Mineral Composition of Durum and Wild Wheat
The 25 wheat genotypes were used to assess the potential similarities and/or differences in the mineral content for the considered two harvesting sites (A, Valenzano, Bari, and B, Policoro, Matera) and among the different cultivars. All samples were grown in both the two harvesting sites and three replicates were collected for each sample, reaching a total of 148 (73 and 75 for site A and B, respectively). According to the wheat type, samples were also classified into durum (63 for each site, for a total of 126 samples) and wild (10 and 12 for site A and B, for a total of 22 samples).
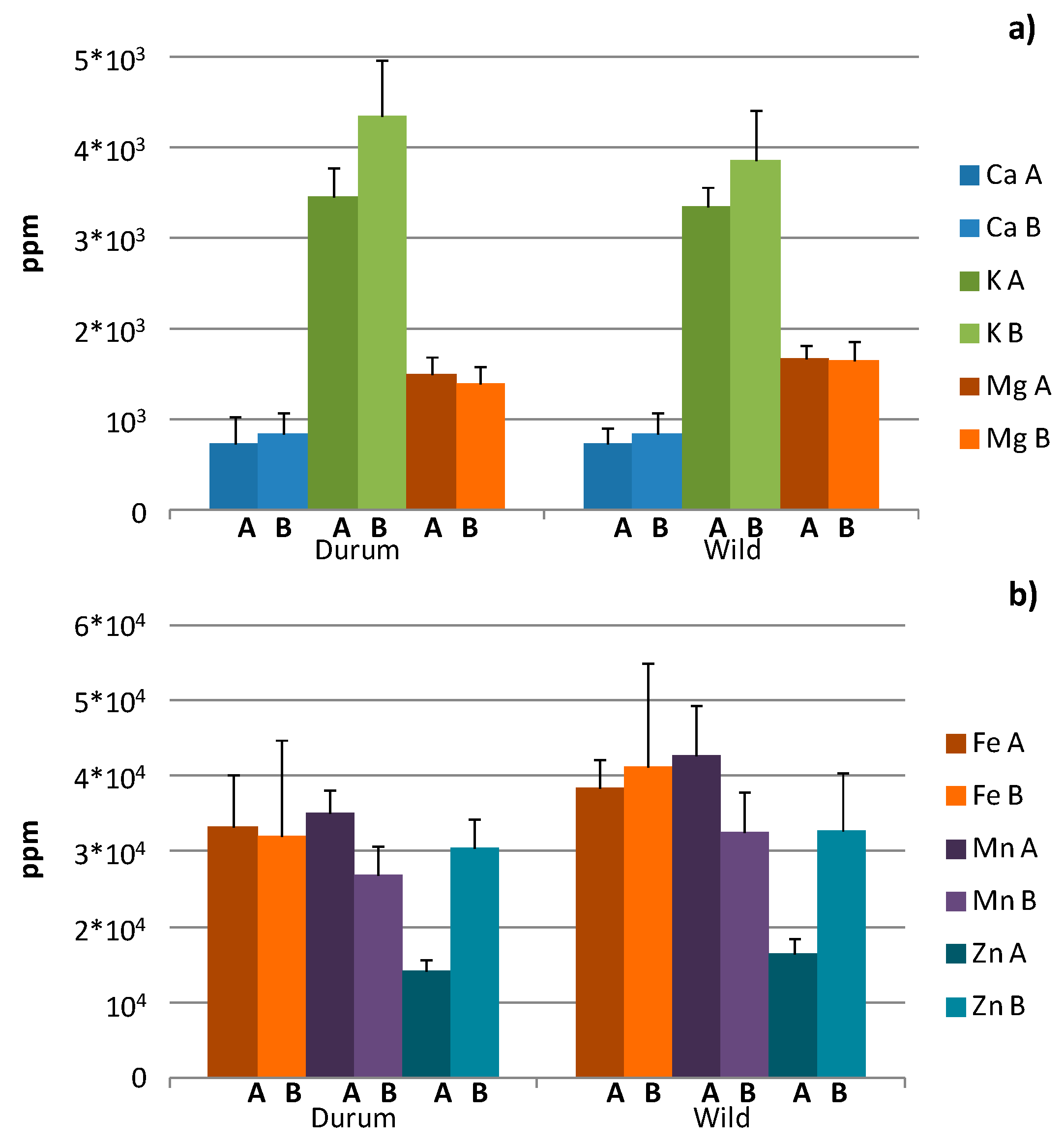
Average and standard deviation values for concentrations of macro- (Ca, K, Mg), micro- (Cr, Cu, Fe, Mn, Zn) and toxic elements (Cd, Pb), which were expressed in part per million (ppm), were measured for all the studied cultivars and sites and are reported in
Supplementary material (Tables S1 and S2). A summary clustering all of the studied cultivars according to the different wheat species (durum and wild) and the two cultivation sites (A and B) is reported in
Table 2 and
Figure 1. Tukey HSD test was applied for multiple comparisons of groups (wheat type) and One-way ANOVA was applied to compare the means between the two cultivation sites. As expected [
44,
45], among all the measured elements, the highest concentration was found for K, followed by Mg and Ca for all the examined wheat types. In particular, the highest K average value was found for durum wheat (4357.19 ± 618.72 ppm), followed by wild wheat (3864.32 ± 561.39 ppm). On the other hand, the highest Mg content resulted for wild wheat, with a similar content for the two cultivation sites (1665.97 ± 147.10 and 1651.89 ± 203.23 ppm for site A and B, respectively). Moreover, for durum wheat, a significantly lower value for Mg content was found in site B with respect to site A. Average Ca values were comparable in the two wheat types, with the highest content for durum wheat samples (833.08 ± 240.16 ppm) being significantly different from the durum wheat of site A (731.11 ± 295.70 ppm). It should be noted that for each type of tetraploid wheats, the average content of the macro-elements had the expected sequence K > Mg > Ca when considering both the harvesting sites [
45]. Regarding micro-elements, in accordance with the literature data [
27,
37,
45,
46], higher levels of Mn, Fe and Zn and lower Cu and Cr, with Ni (Ni data not shown), in very low traces for all the samples were found in wild wheat. Interestingly, the highest Mn and Fe values were measured in wild wheat, followed by durum genotypes for both the harvesting sites. Generally higher values of Mn resulted in site A with respect to site B, while similar content of Fe was measured in the two sites. Very significant and site-dependent differences were found only for Zn, with significantly higher levels for the three tetraploid subspecies in site B with respect to site A. Moreover, the highest Zn content was found in wild (32.86 ± 7.50 ppm), followed by durum (30.59 ± 3.69 ppm), all from site B. Finally, higher Cd and Pb levels were found in durum wheat genotypes, with the highest in samples of site A (Cd = 0.10 ± 0.04 ppm, Pb = 0.09 ± 0.04 ppm), followed by wild for both the two harvesting sites.
A further level of investigation was performed by calculating the correlation matrix based on Pearson’s coefficient for all the measured elements. An overview about the potential linear relationship between the metals (macronutrients, Ca, Mg, K, micronutrients, Zn, Fe, Cu, Mn and toxic trace elements, Cd, Pb) was obtained for each tetraploid genotype (durum and wild) and for each cultivation site, A and B (
Table 3,
Table 4,
Table 5 and
Table 6). Although a simple correlation analysis, especially considering that a large data set should be carefully used for expressing nutrient relationships, we could find and will discuss a limited range of significant correlations that were obtained in this study. A high level of correlation was observed for durum wheat for both the two sites: in particular six couples of elements (Cu/Zn, Mg/Zn with significance at
p < 0.001 and Cu/Mg, Cu/Mn, Zn/Mn, Cr/Fe with significance at
p < 0.01) for site A and seven couples of elements (K/Cu, K/Mg, K/Mg, Cu/Mg, Cu/Mn, Cr/Fe with significance at
p < 0.001 and Ca/Cd with significance at
p < 0.01) for site B (
Table 3 and
Table 5), while wild wheat showed a low number of correlations in both the two sites (Cr/Cu, Fe/Mn with significance at
p < 0.001 and Cr/Fe, Cr/Mg, Cd/Mn and Ca/Mg with significance at
p < 0.01 for site A; Cr/Fe with significance at
p < 0.001 and Ca/Cd, K/Zn, K/Cr, K/Fe, Cr/Fe with significance at
p < 0.01 for site B,
Table 4 and
Table 6). It should also be noted that in some cases, more significant or positive correlation values for other elements (i.e., Mn/Mg) were expected, although comparable results to those that were obtained in this work were also reported in literature [
28,
37,
47,
48].
Multivariate statistical analysis (PCA and OPLS-DA) was used to deeply investigate the variation in macronutrients (Ca, Mg, K), micronutrients (Zn, Fe, Cu, Mn) and toxic trace elements (Cd, Pb) for the whole dataset of 25 wheat accessions and cultivars (three replicates for each cultivar studied) that is representative of durum and wild species in the two cultivation sites. The whole data were studied by OPLS-DA in order to evaluate the potential effect of the different pedoclimatic conditions on the wheat species (
Figure 2). In particular, two OPLS-DA models were built using the same number of components (OPLS-DA model of site A: 1 + 2 + 0, R
2X = 0.48, R
2Y = 0.59, Q
2 = 0.43, p[CV-ANOVA] = 2.40612 × 10
−6; for OPLS-DA model of site B: 1 + 2 + 0, R
2X = 0.56, R
2Y = 0.83, Q
2 = 0.78, p[CV-ANOVA] = 2.10965 × 10
−19). This approach showed that different pedoclimatic conditions characterize the two sites. Samples cultivated in site A (Valenzano, Bari) were homogeneously distributed in the space of the OPLS-DA graph, while those cultivated in site B (Policoro, Matera) appeared well differentiated in two groups (durum, wild).
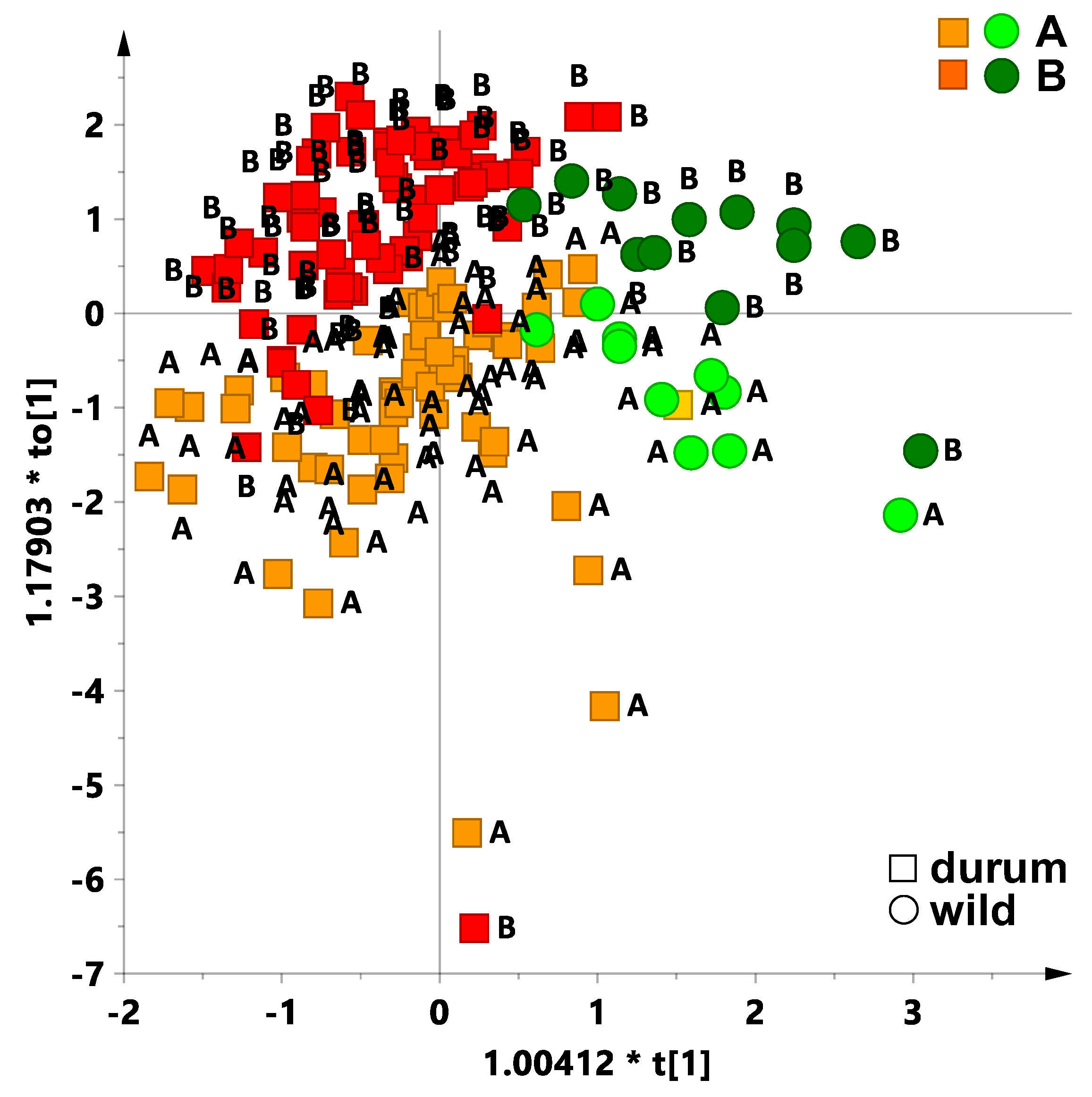
With the aim to assess the different element uptake ability of the studied tetraploid wheat subspecies, an OPLS-DA analysis was performed for the whole data. In the first place, the dataset was analysed by differentiating in the model two different categories, the durum and the wild wheat types. The resulting OPLS-DA model, which was built with one predictive (t[1]) and three orthogonal components (1 + 3 + 0) beside the observed classification parameters for the two categories (R
2X = 0.58, Q
2 = 0.49) gave an interesting hint for the sample distribution among the two sites (
Figure 3). Indeed, this appears to be the natural discrimination (R
2Y = 0.53) observed in the first orthogonal component (to[1]). The colour encoded sample distribution among the two sites is clearly observed in
Figure 3. The OPLS-DA scoreplot showed that durum wheat genotypes were mainly distributed at negative values of the predictive component t[1], while the intra-class variation resulted in a large distribution of the data along the predictive component to[1] in a wide range of values (from −7 to 3). On the other hand, wild wheat accessions were clustered both at positive values of t[1] and in a range of −2 and +2 of to[1].
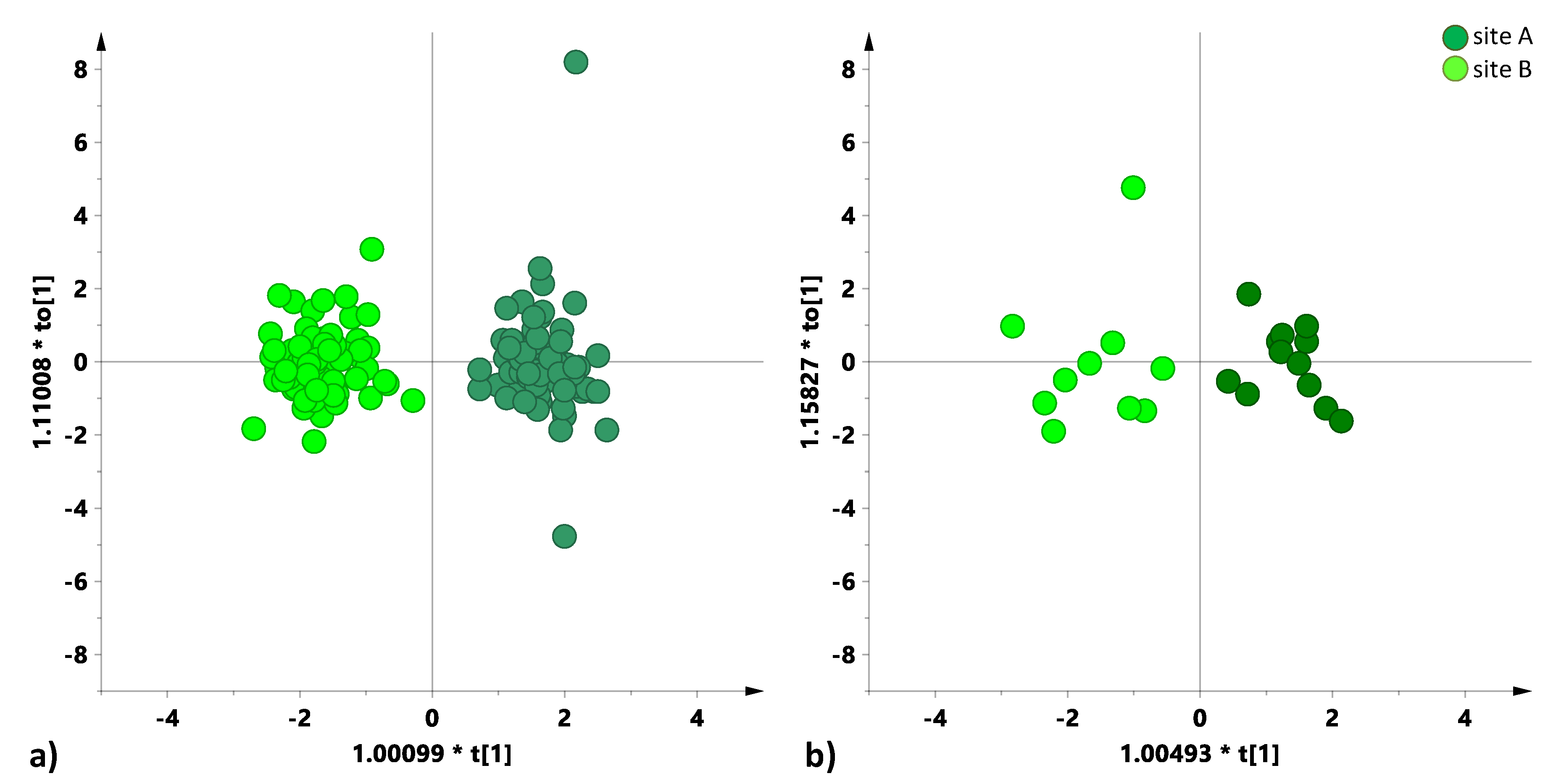
A further MVA was then performed with the aim to differentiate, according to the cultivation site, durum and wild wheat types. For this purpose, two independent PCA and OPLS-DA models were built using the cultivation site as discrimination class. Both for the PCA (data not shown) and OPLS-DA (
Figure 4) models, the two wheat groups resulted differently for the two cultivation sites. According to the OPLS-DA models that are depicted in
Figure 4, and relative model parameters (in particular Q
2 values), the more pronounced separation among the two cultivation sites was observed for durum (Q
2 = 0.92) with respect to wild type (Q
2 = 0.75) wheat. Moreover, due to a different sample size of durum (126 samples) in comparison with wild (22 samples) statistical models, a further cross check was applied to assess the soundness of the obtained parameters for the OPLS-DA models that are reported in
Figure 4. By using the Weka open-source data mining software (v. 3.8.3, University of Waikato New Zealand) [
49], a filter was applied to randomly remove a given percentage of samples from the durum sample set. For this purpose, 80% of durum samples were randomly excluded in order to obtain a comparable sample size for durum set (25) with respect to wild (22) samples. Subsequently, a Naïve Bayes classification was applied separately for both the durum and wild sets. The models’ reliability (indicated with “Correctly Classified Instances”
Tables S3 and S4), resulted with 87.5% and 71.4% for durum and wild samples, respectively, confirming our hypothesis that wild wheat is the less site sensitive species. This result suggested that a profitable search for the less site sensitive species had to be performed within the considered wild wheat cultivars.
Finally, new OPLS-DA models were then calculated by using the four available data sets (three replicates for each of the two cultivation sites) for the four wild wheat accessions. The cultivation site was chosen as the discriminating class for each of the four OPLS-DA models. The obtained calculated quality model parameters are reported in
Table 7, showing the lowest (MG29896) and highest (MG5323) cultivation site differentiation. Among the genotypes of wild wheat tetraploids, MG29896 resulted as the least affected from the cultivation site, having the lowest predictive ability, expressed as Q
2 value, followed by MG4330, MG5323.

 ,
, 






{kind=link}
{kind=link}
{kind=link}
{kind=link}