

Increasing Risk of Tick-Borne Disease through Growth Stages in Ticks

 , ,
, , {kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
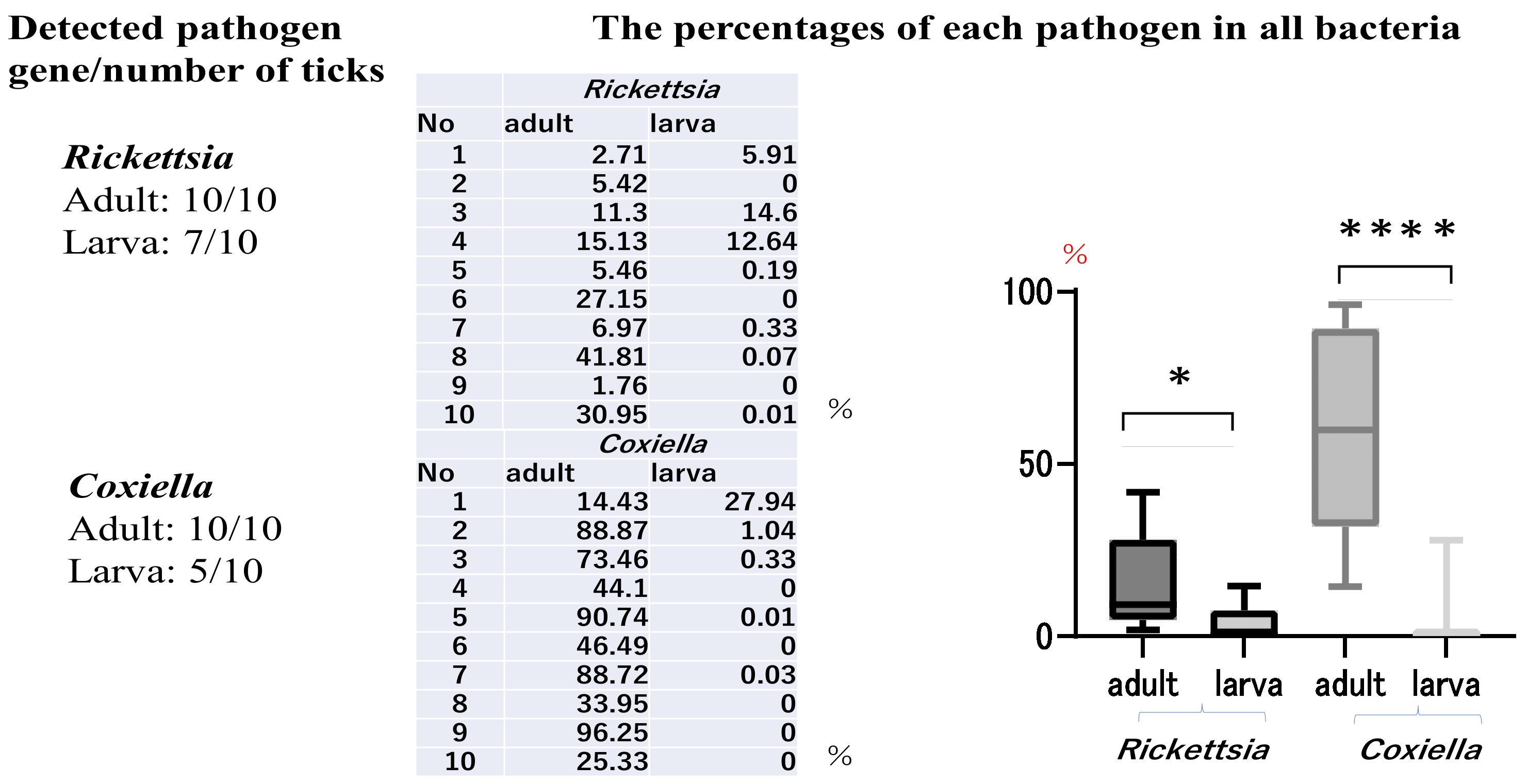
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kondo, M.; Nishii, M.; Gabazza, E.C.; Kurokawa, I.; Akachi, S. Nine cases of Japan spotted fever diagnosed at our hospital in 2008. Int. J. Dermatol. 2010, 49, 430–434. [Google Scholar] [CrossRef] [PubMed]
- Nakata, R.; Motomura, M.; Tokuda, M.; Nakajima, H.; Masuda, T.; Fukuda, T.; Tsujino, A.; Yoshimura, T.; Kawakami, A. A case of Japanese spotted fever complicated with central nervous system involvement and multiple organ failure. Intern Med. 2012, 51, 783–786. [Google Scholar] [CrossRef] [PubMed]
- Angelakis, E.; Raoult, D. Q fever. Vet. Microbiol. 2010, 140, 297–309. [Google Scholar] [CrossRef] [PubMed]
- Graves, S.R.; Gerrard, J.; Coghill, S. Q fever following a tick bite. Aust. J. Gen. Pract. 2020, 49, 823–825. [Google Scholar] [CrossRef] [PubMed]
- Thu, M.J.; Qiu, Y.; Matsuno, K.; Kajihara, M.; Mori-Kajihara, A.; Omori, R.; Monma, N.; Chiba, K.; Seto, J.; Gokuden, M.; et al. Diversity of spotted fever group rickettsiae and their association with host ticks in Japan. Sci. Rep. 2019, 9, 1500. [Google Scholar] [CrossRef] [PubMed]
- Krishnamoorthy, P.; Sudhagar, S.; Goudar, A.L.; Jacob, S.S.; Suresh, K.P. Molecular survey and phylogenetic analysis of tick-borne pathogens in ticks infesting cattle from two South Indian states. Vet. Parasitol. Reg. Stud. Rep. 2021, 25, 100595. [Google Scholar] [CrossRef] [PubMed]
- Sato, M.; Ikeda, S.; Arai, R.; Kato, M.; Aoki, J.; Nishida, A.; Watanabe, K.; Hirokawa, C.; Watanabe, K.; Regilme, M.A.F.; et al. Diversity and distribution of ticks in Niigata Prefecture, Japan (2016–2018): Changes since 1950. Ticks Tick Borne Dis. 2021, 12, 101683. [Google Scholar] [CrossRef] [PubMed]
- Chalada, M.J.; Stenos, J.; Vincent, G.; Barker, D.; Bradbury, R.S. A molecular survey of tick-borne pathogens from ticks collected in central Queensland, Australia. Vector Borne Zoonotic Dis. 2018, 18, 151–163. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Nakao, R.; Amin-Babjee, S.M.; Maizatul, A.M.; Youn, J.H.; Qiu, Y.; Sugimoto, C.; Watanabe, M. Molecular screening for Rickettsia, Anaplasmataceae and Coxiella burnetii in Rhipicephalus sanguineus ticks from Malaysia. Trop. Biomed. 2015, 32, 390–398. [Google Scholar] [PubMed]
- Morshed, M.G.; Lee, M.K.; Boyd, E.; Mak, S.; Fraser, E.; Nguyen, J.; Dykstra, E.A.; Otterstatter, M.; Henry, B.; Galanis, E. Passive Tick Surveillance and Detection of Borrelia Species in Ticks from British Columbia, Canada: 2002–2018. Vector Borne Zoonotic Dis. 2021, 21, 490–497. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, J.; Hashino, M.; Matsumoto, S.; Takano, A.; Kawabata, H.; Takada, N.; Andoh, M.; Oikawa, Y.; Kajita, H.; Uda, A.; et al. Detection of Francisella tularensis and analysis of bacterial growth in ticks in Japan. Lett. Appl. Microbiol. 2016, 63, 240–246. [Google Scholar] [CrossRef] [PubMed]
- Ushijima, Y.; Oliver , J.H., Jr.; Keirans, J.E.; Tsurumi, M.; Kawabata, H.; Watanabe, H.; Fukunaga, M. Mitochondrial sequence variation in Carlos capensis (Neumann), a parasite of seabirds, collected on Torishima Island in Japan. J. Parasitol. 2003, 89, 196–198. [Google Scholar] [CrossRef] [PubMed]
- Kondo, M.; Matsushima, Y.; Nakanishi, T.; Iida, S.; Habe, K.; Yamanaka, K. A novel transmission of Aeromonas hydrophila via tick bites. J. Dermatol. 2022, 49, e243–e245. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, M.; Oshita, S.; Yamazoe, N.; Miyazaki, M.; Takemura, Y.C. Important clinical features of Japanese spotted fever. Am. J. Trop. Med. Hyg. 2018, 99, 466–469. [Google Scholar] [CrossRef] [PubMed]
- Kondo, M.; Ando, K.; Yamanaka, K.; Mizutani, H. Rickettsia species AMONG ticks in an area of Japan endemic for Japanese spotted fever. Southeast Asian J. Trop. Med. Public Health 2017, 48, 585–589. [Google Scholar]
- Uchida, T.; Yan, Y.; Kitaoka, S. Detection of Rickettsia japonica in Haemaphysalis longicornis ticks by restriction fragment length polymorphism of PCR product. J. Clin. Microbiol. 1995, 33, 824–828. [Google Scholar] [CrossRef] [PubMed]
- Kondo, M.; Matsushima, Y.; Nakanishi, T.; Habe, K.; Yamanaka, K. Investigation of pathogens injected into the human body by ticks. Exp. Dermatol. 2022, 31, 1014–1015. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kondo, M.; Matsushima, Y.; Nakanishi, T.; Iida, S.; Koji, H.; Yamanaka, K. Increasing Risk of Tick-Borne Disease through Growth Stages in Ticks. Clin. Pract. 2023, 13, 246-250. https://doi.org/10.3390/clinpract13010022
Kondo M, Matsushima Y, Nakanishi T, Iida S, Koji H, Yamanaka K. Increasing Risk of Tick-Borne Disease through Growth Stages in Ticks. Clinics and Practice. 2023; 13(1):246-250. https://doi.org/10.3390/clinpract13010022
Chicago/Turabian StyleKondo, Makoto, Yoshiaki Matsushima, Takehisa Nakanishi, Shohei Iida, Habe Koji, and Keiichi Yamanaka. 2023. "Increasing Risk of Tick-Borne Disease through Growth Stages in Ticks" Clinics and Practice 13, no. 1: 246-250. https://doi.org/10.3390/clinpract13010022
APA StyleKondo, M., Matsushima, Y., Nakanishi, T., Iida, S., Koji, H., & Yamanaka, K. (2023). Increasing Risk of Tick-Borne Disease through Growth Stages in Ticks. Clinics and Practice, 13(1), 246-250. https://doi.org/10.3390/clinpract13010022