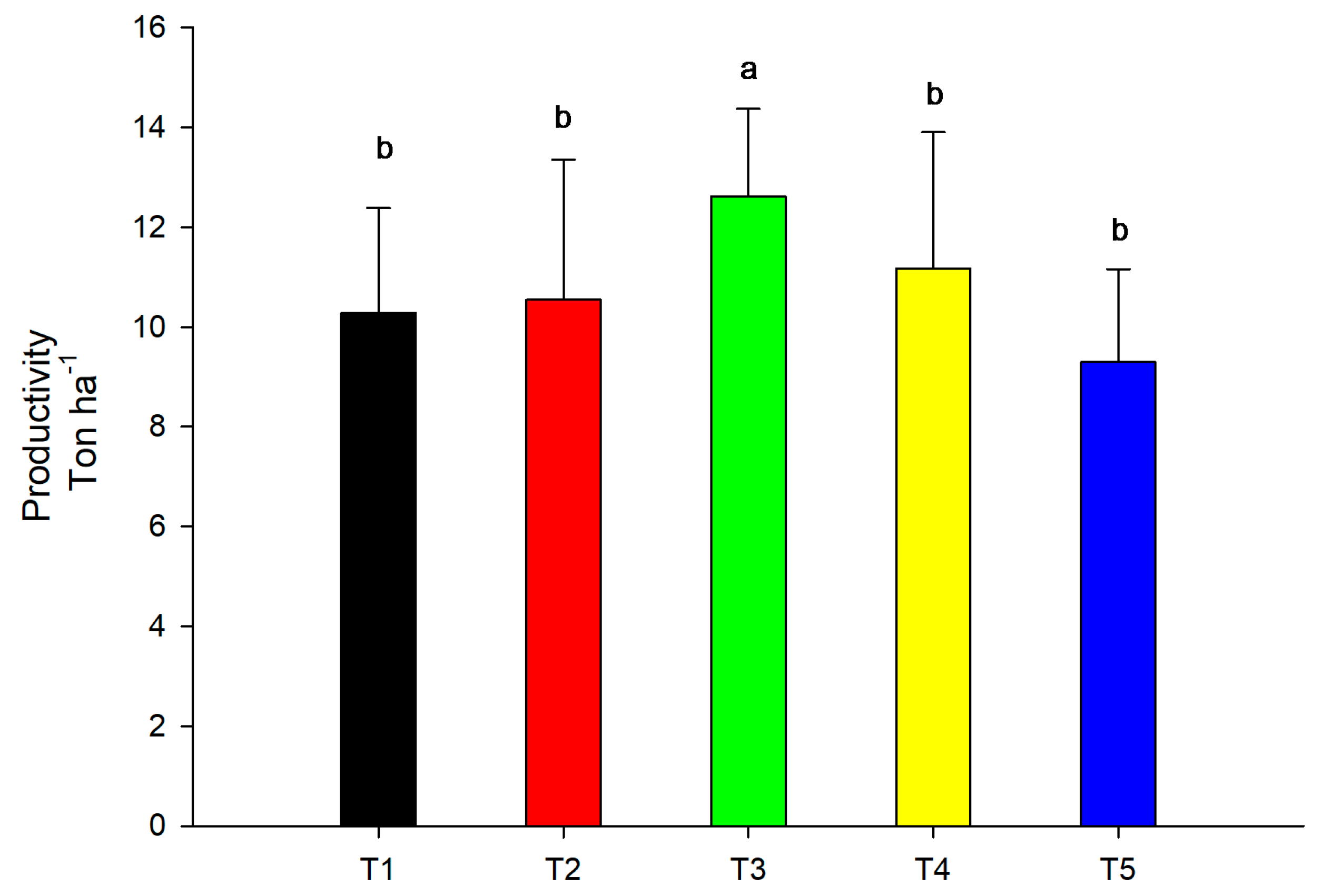
3.5. Productivity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rao, V.T.; Venkanna, V.; Bhadru, D.; Bharathi, D. Studies on variability, character association and path analysis on groundnut (Arachis hypogaea L.). Int. J. Pure Appl. Biosci. 2014, 2, 194–197. [Google Scholar]
- Hamza, M.; Abbas, M.; Abd Elrahman, A.; Helal, M.; Shahba, M. Conventional versus nano calcium forms on peanut production under sandy soil conditions. Agriculture 2021, 11, 767. [Google Scholar] [CrossRef]
- Zhao, X.; Chen, J.; Du, F. Potential use of peanut by-products in food processing: A review. J. Food Sci. Technol. 2012, 49, 521–529. [Google Scholar] [CrossRef]
- Diarra, K.; Nong, Z.G.; Jie, C. Peanut milk and peanut milk based products production: A review. Crit. Rev. Food Sci. Nutr. 2005, 45, 405–423. [Google Scholar] [CrossRef]
- Wang, S.; Hu, Z.; Chen, Y.; Wu, H.; Wang, Y.; Wu, F.; Gu, F. Integration of agricultural machinery and agronomy for mechanised peanut production using the vine for animal feed. Biosyst. Eng. 2022, 219, 135–152. [Google Scholar] [CrossRef]
- Das, A.; Goyali, J.C.; Ferdausi, A. Tissue culture approaches to improve nutritional quality and stress response in peanut. Eur. J. Biol. Res. 2021, 11, 332–347. [Google Scholar]
- Babalola, O.O. Beneficial bacteria of agricultural importance. Biotechnol. Lett. 2010, 32, 1559–1570. [Google Scholar] [CrossRef] [PubMed]
- Patel, T.; Minocheherhomji, F. Plant growth promoting Rhizobacteria: Blessing to agriculture. Int. J. Pure Appl. Biosci. 2018, 6, 481–492. [Google Scholar] [CrossRef]
- Thiebaut, F.; Urquiaga, M.C.d.O.; Rosman, A.C.; da Silva, M.L.; Hemerly, A.S. The Impact of Non-Nodulating Diazotrophic Bacteria in Agriculture: Understanding the Molecular Mechanisms That Benefit Crops. Int. J. Mol. Sci. 2022, 23, 11301. [Google Scholar] [CrossRef] [PubMed]
- Pankievicz, V.C.S.; do Amaral, F.P.; Ané, J.-M.; Stacey, G. Diazotrophic bacteria and their mechanisms to interact and benefit cereals. Mol. Plant-Microbe Interact. 2021, 34, 491–498. [Google Scholar] [CrossRef]
- Kubar, M.S.; Feng, M.; Sayed, S.; Shar, A.H.; Rind, N.A.; Ullah, H.; Kalhoro, S.A.; Xie, Y.; Yang, C.; Yang, W. Agronomical traits associated with yield and yield components of winter wheat as affected by nitrogen managements. Saudi J. Biol. Sci. 2021, 28, 4852–4858. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.; Chen, D.; Wu, F.; Tang, L.; He, S.; Zhou, W. Function of aquatic plants on nitrogen removal and greenhouse gas emission in enhanced denitrification constructed wetlands: Iris pseudacorus for example. J. Clean. Prod. 2022, 330, 129842. [Google Scholar] [CrossRef]
- Cui, X.; Guo, L.; Li, C.; Liu, M.; Wu, G.; Jiang, G. The total biomass nitrogen reservoir and its potential of replacing chemical fertilizers in China. Renew. Sustain. Energy Rev. 2021, 135, 110215. [Google Scholar] [CrossRef]
- de França, A.A.; von Tucher, S.; Schmidhalter, U. Effects of combined application of acidified biogas slurry and chemical fertilizer on crop production and N soil fertility. Eur. J. Agron. 2021, 123, 126224. [Google Scholar] [CrossRef]
- Gao, Y.; Fang, Z.; Van Zwieten, L.; Bolan, N.; Dong, D.; Quin, B.F.; Meng, J.; Li, F.; Wu, F.; Wang, H. A critical review of biochar-based nitrogen fertilizers and their effects on crop production and the environment. Biochar 2022, 4, 36. [Google Scholar] [CrossRef]
- Liu, L.; Zheng, X.; Wei, X.; Kai, Z.; Xu, Y. Excessive application of chemical fertilizer and organophosphorus pesticides induced total phosphorus loss from planting causing surface water eutrophication. Sci. Rep. 2021, 11, 23015. [Google Scholar] [CrossRef]
- Li, F.; Jin, Y.; He, S.; Jin, J.; Wang, Z.; Khan, S.; Tian, G.; Liang, X. Use of polyacrylamide modified biochar coupled with organic and chemical fertilizers for reducing phosphorus loss under different cropping systems. Agric. Ecosyst. Environ. 2021, 310, 107306. [Google Scholar] [CrossRef]
- Meena, S.K.; Dwivedi, B.S.; Meena, M.C.; Datta, S.P.; Singh, V.K.; Mishra, R.P.; Chakraborty, D.; Dey, A.; Meena, V.S. Long-Term Nutrient Supply Options: Strategies to Improve Soil Phosphorus Availability in the Rice-Wheat System. Sustainability 2022, 14, 8629. [Google Scholar] [CrossRef]
- Basu, A.; Prasad, P.; Das, S.N.; Kalam, S.; Sayyed, R.; Reddy, M.; El Enshasy, H. Plant growth promoting rhizobacteria (PGPR) as green bioinoculants: Recent developments, constraints, and prospects. Sustainability 2021, 13, 1140. [Google Scholar] [CrossRef]
- Chandran, H.; Meena, M.; Swapnil, P. Plant growth-promoting rhizobacteria as a green alternative for sustainable agriculture. Sustainability 2021, 13, 10986. [Google Scholar] [CrossRef]
- Ahluwalia, O.; Singh, P.C.; Bhatia, R. A review on drought stress in plants: Implications, mitigation and the role of plant growth promoting rhizobacteria. Resour. Environ. Sustain. 2021, 5, 100032. [Google Scholar] [CrossRef]
- Yagmur, B.; Gunes, A. Evaluation of the effects of plant growth promoting rhizobacteria (PGPR) on yield and quality parameters of tomato plants in organic agriculture by principal component analysis (PCA). Gesunde Pflanz. 2021, 73, 219–228. [Google Scholar] [CrossRef]
- El-Sawah, A.M.; El-Keblawy, A.; Ali, D.F.I.; Ibrahim, H.M.; El-Sheikh, M.A.; Sharma, A.; Alhaj Hamoud, Y.; Shaghaleh, H.; Brestic, M.; Skalicky, M. Arbuscular mycorrhizal fungi and plant growth-promoting rhizobacteria enhance soil key enzymes, plant growth, seed yield, and qualitative attributes of guar. Agriculture 2021, 11, 194. [Google Scholar] [CrossRef]
- El-Sharkawy, H.H.; Abbas, M.S.; Soliman, A.S.; Ibrahim, S.A.; El-Nady, I.A. Synergistic effect of growth-promoting microorganisms on bio-control of Fusarium oxysporum f. sp. pisi, growth, yield, physiological and anatomical characteristics of pea plants. Pestic. Biochem. Physiol. 2021, 178, 104939. [Google Scholar] [CrossRef]
- Miljaković, D.; Marinković, J.; Tamindžić, G.; Đorđević, V.; Tintor, B.; Milošević, D.; Ignjatov, M.; Nikolić, Z. Bio-priming of soybean with Bradyrhizobium japonicum and Bacillus megaterium: Strategy to improve seed germination and the initial seedling growth. Plants 2022, 11, 1927. [Google Scholar] [CrossRef]
- Cavalcanti, M.I.P.; de Carvalho Nascimento, R.; Rodrigues, D.R.; Escobar, I.E.C.; Fraiz, A.C.R.; de Souza, A.P.; de Freitas, A.D.S.; Nobrega, R.S.A.; Fernandes-Junior, P.I. Maize growth and yield promoting endophytes isolated into a legume root nodule by a cross-over approach. Rhizosphere 2020, 15, 100211. [Google Scholar] [CrossRef]
- Piromyou, P.; Greetatorn, T.; Teamtisong, K.; Okubo, T.; Shinoda, R.; Nuntakij, A.; Tittabutr, P.; Boonkerd, N.; Minamisawa, K.; Teaumroong, N. Preferential association of endophytic bradyrhizobia with different rice cultivars and its implications for rice endophyte evolution. Appl. Environ. Microbiol. 2015, 81, 3049–3061. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Chater, C.C.; Yu, F.; Perez-Moreno, J. Tuber pseudohimalayense ascomata-compartments strongly select their associated bacterial microbiome from nearby pine forest soils independently of their maturation stage. Pedobiologia 2021, 87, 150743. [Google Scholar] [CrossRef]
- Shang, J.Y.; Zhang, P.; Jia, Y.W.; Lu, Y.N.; Wu, Y.; Ji, S.; Chen, L.; Wang, E.T.; Chen, W.X.; Sui, X.H. Coordinated regulation of symbiotic adaptation by NodD proteins and NolA in the type I peanut bradyrhizobial strain Bradyrhizobium zhanjiangense CCBAU51778. Microbiol. Res. 2022, 265, 127188. [Google Scholar] [CrossRef]
- Orozco-Mosqueda, M.d.C.; Flores, A.; Rojas-Sánchez, B.; Urtis-Flores, C.A.; Morales-Cedeño, L.R.; Valencia-Marin, M.F.; Chávez-Avila, S.; Rojas-Solis, D.; Santoyo, G. Plant growth-promoting bacteria as bioinoculants: Attributes and challenges for sustainable crop improvement. Agronomy 2021, 11, 1167. [Google Scholar] [CrossRef]
- Saeed, Q.; Xiukang, W.; Haider, F.U.; Kučerik, J.; Mumtaz, M.Z.; Holatko, J.; Naseem, M.; Kintl, A.; Ejaz, M.; Naveed, M. Rhizosphere bacteria in plant growth promotion, biocontrol, and bioremediation of contaminated sites: A comprehensive review of effects and mechanisms. Int. J. Mol. Sci. 2021, 22, 10529. [Google Scholar] [CrossRef]
- Cruz-Hernández, M.A.; Mendoza-Herrera, A.; Bocanegra-García, V.; Rivera, G. Azospirillum spp. from Plant Growth-Promoting Bacteria to Their Use in Bioremediation. Microorganisms 2022, 10, 1057. [Google Scholar] [CrossRef]
- Degani, O.; Rabinovitz, O.; Becher, P.; Gordani, A.; Chen, A. Trichoderma longibrachiatum and Trichoderma asperellum confer growth promotion and protection against late wilt disease in the field. J. Fungi 2021, 7, 444. [Google Scholar] [CrossRef]
- Alfiky, A.; Weisskopf, L. Deciphering Trichoderma–plant–pathogen interactions for better development of biocontrol applications. J. Fungi 2021, 7, 61. [Google Scholar] [CrossRef]
- Saravanakumar, K.; Li, Y.; Yu, C.; Wang, Q.-q.; Wang, M.; Sun, J.; Gao, J.-x.; Chen, J. Effect of Trichoderma harzianum on maize rhizosphere microbiome and biocontrol of Fusarium Stalk rot. Sci. Rep. 2017, 7, 1771. [Google Scholar] [CrossRef]
- Steiner, F.; de Queiroz, L.F.M.; Zuffo, A.M.; da Silva, K.C.; Lima, I.M.d.O. Peanut response to co-inoculation of Bradyrhizobium spp. and Azospirillum brasilense and molybdenum application in sandy soil of the Brazilian Cerrado. Agron. J. 2021, 113, 623–632. [Google Scholar] [CrossRef]
- Carlos, L.; Venturin, N.; Macedo, R.L.G.; Higashikawa, E.M.; Garcia, M.B.; Farias, E.d.S. Crescimento e nutrição mineral de mudas de pequi sob efeito da omissão de nutrientes. Ciência Florest. 2014, 24, 13–21. [Google Scholar] [CrossRef] [Green Version]
- Asamatdinov, M.; Zhukov, A.; Medvedev, A.; Mukhametzyanov, V. Fire protection systems using clay-gypsum plaster in agriculture. E3S Web Conf. EDP Sci. 2020, 175, 11008. (In Russian) [Google Scholar] [CrossRef]
- Badawi, F.S.F.; Biomy, A.; Desoky, A. Peanut plant growth and yield as influenced by co-inoculation with Bradyrhizobium and some rhizo-microorganisms under sandy loam soil conditions. Ann. Agric. Sci. 2011, 56, 17–25. [Google Scholar] [CrossRef] [Green Version]
- Araújo, F.F.d.; Hungria, M. Soybean nodulation and yield when co-inoculated with Bacillus subtilis and Bradyrhizobium japonicum/Bradyrhizobium elkanii. Pesqui. Agropecuária Bras. 1999, 34, 1633–1643. [Google Scholar] [CrossRef]
- Carvalho, D.D.C.; Mello, S.C.M.d.; Lobo Júnior, M.; Geraldine, A.M. Biocontrol of seed pathogens and growth promotion of common bean seedlings by Trichoderma harzianum. Pesqui. Agropecuária Bras. 2011, 46, 822–828. [Google Scholar] [CrossRef] [Green Version]
- Malavolta, E.; Damião Filho, C.; Volpe, C.; Machado Jr, C.; Velho, L.; Rosa, P.; Laurentiz, S.d. Deficiências e excessos minerais no feijoeiro (Phaseolus vulgaris L., cv. Carioca). An. Esc. Super. Agric. Luiz Queiroz 1980, 37, 701–718. [Google Scholar] [CrossRef]
- Mohamed, T.A.; Wu, J.; Zhao, Y.; Elgizawy, N.; El Kholy, M.; Yang, H.; Zheng, G.; Mu, D.; Wei, Z. Insights into enzyme activity and phosphorus conversion during kitchen waste composting utilizing phosphorus-solubilizing bacterial inoculation. Bioresour. Technol. 2022, 362, 127823. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Chen, G.; Qin, Y.; Li, J.; Hu, L.; Gu, W.; Zhu, C. Proton-regulated catalytic activity of nanozymes for dual-modal bioassay of urease activity. Anal. Chem. 2021, 93, 9897–9903. [Google Scholar] [CrossRef]
- Barbosa, J.; Maldonado Júnior, W. AgroEstat: Sistema para Análises Estatísticas de Ensaios Agronômicos; Fac. Ciências Agrárias Veterinárias Unesp: Jaboticabal, Brazil, 2010. [Google Scholar]
- Johnson III, W.C. A review of weed management challenges in organic peanut production. Peanut Sci. 2019, 46, 56–66. [Google Scholar] [CrossRef]
- Santos, R.M.d.; Desoignies, N.; Rigobelo, E.C. The bacterial world inside the plant. Front. Sustain. Food Syst. 2022, 6, 319. [Google Scholar] [CrossRef]
- Aioub, A.A.; Elesawy, A.E.; Ammar, E.E. Plant growth promoting rhizobacteria (PGPR) and their role in plant-parasitic nematodes control: A fresh look at an old issue. J. Plant Dis. Prot. 2022, 129, 1305–1321. [Google Scholar] [CrossRef]
- Altaf, M. Functional diversity of nitrogen-fixing plant growth-promoting Rhizobacteria: The story so far. In Soil Nitrogen Ecology; Springer: Berlin/Heidelberg, Germany, 2021; Volume 1, pp. 327–348. [Google Scholar]
- Duff, A.M.; Forrestal, P.; Ikoyi, I.; Brennan, F. Assessing the long-term impact of urease and nitrification inhibitor use on microbial community composition, diversity and function in grassland soil. Soil Biol. Biochem. 2022, 170, 108709. [Google Scholar] [CrossRef]
- Huang, Y.; Dai, Z.; Lin, J.; Li, D.; Ye, H.; Dahlgren, R.A.; Xu, J. Labile carbon facilitated phosphorus solubilization as regulated by bacterial and fungal communities in Zea mays. Soil Biol. Biochem. 2021, 163, 108465. [Google Scholar] [CrossRef]
- Escobar Diaz, P.A.; Santos, R.M.d.; Baron Cozentino, N.C.; Oniel, J.A.; Rigobelo, E.C. Effect of Aspergillus and Bacillus concentration on cotton growth promotion. Front. Microbiol. 2021, 12, 2734. [Google Scholar] [CrossRef]
- Yang, J.K.; Zhou, J.C. Diversity, phylogeny and host specificity of soybean and peanut bradyrhizobia. Biol. Fertil. Soils 2008, 44, 843–851. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, H.-W.; Wang, X.-X.; Xie, X.-G.; Siddikee, M.A.; Xu, R.-S.; Dai, C.-C. Enhanced nodulation of peanut when co-inoculated with fungal endophyte Phomopsis liquidambari and bradyrhizobium. Plant Physiol. Biochem. 2016, 98, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Mathesius, U. Are legumes different? Origins and consequences of evolving nitrogen fixing symbioses. J. Plant Physiol. 2022, 276, 153765. [Google Scholar] [CrossRef]
- Lobo, L.L.B.; da Silva, M.S.R.d.A.; Carvalho, R.F.; Rigobelo, E.C. The negative effect of coinoculation of plant growth-promoting bacteria is not related to indole-3-acetic acid synthesis. J. Plant Growth Regul. 2022, 5, 1–10. [Google Scholar] [CrossRef]
- Lobo, L.L.B.; de Andrade da Silva, M.S.R.; Castellane, T.C.L.; Carvalho, R.F.; Rigobelo, E.C. Effect of Indole-3-Acetic Acid on Tomato Plant Growth. Microorganisms 2022, 10, 2212. [Google Scholar] [CrossRef] [PubMed]
- Lopes, M.J.d.S.; Dias-Filho, M.B.; Gurgel, E.S.C. Successful plant growth-promoting microbes: Inoculation methods and abiotic factors. Front. Sustain. Food Syst. 2021, 5, 606454. [Google Scholar] [CrossRef]
- Mitter, E.K.; Kataoka, R.; de Freitas, J.R.; Germida, J.J. Potential use of endophytic root bacteria and host plants to degrade hydrocarbons. Int. J. Phytoremediation 2019, 21, 928–938. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Inoculants |
|---|---|
| 1 | Control (Without inoculant) |
| 2 | Azospirillum brasilense |
| 3 | Bacillus subtilis |
| 4 | Bradyrhizobium japonicum |
| 5 | Trichoderma harzianum |
| 6 | A. brasilense + B. subtilis |
| 7 | A. brasilense + B. japonicum |
| 8 | A. brasilense + T. harzianum |
| 9 | B. subtilis + B. japonicum |
| 10 | B. subtilis + T. harzianum |
| 11 | B. japonicum + T. harzianum |
| 12 | Consortium (A. brasilense + B. subtilis + B. japonicum + T. harzianum) |
| Treatment | Inoculant |
|---|---|
| 1 | Control-No inoculant |
| 2 | Azospirillum brasilense |
| 3 | Bradyrhizobium japonicum |
| 4 | A. brasilense + B. japonicum |
| 5 | A. brasilense + Trichoderma harzianum |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Frezarin, E.T.; Santos, C.H.B.; Sales, L.R.; dos Santos, R.M.; de Carvalho, L.A.L.; Rigobelo, E.C. Promotion of Peanut (Arachis hypogaea L.) Growth by Plant Growth-Promoting Microorganisms. Microbiol. Res. 2023, 14, 316-332. https://doi.org/10.3390/microbiolres14010025
Frezarin ET, Santos CHB, Sales LR, dos Santos RM, de Carvalho LAL, Rigobelo EC. Promotion of Peanut (Arachis hypogaea L.) Growth by Plant Growth-Promoting Microorganisms. Microbiology Research. 2023; 14(1):316-332. https://doi.org/10.3390/microbiolres14010025
Chicago/Turabian StyleFrezarin, Edvan Teciano, Carlos Henrique Barbosa Santos, Luziane Ramos Sales, Roberta Mendes dos Santos, Lucas Amoroso Lopes de Carvalho, and Everlon Cid Rigobelo. 2023. "Promotion of Peanut (Arachis hypogaea L.) Growth by Plant Growth-Promoting Microorganisms" Microbiology Research 14, no. 1: 316-332. https://doi.org/10.3390/microbiolres14010025
APA StyleFrezarin, E. T., Santos, C. H. B., Sales, L. R., dos Santos, R. M., de Carvalho, L. A. L., & Rigobelo, E. C. (2023). Promotion of Peanut (Arachis hypogaea L.) Growth by Plant Growth-Promoting Microorganisms. Microbiology Research, 14(1), 316-332. https://doi.org/10.3390/microbiolres14010025