Comparative Analysis of Perennial and Annual Phaseolus Seed Nutrient Concentrations

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study System
2.2. Sample Preparation and Analysis
2.3. Statistical Analysis
3. Results
3.1. Differences among Perennials and Annuals in Ion and Amino Acid Concentrations
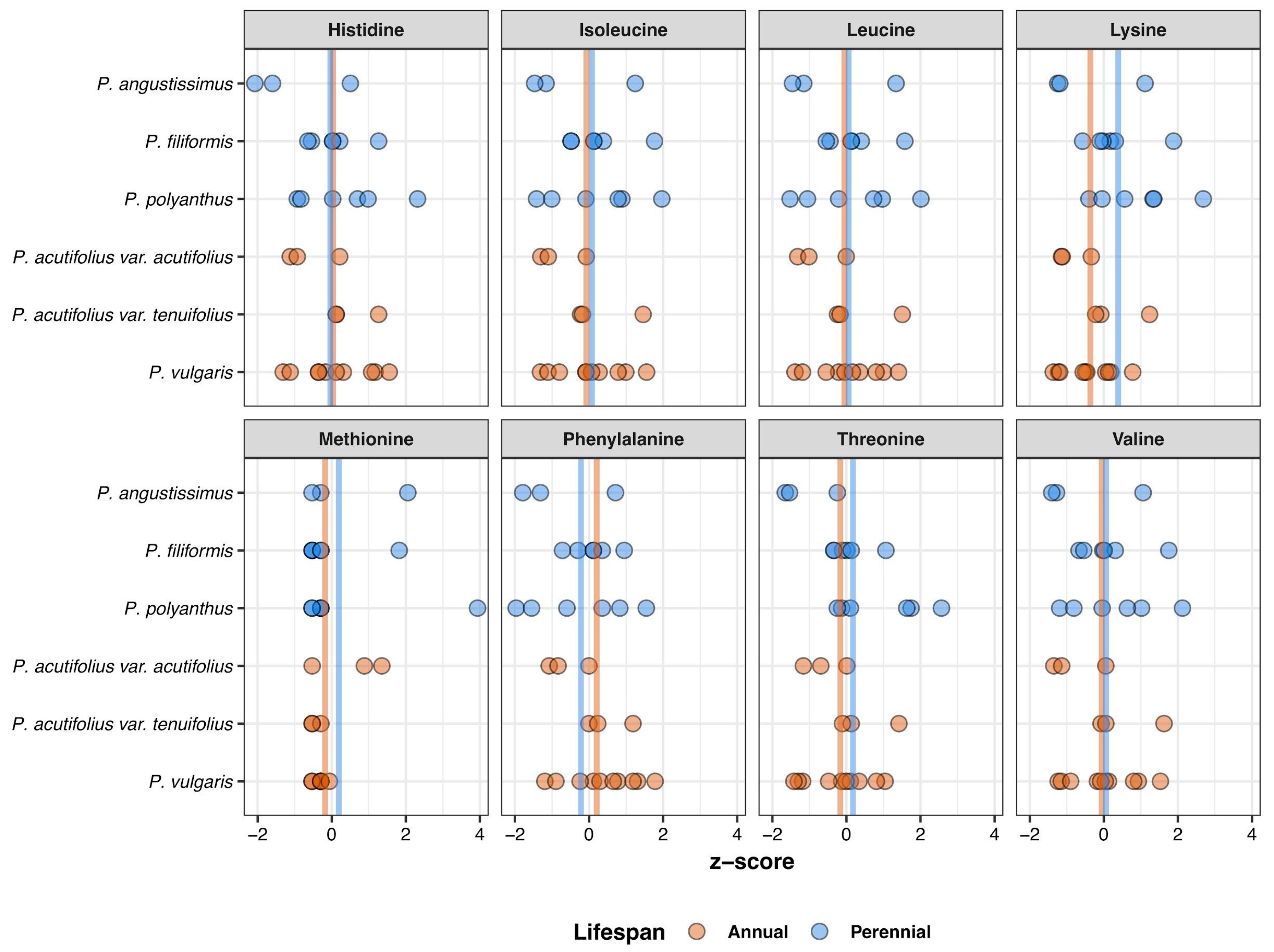
3.2. Variation within Species in Ion and Amino Acid Concentrations
3.3. Correlations among Ions and Amino Acids
4. Discussion
4.1. Comparisons of Ion and Amino Acid Concentrations in Annual and Perennial Phaseolus
4.2. Natural Variation in Mineral and Amino Acid Concentrations within Six Wild Phaseolus Species
4.3. Correlations in Mineral and Amino Acid Concentrations across Phaseolus Species
4.4. Future Directions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Global Nutrition Targets 2025: Policy Brief Series; World Health Organization: Geneva, Switzerland, 2012. [Google Scholar]
- Welch, R.M.; Graham, R.D. Breeding crops for enhanced micronutrient content. In Food Security in Nutrient-Stressed Environments: Exploiting Plants’ Genetic Capabilities; Adu-Gyamfi, J.J., Ed.; Springer: Dordrecht, The Netherlands, 2002; pp. 267–276. ISBN 9789401715706. [Google Scholar]
- Black, R.E.; Victora, C.G.; Walker, S.P.; Bhutta, Z.A.; Christian, P.; de Onis, M.; Ezzati, M.; Grantham-McGregor, S.; Katz, J.; Martorell, R.; et al. Maternal and child undernutrition and overweight in low-income and middle-income countries. Lancet 2013, 382, 427–451. [Google Scholar] [CrossRef]
- Smith, M.R.; Golden, C.D.; Myers, S.S. Potential rise in iron deficiency due to future anthropogenic carbon dioxide emissions. GeoHealth 2017, 1, 248–257. [Google Scholar] [CrossRef] [Green Version]
- United States Department of Agriculture. USDA Agroforestry Fact Sheet; United States Department of Agriculture: Washington, DC, USA, 2011.
- Frison, E.A.; Cherfas, J.; Hodgkin, T. Agricultural biodiversity is essential for a sustainable improvement in food and nutrition security. Sustain. Sci. Pract. Policy 2011, 3, 238–253. [Google Scholar] [CrossRef]
- Jamnadass, R.H.; Place, F.; Torquebiau, E.F.; Malezieux, E.; Iiyama, M.; Sileshi, G.W.; Kehlenbeck, K.; Masters, E.; Mcmullin, S.; Weber, J.C.; et al. Agroforestry, Food And Nutritional Security; ICRAF: Working Paper, No. 170; World Agroforestry Centre: Nairobi, Kenya, 2013. [Google Scholar] [CrossRef]
- Callo-Concha, D.; Jemal, O.M.; Aragaw, H.S. Local alternatives to local problems: The contribution of agroforestry system by-products to food and nutrition security of communities in southwestern Ethiopia. Food Stud. Interdiscip. J. 2019, 9, 29–42. [Google Scholar] [CrossRef]
- Kane, D.A.; Rogé, P.; Snapp, S.S. A systematic review of perennial staple crops literature using topic modeling and bibliometric analysis. PLoS ONE 2016, 11, e0155788. [Google Scholar] [CrossRef]
- Crews, T.E.; Cattani, D.J. Strategies, advances, and challenges in breeding perennial grain crops. Sustain. Sci. Pract. Policy 2018, 10, 2192. [Google Scholar] [CrossRef]
- DeHaan, L.R.; Van Tassel, D.L.; Cox, T.S. Perennial grain crops: A synthesis of ecology and plant breeding. Renew. Agric. Food Syst. 2005, 20, 5–14. [Google Scholar] [CrossRef]
- Pimentel, D.; Cerasale, D.; Stanley, R.C.; Perlman, R.; Newman, E.M.; Brent, L.C.; Mullan, A.; Chang, D.T.-I. Annual vs. perennial grain production. Agric. Ecosyst. Environ. 2012, 161, 1–9. [Google Scholar] [CrossRef]
- Cox, T.S.; Glover, J.D.; Van Tassel, D.L.; Cox, C.M.; DeHaan, L.R. Prospects for developing perennial grain crops. Bioscience 2006, 56, 649–659. [Google Scholar] [CrossRef]
- Ciotir, C.; Applequist, W.; Crews, T.E.; Cristea, N.; DeHaan, L.R.; Frawley, E.; Herron, S.A.; Magill, R.; Miller, J.S.; Roskov, Y.; et al. Building a botanical foundation for perennial agriculture: Global inventory of wild, perennial herbaceous Fabaceae species. bioRxiv 2019. bioRxiv: 515189. [Google Scholar]
- Mossé, J.; Baudet, J. Crude protein content and amino acid composition of seeds: variability and correlations. Plant Foods Hum. Nutr. 1983, 32, 225–245. [Google Scholar] [CrossRef]
- Beebe, S.; Gonzalez, A.V.; Rengifo, J. Research on trace minerals in the common bean. Food Nutr. Bull. 2000, 21, 387–391. [Google Scholar] [CrossRef]
- González, A.M.; Monteagudo, A.B.; Casquero, P.A.; De Ron, A.M.; Santalla, M. Genetic variation and environmental effects on agronomical and commercial quality traits in the main European market classes of dry bean. Field Crops Res. 2006, 95, 336–347. [Google Scholar] [CrossRef]
- Bonfil, D.J.; Goren, O.; Mufradi, I.; Lichtenzveig, J.; Abbo, S. Development of early-flowering Kabuli chickpea with compound and simple leaves. Plant Breed. 2007, 126, 125–129. [Google Scholar] [CrossRef] [Green Version]
- Florez, A.; Pujolà, M.; Valero, J.; Centelles, E.; Almirall, A.; Casañas, F. Genetic and environmental effects on chemical composition related to sensory traits in common beans (Phaseolus vulgaris L.). Food Chem. 2009, 113, 950–956. [Google Scholar] [CrossRef]
- Buescher, E.; Achberger, T.; Amusan, I.; Giannini, A.; Ochsenfeld, C.; Rus, A.; Lahner, B.; Hoekenga, O.; Yakubova, E.; Harper, J.F.; et al. Natural genetic variation in selected populations of Arabidopsis thaliana is associated with ionomic differences. PLoS ONE 2010, 5, e11081. [Google Scholar] [CrossRef]
- Pinson, S.R.M.; Tarpley, L.; Yan, W.; Yeater, K.; Lahner, B.; Yakubova, E.; Huang, X.-Y.; Zhang, M.; Guerinot, M.L.; Salt, D.E. Worldwide genetic diversity for mineral element concentrations in rice grain. Crop Sci. 2015, 55, 294–311. [Google Scholar] [CrossRef]
- Migicovsky, Z.; Harris, Z.N.; Klein, L.L.; Li, M.; McDermaid, A.; Chitwood, D.H.; Fennell, A.; Kovacs, L.G.; Kwasniewski, M.; Londo, J.P.; et al. Rootstock effects on scion phenotypes in a “Chambourcin” experimental vineyard. Hortic. Res. 2019, 6, 64. [Google Scholar] [CrossRef] [PubMed]
- Huber, S.C.; Li, K.; Nelson, R.; Ulanov, A.; DeMuro, C.M.; Baxter, I. Canopy position has a profound effect on soybean seed composition. PeerJ 2016, 4, e2452. [Google Scholar] [CrossRef]
- Kell, D.B. Breeding crop plants with deep roots: their role in sustainable carbon, nutrient and water sequestration. Ann. Bot. 2011, 108, 407–418. [Google Scholar] [CrossRef] [Green Version]
- Pierret, A.; Maeght, J.-L.; Clément, C.; Montoroi, J.-P.; Hartmann, C.; Gonkhamdee, S. Understanding deep roots and their functions in ecosystems: An advocacy for more unconventional research. Ann. Bot. 2016, 118, 621–635. [Google Scholar] [CrossRef]
- Murphy, K.M.; Hoagland, L.A.; Reeves, P.G.; Baik, B.-K.; Jones, S.S. Nutritional and quality characteristics expressed in 31 perennial wheat breeding lines. Renew. Agric. Food Syst. 2009, 24, 285–292. [Google Scholar] [CrossRef]
- Rendón-Anaya, M.; Montero-Vargas, J.M.; Saburido-Álvarez, S.; Vlasova, A.; Capella-Gutierrez, S.; Ordaz-Ortiz, J.J.; Aguilar, O.M.; Vianello-Brondani, R.P.; Santalla, M.; Delaye, L.; et al. Genomic history of the origin and domestication of common bean unveils its closest sister species. Genome Biol. 2017, 18, 60. [Google Scholar] [CrossRef]
- Welsh, W.; Bushuk, W.; Roca, W.; Singh, S.P. Characterization of agronomic traits and markers of recombinant inbred lines from intra- and interracial populations of Phaseolus vulgaris L. Theor. Appl. Genet. 1995, 91, 169–177. [Google Scholar] [CrossRef]
- Broughton, W.J.; Hernández, G.; Blair, M.; Beebe, S.; Gepts, P.; Vanderleyden, J. Beans (Phaseolus spp.)—Model food legumes. Plant Soil 2003, 252, 55–128. [Google Scholar] [CrossRef]
- Graham, P.H.; Vance, C.P. Legumes: Importance and constraints to greater use. Plant Physiol. 2003, 131, 872–877. [Google Scholar] [CrossRef]
- Statistical Yearbook of the Food And Agricultural Organization for the United Nations. FAOSTAT: Rome, Italy, 2013. Available online: http://www.fao.org/3/i3107e/i3107e.pdf (accessed on 14 May 2019).
- Porch, T.G.; Beaver, J.S.; Debouck, D.G.; Jackson, S.A.; Kelly, J.D.; Dempewolf, H. Use of wild relatives and closely related species to adapt common bean to climate change. Agronomy 2013, 3, 433–461. [Google Scholar] [CrossRef]
- Ramirez-Cabral, N.Y.Z.; Kumar, L.; Taylor, S. Crop niche modeling projects major shifts in common bean growing areas. Agric. For. Meteorol. 2016, 218–219, 102–113. [Google Scholar] [CrossRef]
- Martínez, J.P.; Silva, H.; Ledent, J.F.; Pinto, M. Effect of drought stress on the osmotic adjustment, cell wall elasticity and cell volume of six cultivars of common beans (Phaseolus vulgaris L.). Eur. J. Agron. 2007, 26, 30–38. [Google Scholar] [CrossRef]
- Dohle, S.; Berny, J.C.; Khoury, C.K.; Kisha, T.; Egan, A.N. Wild beans (Phaseolus L.) of North America. In North American Crop Wild Relatives, Volume 2: Important Species; Greene, S.L., Williams, K.A., Khoury, C.K., Kantar, M.B., Marek, L.F., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 99–127. ISBN 9783319971216. [Google Scholar]
- Nabhan, G.P.; Felger, R.S. Wild desert relatives of crops: Their direct uses as food. In Plants for Arid Lands; Wickens, G.E., Field, D.V., Goodin, J.R., Eds.; Springer: Cham, The Netherlands, 1985; pp. 19–33. ISBN 9780044453307. [Google Scholar]
- Arrifin, R. Proximate Analysis of Sonoran Desert Food Plants; The University of Arizona: Tucson, AZ, USA, 1984. [Google Scholar]
- Gepts, P.; Debouck, D. Origin, domestication, and evolution of the common bean (Phaseolus vulgaris L.). Common Beans Res. Crop Improv. 1991, 7, 53. [Google Scholar]
- Ziegler, G.; Terauchi, A.; Becker, A.; Armstrong, P.; Hudson, K.; Baxter, I. Ionomic screening of field-grown soybean identifies mutants with altered seed elemental composition. Plant Genome 2013, 6, 1–9. [Google Scholar] [CrossRef]
- Baxter, I. Ionomics: The functional genomics of elements. Brief. Funct. Genom. 2010, 9, 149–156. [Google Scholar] [CrossRef] [Green Version]
- Hacham, Y.; Avraham, T.; Amir, R. The N-terminal region of Arabidopsis cystathionine gamma-synthase plays an important regulatory role in methionine metabolism. Plant Physiol. 2002, 128, 454–462. [Google Scholar] [CrossRef]
- Wei, T.; Simko, V. R Package “Corrplot”: Visualization of a Correlation Matrix. Available online: https://github. com/taiyun/corrplot (accessed on 17 October 2017).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: Cham, Switzerland, 2016; ISBN 9783319242774. [Google Scholar]
- SAS Institute. Base SAS 9.4 Procedures Guide: Statistical Procedures, 5th ed.; SAS Institute: Cary, NC, USA, 2017; ISBN 9781635260205. [Google Scholar]
- Velu, G.; Crespo Herrera, L.; Guzman, C.; Huerta, J.; Payne, T.; Singh, R.P. Assessing genetic diversity to breed competitive biofortified wheat with enhanced grain Zn and Fe concentrations. Front. Plant Sci. 2018, 9, 1971. [Google Scholar] [CrossRef] [PubMed]
- Baldi, G.; Salamini, F. Variability of essential amino acid content in seeds of 22 Phaseolus species. Theor. Appl. Genet. 1973, 43, 75–78. [Google Scholar] [CrossRef] [PubMed]
- Brock, M.A. Reproductive allocation in annual and perennial species of the submerged aquatic halophyte Ruppia. J. Ecol. 1983, 71, 811–818. [Google Scholar] [CrossRef]
- Bazzaz, F.A.; Chiariello, N.R.; Coley, P.D.; Pitelka, L.F. Allocating resources to reproduction and defense: New assessments of the costs and benefits of allocation patterns in plants are relating ecological roles to resource use. Bioscience 1987, 37, 58–67. [Google Scholar] [CrossRef]
- Van Tassel, D.L.; DeHaan, L.R.; Cox, T.S. Missing domesticated plant forms: Can artificial selection fill the gap? Evol. Appl. 2010, 3, 434–452. [Google Scholar] [CrossRef] [PubMed]
- Vico, G.; Manzoni, S.; Nkurunziza, L.; Murphy, K. Trade-offs between seed output and life span–a quantitative comparison of traits between annual and perennial congeneric species. New Phytol. 2016, 209, 104–114. [Google Scholar] [CrossRef]
- Roumet, C.; Urcelay, C.; Díaz, S. Suites of root traits differ between annual and perennial species growing in the field. New Phytol. 2006, 170, 357–368. [Google Scholar] [CrossRef] [Green Version]
- Garcia, C.B.; Grusak, M.A. Mineral accumulation in vegetative and reproductive tissues during seed development in Medicago truncatula. Front. Plant Sci. 2015, 6, 622. [Google Scholar] [CrossRef] [PubMed]
- Khan, J.A.; Narayana, K.K.; Holla, S.; Shrinivas, S.M.; Dar, Z.A.; Shashidhar, H.E. Micronutrient productivity: A comprehensive parameter for biofortification in rice (Oryza sativa L.) grain. J. Sci. Food Agric. 2019, 99, 1311–1321. [Google Scholar] [CrossRef] [PubMed]
- Porch, T.G.; Cichy, K.; Wang, W.; Brick, M.; Beaver, J.S.; Santana-Morant, D.; Grusak, M.A. Nutritional composition and cooking characteristics of tepary bean (Phaseolus acutifolius Gray) in comparison with common bean (Phaseolus vulgaris L.). Genet. Resour. Crop Evol. 2017, 64, 935–953. [Google Scholar] [CrossRef]
- Celmeli, T.; Sari, H.; Canci, H.; Sari, D.; Adak, A.; Eker, T.; Toker, C. The nutritional content of common bean (Phaseolus vulgaris L.) landraces in comparison to modern varieties. Agronomy 2018, 8, 166. [Google Scholar] [CrossRef]
- Bhardwaj, H.L.; Hamama, A.A. Protein and mineral composition of tepary bean seed. HortScience 2004, 39, 1363–1365. [Google Scholar] [CrossRef]
- Mhlaba, Z.B.; Mashilo, J.; Shimelis, H.; Assefa, A.B.; Modi, A.T. Progress in genetic analysis and breeding of tepary bean (Phaseolus acutifolius A. Gray): A review. Sci. Hortic. 2018, 237, 112–119. [Google Scholar] [CrossRef]
- Souter, J.R.; Gurusamy, V.; Porch, T.G.; Bett, K.E. Successful introgression of abiotic stress tolerance from wild tepary bean to common bean. Crop Sci. 2017, 57, 1160–1171. [Google Scholar] [CrossRef]
- Baxter, I.; Dilkes, B.P. Elemental profiles reflect plant adaptations to the environment. Science 2012, 336, 1661–1663. [Google Scholar] [CrossRef]
- Horton, M.W.; Hancock, A.M.; Huang, Y.S.; Toomajian, C.; Atwell, S.; Auton, A.; Muliyati, N.W.; Platt, A.; Sperone, F.G.; Vilhjálmsson, B.J.; et al. Genome-wide patterns of genetic variation in worldwide Arabidopsis thaliana accessions from the RegMap panel. Nat. Genet. 2012, 44, 212–216. [Google Scholar] [CrossRef]
- Sotelo, A.; Sousa, H.; Sánchez, M. Comparative study of the chemical composition of wild and cultivated beans (Phaseolus vulgaris). Plant Foods Hum. Nutr. 1995, 47, 93–100. [Google Scholar] [CrossRef]
- Guzmán-Maldonado, S.H.; Acosta-Gallegos, J.; Paredes-López, O. Protein and mineral content of a novel collection of wild and weedy common bean (Phaseolus vulgaris L). J. Sci. Food Agric. 2000, 80, 1874–1881. [Google Scholar] [CrossRef]
- Baxter, I. Should we treat the ionome as a combination of individual elements, or should we be deriving novel combined traits? J. Exp. Bot. 2015, 66, 2127–2131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maillard, A.; Etienne, P.; Diquélou, S.; Trouverie, J.; Billard, V.; Yvin, J.-C.; Ourry, A. Nutrient deficiencies modify the ionomic composition of plant tissues: A focus on cross-talk between molybdenum and other nutrients in Brassica napus. J. Exp. Bot. 2016, 67, 5631–5641. [Google Scholar] [CrossRef] [PubMed]
- Hacisalihoglu, G.; Settles, A.M. Quantification of seed ionome variation in 90 diverse soybean (Glycine max) lines. J. Plant Nutr. 2017, 40, 2808–2817. [Google Scholar] [CrossRef]
- Ghandilyan, A.; Barboza, L.; Tisné, S.; Granier, C.; Reymond, M.; Koornneef, M.; Schat, H.; Aarts, M.G.M. Genetic analysis identifies quantitative trait loci controlling rosette mineral concentrations in Arabidopsis thaliana under drought. New Phytol. 2009, 184, 180–192. [Google Scholar] [CrossRef] [PubMed]
- Pineau, C.; Loubet, S.; Lefoulon, C.; Chalies, C.; Fizames, C.; Lacombe, B.; Ferrand, M.; Loudet, O.; Berthomieu, P.; Richard, O. Natural variation at the FRD3 MATE transporter locus reveals cross-talk between Fe homeostasis and Zn tolerance in Arabidopsis thaliana. PLoS Genet. 2012, 8, e1003120. [Google Scholar] [CrossRef]
- Broadley, M.R.; White, P.J. Some elements are more equal than others: Soil-to-plant transfer of radiocaesium and radiostrontium, revisited. Plant Soil 2012, 355, 23–27. [Google Scholar] [CrossRef]
- Watanabe, T.; Urayama, M.; Shinano, T.; Okada, R.; Osaki, M. Application of ionomics to plant and soil in fields under long-term fertilizer trials. Springerplus 2015, 4, 781. [Google Scholar] [CrossRef]
- Broadley, M.R.; Willey, N.J.; Wilkins, J.C.; Baker, A.J.M.; Mead, A.; White, P.J. Phylogenetic variation in heavy metal accumulation in angiosperms. New Phytol. 2001, 152, 9–27. [Google Scholar] [CrossRef]
- Neugebauer, K.; Broadley, M.R.; El-Serehy, H.A.; George, T.S.; McNicol, J.W.; Moraes, M.F.; White, P.J. Variation in the angiosperm ionome. Physiol. Plant. 2018, 163, 306–322. [Google Scholar] [CrossRef] [Green Version]
- Du, Q.; Wei, D.; Chou, K.-C. Correlations of amino acids in proteins. Peptides 2003, 24, 1863–1869. [Google Scholar] [CrossRef] [PubMed]
- Tinsley, A.M.; Scheerens, J.C.; Alegbejo, J.O.; Adan, F.H.; Krumhar, K.C.; Butler, L.E.; Kopplin, M.J. Tepary beans (Phaseolus acutifolius var. latifolius): A potential food source for African and Middle Eastern cultures. Qualitas Plantarum. Plant Foods Hum. Nutr. 1985, 35, 87–101. [Google Scholar] [CrossRef]
- Mahe, S.; Gausseres, N.; Tome, D. Legume proteins for human requirements. Grain Legumes (AEP) 1994, 7, 15–17. [Google Scholar]
- Myers, S.S.; Zanobetti, A.; Kloog, I.; Huybers, P.; Leakey, A.D.B.; Bloom, A.J.; Carlisle, E.; Dietterich, L.H.; Fitzgerald, G.; Hasegawa, T.; et al. Increasing CO2 threatens human nutrition. Nature 2014, 510, 139. [Google Scholar] [CrossRef]
- Pilbeam, D.J. Breeding crops for improved mineral nutrition under climate change conditions. J. Exp. Bot. 2015, 66, 3511–3521. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lifespan | Species | Wild/Cultivated | Variety | Geographic Provenance | Accession |
|---|---|---|---|---|---|
| Perennial | Phaseolus angustissimus A. Gray | Wild | Arizona, USA | NI 788 | |
| New Mexico, USA | *NI 878 | ||||
| New Mexico, USA | NI 1303 | ||||
| Phaseolus filiformis Benth. | Wild | Arizona, USA | *PI 535294 | ||
| Arizona, USA | PI 535299 | ||||
| Arizona, USA | PI 535300 | ||||
| Arizona, USA | PI 535303 | ||||
| Arizona, USA | PI 535306 | ||||
| Sonora, Mexico | PI 535307 | ||||
| Phaseolus maculatus ssp. ritensis Freytag† | Wild | Mexico | *PI 661844 | ||
| Jalisco, Mexico | PI 494138 | ||||
| Arizona, USA | PI 535372 | ||||
| Phaseolus polyanthus Greenm. | Wild | Guatemala | *NI 1112 | ||
| Cultivated | Guatemala | NI 1015 | |||
| Mexico | NI 1340 | ||||
| Costa Rica | NI 429 | ||||
| Mexico | NI 519 | ||||
| Guatemala | NI 758 | ||||
| Colombia | NI 553 | ||||
| Guatemala | NI 913 | ||||
| Annual | Phaseolus acutifolius A. Gray | Wild | var. acutifolius | Mexico | PI 535200 |
| Arizona, USA | *PI 535202 | ||||
| Mexico | PI 535205 | ||||
| Phaseolus acutifolius A. Gray | Wild | var. tenuifolius | Arizona, USA | *PI 535236 | |
| Arizona, USA | PI 535248 | ||||
| Mexico | PI 638911 | ||||
| Phaseolus vulgaris L. | Wild | Mexico | NI 1192 | ||
| Mexico | NI 1189 | ||||
| Mexico | NI 1218 | ||||
| Mexico | *NI 1248 | ||||
| Mexico | NI 1205 | ||||
| Mexico | NI 1193 | ||||
| Mexico | NI 1249 | ||||
| Mexico | NI 1253 | ||||
| Mexico | NI 1286 | ||||
| Mexico | W6 18752 |
| Ion | Lifespan | Species (Lifespan) | Variety | Cultivation Status |
|---|---|---|---|---|
| Annual Mean ± SD | var. acutifolius | Wild Mean ± SD | ||
| Perennial Mean ± SD | var. tenuifolius | Cultivated Mean ± SD | ||
| Aluminum (Al) | 0.04 | 0.39 | 0.02 | 0.03 |
| Arsenic (As) | 1.88 | 1.63 | 0.01 | 0.43 |
| Boron (B) | 3.47 | 4.46 ** | 0.08 | 9.24 ** |
| W: 2.06 ± 2.25 | ||||
| C: 0.06 ± 0.99 | ||||
| Cadmium (Cd) | 1.39 | 0.50 | 0.00 | 0.01 |
| Calcium (Ca) | 19.33 *** | 0.65 | 1.02 | 3.46 |
| A: 0.67 ± 0.76 | ||||
| P: −0.56 ± 0.82 | ||||
| Cobalt (Co) | 2.02 | 2.20 | 0.03 | 0.33 |
| Copper (Cu) | 15.26 *** | 2.00 | 2.49 | 0.06 |
| A: 0.56 ± 0.86 | ||||
| P: −0.47 ± 0.87 | ||||
| Iron (Fe) | 5.03 * | 1.63 | 0.15 | 1.57 |
| A: 0.48 ± 1.14 | ||||
| P: −0.41 ± 0.64 | ||||
| Lithium (Li) | 2.38 | 5.94 ** | 0.10 | 0.00 |
| Magnesium (Mg) | 7.14 * | 1.18 | 2.56 | 0.36 |
| A: 0.41 ± 1.09 | ||||
| P: −0.35 ± 0.79 | ||||
| Manganese (Mn) | 6.37 * | 0.82 | 0.21 | 0.61 |
| A: 0.51 ± 1.16 | ||||
| P: −0.43 ± 0.57 | ||||
| Molybdenum (Mo) | 0.28 | 1.12 | 0.23 | 0.79 |
| Nickel (Ni) | 0.88 | 1.74 | 0.86 | 0.39 |
| Phosphorus (P) | 13.93 *** | 1.23 | 1.05 | 4.64 * W: 0.59 ± 2.20 C: −0.83 ± 0.46 |
| A: 0.62 ± 0.73 | ||||
| P: −0.52 ± 0.91 | ||||
| Potassium (K) | 3.54 | 6.71 *** | 0.46 | 0.22 |
| Rubidium (Rb) | 1.02 | 2.03 | 0.10 | 0.00 |
| Selenium (Se) | 0.41 | 5.92 ** | 1.69 | 0.03 |
| Sodium (Na) | 20.62 *** A: −0.36 ± 0.61 P: 0.31 ± 1.17 | 33.54 *** | 0.24 | 0.00 |
| Strontium (Sr) | 9.28 ** | 2.33 | 0.12 | 5.94 * |
| A: 0.56 ± 0.71 | W: 0.73 ± 0.44 | |||
| P: −0.47 ± 0.98 | C: −0.86 ± 0.31 | |||
| Sulfur (S) | 14.47 *** | 13.15 *** | 2.41 | 3.31 |
| A: 0.37 ± 0.90 | ||||
| P: −0.31 ± 1.00 | ||||
| Zinc (Zn) | 0.15 | 0.97 | 4.87 * | 3.00 |
| a: 1.24 ± 1.61 | ||||
| t: −0.69 ± 0.59 |
| Amino Acid | Lifespan | Species (Lifespan) | Variety | Cultivation Status |
|---|---|---|---|---|
| Annual Mean ± SD | ||||
| Perennial Mean ± SD | ||||
| Alanine (Ala) | 0.07 | 0.74 | 3.15 | 0.13 |
| Arginine (Arg) | 0.01 | 0.32 | 1.83 | 0.03 |
| Aspartic acid (Asp) | 0.01 | 1.4 | 2.64 | 0.57 |
| Glutamic acid (Glu) | 0.00 | 0.07 | 3.42 | 0.32 |
| Glycine (Gly) | 0.06 | 0.37 | 1.98 | 0.36 |
| Histidine (His) | 0.06 | 1.60 | 1.91 | 0.45 |
| Isoleucine (Ile) | 0.25 | 0.52 | 1.93 | 0.40 |
| Leucine (Leu) | 0.17 | 0.42 | 1.79 | 0.36 |
| Lysine (Lys) | 3.73 | 1.56 | 2.58 | 0.28 |
| Methionine (Met) | 0.37 | 0.30 | 1.45 | 0.44 |
| Phenylalanine (Phe) | 0.75 | 0.82 | 1.86 | 0.41 |
| Proline (Pro) | 0.75 | 0.19 | 2.38 | 0.60 |
| Serine (Ser) | 5.92 * | 3.97 * | 0.76 | 2.03 |
| A: −0.39 ± 0.77 | ||||
| P: 0.42 ± 1.07 | ||||
| Threonine (Thr) | 0.56 | 4.00 * | 2.42 | 0.76 |
| Tyrosine (Tyr) | 5.74 * | 3.11 * | 0.92 | 1.66 |
| A: 0.44 ± 0.87 | ||||
| P: −0.47 ± 0.94 | ||||
| Valine (Val) | 0.08 | 0.51 | 2.56 | 0.14 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schier, H.E.; Eliot, K.A.; Herron, S.A.; Landfried, L.K.; Migicovsky, Z.; Rubin, M.J.; Miller, A.J. Comparative Analysis of Perennial and Annual Phaseolus Seed Nutrient Concentrations. Sustainability 2019, 11, 2787. https://doi.org/10.3390/su11102787
Schier HE, Eliot KA, Herron SA, Landfried LK, Migicovsky Z, Rubin MJ, Miller AJ. Comparative Analysis of Perennial and Annual Phaseolus Seed Nutrient Concentrations. Sustainability. 2019; 11(10):2787. https://doi.org/10.3390/su11102787
Chicago/Turabian StyleSchier, Heather E., Kathrin A. Eliot, Sterling A. Herron, Lauren K. Landfried, Zoë Migicovsky, Matthew J. Rubin, and Allison J. Miller. 2019. "Comparative Analysis of Perennial and Annual Phaseolus Seed Nutrient Concentrations" Sustainability 11, no. 10: 2787. https://doi.org/10.3390/su11102787
APA StyleSchier, H. E., Eliot, K. A., Herron, S. A., Landfried, L. K., Migicovsky, Z., Rubin, M. J., & Miller, A. J. (2019). Comparative Analysis of Perennial and Annual Phaseolus Seed Nutrient Concentrations. Sustainability, 11(10), 2787. https://doi.org/10.3390/su11102787