
The Structural–Rhythmological Organization of Coelogyne (Orchidaceae Juss.) Inflorescences



{kind=link}
{kind=link}
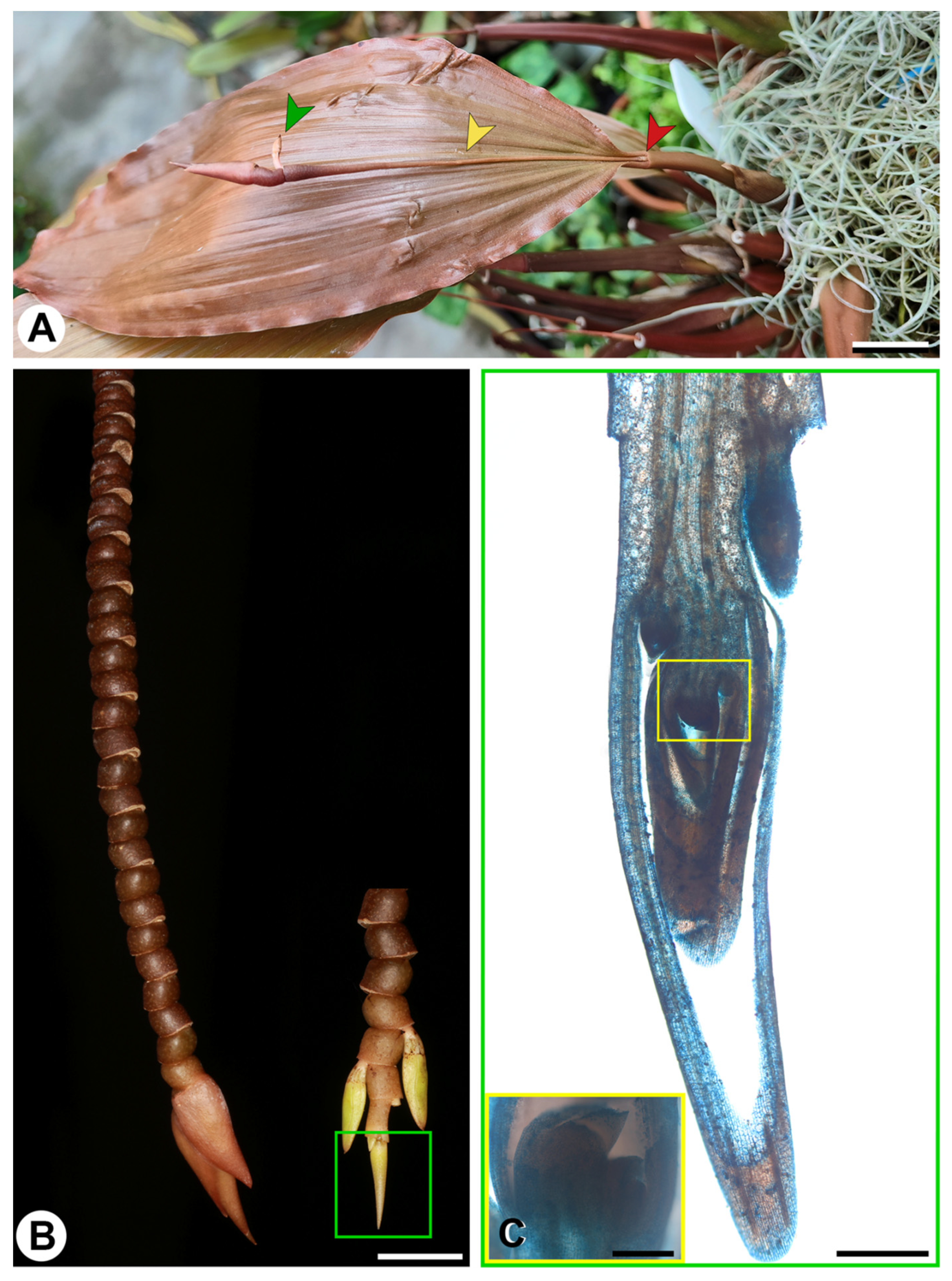{kind=link}
{kind=link}
{kind=link}
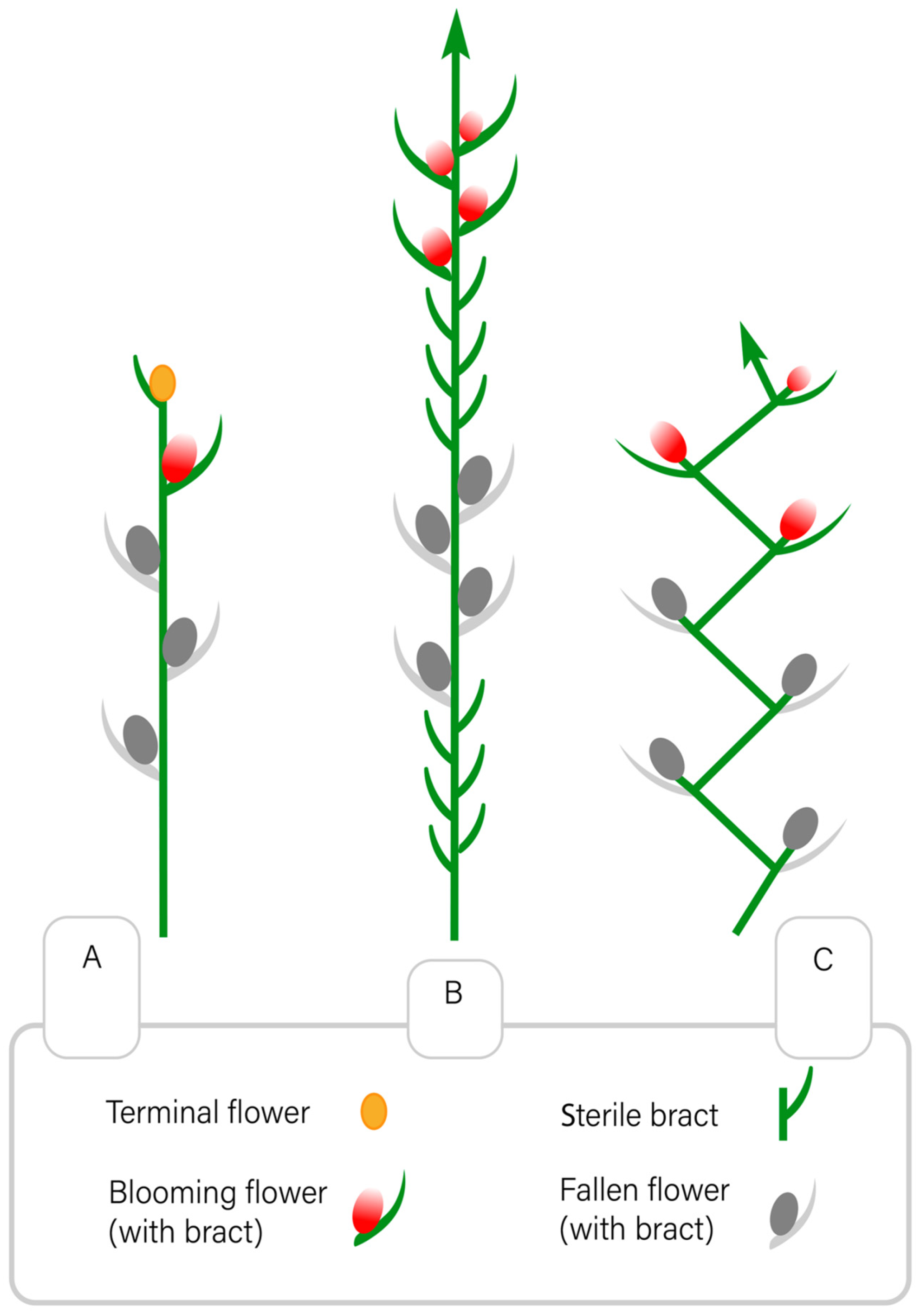{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. Structural Organization and Rhythmological Features of Coelogyne ovalis Inflorescence
3.2. Structural Organization and Rhythmological Features of Coelogyne prolifera Inflorescence
3.3. Structural Organization and Rhythmological Features of Coelogyne monilirachis Inflorescence
4. Discussion
4.1. Structural–Rhythmological Patterns for Coelogyne Inflorescences
4.2. One-Season Inflorescences of C. ovalis
4.3. Intercalary Inflorescences of C. prolifera
4.4. All-Season Inflorescences of C. monilirachis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Troll, W. Die Infloreszenzen. In Typologie und Stellung im Aufbau des Vegetationskörpers, 1st ed.; Gustav Fischer Verlag: Jena, Germany, 1964; p. 615. [Google Scholar]
- Kuznetsova, T.V.; Pryakhina, N.I.; Yakovlev, G.P. Inflorescence. In Morphological Classification; Chemical and Pharmaceutical Institute Press: St. Petersburg, Russia, 1992; p. 127. (In Russian) [Google Scholar]
- Tikhonova, M.N. The structure and formation of shoots in some species of tropical orchids of the genus Coelogyne Lindl. Bot. J. 1970, 55, 422–430. [Google Scholar]
- Arditti, J. Fundamentals of Orchid Biology; John Wiley & Sons: New York, NY, USA, 1992; pp. 356–456. ISBN 978-047-154-906-2. [Google Scholar]
- Bell, A.D. Plant form. In An Illustrated Guide to Flowering plant Morphology; Timber Press: London, UK, 2008; pp. 213–432. [Google Scholar]
- Troll, W. Die Infloreszenzen, 2nd ed.; Gustav Fischer Verlag: Jena, Germany, 1969; p. 630. [Google Scholar]
- Weberling, F. Morphology of Flowers and Inflorescences; Cambridge University Press: Cambridge, UK, 1989; pp. 156–405. ISBN 978-052-143-832-2. [Google Scholar]
- Prenner, G.; Vergara-Silva, F.; Rudall, P.J. The key role of morphology in modelling inflorescence architecture. Trends Plant Sci. 2009, 14, 302–309. [Google Scholar] [CrossRef] [PubMed]
- Kuznetsova, T.V. Morphology of Inflorescences; Results of Science and Technology; VINITI. Ser. Botany: Moscow, Russia, 1991; p. 127. (In Russian) [Google Scholar]
- Pridgeon, A.M.; Cribb, P.J.; Chase, M.W.; Rasmussen, F.N. (Eds.) Genera Orchidacearum, 4, Epidendroideae (Part One); Oxford University Press: Oxford, UK, 2005; ISBN 978-019-850-713-0. [Google Scholar]
- Van der Cingel, N.A. An Atlas of Orchid Pollination. In America, Africa, Asia and Australia; A.A. Balkema: Rotterdam, The Netherlands, 2001; p. 296. ISBN 905-410-486-4. [Google Scholar]
- Shi-Shun, Z.; Yunhong, T.; Xiao-Hua, J.; Win Maung, K.; Myint, Z.; Ren, L.; Rui-Chang, Q. Coelogyne victoria-reginae (Orchidaceae, Epidendroideae, Arethuseae), a new species from Chin State, Myanmar. PhytoKeys 2018, 98, 123–133. [Google Scholar] [CrossRef] [Green Version]
- Teoh, E.S. Medicinal Orchids of Thailand and Myanmar. Orchid. Aphrodisiac Med. Food 2019, 729, 245–254. [Google Scholar] [CrossRef]
- Nutan, S.; Suman, K. Molecular cloning and characterization of chalcone synthase gene from Coelogyne ovalis Lindl. and its stress-dependent expression. Gene 2020, 762, 145104. [Google Scholar] [CrossRef]
- Smirnova, E.S. Morphology of Orchid Shoot Systems; Golovkin, B.N., Ed.; Science: Moscow, Russia, 1990; pp. 50–209. (In Russian) [Google Scholar]
- Clayton, D. The Genus Coelogyne: A Synopsis; Natural History Publications:: Borneo; Kew, London, UK; The Royal Botanic Gardens Press: London, UK, 2002; ISBN 983-812-048-0. [Google Scholar]
- Koval, V.A.; Kolomeitseva, G.L. Types of inflorescence development in the genus Coelogyne Lindl. (Orchidaceae Juss.). In Proceedings of the International Scientific Conference “Plant Biomorphology: Traditions and Modernity”, Kirov, Russian, 19–21 October 2022; Vyatka State University Press: Kirov, Russian, 2022; pp. 239–244. (In Russian). [Google Scholar]
- Prozina, M.N. Botanical Microtechnology; Higher School: Moscow, Russia, 1960. (In Russian) [Google Scholar]
- Weberling, F. Typology of inflorescences. J. Linn. Soc. Bot. 1965, 59, 215–221. [Google Scholar] [CrossRef]
- Harder, L.D.; Prusinkiewicz, P. The interplay between inflorescence development and function as the crucible of architectural diversity. Ann. Bot. 2013, 112, 1477–1493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darwin, C.R. On the Various Contrivances by Which British and Foreign Orchids Are Fertilized by Insects; John Murray: London, UK, 1862. [Google Scholar]
- Ishii, H.S.; Hirabayashi, Y.; Kudo, G. Combined effects of inflorescence architecture, display size, plant density and empty flowers on bumble bee behaviour: Experimental study with artificial inflorescences. Oecologia 2008, 156, 341–350. [Google Scholar] [CrossRef] [PubMed]
- Troll, W. Organisation und Gestalt im Bereich der Blute, 1st ed.; Springer: Berlin, Germany, 1928; pp. 21–413. [Google Scholar]
- Prusinkiewicz, P.; Erasmus, Y.; Lane, B.; Harder, L.D.; Coen, E. Evolution and development of inflorescence architectures. Science 2007, 316, 1452–1456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Endress, P.K. Disentangling confusions in inflorescence morphology: Patterns and diversity of reproductive shoot ramification in angiosperms. J. Syst. Evol. 2010, 48, 225–239. [Google Scholar] [CrossRef]
- Linnaeus, K. Philosophy of Botany. AN SSSR (transl.from latin); M.: Science: Moscow, Russia, 1989. (In Russian) [Google Scholar]
- Parkin, J.M.A. The Evolution of the Inflorescence; J. Linn. Soc. Botany: London. UK, 1914; Volume 42, pp. 511–563. [Google Scholar]
- Maresquelle, H. Le theme évolutif des complexes d’inflorescences. Son aptitude a susciter des problèmes nouveaux. Bull. Soc. Bot. Fr. 1970, 117, 1–4. [Google Scholar] [CrossRef]
- Stebbins, G.L. Evolutionary trends in the inflorescence of angiosperms. Flora 1973, 162, 501–528. [Google Scholar] [CrossRef]
- Zhitkov, V.S.; Mamedova, E.T. Features of the structure of partial inflorescences in the family Gesneriaceae. Bull. Main Bot. Gard. 1982, 126, 78–83. (In Russian) [Google Scholar]
- Ricket, H. The European Garden Flora Flowering Plants. A Manual for the Identification; Cambridge University Press: Cambridge, UK, 2011. [Google Scholar]
- Nukhimovsky, E.D. Peculiarities of phenetic organization of biomorphs of seed plants. Mod. Biol. Successes 1986, 102, 289–306. (In Russian) [Google Scholar]
- Cremers, G. Architecture Vegetative et Structure Inflorescentielle de Quelques Melastomaceae Guyanaise; ORSTOM: Paris, France, 1986; ISBN 270-990-808-5. [Google Scholar]
- Weberling, F. Current Problems of Modern Inflorescence Morphology; Australasian Systematic Botany Society, Newsletter: Perth, Western Australia, 1982; pp. 3–21. [Google Scholar]
- Tucker, S.C.; Grimes, J. The Inflorescence: Introduction; The Botanical Review: Santa Barbara, CA, USA, 1999; Volume 65, pp. 303–316. [Google Scholar]
- Choob, V.V.; Mavrodiev, E.V. Morphological characters of the leaf series in the Commelinaceae family with special emphasis on the number of prophylls and their homology in monocots. Bot. J. 2001, 86, 1–11. (In Russian) [Google Scholar]
- Remizowa, M.V.; Sokoloff, D.D.; Rudall, P.J. Patterns of bract reduction inracemose inflorescences of early-divergent monocots. In Early Events in Monocot Evolution; Cambridge University Press: Cambridge, UK, 2013; pp. 185–207. [Google Scholar] [CrossRef]
- De Lange, P.J. A revision of the New Zeland Kunzea ericoides (Myrtaceae) complex. PhytoKeys 2014, 40, 1–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawler, L.J. Ethnobotany of the Orchidaceae. In Orchid Biology and Perspectives, III; Arditti, J., Ed.; Cornell University Press: Ithaca, NY, USA, 1984; pp. 27–149. [Google Scholar]
- Seidenfaden, G. Contributions to a Revision of the Orchid Flora of Cambodia, Laos and Vietnam; Kai Olsen: Copenhagen, Denmark, 1975. [Google Scholar]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kolomeitseva, G.; Koval, V.; Ryabchenko, A. The Structural–Rhythmological Organization of Coelogyne (Orchidaceae Juss.) Inflorescences. Int. J. Plant Biol. 2023, 14, 286-298. https://doi.org/10.3390/ijpb14010024
Kolomeitseva G, Koval V, Ryabchenko A. The Structural–Rhythmological Organization of Coelogyne (Orchidaceae Juss.) Inflorescences. International Journal of Plant Biology. 2023; 14(1):286-298. https://doi.org/10.3390/ijpb14010024
Chicago/Turabian StyleKolomeitseva, Galina, Vladimir Koval, and Andrey Ryabchenko. 2023. "The Structural–Rhythmological Organization of Coelogyne (Orchidaceae Juss.) Inflorescences" International Journal of Plant Biology 14, no. 1: 286-298. https://doi.org/10.3390/ijpb14010024
APA StyleKolomeitseva, G., Koval, V., & Ryabchenko, A. (2023). The Structural–Rhythmological Organization of Coelogyne (Orchidaceae Juss.) Inflorescences. International Journal of Plant Biology, 14(1), 286-298. https://doi.org/10.3390/ijpb14010024