1. Introduction
Plants experience stress facing unfavourable growth conditions. This is not only crucial in understanding the impact of unfavorable conditions on plant growth but also important for agriculture and food security. The unfavorable conditions may lead to yield loss of up to 70% for major crops, reflecting that average yields are limited to around 30% of their genetic potential [
1]. Abiotic stress caused by excesses or deficiencies in water, salt, light, temperature and nutrients, significantly lowers plant growth and productivity and even poses a threat to survival. Stressed plants have less-than-ideal environmental circumstances for development processes like cell division and expansion, which restricts plant advancement; dry-season stretch restrains plant growth since water is required for cell turgor, which applies weight on the cell development. Cold stress limits enzyme activities and other proteins, thus reducing plant growth.
The decelerated growth of plants under stressful conditions is not merely a passive outcome of unfavorable surroundings. In fact, stressed plants actively reduce their growth rate in an attempt to adapt to the stressor through stress-triggered cell signaling. When exposed to environmental stressors, plant cells exhibit upregulated expression levels of defense genes that encode for proteins such as pathogenesis-related (PR) and trypsin protease inhibitors (PI), which serve as protection against herbivory and pathogen attacks, respectively. Furthermore, secondary metabolites like osmoprotectants and toxins are synthesized at higher rates by these same plants with deterrent properties [
2]. Plants that swiftly respond to external stimuli can benefit from enhanced fitness due to timely defense reaction mechanisms against abiotic and biotic challenges activated via various defense signaling pathways resulting in gene expression changes [
3].
Volatile organic compounds (VOCs), a type of phytochemical among secondary metabolites, are responsible for chemical signaling molecules that are involved in intra- and interplant communication, benefiting the health of both the emitter and adjacent beneficiary plants [
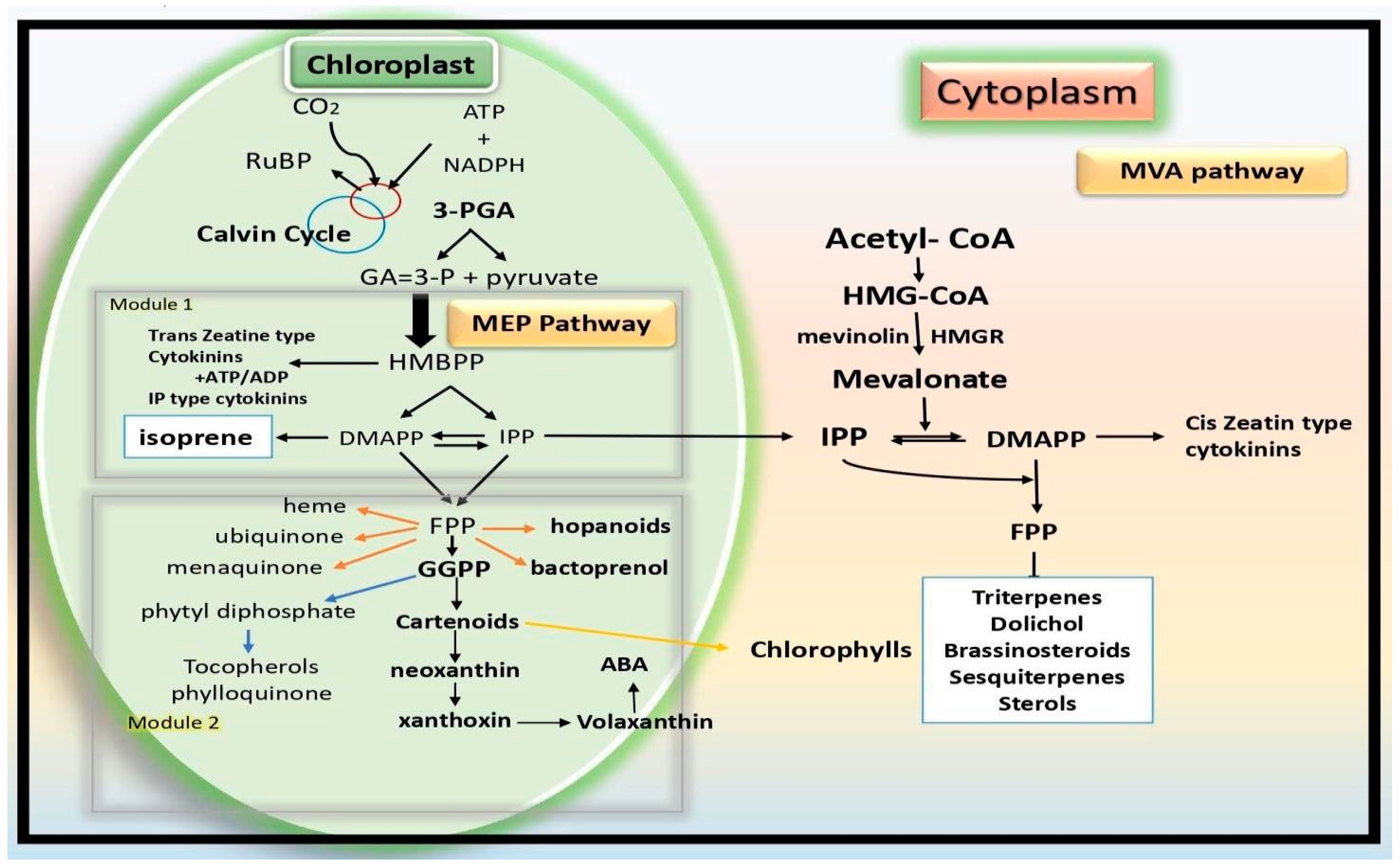
4]. Isoprene (C5H8), which is small and volatile, is produced at the chloroplast through the 2-C-methyl-
d-erythritol 4-phosphate (MEP) pathway. The synthesis of one isoprene molecule requires 14 NADPH and 21 ATP, indicating that it is energy- and carbon-intensive. The final step in the production of isoprene involves converting dimethylallyl diphosphate (DMADP) to isoprene, which is catalyzed by the enzyme known as “isoprene synthase” (IspS). In addition to producing hemiterpene isoprene, DMADP serves as a substrate for synthesizing several other important molecules such as monoterpenes, pigments including carotenoids, chlorophyll prenyl chains and cytokinins; hormones like abscisic acid (ABA); and pigments too [
5].
The production of isoprene in higher plants is reliant on light. Nonetheless, a recent study showing that algae can generate and disseminate isoprene heterotrophically challenges the notion that all photosynthetic organisms are only capable of producing isoprene under light-induced conditions. Bacteria have the capacity to manufacture isoprene via both mixotrophic and heterotrophic pathways through MEP, while nonphotosynthetic creatures such as animals also possess this ability. Isoprene emission is significantly influenced by temperature due to its low evaporation point of 34 °C, which accounts for its highly volatile nature. Within physiological limits, an exponential rise in the synthesis and emission of isoprene can be attributed to this factor along with elevated Q10 values associated with IspS [
5].
Due to its conjugated double bonds, this isoprene exhibits rapid reactivity towards various reactive chemical species, predominantly producing macrolein (MAC) and methyl vinyl ketone (MVK). These substances are toxic and swiftly absorbed by plants, thereby necessitating their detoxification. Generally, the concentration of isoprene in intercellular airspaces of highly emitting leaves ranges from 20 to 60 µL L
−1; however, it appears notably low in thylakoid membranes. At a global level, the amount of isoprene released into the atmosphere is approximately 500–600 Tg C yr
−1, whereas during summer days, simulated daily isoprene emission from a single leaf amounts to approximately 1–2 mg C m
−2 leaf d
−1 [
6].
The catalysis of isoprene synthesis, which is produced from DMADP via the MEP pathway, relies on the presence of isoprene synthase (IspS). This specific enzyme serves as the sole dedicated catalyst within this sub-branch of the primary terpene biosynthesis pathway. Henceforth, a plant can emit isoprene by simply possessing this chloroplast-localized IspS enzyme (
Figure 1). Since the initial isolation of the isoprene synthase sequence, numerous functional investigations have utilized this characteristic of IspS to either modulate isoprene emission in natural non-emitters or entirely eliminate it from natural emitters. These studies concentrate on upregulating or downregulating IspS genes from different species.
The first isolated IspS gene originated from Arundo donax L., a monocot species that belongs to the Poaceae family (AdoIspS), which is an excellent energy crop with increased growth rate along with resistance to both types of stressors. Li and colleagues discovered that when exposed to heat and osmotic pressure, the AdoIspS gene exhibits multiple upregulations. Additional research revealed differences in isoprene emission rates between the two ecotypes of Arundo donax, where higher levels of emitted isoprene were associated with greater tolerance to drought and faster recovery [
7]. These findings suggest that A. donax’s isoprene emissions may have positive contributions towards plant adaptation under abiotic stresses. Extensive research has primarily compared physiological responses among transgenic isoprene emitters and corresponding plants, as well as natural and non-emitting counterparts under various abiotic stresses [
6]. GA is composed of 19 or 20 carbon atoms in their tetracyclic structure. Their de novo synthesis begins with GGDPD, a typical precursor of C20 diterpenoid compounds. At first, it was thought that the MVA pathway is the source of the isoprene units required for the synthesis of GGDP. However, recent research employing isotope-labelled precursors revealed that the cytosolic MVA pathway contributes very little to the production of GA, with the majority coming from the plastidial MEP pathway [
8].
The initial stages of GA biosynthesis are identical to those of ABA biosynthesis. Specifically, C5 isoprenoid units are generated in plastids through the MEP pathway and then joined head-to-tail to form the linear isoprenoids GPP, FDP and finally GGDP. Experiments with plants that disrupted the normal transformation of 1-deoxy-d-xylulose 5-phosphate by 1-deoxy-d-xylulose 5-phosphate reductoisomerase into the branched compound 2-C-methyl-d-erythritol 4-phosphate (MEP) demonstrated the significance of the MEP pathway in GA and ABA biosynthesis. In the current review, we are trying to explore the unknown plant physiological and molecular-level functions of isoprene in plant systems such as isoprene-derived signaling molecules, volatile communications between plants and molecular aspects; future prospective studies will benefit from this manuscript.
2. Isoprene-Derived Signaling Molecules in Plant System
Plants use volatile organic compounds (VOCs) to communicate with other living things. Isoprene, monoterpenes and sesquiterpenes make up the largest class of volatile organic compounds (VOCs) emitted by plants terpenes. In plant–plant interactions, mono- and sesquiterpenes are well-known communication molecules. On the other hand, isoprene, the smallest and most often released terpene, is instead given a role in fighting abiotic stressors. Different isoprene-derived signaling molecules in plant systems are featured in this part.
2.1. Brassinosteroids
Brassinosteroids (BRs) are a class of signaling molecules with a steroidal skeleton with four rings. They play a noticeable role in the hormonal network that regulates numerous critical physiological processes in the plant life cycle. BRs play a role in male fertility, flowering and germination and control the lengthening of organs, the onset of senility and the plant’s capacity to withstand stresses like heat, salinity and water during the vegetative growth phase [
9]. BRs are steroid alcohols similar to other plant sterols. They share a structural resemblance with cholesterol, the main C27 animal sterol. BRs, along with other phytosterols, can have 27–29 carbon atoms, depending on how side chains are substituted (i.e., the type of C-24 alkyl substituent). According to [
10], C28 phytosterols, like BL, are the most abundant in plants. The entire steroid family requires the linear C30 hydrocarbon squalene, which insects cannot cyclize. In contrast, plants, animals and humans have access to the full sterol biosynthesis pathway. The cytoplasmic mevalonic acid pathway is the source of the isoprenoid building blocks IPP and DMAPP, which are used in BR synthesis.
The carbon skeleton of BR C27, an analog of BR C28 without the methyl group at C24 (norBRs), is identical to that of cholesterol, suggesting that they are synthesized from cholesterol via the same pathway as CS from CR. Additionally, the methylation of norCS at C24 in the presence of NADPH and S-adenosyl-l-methionine was shown to induce CS formation in cell-free enzymatic extracts of
Arabidopsis thaliana [
11].
Sitosterol, a naturally occurring C29 phytosterol, can form C29 BRs and 24-ethylBRs (also known as homoBRs) when combined with an enzyme extract from cell-free rice seedlings [
12]. The final biosynthetic product in this instance is homoCS, which is produced by homoTE and homoTY. Strangely, no homoBL was observed in plant tissues, even though similar processes described above result in the synthesis of either BL or its equivalent analog. Additionally, it appears that C28 demethylation of C29 BRs can produce C28 BRs, and this interconversion may enable plants to restore a BR bioactivity level that is ideal for different physiological procedures, as C29 BRs are biologically less active than C28 BRs.
2.2. Abscisic Acid
A nonvolatile C15 terpenoid is ABA carboxylic acid, and it was initially presumed that it is made directly from the precursor of the C15 sesquiterpene FDP, or farnesyl diphosphate. Still, trials involving precursors with 18O labels showed that ABA is produced by the C40 carotenoids cleaved from the MEP pathway. Violaxanthin is either converted to 9-
cis-violaxanthin or 9′-
cis-neoxanthin by forming transneoxanthin (C40) [
13,
14]. This process aids in protecting plant tissues from photooxidative stress [
15]. The neoxanthin-deficient Arabidopsis mutant aba4 showed a decrease in ABA production, indicating that neoxanthin is the source of ABA, particularly under water-stress situations. The process of xanthophyll neoxanthin production is still unknown, despite the substantial advancements in our knowledge of the carotenoid biosynthesis pathway. The 2-C-methyl-
d-erythritol 4-phosphate (MEP) route of isopentenyl diphosphate (IPP) synthesis produces carotenoids inside plastids. Plants have virtually figured out the production process for carotenoid pigments. But ABA was not entirely eliminated in the mutant, suggesting that cisviolaxanthin could potentially be used to produce ABA. Furthermore, although lacking 90-cis-neoxanthin, the parasitic plant
Cuscuta reflexa can nevertheless produce ABA [
16]. The cleavage of both xanthins by 9-cisepoxycarotenoid dioxygenase (NCED) to release the C15 compound xanthoxin (C15H22O3) into the cytosol is the last plastidial step in ABA biosynthesis, and this step limits the rate of ABA biosynthesis.
2.3. Cytokinins
Another class of naturally occurring low-molecular-weight substances, cytokinins (CKs) bind to specific receptors and set off a series of reactions that control plant growth and development. They are mostly recognized by biologists as the chemicals that, along with another hormone called auxin, regulate cell division (cytokinesis). In addition, they play a role in the development of lateral shoots, vascular differentiation, apical dominance, senility and other processes. CKs can have an aromatic side chain or an isoprenoid side chain at the N-6 position of their adenine moiety, as indicated in the introduction. Plant cells can produce isoprenoid CKs through the transfer of a donor C5 isoprenoid unit to an adenine molecule that serves as an acceptor, which can be a tRNA-bound species or a free nucleotide [
17]. DMAPP and HMBPP are the currently recognized viable isoprenoid donors. Both the MEP and MVA pathways in plastids and the cytosol of eukaryotes can produce DMAPP.
Transferring the isoprenoid moiety to adenine, either in its nucleotide forms or coupled to RNA, is the first step in the production of isoprenoid cytokinins. Dimethylallyl pyrophosphate (DMAPP) and (E)-4-hydroxy-3-methyl-but-2-enyl diphosphate are the isoprenoid side chain donors that have been discovered to date. Cytochrome P450 monooxygenase has the ability to further hydroxylate the side chain in the event of DMAPP attachment and isopentenyladenine-type cytokinin production [
18]. Following their eventual liberation from tRNA, the cytokinin nucleotides are hydrolyzed to produce free bases. DMAPP and HMBPP are the currently recognized viable isoprenoid donors. Both the MEP and MVA pathways in plastids and the cytosol of eukaryotes can produce DMAPP.
The MEP pathway active in bacteria and parasites produces only HMBPP [
19]. Adenylate isopentenyl transferases (IPTs) are responsible for catalyzing the N-prenylation of adenine nucleotides. Two types of IPTs are identified: (1) An IPT that functions by adding an isopentenyl unit to a free adenine nucleotide (EC 2.5.1.27) [
20]; and (2) an IPT that performs the same function for tRNA-binding adenine units (EC 2.5.1.8). The isopentenyladenine (iP) nucleotide is the main product of reactions in which DMAPP acts as the IPT’s substrate. On the other hand, the trans-zeatin (tZ) nucleotide is produced by reactions involving the hydroxylated isoprenoid substrate HMBPP (
Figure 2) [
21,
22].
2.4. Strigolactones
Of all the groups, strigolactones (SLs) are newly identified signaling molecules in plants that have hormonal activity [
23]. The primary cause for this discovery was their apparent part in preventing the division of shoots [
24,
25], which seems to be governed by a mechanism encompassing the interaction of auxins, CKs and SLs. Together, SLs also seem to control leaf senility via ethylene, which is gaseous [
26], and to strategically modify overall development to regulate other physiological processes like the distribution of resources in balance. To increase phosphate intake, for example, phosphate scarcity causes plants to uplift SL biosynthesis. This results in modifications to root elongation and the encouragement of fungal symbiosis, indicating that some SLs serve as rhizosphere-signaling molecules [
27]. As per [
28], about 20 SLs are identified and chemically characterized from root exudates of different terrestrial plants. According to [
29], they are tricyclic lactones with three fused rings, A, B and C, along with a butenolide ring (D) joined by an enol ether bond.
3. Volatile-Mediated Communication within and between Plants
Actually, rather than between plants, it is possible that VOC-based plant-to-plant communication has generally evolved to speed up signaling within plants. The air phase appears to be a desirable substitute for phloem-based transport, especially for larger trees, and it may aid in the transmission of information from one area of the tree to another. The observation that volatile-mediated transmission works better between genetically related plants than between different species supports the evolutionary genesis of plant-to-plant communication.
Plants have developed special means of communication between different organs for the improvement of their growth and the provision of stress resistance. When a plant senses environmental stress, it will send out multiple long-range signals to initiate the plant’s vasculature to assist in systemic stress responses. Multiple stresses cause these signals to overlap in frequency and amplitude, which makes it challenging to link a given response to a particular stressor (
Table 1).
Information about the surroundings can travel from the plant’s roots to its aerial portions and vice versa through carefully planned long-distance and/or cell-to-cell communications. To activate defense responses in the target tissues and enable plant adaptation to overall stresses at the overall plant level, root-to-shoot or shoot-to-root signaling is essential. The modes of action and propagation speeds of these potentially related signals, which comprise chemical, electrical and hydraulic signals, vary [
33].
It is widely known how volatiles function in both intra- and interspecific communication, as is information about their mode of action along with chemical diversity [
34]. Additionally, they act as signaling molecules in intra- and interplant communication, enhancing the fitness of the “emitter” plant and altering the behavior of nearby “receiver” plants. Plants hold the potential to discriminate among various volatile blends. Since VOCs are not species-specific, plants thus create distinct messages by manipulating the constituent parts of a volatile bouquet [
35]. These unique messages then elicit particular reactions from the recipient. For example, the parasitic dodder plant Cuscuta pentagonia uses a specific mixture of VOCs as signals to find host plants [
36]. Nonhost dodder plant (Z)-3-hexenyl acetate was reported to be absent in wheat and tomato VOC mixtures, indicating that it acted as a dodder repellent.
Herbivore-induced plant volatiles (HIPVs) and pathogen-induced volatiles (PIPVs) are the responses that plants release in response to insect and pathogen attacks during biotic stress. Neighboring hetero- or conspecifics may take advantage of these volatiles, to the advantage or jeopardizing the health of the emitter plant. The PIPVs or HIPVs can be advantageous to the emitting plant by drawing pollinators and natural enemies of its herbivores, or they can be detrimental to the plant by serving as cues for parasitic plants to forage and for herbivores to locate their host [
36].
It is noteworthy to consider that in certain HIPV-mediated plant interactions, receiver plants have been observed to absorb volatile organic compounds (VOCs) from their adjacent stressed neighbors and transform them into deadly derivatives that are harmful to the growth and survival of herbivores. As a form of associational resistance (AR), the receiving plant may occasionally rerelease volatile organic compounds (VOCs) that it has adsorbed from the parent plant to repel insects and pests [
37]. VOC emissions in plants have also been demonstrated to be induced by multiple abiotic factors, including nutrient availability, higher light intensity, UV radiation, salinity, temperature, ozone exposure and wind, as well as mechanical damage [
38].
VOCs improve the thermotolerance of photosynthetic tissues in reply to abiotic stress and protect against exposure to high temperatures. By either quenching ROS species or changing ROS signaling, plant volatile organic compounds (VOCs) can stabilize and protect cellular membranes against the harmful effects of oxidative stress caused as a consequence of stimuli in the environment. In reducing the negative consequences of abiotic and biotic stressors, the function and mode of action of stress-induced VOCs are encapsulated [
39].
4. Isoprene as a Hormone Controlling the Growth and Antioxidant System of Plants
The origins of isoprenoids are shared among various phytohormones and signaling molecules in plants [
40]. While monoterpenes (C10) have recently demonstrated a significant role in transmitting plant-to-plant defense signals and contributing to systemic acquired resistance (SAR) against pathogens [
41], isoprene (C5) was initially overlooked for its signaling role. However, recent recognition has established isoprene as a plausible mechanism of action for its effects on plants. The discovery that Arabidopsis plants, which do not naturally release isoprene, undergo transcriptome reprogramming when exposed to isoprene fumigation [
42] has garnered recognition for investigating isoprene’s role. Despite the challenge in perceiving isoprene’s significance, numerous reports linking isoprene to plant development suggest its likely role as a stress-signaling molecule. Recent findings in Arabidopsis and poplar plants revealed the active IspS gene promoter in various plant areas, including roots, developing lateral roots, root hairs, vascular tissue and root caps [
43].
Treatment with indole-3-acetic acid increased the promoter activity, and engineered poplar plants lacking the isoprene gene exhibited enhanced lateral root formation. Isoprene-producing Arabidopsis plants, engineered with the IspS gene from Eucalyptus globulus, displayed increased cotyledon, hypocotyl, leaf and inflorescence growth, indicating that isoprene may directly influence growth through gene expression regulation [
44]. In contrast, another study showed that Arabidopsis plants transformed with the IspS gene from Arundo donax, producing isoprene, exhibited reduced growth inhibition after receiving exogenous ABA treatment. On the contrary, tobacco lines emitting isoprene exhibited detrimental effects on the growth of their leaves and stems. However, these plants displayed increased hypocotyl growth, a phenomenon also observed in Arabidopsis. Consequently, isoprene appears to exert regulatory influences on plant development, manifesting both positive and negative effects. Isoprene demonstrates a genuine hormonal impact owing to its structural resemblance to zeatin and other chloroplast cytokinins produced through the MEP pathway. The collaboration between cytokinins and isoprene may play a crucial role in governing leaf development and lifespan, despite isoprene being released in quantities that might be considered excessively high for a conventional hormonal action [
5].
5. The Interplay between ROS and Isoprene
Isoprene is recognized for its protective role against various abiotic stresses in plants. Plants releasing isoprene exhibit enhanced resilience to oxidative stressors like ozone, heat stress and other reactive oxygen species (ROS). The ability of isoprene to quench nitric oxide, hydrogen peroxide, ozone and singlet oxygen has fueled speculation about its direct antioxidant activity. However, uncertainties persist regarding whether isoprene scavenges ROS within the leaf boundary layer or within leaves after exiting through stomata [
45].
An indirect assessment of the interplay between isoprene and ROS in leaves involved monitoring increased emissions of mevalonic acid (MVA) and methyl anthranilate (MAA) from isoprene-producing plants exposed to elevated temperatures. It is now recognized that sources other than isoprene can generate similar secondary oxidants within leaves [
44]. Isoprene-emitting plants demonstrate prolonged preservation of high photosynthetic activity under abiotic stresses, suggesting a direct shielding effect of isoprene on the photosynthetic machinery. Beyond ROS quenching, isoprene is shown to enhance the stability of thylakoid membranes [
46].
Numerous studies have aimed to elucidate the precise mechanism through which isoprene protects photosynthesis. Thylakoid membranes in the chloroplasts of isoprene-emitting plants exhibit increased resistance to denaturation, as evidenced by a smaller rise in the nonphotochemical quenching of chlorophyll fluorescence (NPQ) compared to non-emitting plants. This suggests that NPQ remains stable because thylakoids are less susceptible to ROS, which are scavenged by isoprene before reaching the membranes during stressful conditions [
44]. Isoprene-emitting plants consistently exhibit lower NPQ compared to non-emitting plants, even under highly efficient photosynthetic conditions. The lipophilic nature of isoprene may contribute to its ability to intercalate within membranes, reinforcing their lipidic layer [
47].
6. Isoprene’s Role as a Signaling Molecule in Gene Networks
The investigation into RNA interference (RNAi) technology’s impact on inhibiting isoprene release in gray poplar [
48] and the examination of isoprene-fumigated Arabidopsis thaliana [
42] offer fresh insights into how isoprene induces alterations in the expression of vital gene regulatory networks crucial for plant development and stress responses. Isoprene’s influence extends to the metabolome [
49] and metabolic fluxes in plants [
48,
49]. Observations indicating a potential impact of isoprene on ATP synthase functionality within thylakoid membranes were noted in Arabidopsis plants expressing IspS, supporting the hypothesis of isoprene interacting with signal transduction networks through membrane-bound proteins [
42].
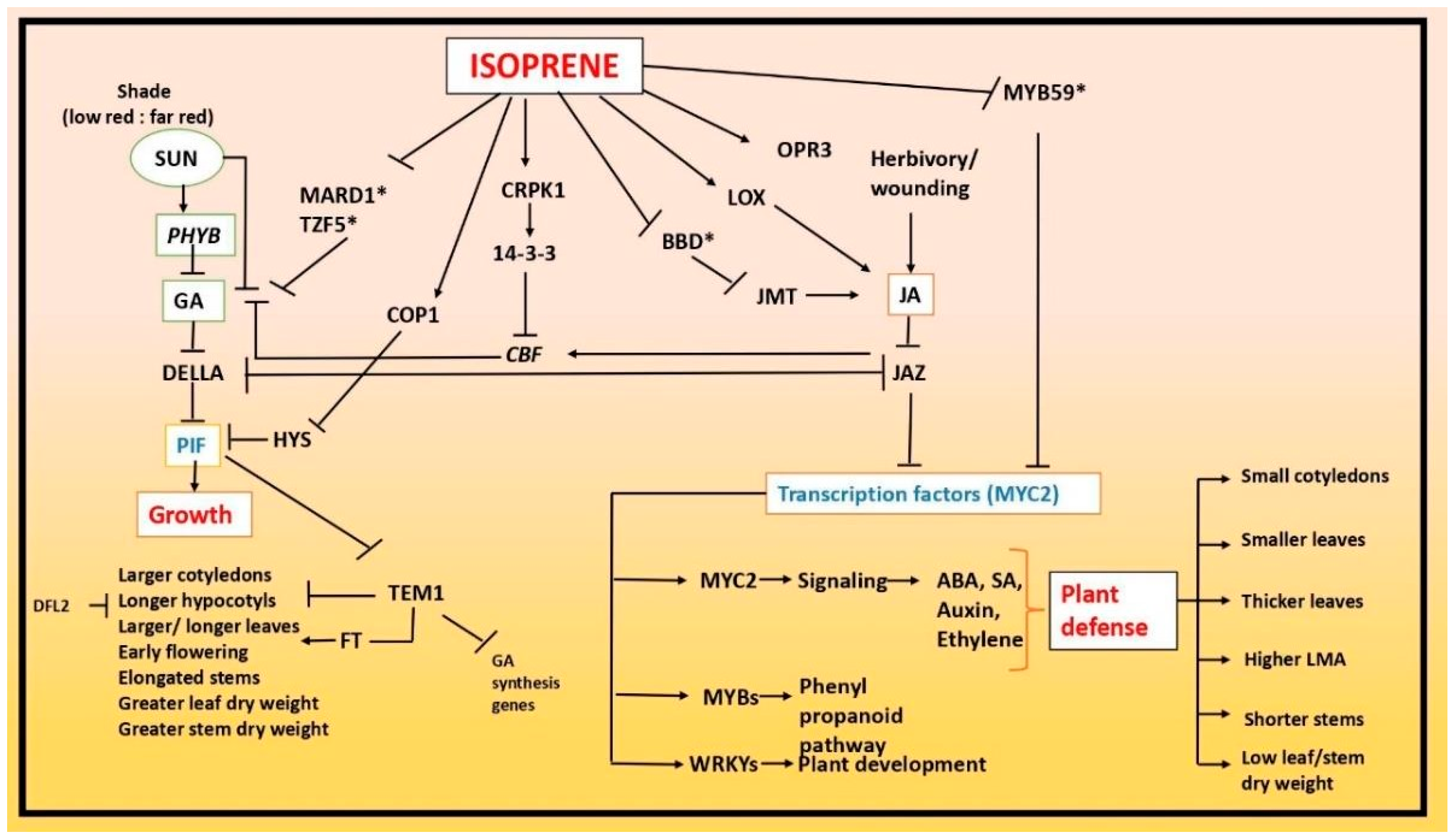
A 24 h fumigation of Arabidopsis plants with a physiologically relevant concentration (20 mL L21) resulted in altered gene expression, as revealed by microarray analysis. The affected genes encode proteins related to photosynthetic light reactions, cell wall synthesis, phenylpropanoid and flavonoid biosynthesis and stress responses. Harvey and Sharkey [
42] postulated that isoprene may function as a signaling molecule, altering gene expression and consequently impacting abiotic stress tolerance. However, the specific genes and pathways responsive to isoprene remain unknown within the growth- and stress-related signaling pathways. Transcriptome analyses demonstrated that plants exposed to isoprene either internally (Arabidopsis and tobacco expressing IspS) or externally (tobacco expressing IspS) had altered expression of key genes important for the synthesis of specific growth regulators as well as key components of these growth-regulator signaling pathways. To promote the amplification of ethylene signaling, gibberellins (GA), cytokinin (CK), jasmonic acid (JA) and gibberellins, isoprene changed the expression of certain genes. The genes BBD1, BBD2, LOX1, LOX3, LOX4, LOX5, OPR3 and MYB59 are among the changed genes linked to JA. Cytokinin-affecting genes include WIND1 and KFB20. Gibberellins are affected by TEM1, DFL2, MARD5, TZF5 and others. The ethylene-related genes ACO4, EBF1 and EBF2 are listed. Key genes involved in the signaling pathways of auxin (MYB73, BPC and KFB20), salicylic acid (SA; CBP60A, ATAF1, EBF1 and EBF2) and brassinosteroid (BR; BRI1) were found to have altered in transcription. This suggests that these pathways are downregulated when isoprene is present. The study revealed that isoprene enhanced the expression of genes from the abscisic acid (ABA) signaling branch that are necessary for drought tolerance (NCED3, NCED5 and ATAF1). Conversely, genes linked to ABA signaling that are associated with seed dormancy (TZF5, MARD1, NCED4 and MYBS2), pathogen resistance (BBD1, BBD2 and MPK7), heat stress (NCED4) and salt stress (CIPK20) appeared to be downregulated [
43].
7. Priming: Defense Strategies in Plants
Plant stress physiology increasingly explores priming as a practical and effective tool, representing hormesis. Priming, linked to induced resistance (IR), renders plants more sensitive to future attacks, providing inherited immunological memory that persists throughout the plant’s life cycle. IR offers broad-spectrum protection against various environmental stresses, placing the plant in a state of alertness for potential future challenges [
44].
Successful defense priming requires resilient defense mechanisms with low fitness costs. Recollection of a prior stressful incident implies a more effective response to the actual stress challenge [
50]. Recognition of specific volatile plumes, influencing focused defense responses, characterizes priming with volatile organic compounds. Factors such as exposure length, distance from the source plant and the plant’s sensitivity impact how recipient plants respond to volatile signals, priming their defense mechanisms. The ability of emitting-power plants to adapt with low costs, triggered by lower airborne volatile organic compound concentrations, suggests the potential ecological fitness benefit of priming, enabling stronger and quicker defenses against impending stress [
51].
8. Exploring the Molecular Dimensions of Isoprene in Plant Systems
The ability to investigate isoprene-induced alterations in plant biology, spanning from genetic to phenotypic levels, has been made feasible by the existence of transgenic plants where isoprene synthesis is either induced or suppressed (
Table 2). In general, these studies have revealed that, in comparison to non-emitting plants cultivated under identical conditions, isoprene-emitting plants display distinctive profiles in gene expression, proteomics and metabolomics. Under conditions of high light and heat stress, Populus spp. with reduced expression of the IspS gene exhibited a decline in the expression of genes associated with the biosynthesis and regulation of phenylpropanoids. Additionally, downregulated genes involved in the synthesis of condensed tannins and anthocyanins were observed. Tolerance of ozone and other reactive oxygen species (ROS) is isoprene’s second function. Isoprene can stop ozone exposure-related visual damage as well as quantifiable losses in photosynthetic ability, and isoprene emissions have the potential to worsen ozone depletion. High temperatures enhance the amount of IspS protein and gene expression, but excessive ozone causes the opposite effect [
52]. Thus, whereas isoprene offers protection against ROS and heat flecks, there seems to be a connection between the physiology of isoprene emission and heat fleck protection.
The ability to synthesize isoprene has undergone several evolutionary changes, most likely from a reservoir of genes encoding monoterpene synthase. The substrate and product specificity of IspS genes can be readily altered by small changes in gene sequence [
53]. That all IspS genes have a common origin is not supported by gene sequences. Rather, it seems that the development of isoprene emission is more akin to that of C4 metabolism, which emerged many times as a response to an environmental limitation.
Table 2.
A summary of biological tasks of isoprene, with associated roles in the plant.
Table 2.
A summary of biological tasks of isoprene, with associated roles in the plant.
| S.No. | Isoprene Role | Functional Expectations | Experimental Evidence | Reference |
|---|
| 1 | Antioxidant | Safeguarding against oxidative harm by directly interacting with oxidative molecules such as reactive oxygen species (ROS) or nitric oxide (NO). | PRO: Isoprene inhibits ROS and reactive nitrogen species, promoting resistance to thermal and oxidative stresses. | [54] |
| 2 | Defense priming stimulus | Enhancing resistance to stress by increasing the perception, transmission and/or initiation of defense mechanisms in response to stress signals. | PRO: Exposure to isoprene triggers resistance for bacterial infection in Arabidopsis through activating defense mechanisms dependent on salicylic acid. | [55] |
| 3 | Stress signal | Elevated production following stress and its function as a signal for transmitting stress to other parts of plant and/or neighboring plants. | PRO: Isoprene emission exhibits swift alterations in reaction to both wounding and temperature changes. | [54] |
| 4 | Hormone | Control at the gene level of plant processes occurs at extremely low concentrations. | PRO: Isoprene works as a suppressor of reactive oxygen species (ROS) production by influencing gene expression. | [56] |
Isoprene significantly influences the expression of genes related to defense against both abiotic and biotic stresses, signaling pathways for growth regulators, photosynthesis, seed germination and the growth of seedlings and plants. This phenomenon has been demonstrated in both non-emitting Arabidopsis exposed to isoprene under unstressed conditions and in Arabidopsis and tobacco plants expressing IspS and emitting isoprene. The majority of these genes are implicated in phenylpropanoid biosynthesis and regulation, cytokinin-mediated wound repair and defense signaling facilitated by jasmonic acid (JA) [
49].
In a specific experiment, Arabidopsis plants were exposed to isoprene for an entire day under nonstressful conditions. This led to the induction of transcription factor-enriched gene networks, phenylpropanoid biosynthesis, translation machinery and chloroplast genes. Another study found that exogenous ABA supplementation heightened the sensitivity of Arabidopsis plants capable of producing isoprene due to IspS insertion, leading to the upregulation of the RD29B gene, which plays a role in the abscisic acid-activated signaling pathway [
44].
However, in contrast to non-emitting Arabidopsis, under conditions of water scarcity, isoprene-emitting plants exhibited downregulation of the P5CS gene, involved in protecting against reactive oxidative species, and the COR15A gene, which protects stromal proteins. While numerous studies have explored transcriptomic variations resulting from isoprene biosynthesis, only two delved into the impact of isoprene production capacity on proteomics. Analysis of chloroplast proteins revealed that transgenic poplars with suppressed isoprene emission displayed reduced expression in chloroplast proteins related to photosynthesis, light reactions, redox regulation, defense against oxidative stress and overall metabolism in comparison to naturally isoprene-emitting poplars.
Recent proteomic research further unveiled distinct protein patterns between isoprene-emitting and non-emitting poplars. Specifically, non-emitting plants exhibited elevated levels of proteins involved in the biosynthesis of carotenoids and flavonoids compared to isoprene-emitting plants. Conversely, proteins associated with phenylpropanoid and flavonoid biosynthesis were found at lower concentrations, suggesting a potential reprogramming of the entire secondary metabolism and a trade-off between metabolites synthesized through the MEP pathway (carotenoids vs. isoprene). However, the veracity of this assertion remains a subject of debate, as elaborated in subsequent sections. Lastly, examinations of the impact of isoprene emission on the plant metabolome have consistently indicated that in poplar plants with inhibited isoprene synthesis, especially under elevated temperatures and intense light conditions, the reduction in total phenolics and condensed tannin concentrations is correlated with the decreased expression of genes involved in secondary metabolism [
57].
9. Technologies for Investigating Plant VOC Signaling Interactions
The enhanced understanding of plant defense responses triggered by volatile organic compounds (VOCs) has become possible due to advancements in bioinformatics, metabolomics, transcriptomics and volatilomics. Over the last decade, there has been noticeable progress in the development of sensitive and cost-effective VOC collection and analysis technologies [
58]. These advancements encompass techniques such as static and dynamic headspace (SHS, DHS) and gas chromatography–mass spectrometry (GC-MS).
Headspace VOC sampling methods, such as collection chambers by [
59], and nonsolvent sorptive extraction techniques like headspace solid-phase microextraction (HS-SPME) and purge-and-trap headspace (P&T-HS) for static and dynamic extraction, respectively, represent these developments [
60]. Through these techniques, live plants can be analyzed, providing a more representative volatile profile compared to conventional methods like steam distillation or solvent extraction. However, it is essential to consider potential alterations to the plant’s microenvironment during headspace collection.
Transpiration rates, along with the composition and emission rates of volatile organic compounds (VOCs), are anticipated to be influenced by rising temperatures, increased humidity and reduced light intensity within chambers. Additionally, to prevent the potential scalping of VOCs by hydrophobic materials, nonreactive substances like polytetrafluoroethylene (PTFE) or fluorinated ethylene propylene (FEP), which have demonstrated success in plant VOC sampling, may be required for VOC adsorption. These materials are commonly found lining collection chambers or sampling bags [
61].
In the past decade, new approaches have emerged. One such trend is direct analysis in real time (DART), an ionization technique facilitating rapid, noncontact detection of analytes on solid or liquid surfaces. Portable and field-appropriate devices include the GC-PID, a portable gas chromatograph measuring photoionization, and the VOC analyzer FROG-4000 [
62]. Additionally, microversions of these detectors, known as µGC–PIDs, have demonstrated the capability for swift vapor analysis in field settings [
63].
Another portable analytical tool is the FlavourSpec, which analyzes headspace volatiles from liquid or solid samples without requiring pretreatment. It comprises a gas chromatograph (GC) linked to an ion mobility spectrometer (IMS). Utilizing silicone tubing filled with polydimethylsiloxane (PDMS) presents a cost-effective method for manipulating organisms while minimizing headspace during field sampling. This involves headspace sampling using small sections of tubing in conjunction with thermal desorption (TD)-GC-MS analysis [
29]. Proton-transfer-reaction mass spectrometry (PTR-MS), a GC-independent technique, has also proven valuable, particularly for real-time VOC monitoring both in the field and in the laboratory.
10. Conclusions and Future Perspectives
Exploring research that assigns various purposes to the isoprene released by plants reveals a narrow understanding. Initially thought to primarily possess an antioxidant effect, isoprene is now recognized for influencing an increasingly complex web of processes and mechanisms. This seemingly simple volatile substance, easily absorbed by plants and released into the surrounding air, emerges as a multipurpose and beneficial molecule functioning as an infochemical, priming defensive pathways, collaborating with hormones and acting as an antioxidant. The widespread release of isoprene by plants globally may find a more fitting explanation in the multitude of mechanisms that support them throughout their life cycle and in the face of adversity.
While certain secondary metabolites (SMs) act as herbivore deterrents, many directly hinder the growth of pests and pathogens, with the majority contributing to maintaining redox balance by scavenging reactive oxygen species (ROS), thereby enhancing stress tolerance in plants. Several primary transcription factors discussed in this review exhibit close relationships with the expression of one or more genes involved in the SM biosynthesis pathway. In controlling the linked genes, specific circumstances may necessitate a network of two or more TFs.
Several tasks remain in the domain of metabolite profiling of volatile organic compounds (VOCs) in plant blends emitted under diverse environmental stress conditions. Identifying the specific receptors for VOCs, along with transcription factors (TFs) and other regulatory proteins participating in signaling pathways, also poses a continuing challenge. The technological advancements witnessed in bioinformatics, transcriptomics, metabolomics and plant physiology over the past decade are poised to significantly aid future endeavors. Integrating metabolomic, transcriptomic and physiological analyses in studies focused on volatile-mediated signaling and defense priming could enhance our understanding of VOC-induced defense responses in plants under stressors comprehensively. While the majority of research has been conducted in controlled settings, combining these findings with studies considering realistic plant growth environments, such as glasshouses and growth chambers, would be advantageous. Natural environments present simultaneous occurrences of multiple stresses, as greenery constantly interacts with both hetero- and conspecifics. In-depth examinations into nearby plants’ reactions to the ensuing VOC emissions could provide more conclusive findings about the viability of using VOC-mediated priming not only against different abiotic stresses in agricultural systems but also against biotic stresses.
There are different purposes for which plants emit isoprene. Plants appear to benefit from isoprene emission, but research that at first only suggested that isoprene had an antioxidant effect is now showing that isoprene influences an ever more complex web of processes and mechanisms. This straightforward volatile substance, which is easily absorbed by plants and released into the surrounding air, is becoming known for being a useful and multipurpose molecule that in addition to functioning as an antioxidant, also primes defense pathways, functions as an infochemical and interacts with hormones. Such a wide range of activities supporting plants throughout their lifespan and when faced with challenges may provide a better explanation for the widespread release of isoprene by plants globally.







{kind=link}
{kind=link}

