Ecological Niche Space of Fish Communities in Impounded Sections of Large Rivers: Its Application to Assessment of the Impact of Weirs on River Ecosystems



Abstract
:1. Introduction
2. Materials and Methods
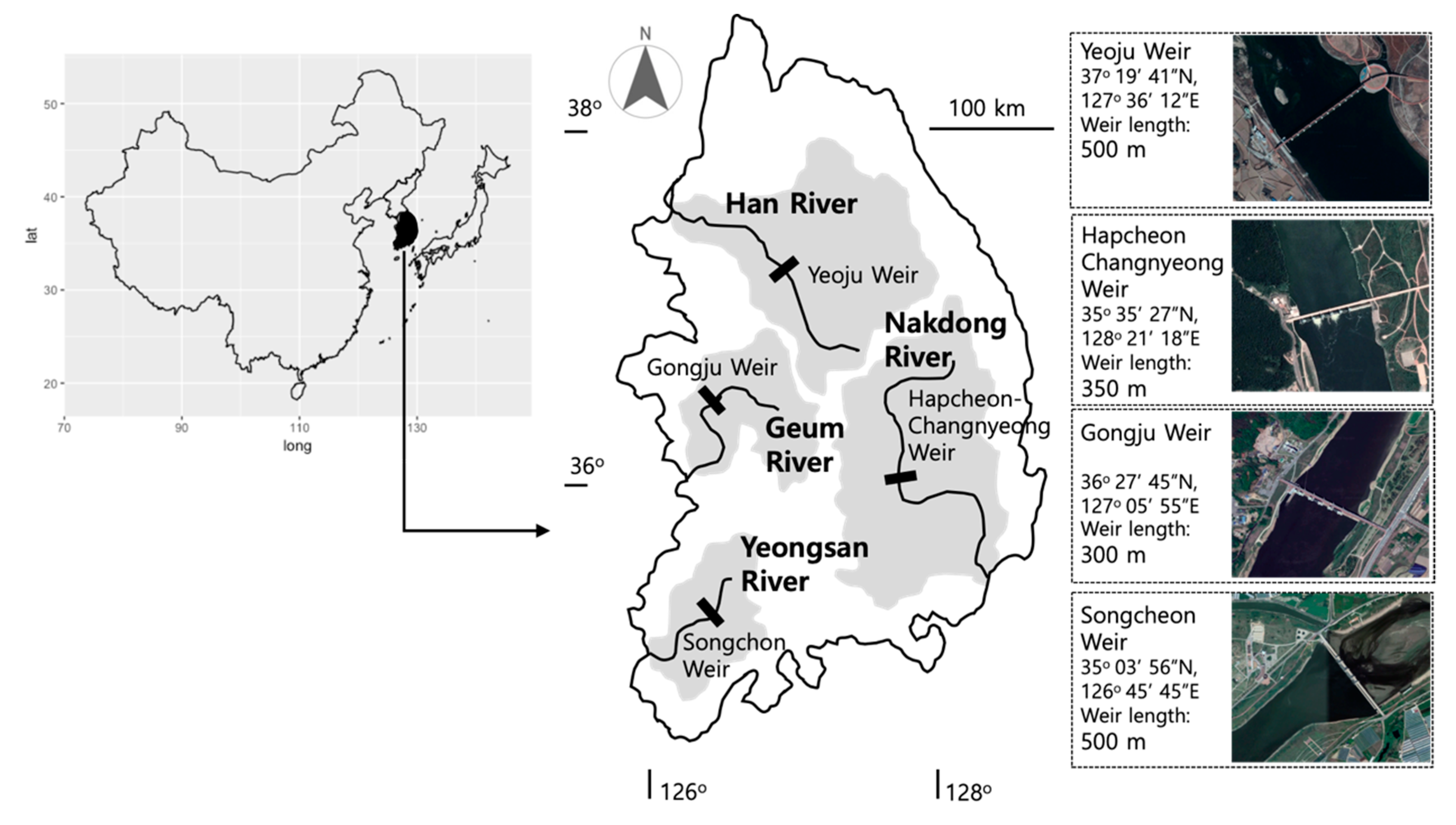
2.1. Study Sites and Fish Sampling
2.2. Community Index and Habitat Index
2.3. Stable Isotope Analysis and Niche Calculations
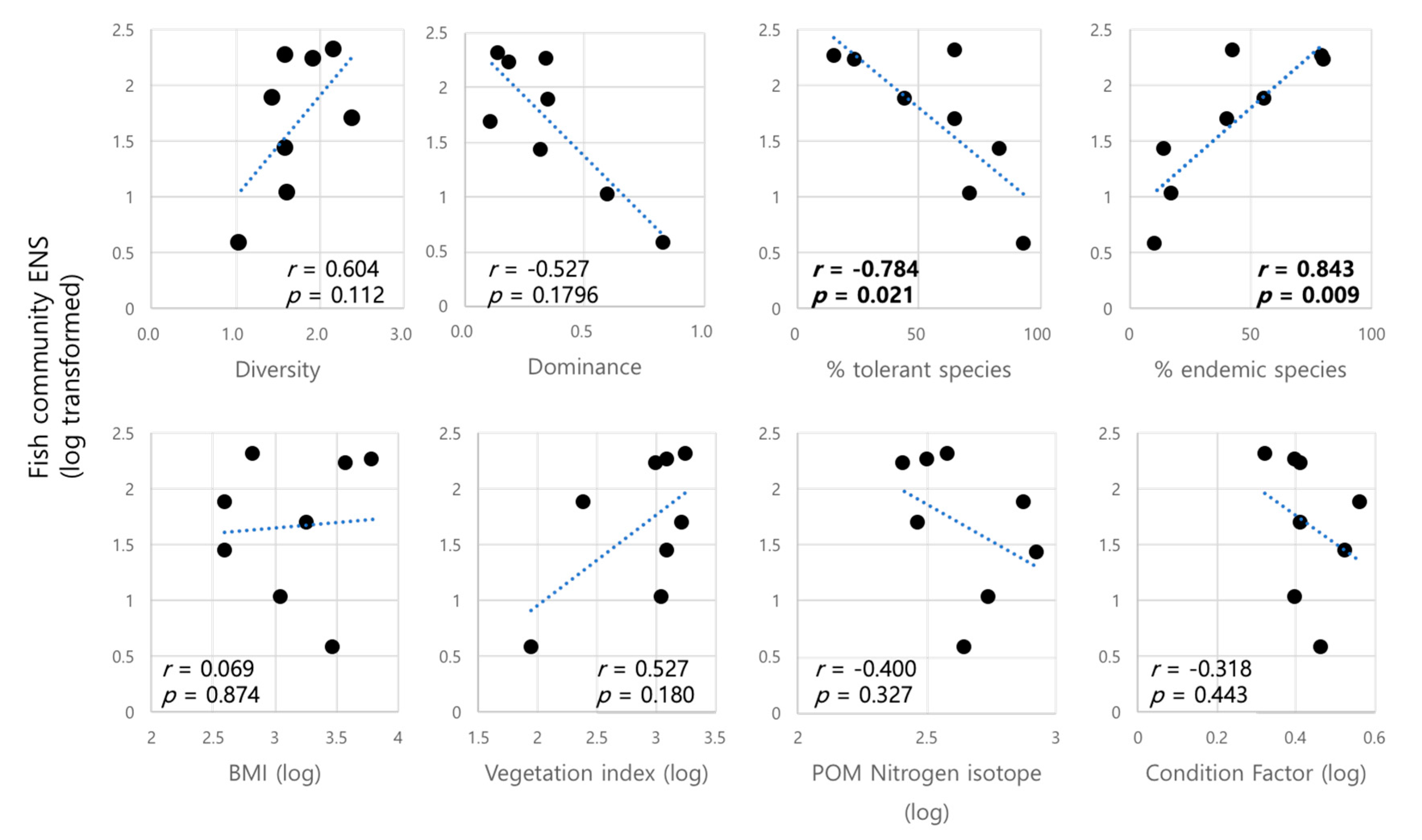
3. Results
3.1. Water Quality
3.2. Fish Community
3.3. Habitat Index
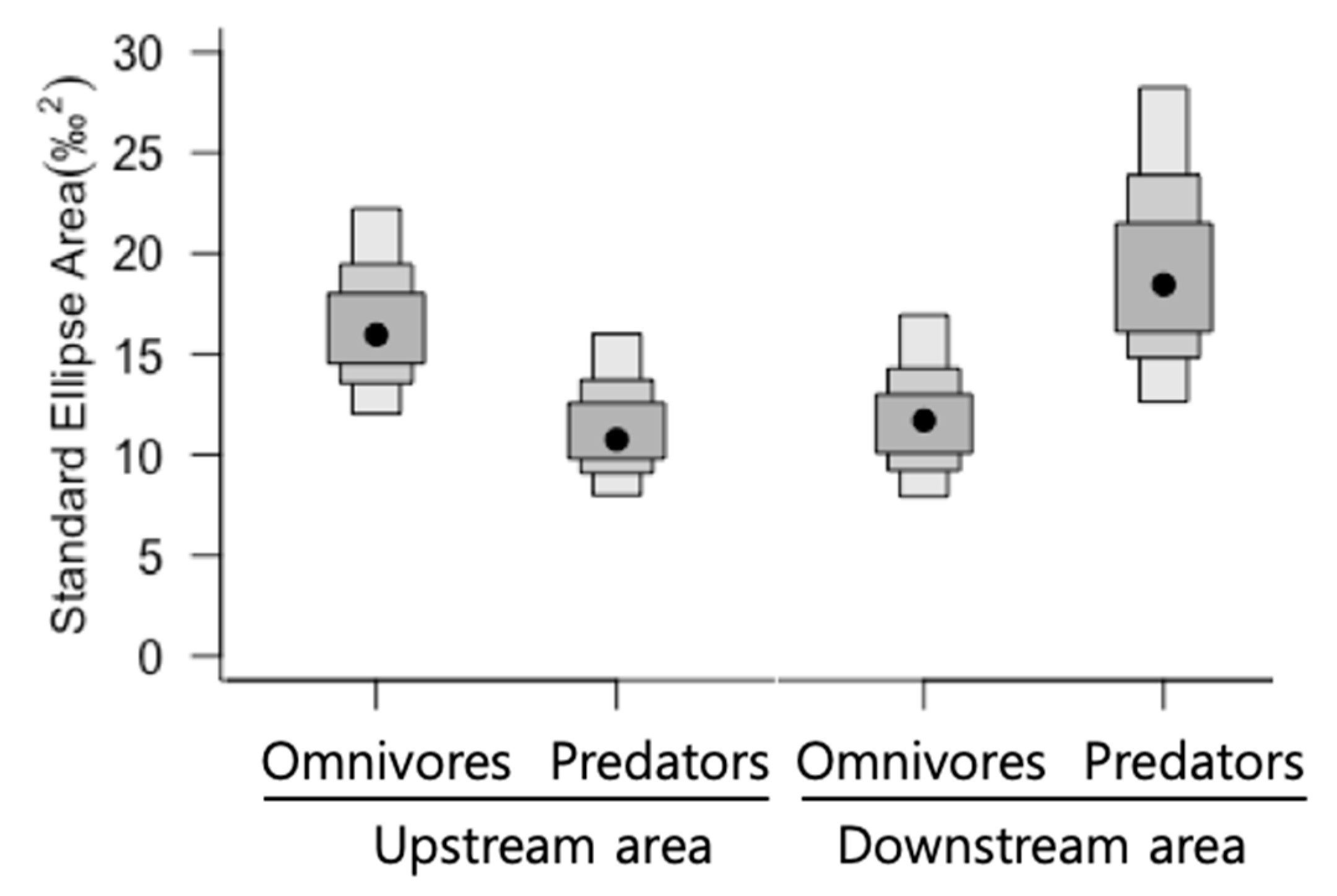
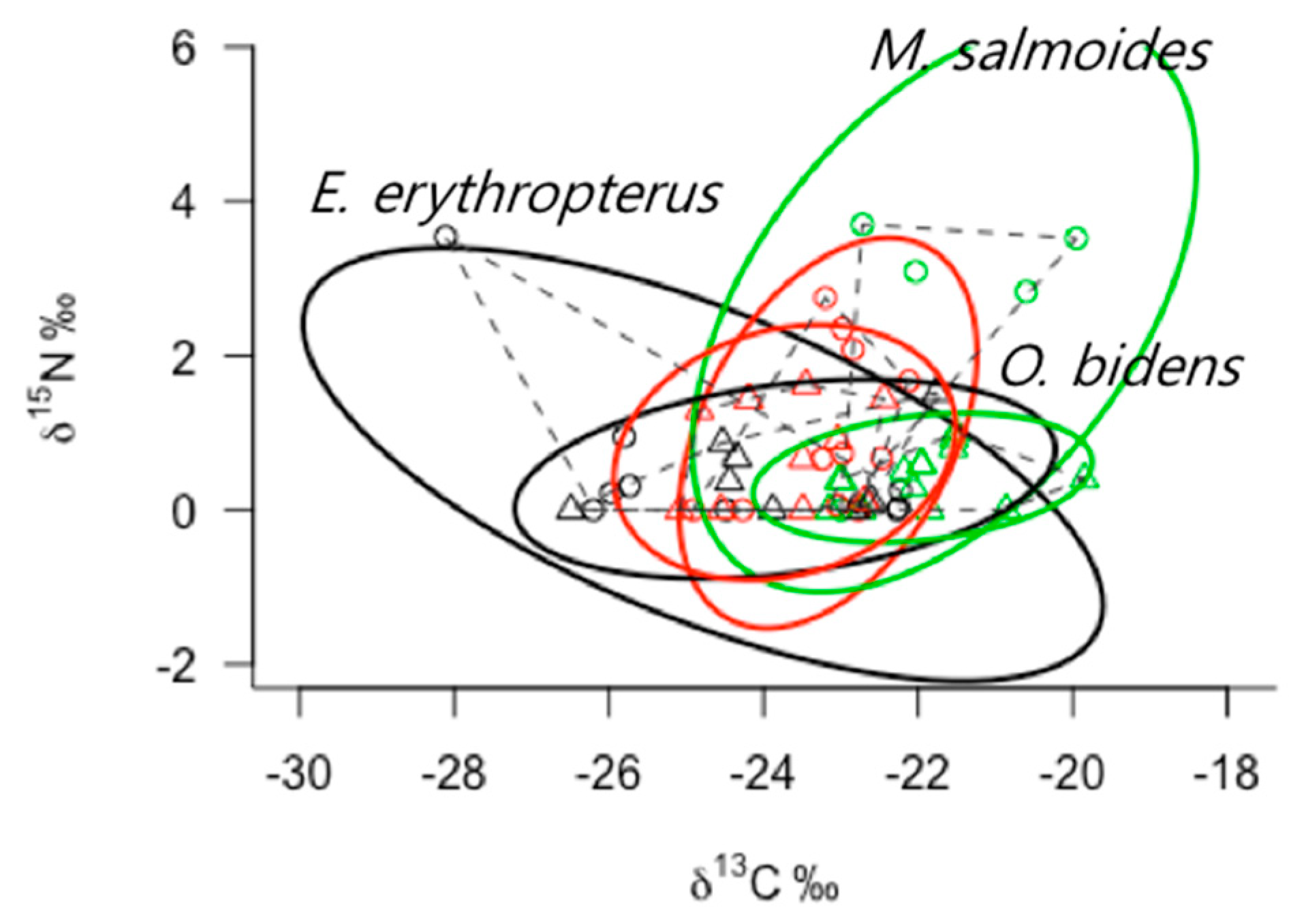
3.4. Ecological Niche Space of Fish and Overlap between Species Different Guild
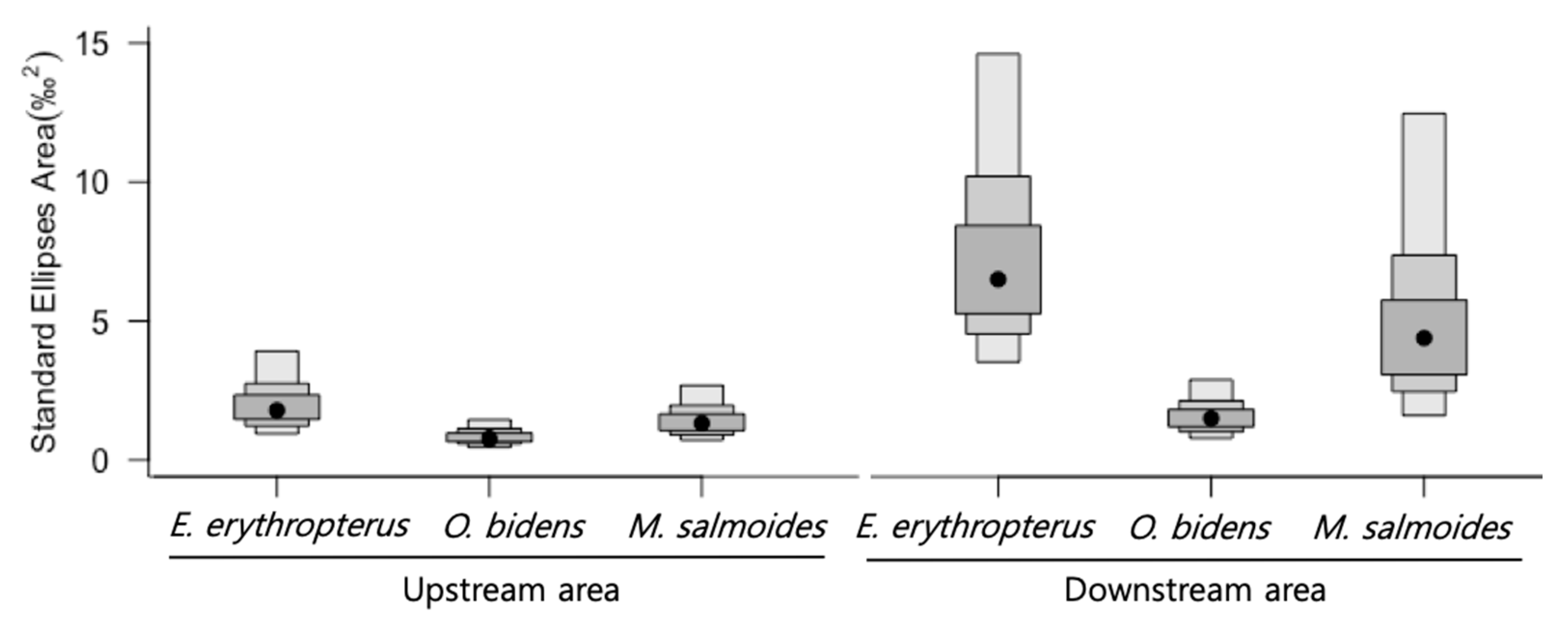
3.5. Comparison of ENSs of Predator Species
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class (Species) | TG 1 | EI 2 | Han River (Yeoju Weir) | Nakdong River (Hapcheon Weir) | Geum River (Gongju Weir) | Yeongsan River (Seungchon Weir) | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| Region | U 3 | D 4 | U | D | U | D | U | D | ||
| Channidae | ||||||||||
| Channa argus | TS | 3 | 1 | 1 | 1 | |||||
| Centrarchidae | ||||||||||
| Micropterus salmoides | TS | I | 9 | 39 | 8 | 68 | 41 | |||
| Lepomis macrochirus | TS | I | 1 | 44 | 2 | 12 | 1 | 233 | 88 | |
| Centropomidae | ||||||||||
| Siniperca scherzeri | IS | 1 | ||||||||
| Odontobutidae | ||||||||||
| Odontobutis interrupta | IS | E | 2 | |||||||
| Bagridae | ||||||||||
| Pseudobagrus koreanus | IS | E | 1 | 1 | ||||||
| Pseudobagrus fulvidraco | TS | 4 | ||||||||
| Gobiidae | ||||||||||
| Rhinogobius brunneus | IS | 1 | 7 | 3 | 4 | |||||
| Rhinogobius giurinus | TS | 1 | ||||||||
| Tridentiger brevispinis | IS | 2 | ||||||||
| Siluridae | ||||||||||
| Silurus asotus | TS | 4 | 1 | 1 | ||||||
| Cobitidae | ||||||||||
| Cobitis rotundicaudata | SS | E | 1 | |||||||
| Misgurnus mizolepis | TS | 3 | ||||||||
| Cyprinidae | ||||||||||
| Abbottina rivularis | TS | 4 | ||||||||
| Acanthorhodeus asmussi | IS | 4 | 18 | 2 | ||||||
| Acanthorhodeus gracilis | IS | E | 191 | 121 | 20 | 5 | ||||
| Acheilognathus lanceolatus | IS | 1 | ||||||||
| Acheilognathus yamatsutae | IS | E | 669 | 80 | ||||||
| Carassius carassius | TS | 34 | 4 | 1 | 3 | 23 | 20 | |||
| Carassius cuvieri | TS | I | 9 | 4 | ||||||
| Cyprinus carpio | TS | I | 1 | 2 | 1 | 4 | ||||
| Erythroculter erythropterus | TS | 4 | 10 | 44 | 4 | 19 | 35 | |||
| Gnathopogon strigatus | IS | 2 | ||||||||
| Hemibarbus labeo | TS | 33 | 27 | 188 | 12 | 1 | 14 | 3 | ||
| Hemiculter eigenmanni | TS | E | 13 | 5 | 16 | 11 | ||||
| Microphysogobio jeoni | IS | E | 1 | 35 | 33 | |||||
| Microphysogobio yaluensis | IS | E | 1 | |||||||
| Opsariichthys uncirostris | TS | 15 | 1 | 406 | 20 | 26 | 36 | 4 | 5 | |
| Paracheilognathus rhombea | IS | 7 | 3 | |||||||
| Pseudogobio esocinus | IS | 2 | 1 | 3 | 1 | 4 | ||||
| Pseudorasbora parva | TS | 12 | 1 | 7 | 1 | 1 | ||||
| Pungtungia herzi | IS | 12 | 4 | |||||||
| Sarcocheilichthys nigripinnis | IS | E | 2 | 61 | 1 | |||||
| Sarcocheilichthys wakiyae | IS | E | 49 | 1 | ||||||
| Squalidus chankaensis tsuchigae | IS | E | 178 | 1 | 51 | 212 | ||||
| Squalidus japonicus Coreanu | TS | E | 59 | 46 | 40 | 42 | ||||
| Squaliobarbus curriculus | IS | 3 | 4 | |||||||
| Zacco platypus | IS | 102 | 15 | 106 | 7 | 19 | 1 | |||
| Total species number | 22 | 17 | 10 | 6 | 18 | 19 | 13 | 15 | ||
| Total individual number | 1211 | 384 | 990 | 29 | 235 | 201 | 446 | 379 | ||
References
- Poff, N.L.; Allan, J.D.; Bain, M.B.; Karr, J.R.; Prestegaard, K.L.; Richter, B.D.; Sparks, R.E.; Stromberg, J.C. The natural flow paradigm. BioScience 1997, 47, 769–784. [Google Scholar] [CrossRef]
- Bain, M.B.; Finn, J.T.; Booke, H.E. Streamflow regulation and fish community structure. Ecology 1988, 69, 382–392. [Google Scholar] [CrossRef]
- Musil, J.; Horky, P.; Slavik, O.; Zboril, A.; Horka, P. The response of the young of the year fish to river obstacles: Functional and numerical linkages between dams, weirs, fish habitat guilds and biotic integrity across large spatial scale. Ecol. Indic. 2012, 23, 634–640. [Google Scholar] [CrossRef]
- Amaral, S.D.; Branco, P.; da Silva, A.T.; Datopodis, C.; Viseu, T.; Ferreira, M.T.; Pinheiro, A.N.; Santos, J.M. Upstream passage of potamodromous cyprinids over small weirs: The influence of key-hydraulic parameters. J. Ecohydraul. 2016, 1, 79–89. [Google Scholar] [CrossRef]
- Vermaat, J.E.; Wagtendok, A.J.; Brouwer, R.; Sheremet, O.; Ansink, E.; Brockhoff, T.; Plug, M.; Hellsten, S.; Aroviita, J.; Tylec, L.; et al. Assessing the sociental benefits of river restoration using the ecosystem services approach. Hydrobiologia 2015, 769, 121–135. [Google Scholar] [CrossRef]
- Doi, H.; Katano, I.; Negishi, J.N.; Sanada, S.; Kayaba, Y. Effects of biodiversity, habitat structure, and water quality on recreational use of rivers. Ecophere 2013, 4, 102. [Google Scholar] [CrossRef]
- Ward, J.V.; Stanford, J.A. The Ecology of regulated streams: Past accomplishment and directions for future research. In Regulated Streams; Craig, J.F., Kemper, J.B., Eds.; Springer: Boston, MA, USA, 1987; pp. 391–409. ISBN 978-1-4684-5394-2. [Google Scholar]
- Branco, P.; Amaral, S.D.; Ferreira, M.T.; Santos, J.M. Do small barriers affect the movement of freshwater fish by increasing residency? Sci. Total Environ. 2017, 581–582, 486–494. [Google Scholar] [CrossRef] [PubMed]
- Poulet, N. Impact of weirs on fish communities in a Piedmont Stream. River Res. Appl. 2007, 23, 1038–1047. [Google Scholar] [CrossRef]
- Amaral, S.D.; Branco, P.; Romao, F.; Viseu, T.; Ferreira, M.T.; Pinheiro, A.N.; Santos, J.M. The effect of weir crest width and discharge on passage performance of a potamodromous cycprinid. Mar. Freshw. Res. 2018. [Google Scholar] [CrossRef]
- Lah, T.J.; Park, Y.; Cho, Y.J. The four major rivers restoration project of South Korea: An assessment of its process, program, and political dimensions. J. Environ. Dev. 2015, 24, 375–394. [Google Scholar] [CrossRef]
- Ahn, J.M.; Lee, S.; Kang, T. Evaluation of dams and weirs operating for water resource management of the Geum River. Sci. Total Environ. 2014, 478, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Han, J.H.; Lim, B.J.; Park, J.H.; Shin, J.K.; An, K.G. Comparative analysis of fish fauna and community structures before and after the artificial weir construction in the mainstreams and tributaries of Yeongsan River watershed. Korean J. Ecol. Environ. 2013, 46, 103–115. [Google Scholar] [CrossRef]
- Cha, S.M.; Kang, M.J.; Park, Y.; Lee, S.W.; Kim, J.H. Water quality changes according to the midstream weir construction in the Yeongsan River, Korea. Desalin. Water Treat. 2014, 53, 3066–3071. [Google Scholar] [CrossRef]
- Kim, J.H.; Yoon, J.D.; Baek, S.H.; Park, S.H.; Lee, J.W.; Lee, J.A.; Jang, M.H. An efficiency analysis of a nature-like fishway for freshwater fish ascending a large Korean river. Water 2016, 8, 3. [Google Scholar] [CrossRef]
- Wang, J.; Chapman, D.; Xu, J.; Wang, Y.; Gu, B. Isotope niche dimension and trophic overlap between bigheaded carps and native fliter-feeding fish in the lower Missouri River, USA. PLoS ONE 2018, 13, e0197584. [Google Scholar] [CrossRef]
- Cohen, J.E. Food webs and the dimensionality of trophic niche space. Proc. Natl. Acad. Sci. USA 1977, 74, 4533–4536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newsome, S.D.; Martinez del Rio, C.; Bearhop, S.; Phillips, D.L. A niche for isotopic ecology. Front. Ecol. Environ. 2007, 5, 429–436. [Google Scholar] [CrossRef]
- Jackson, A.L.; Inger, R.; Parnell, A.C.; Bearhop, S. Comparing isotopic niche widths among and within communities: SIBER-stable isotope Bayesian ellipses in R. J. Anim. Ecol. 2011, 80, 595–602. [Google Scholar] [CrossRef]
- Ministry of Environment, Korea. Survey and Evaluation Method for River and Stream Ecosystem Health Assessment. In Biomonitoring Survey and Assessment Manual; NIER/2016-372; Ministry of Environment: Sejong, Korea, 2016. [Google Scholar]
- Pielou, E.C. Species-diversity and pattern-diversity in the study of ecological succession. J. Theor. Biol. 1966, 10, 370–383. [Google Scholar] [CrossRef]
- Simpson, E.H. Measurement of diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Kong, D.; Son, S.H.; Hwang, S.J.; Won, D.H.; Kim, M.C.; Park, J.H.; Jeon, T.S.; Lee, J.E.; Kim, J.H.; Kim, J.S.; et al. Development of benthic macroinvertebrates index (BMI) for biological assessment on stream environment. J. Korean Soc. Water Environ. 2018, 34, 183–201. [Google Scholar] [CrossRef]
- Ministry of Environment, Korea. The Improvement of Mid-Long Term Aquatic Ecosystem Monitoring on Weir Section; NIER-SP2015-189; Ministry of Environment: Se-jong, Korea, 2015; ISBN 11-1480523-002465-01.
- Barbour, M.T.; Gerritsen, J.; Snyder, B.D.; Stribling, J.B. Rapid Bioassessment Protocols for Use in Streams and Wadeable Rivers: Periphyton, Benthic Macroinvertebrates and Fish, 2nd ed.; EPA/841-B-99-002; US Environmental Protection Agency: Washington, DC, USA, 1999.
- Bannon, R.O.; Roman, C.T. Using stable isotopes to monitor anthropogenic nitrogen inputs to estuaries. Ecol. Appl. 2008, 18, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Froese, R. Cube law, condition factor and weight-length relationships: History, meta-analysis and recommendations. J. Appl. Ichthyol. 2006, 22, 241–253. [Google Scholar] [CrossRef]
- Bolger, T.; Connolly, P.L. The selection of suitable indices for the measurement and analysis of fish condition. J. Fish Biol. 1989, 34, 171–182. [Google Scholar] [CrossRef]
- Lee, W.S.; Kim, M.S.; Hwang, J.Y.; Kwon, O.S. Analytical methodology of stable isotopes ratios: Sample pretreatment, analysis and application. Korean J. Ecol. Environ. 2013, 46, 471–487. [Google Scholar] [CrossRef]
- Layman, C.A.; Arrington, D.A.; Montana, C.G.; Post, D.M. Can stable isotope ratios provide for community-wide measures of trophic structure? Ecology 2007, 88, 42–48. [Google Scholar] [CrossRef]
- Coulter, A.A.; Swanson, H.K.; Goforth, R.R. Seasonal variation in resource overlap of invasive and native fishes revealed by stable isotopes. Biol. Invasions 2018. [Google Scholar] [CrossRef]
- Cabana, G.; Rasmussen, J.B. Comparison of aquatic food chains using nitrogen isotopes. Proc. Natl. Acad. Sci. USA 1996, 93, 10844–10847. [Google Scholar] [CrossRef]
- Tummers, J.S.; Hudson, S.; Lucas, M.C. Evaluating the effectiveness of restoring longitudinal connectivity for stream fish communities: Towards a more holistic approach. Sci. Total Environ. 2016, 569–570, 850–860. [Google Scholar] [CrossRef]
- Mbaka, J.G.; Mwaniki, M.W. A global review of the downstream effects of small impoundments on stream habitat conditions and macroinvertebrates. Environ. Rev. 2015, 23, 257–262. [Google Scholar] [CrossRef]
- Smith, S.C.F.; Meiners, S.J.; Hastings, R.P.; Thomas, T.; Colombo, R.E. Low-head dam impacts on habitat and the functional composition of fish communities. River Res. Appl. 2017, 33, 680–689. [Google Scholar] [CrossRef]
- France, R.L. Differentiation between littoral and pelagic food webs in lakes using stable carbon isotopes. Limnol. Oceanogr. 1995, 40, 1310–1313. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.J.; Lee, K.L.; Lee, H.J.; Hwang, J.W.; Lyu, H.S.; Shin, L.Y.; Park, A.R.; Chen, S.U. Relations of nutrient concentrations on the seasonality of algal community in the Nakdong River, Korea. J. Korean Soc. Water Environ. 2015, 31, 110–119. [Google Scholar] [CrossRef]
- Chang, K.H.; Doi, H.; Imai, H.; Gunji, H.; Nakano, S. Longitudinal changes in zooplankton distribution below a reservoir outfall with reference to river planktivory. Limnology 2008, 9, 125–133. [Google Scholar] [CrossRef]
- Jo, H.; Yoon, J.D.; Kim, J.H.; Jeong, K.S.; Do, Y.; Joo, G.J. Diet shifts and delayed piscivory specialization during the ontegenesis of the largemouth bass Micropterus salmoides (Lacepede, 1802) in the Nakdong River and Upo Wetlands, South Korea. Indian J. Fish. 2016, 63, 48–54. [Google Scholar] [CrossRef]
- Doi, H.; Chang, K.H.; Ando, T.; Imai, H.; Nakano, S.; Kajimoto, A.; Katano, I. Drifting plankton from a reservoir subsidize downstream food webs and alter community structure. Oecologia 2008, 156, 363–371. [Google Scholar] [CrossRef]
- Lombard, R.J.; Chimimba, T.; Zengeya, T.A. Niche complementary between an alien predator and native omnivorous fish in the Wilge River, South Africa. Hydrobiologia 2018, 817, 329–340. [Google Scholar] [CrossRef]
- Yoon, J.D.; Park, S.H.; Chang, K.H.; Choi, J.Y.; Joo, G.J.; Nam, G.S.; Yoon, J.; Jang, M.H. Characteristics of fish fauna in the lower Geum River and identification of trophic guilds using stable isotopes analysis. Korean J. Environ. Biol. 2015, 33, 34–44. [Google Scholar] [CrossRef]






| River (Weir) | Region | Predators | Omnivores |
|---|---|---|---|
| Han River (Yeoju Weir) | upstream | Erythroculter erythropterus (3) Micropterus salmoides (3) | Acheilognathus yamatsutae (3) 1 Acanthorhodeus gracilis (3) Pseudogobio esocinus (3) |
| downstream | Erythroculter erythropterus (3) | Acheilognathus yamatsutae (3) Acanthorhodeus gracilis (3) 1 | |
| Nakdong River (Hapcheon Weir) | upstream | Erythroculter erythropterus (3) Micropterus salmoides (3) Opsariichthys bidens (3) 1 | Lepomis macrochirus (3) Squalidus chankaensis tsuchigae (3) Pseudogobio esocinus (3) |
| downstream | Erythroculter erythropterus (3) Opsariichthys bidens (3)1 | Lepomis macrochirus (3) Squalidus chankaensis tsuchigae (3) Hemiculter eigenmanni (2) | |
| Geum River (Gongju Weir) | upstream | Erythroculter erythropterus (3) Micropterus salmoides (3) Opsariichthys bidens (3) | Squalidus japonicus Coreanus(3)1 Microphysogobio jeoni (3) Pseudogobio esocinus (3) |
| downstream | Erythroculter erythropterus (3) Opsariichthys bidens (3) | Squalidus japonicus Coreanus (3) 1 | |
| Yeongsan River (Seungchon Weir) | upstream | Micropterus salmoides (5) Opsariichthys bidens (5) | Squalidus chankaensis tsuchigae (5) Lepomis macrochirus (5) 1 Pseudogobio esocinus (5) |
| downstream | Micropterus salmoides (5) Opsariichthys bidens (5) | Squalidus chankaensis tsuchigae (5) 1 Lepomis macrochirus (5) Pseudogobio esocinus (5) |
| River (Weir) | Region | Number of Captured Individuals | Diversity Index | Dominance Index | Tolerant Species % | Endemic Species % |
|---|---|---|---|---|---|---|
| Han River (Yeoju Weir) | upstream | 1211 | 1.62 | 0.34 | 14.9 | 80.3 |
| downstream | 384 | 1.95 | 0.19 | 23.7 | 80.5 | |
| Nakdong River (Hapcheon Weir) | upstream | 990 | 2.40 | 0.11 | 65.1 | 40.9 |
| downstream | 29 | 2.19 | 0.14 | 64.7 | 43.3 | |
| Geum River (Gongju Weir) | upstream | 235 | 1.62 | 0.60 | 71.3 | 17.8 |
| downstream | 201 | 1.06 | 0.83 | 93.1 | 11.1 | |
| Yeongsan River (Seungchon Weir) | upstream | 446 | 1.62 | 0.32 | 83.6 | 15.0 |
| downstream | 397 | 1.46 | 0.35 | 44.6 | 56.4 |
| River (weir) | Region | BMI | Vegetation Index | Delta 15N POM | CF |
|---|---|---|---|---|---|
| Han River (Yeoju Weir) | upstream | 43.2 | 21 | 11.23 | 0.49 |
| downstream | 34.6 | 19 | 10.16 | 0.51 | |
| Nakdong River (Hapcheon Weir) | upstream | 25.2 | 24 | 10.77 | 0.51 |
| downstream | 15.8 | 25 | 12.20 | 0.38 | |
| Geum River (Gongju Weir) | upstream | 20.1 | 20 | 14.50 | 0.49 |
| downstream | 31.0 | 6 | 13.08 | 0.59 | |
| Yeongsan River (Seungchon Weir) | upstream | 12.5 | 21 | 17.70 | 0.69 |
| downstream | 12.4 | 10 | 16.81 | 0.76 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hur, J.W.; Jang, M.-H.; Shin, K.-H.; Lee, K.-L.; Chang, K.-H. Ecological Niche Space of Fish Communities in Impounded Sections of Large Rivers: Its Application to Assessment of the Impact of Weirs on River Ecosystems. Sustainability 2018, 10, 4784. https://doi.org/10.3390/su10124784
Hur JW, Jang M-H, Shin K-H, Lee K-L, Chang K-H. Ecological Niche Space of Fish Communities in Impounded Sections of Large Rivers: Its Application to Assessment of the Impact of Weirs on River Ecosystems. Sustainability. 2018; 10(12):4784. https://doi.org/10.3390/su10124784
Chicago/Turabian StyleHur, Jun Wook, Min-Ho Jang, Kyung-Hoon Shin, Kyung-Lak Lee, and Kwang-Hyeon Chang. 2018. "Ecological Niche Space of Fish Communities in Impounded Sections of Large Rivers: Its Application to Assessment of the Impact of Weirs on River Ecosystems" Sustainability 10, no. 12: 4784. https://doi.org/10.3390/su10124784
APA StyleHur, J. W., Jang, M. -H., Shin, K. -H., Lee, K. -L., & Chang, K. -H. (2018). Ecological Niche Space of Fish Communities in Impounded Sections of Large Rivers: Its Application to Assessment of the Impact of Weirs on River Ecosystems. Sustainability, 10(12), 4784. https://doi.org/10.3390/su10124784