
In Vitro Screening of Antiviral Activity of Lactic Acid Bacteria Isolated from Traditional Fermented Foods

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Studied LAB Strains and CFS Preparation
2.2. Cytotoxicity Test
2.3. Antiviral Activity of the Bacterial Supernatant
2.3.1. MTT-Based Colorimetric Assay for Detection of HHV Replication Inhibition
2.3.2. Virucidal Activity
2.4. Data Analyses
3. Results
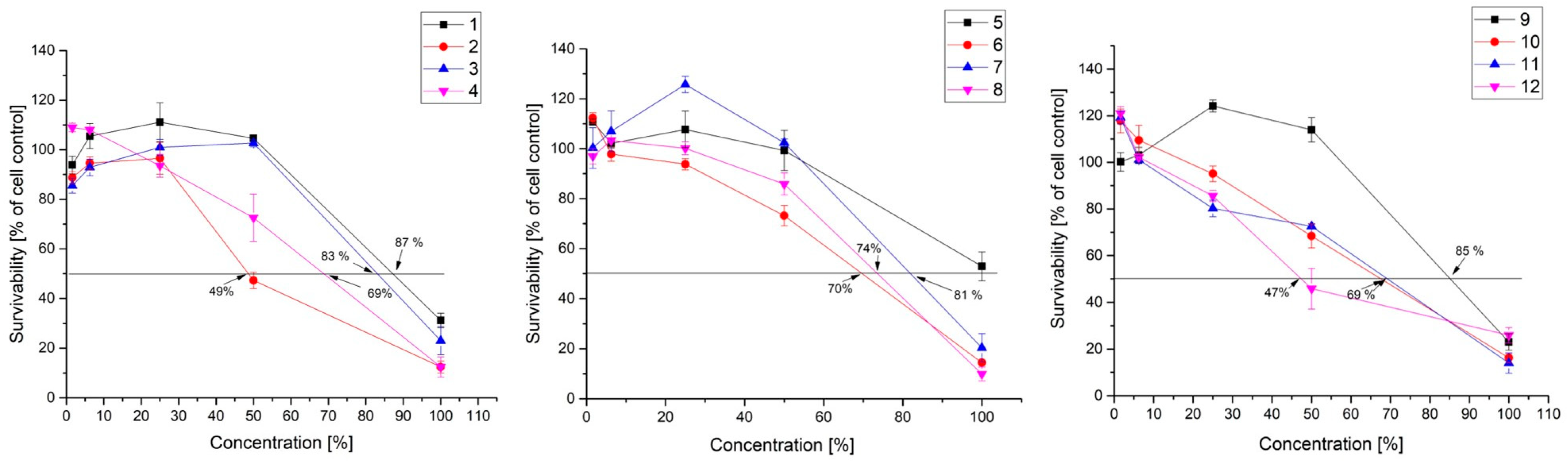
3.1. Cytotoxicity
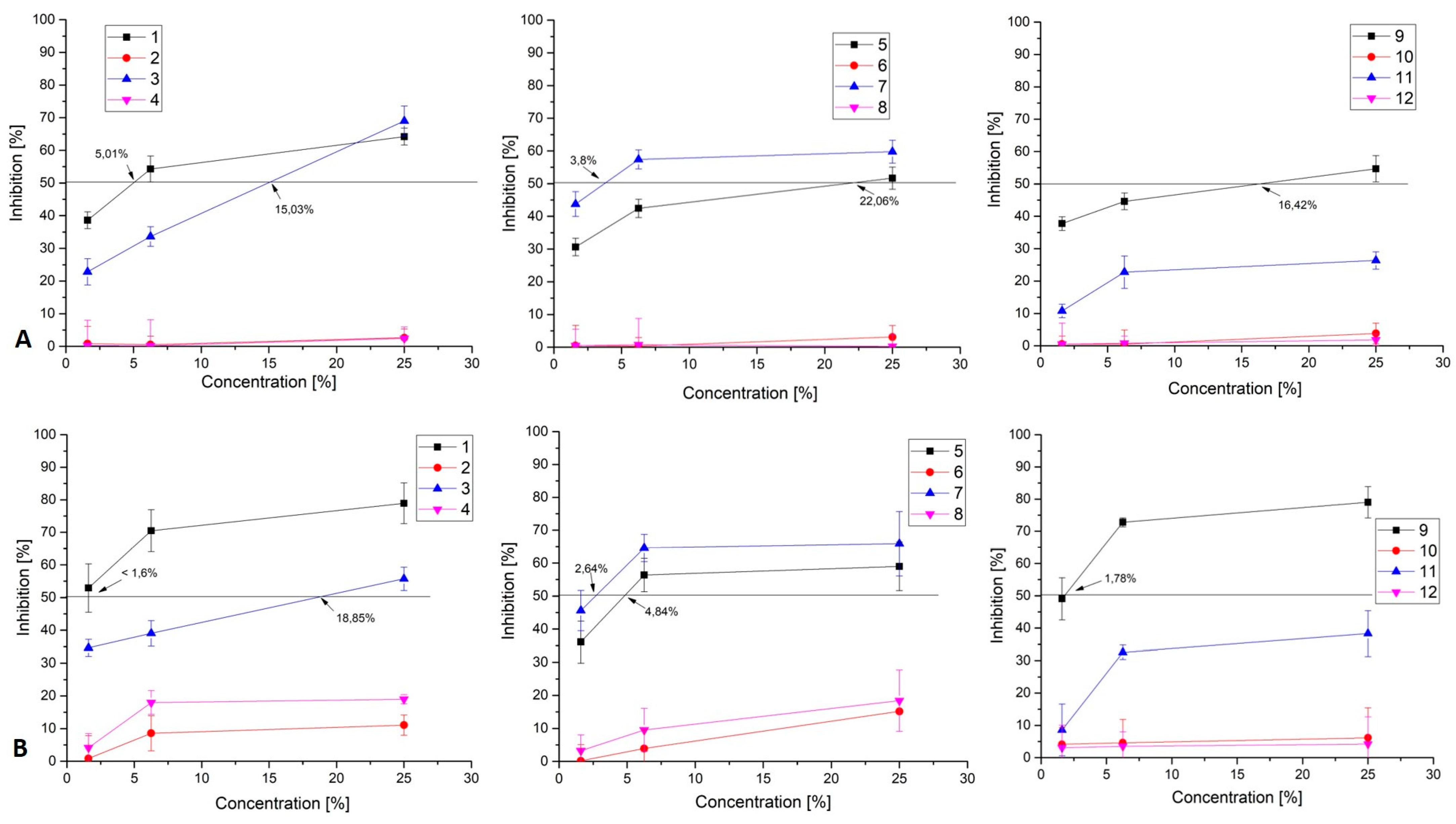
3.2. Evaluation of HHV Replication Inhibition from the Bacterial CFSs
3.3. Selective Index (SI)
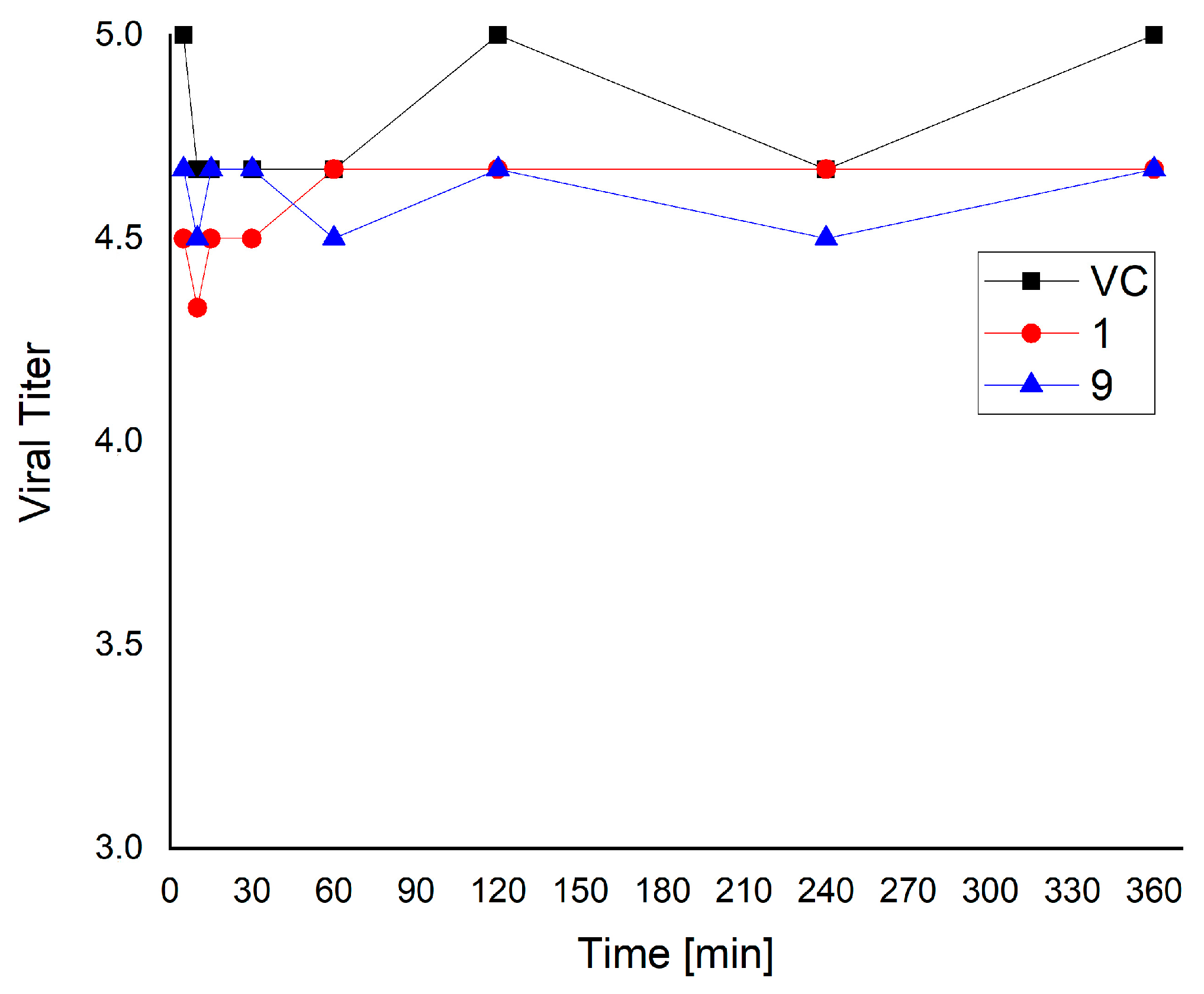
3.4. Virucidal Activity
4. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhou, T.; Huo, R.; Lai-Yu, K.; Li, C.; Ma, Y.; Mi, Z.; Chen, Y. Effects of applying Lactobacillus helveticus H9 as adjunct starter culture in yogurt fermentation and storage. J. Dairy Sci. 2019, 102, 223–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassan, A.N.; Frank, J.F. Starter Cultures and Their Use. In Applied Dairy Microbiology, 2nd ed.; revised and expanded; Marth, E.H., Steele, J.L., Eds.; CRC Press: Boca Raton, FL, USA, 2001; Volume 6, p. 736. [Google Scholar]
- Russo, P.; Fares, C.; Longo, A.; Spano, G.; Capozzi, V. Lactobacillus plantarum with Broad Antifungal Activity as a Protective Starter Culture for Bread Production. Foods 2017, 6, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bintsis, T. Lactic acid bacteria: Their applications in foods. J. Bacteriol. Mycol. 2018, 6, 89–94. [Google Scholar]
- Bai, M.; Huang, T.; Guo, S.; Wang, Y.; Wang, J.; Kwok, L.; Dan, T.; Zhang, H.; Bilige, M. Probiotic Lactobacillus casei Zhang improved the properties of stirred yogurt. Food Biosci. 2020, 37, 100718. [Google Scholar] [CrossRef]
- Jing, W.; Haifeng, J.; Sixin, W.; Hui, L.; Wei, Z.; Dongyan, Z.; Yamin, W. Probiotic Lactobacillus plantarum Promotes Intestinal Barrier Function by Strengthening the Epithelium and Modulating Gut Microbiota. Front. Microbiol. 2018, 9, 1953. [Google Scholar]
- Smit, G.; Smit, B.A.; Engels, W.J.M. Flavour formation by lactic acid bacteria and biochemical flavour profiling of cheese products. FEMS Microbiol. Rev. 2005, 29, 591–610. [Google Scholar] [CrossRef]
- Ramzi, A.; Alsaheb, A.; Aladdin, A.; Othman, N.Z.; Malek, R.M.; Leng, O.M.; Aziz, R.; El Enshasy, H.A. Lactic acid applications in pharmaceutical and cosmeceutical industries. J. Chem. Pharm. Res. 2015, 7, 729–735. [Google Scholar]
- Ren, C.; Faas, M.M.; de Vos, P. Disease managing capacities and mechanisms of host effects of lactic acid bacteria. Critical Rev. Food Sci. Nutr. 2021, 61, 1365–1393. [Google Scholar] [CrossRef]
- Tiwari, S.K.; Dicks, L.M.T.; Popov, I.V.; Karaseva, A.; Ermakov, A.M.; Suvorov, A.; Tagg, J.R.; Weeks, R.; Chikindas, M.L. Probiotics at War Against Viruses: What Is Missing from the Picture? Front. Microbiol. 2020, 11, 1877. [Google Scholar] [CrossRef]
- Gaspar, C.; Donders, G.G.; Palmeira-de-Oliveira, R.; Queiroz, J.A.; Tomaz, C.; Martinez-de-Oliveira, J.; Palmeira-de-Oliveira, A. Bacteriocin production of the probiotic Lactobacillus acidophilus KS400. AMB Expr. 2018, 8, 153. [Google Scholar] [CrossRef]
- Zhu, Y.; Wang, X.; Pan, W.; Shen, X.; He, Y.; Yin, H.; Zhou, K.; Zou, L.; Chen, S.; Liu, S. Exopolysaccharides produced by yogurt-texture improving Lactobacillus plantarum RS20D and the immunoregulatory activity. Int. J. Biol. Macromol. 2019, 121, 342–349. [Google Scholar] [CrossRef]
- Nes, I.F.; Kjos, M.; Diep, D.B. Antimicrobial Compounds of Lactic Acid Bacteria. In Lactic Acid Bacteria Microbiological and Functional Aspects, 4th ed.; Lahtinen, S., Ouwehand, A.C., Salminen, S., von Wright, A., Eds.; CRC Press: Boca Raton, FL, USA, 2012; Volume 5, pp. 285–329. [Google Scholar]
- Conti, C.; Malacrino, C.; Mastromarino, P. Inhibition of herpes simplex virus type 2 by vaginal lactobacilli. J. Physiol. Pharmacol. 2009, 6, 19–26. [Google Scholar]
- Akhtar, J.; Shukla, D. Viral entry mechanisms: Cellular and viral mediators of herpes simplex virus entry. FEBS J. 2009, 276, 7228–7236. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://www.who.int/news-room/fact-sheets/detail/herpes-simplex-virus (accessed on 10 March 2022).
- Morfin, F.; Thouvenot, D. Herpes simplex virus resistance to antiviral drugs. J. Clin. Virol. 2003, 26, 29–37. [Google Scholar] [CrossRef]
- Al Kassaa, I.; Hober, D.; Hamze, M.; Chihib, N.E.; Drider, D. Antiviral potential of lactic acid bacteria and their bacteriocins. Probiotics Antimicrob. Proteins 2014, 6, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Carver, D.H.; Naficy, K. Inhibition of arbor viruses (group A) by a protein-like constituent of a Corynebacterium. Proc. Soc. Exp. Biol. Med. 1962, 111, 356–360. [Google Scholar] [CrossRef]
- Cliver, D.O.; Herrmann, J.E. Proteolytic and microbial inactivation of enteroviruses. Water Res. 1972, 6, 797–805. [Google Scholar] [CrossRef]
- Deng, M.Y.; Cliver, D.O. Inactivation of poliovirus type 1 in mixed human and swine wastes and by bacteria from swine manure. Appl. Environ. Microbiol. 1992, 58, 2016–2021. [Google Scholar] [CrossRef] [Green Version]
- Deng, M.Y.; Cliver, D.O. Antiviral effects of bacteria isolated from manure. Microb. Ecol. 1995, 30, 43–54. [Google Scholar] [CrossRef]
- Munoz, J.A.; Chenoll, E.; Casinos, B.; Bataller, E.; Ramon, D.; Genoves, S.; Montava, R.; Ribes, J.M.; Buesa, J.; Fabrega, J.; et al. Novel probiotic Bifidobacterium longum subsp. infantis CECT 7210 strain active against rotavirus infections. Appl. Environ. Microbiol. 2011, 77, 8775–8783. [Google Scholar] [CrossRef] [Green Version]
- Muhialdin, B.J.; Zawawi, N.; Abdul Razis, A.F.; Bakar, J.; Zarei, M. Review Antiviral activity of fermented foods and their probiotics bacteria towards respiratory and alimentary tracts viruses. Food Control 2021, 127, 108140. [Google Scholar] [CrossRef]
- Hotta, S.; Kojima, M.; Fujisaki, M.; Uchida, S.; Kuroda, H.; Hamada, C. An antiviral substance extracted from Streptococcus faecalis. Nature 1977, 268, 733–734. [Google Scholar] [CrossRef] [PubMed]
- Botić, T.; Danø, T.; Weingartl, H.; Cencič, A. A novel eukaryotic cell culture model to study antiviral activity of potential probiotic bacteria. Int. J. Food Microbiol. 2007, 115, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.J.; Song, J.H.; Ahn, Y.J.; Baek, S.H.; Kwon, D.H. Antiviral activities of cell-free supernatants of yogurts metabolites against some RNA viruses. Eur. Food Res. Technol. 2009, 228, 945–950. [Google Scholar] [CrossRef]
- Shearer, A.E.; Hoover, D.G.; Kniel, K.E. Effect of bacterial cell-free supernatants on infectivity of norovirus surrogates. J. Food Prot. 2014, 77, 145–149. [Google Scholar] [PubMed]
- Sirichokchatchawan, W.; Temeeyasen, G.; Nilubol, D.; Prapasarakul, N. Protective effects of cell-free supernatant and live lactic acid bacteria isolated from Thai pigs against a pandemic strain of porcine epidemic diarrhea virus. Probiotics Antimicrob. Proteins 2018, 10, 383–390. [Google Scholar] [CrossRef]
- Hoxha, R.; Evstatieva, Y.; Nikolova, D. New lactic acid bacterial strains from traditional fermented foods—Bioprotective and probiotic potential. J. Chem. Technol. Metall. 2023, 58, 252–269. [Google Scholar]
- Mastromarino, P.; Cacciotti, F.; Masci, A.; Mosca, L. Antiviral activity of Lactobacillus brevis towards herpes simplex virus type 2: Role of cell-wall associated components. Anaerobe 2011, 17, 334–336. [Google Scholar] [CrossRef]
- Li, W.; Zhou, J.; Xu, Y. Study of the in vitro cytotoxicity testing of medical devices. Biomed. Rep. 2015, 3, 617–620. [Google Scholar] [CrossRef] [Green Version]
- Mani-López, E.; Arrioja-Bretón, D.; López-Malo, A. The impacts of antimicrobial and antifungal activity of cell-free supernatants from lactic acid bacteria in vitro and foods. Compr. Rev. Food Sci Food Saf. 2022, 21, 604–641. [Google Scholar] [CrossRef]
- Vilhelmova-Ilieva, N.; Atanasov, G.; Simeonova, L.; Dobreva, L.; Mancheva, K.; Trepechova, M.; Danova, S. Anti-herpes virus activity of lactobacillus’ postbiotics. BioMedicine 2022, 12, 21. [Google Scholar] [CrossRef]
- Möller, N.P.; Scholz-Ahrens, K.E.; Roos, N.; Schrezenmeir, J. Bioactive peptides and proteins from foods: Indication for health effects. Eur. J. Nutr. 2008, 47, 171–182. [Google Scholar] [CrossRef]
- Khani, S.; Motamedifar, M.; Golmoghaddam, H.; Hosseini, H.M.; Hashemizadeh, Z. In vitro study of the effect of a probiotic bacterium Lactobacillus rhamnosus against herpes simplex virus type 1. Braz. J. Infect. Dis. 2012, 16, 129–135. [Google Scholar] [PubMed] [Green Version]
- Cirrincione, S.; Luganini, A.; Lamberti, C.; Manfredi, M.; Cavallarin, L.; Giuffrida, M.G.; Pessione, E. Donkey Milk Fermentation by Lactococcus lactis subsp. cremoris and Lactobacillus rhamnosus Affects the Antiviral and Antibacterial Milk Properties. Molecules 2021, 26, 5100. [Google Scholar] [CrossRef] [PubMed]
- Mousavi, E.; Makvandi, M.; Teimoori, A.; Ataei, A.; Ghafari, S.; Samarbaf-Zadeh, A. Antiviral effects of Lactobacillus crispatus against HSV-2 in mammalian cell lines. J. Chin. Med. Assoc. 2018, 81, 262–267. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Lee, G.; Thanh, H.D.; Kim, J.; Konkit, M.; Yoon, S.; Park, M.; Yang, S.; Park, E.; Kim, W. Exopolysaccharide from Lactobacillus plantarum LRCC5310 offers protection against rotavirus-induced diarrhea and regulates inflammatory response. J. Dairy Sci. 2017, 101, 5702–5712. [Google Scholar] [CrossRef]
- Biliavska, L.; Pankivska, Y.; Povnitsa, O.; Zagorodnya, S. Antiviral Activity of Exopolysaccharides Produced by Lactic Acid Bacteria of the genera Pediococcus, Leuconostoc and Lactobacillus against Human Adenovirus Type 5. Medicina 2019, 55, 519. [Google Scholar] [CrossRef] [Green Version]
- Kwak, M.; Liu, R.; Kwon, J.; Kim, M.; Kim, A.H.; Kang, S. Cyclic Dipeptides from Lactic Acid Bacteria Inhibit Proliferation of the Influenza A Virus. J. Microbiol. 2013, 51, 836–843. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Li, S.; Lu, Y.; Jansen, C.h.A.; Savelkoul, H.F.J.; Liu, G. Oral administration of Lactic acid bacteria inhibits PEDV infection in young piglets. Virology 2023, 579, 1–8. [Google Scholar] [CrossRef]
- Lopez-Santamarina, A.; Lamas, A.; del Carmen Mondragón, A.; Cardelle-Cobas, A.; Regal, P.; Rodriguez-Avila, J.A.; Miranda, J.M.; Franco, C.M.; Cepeda, A. Probiotic Effects against Virus Infections: New Weapons for an Old War. Foods 2021, 10, 130. [Google Scholar] [CrossRef]
- Rather, I.A.; Kamli, M.R.; Sabir, J.S.M.; Paray, B.A. Potential Antiviral Activity of Lactiplantibacillus plantarum KAU007 against Influenza Virus H1N1. Vaccines 2022, 10, 456. [Google Scholar] [CrossRef]
- Rather, I.A.; Choi, S.-B.; Kamli, M.R.; Hakeem, K.R.; Sabir, J.S.M.; Park, Y.-H.; Hor, Y.-Y. Potential Adjuvant Therapeutic Effect of Lactobacillus plantarum Probio-88 Postbiotics against SARS-COV-2. Vaccines 2021, 9, 1067. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, E.; Sawabuchi, T.; Kimoto, T.; Sakai, S.; Kido, H. Lactobacillus delbrueckii ssp. bulgaricus OLL1073R-1 feeding enhances humoral immune responses, which are suppressed by the antiviral neuraminidase inhibitor oseltamivir in influenza A virus–infected mice. J. Dairy Sci. 2019, 102, 9559–9569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serkedjieva, J.; Danova, S.; Ivanova, I. Anti-influenza virus activity of a bacteriocin produced by Lactobacillus delbrueckii. Appl. Biochem. Biotechnol. 2000, 88, 285–298. [Google Scholar] [CrossRef]
- Wahab, S.; Almaghaslah, D.; Mahmood, S.E.; Ahmad, M.F.; Alsayegh, A.A.; Abu Haddash, Y.M.; Rahman, M.A.; Ahamd, I.; Ahmad, W.; Khalid, M.; et al. Pharmacological Efficacy of Probiotics in Respiratory Viral Infections: A Comprehensive Review. J. Pers. Med. 2022, 12, 1292. [Google Scholar] [CrossRef] [PubMed]
- Lehtoranta, L.; Pitkäranta, A.; Korpela, R. Probiotics in respiratory virus infections. Eur. J. Clin. Microbiol. Infect. Dis. 2014, 33, 1289–1302. [Google Scholar] [CrossRef] [PubMed]
- Martín, V.; Maldonado, A.; Fernández, L.; Rodríguez, J.M.; Connor, R.I. Inhibition of human immunodeficiency virus type 1 by lactic acid bacteria from human breastmilk. Breastfeed Med. 2010, 5, 153–158. [Google Scholar] [CrossRef]
- Chourasia, R.; Padhi, S.; Chiring Phukon, L.; Abedin, M.M.; Singh, S.P.; Rai, A.K. A Potential Peptide from Soy Cheese Produced Using Lactobacillus delbrueckii WS4 for Effective Inhibition of SARS-CoV-2 Main Protease and S1 Glycoprotein. Front. Mol. Biosci. 2020, 7, 601753. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Strains | Origin | CFS Sample Numbering |
|---|---|---|
| L. plantarum KC 5-12 | White cow cheese | 1 |
| P. pentosaceus KC 5-13 | White cow cheese | 2 |
| L. plantarum KC 5-14 | White cow cheese | 3 |
| L. plantarum KZC 8-21-1 | White goat cheese | 4 |
| L. plantarum KZC 8-23-5 | White goat cheese | 5 |
| L. sakei C 10-31-3 | Sausage | 6 |
| L. plantarum KO 4-4 | Yellow ewe cheese | 7 |
| L. coryniformis KO 3-7-5 | Yellow ewe cheese | 8 |
| L. delbrueckii subsp. bulgaricus KZM 2-11-3 | Yogurt | 9 |
| L. delbrueckii subsp. bulgaricus KZM 2-11-1 | Yogurt | 10 |
| L. delbrueckii subsp. bulgaricus KZM 2-11-3 (SM) | Yogurt | 11 |
| L. delbrueckii subsp. bulgaricus KZM 2-11-1 (SM) | Yogurt | 12 |
| Sample No. | CFSs from LAB Strains | HHV-1 | HHV-2 | |||
|---|---|---|---|---|---|---|
| CC50 | IC50 | SI | IC50 | SI | ||
| 1 | KC 5-12 | 87 | 5.01 | 17.37 | <1.6 | >54 |
| 3 | KC 5-14 | 83 | 15.03 | 5.52 | 18.85 | 4.40 |
| 5 | KZC 8-23-5 | >100 | 22.06 | 4.53 | 4.84 | 20.66 |
| 7 | KO 4-4 | 81 | 3.8 | 21.32 | 2.64 | 30.68 |
| 9 | KZM 2-11-3 | 85 | 16.42 | 5.18 | 1.78 | 47.75 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hoxha, R.; Todorov, D.; Hinkov, A.; Shishkova, K.; Evstatieva, Y.; Nikolova, D. In Vitro Screening of Antiviral Activity of Lactic Acid Bacteria Isolated from Traditional Fermented Foods. Microbiol. Res. 2023, 14, 333-342. https://doi.org/10.3390/microbiolres14010026
Hoxha R, Todorov D, Hinkov A, Shishkova K, Evstatieva Y, Nikolova D. In Vitro Screening of Antiviral Activity of Lactic Acid Bacteria Isolated from Traditional Fermented Foods. Microbiology Research. 2023; 14(1):333-342. https://doi.org/10.3390/microbiolres14010026
Chicago/Turabian StyleHoxha, Ramize, Daniel Todorov, Anton Hinkov, Kalina Shishkova, Yana Evstatieva, and Dilyana Nikolova. 2023. "In Vitro Screening of Antiviral Activity of Lactic Acid Bacteria Isolated from Traditional Fermented Foods" Microbiology Research 14, no. 1: 333-342. https://doi.org/10.3390/microbiolres14010026
APA StyleHoxha, R., Todorov, D., Hinkov, A., Shishkova, K., Evstatieva, Y., & Nikolova, D. (2023). In Vitro Screening of Antiviral Activity of Lactic Acid Bacteria Isolated from Traditional Fermented Foods. Microbiology Research, 14(1), 333-342. https://doi.org/10.3390/microbiolres14010026