Effects of Aquatic Macrophytes on Spatial Distribution and Feeding Habits of Exotic Fish Species Lepomis macrochirus and Micropterus salmoides in Shallow Reservoirs in South Korea


Abstract
:1. Introduction
2. Materials and Methods
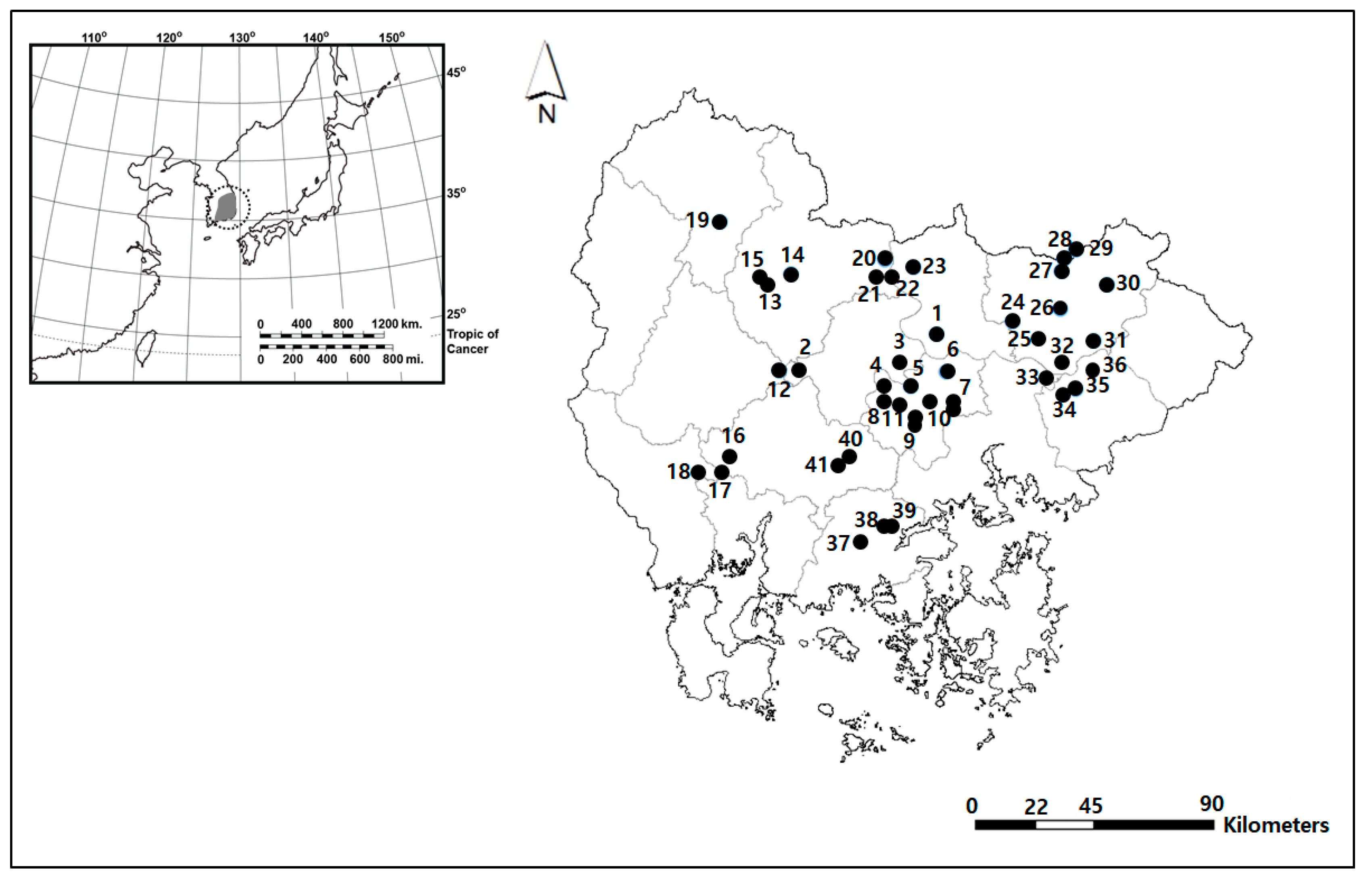
2.1. Study Area
2.2. Monitoring Strategy
2.3. Data Analysis
3. Results
3.1. Environmental Parameters and Invertebrate Distribution
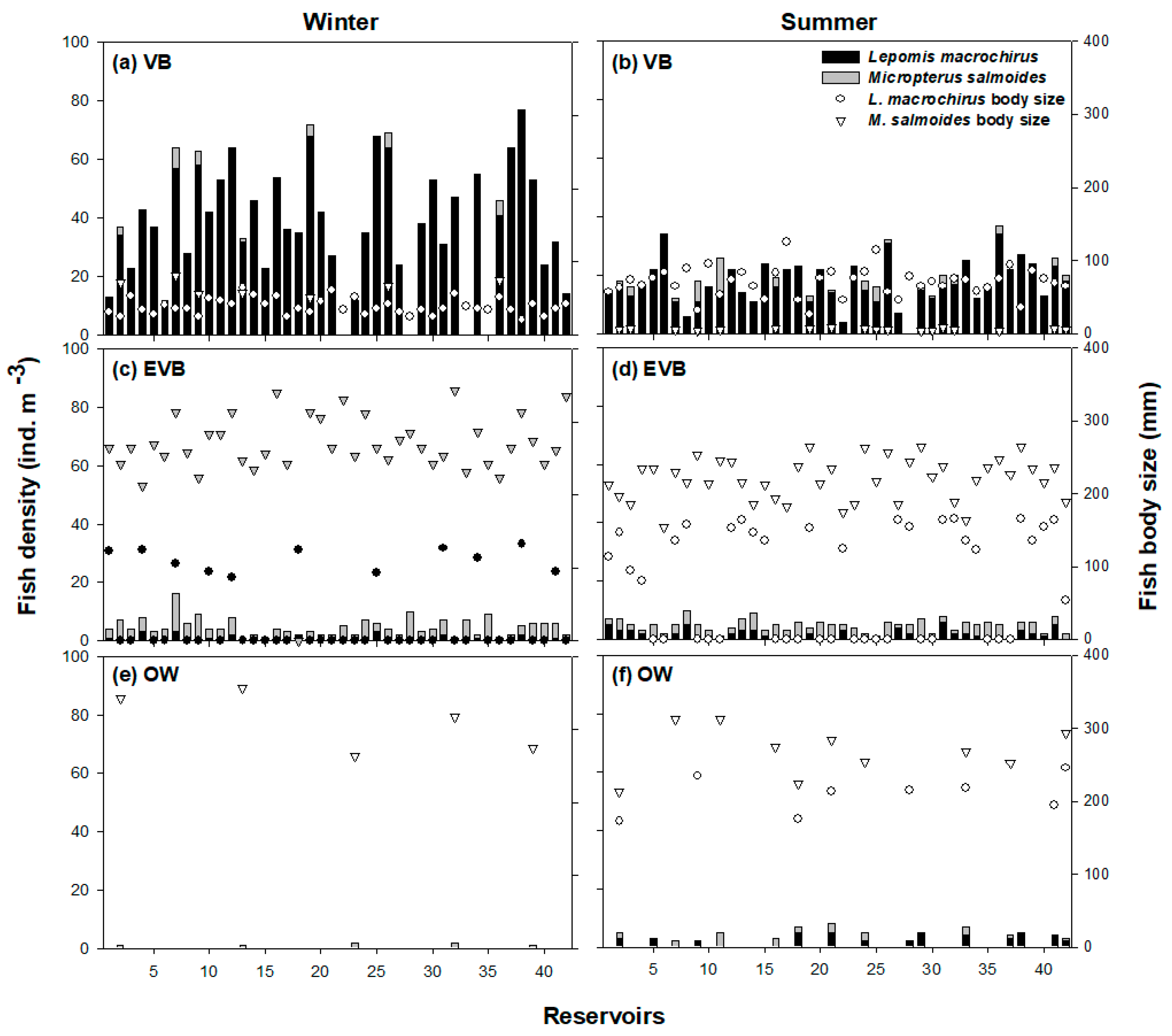
3.2. Distribution of Species among Different Zones
3.3. Prey Consumption Patterns
4. Discussion
4.1. Microhabitat Characteristics Influence Fish Distribution
4.2. Role of Winter Macrophytes for Lepomis macrochirus
4.3. Absence of Predation on L. macrochirus and M. salmoides
4.4. Strategy for Managing Exotic Fish Species
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Bazzaz, F.A. Plant species diversity in old-field successional ecosystems in southern Illinois. Ecology 1975, 56, 485–488. [Google Scholar] [CrossRef]
- Chick, J.H.; McIvor, C.C. Patterns in the abundance and composition of fishes among beds of different macrophytes: Viewing a littoral zone as a landscape. Can. J. Fish. Aquat. Sci. 1994, 51, 2873–2882. [Google Scholar] [CrossRef]
- Manatunge, J.; Asaeda, T.; Priyadarshana, T. The influence of structural complexity on fish–zooplankton interactions: A study using artificial submerged macrophytes. Environ. Biol. Fishes 2000, 58, 425–438. [Google Scholar] [CrossRef]
- Ruetz, C.R.; Breen, M.J.; Vanhaitsma, D.L. Habitat structure and fish predation: Effects on invertebrate colonization and breakdown of stream leaf packs. Freshw. Biol. 2006, 51, 797–806. [Google Scholar] [CrossRef]
- Genkai-Kato, M. Macrophyte refuges, prey behaviour and trophic interactions: Consequences for lake water clarity. Ecol. Lett. 2007, 10, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Declerck, S.; Vandekerkhove, J.; Johansson, L.; Muylaert, K.; Conde-Porcuna, J.M.; Van der Gucht, K.; Pérez-Martínez, C.; Lauridsen, T.; Schwenk, K.; Zwart, G.; et al. Multi-group biodiversity in shallow lakes along gradients of phosphorus and water plant cover. Ecology 2005, 86, 1905–1915. [Google Scholar] [CrossRef] [Green Version]
- Vieira, L.C.G.; Bini, L.M.; Velho, L.F.M.; Mazão, G.R. Influence of spatial complexity on the density and diversity of periphytic rotifers, microcrustaceans and testate amoebae. Fundam. Appl. Limnol. 2007, 170, 77–85. [Google Scholar] [CrossRef]
- Meerhoff, M.; Mazzeo, N.; Moss, B.; Rodríguez-Gallego, L. The structuring role of free-floating versus submerged plants in a subtropical shallow lake. Aquat. Ecol. 2003, 37, 377–391. [Google Scholar] [CrossRef]
- Choi, J.Y.; Jeong, K.S.; La, G.H.; Kim, S.K.; Joo, G.J. Sustainment of epiphytic microinvertebrate assemblage in relation with different aquatic plant microhabitats in freshwater wetlands (South Korea). J. Limnol. 2014, 73, 11–16. [Google Scholar] [CrossRef] [Green Version]
- Jeppesen, E.; Lauridsen, T.L.; Kairesalo, T.; Perrow, M.R. Impact of submerged macrophytes on fish–zooplankton interactions in lakes. In The Structuring Role of Submerged Macrophytes in Lakes; Springer Verlag: New York, NY, USA, 1998; pp. 91–114. [Google Scholar]
- Blindow, I.; Hargeby, A.; Bálint, M.A.; Andersson, G. How important is the crustacean plankton for the maintenance of water clarity in shallow lakes with abundant submerged vegetation? Freshw. Biol. 2000, 44, 185–197. [Google Scholar] [CrossRef]
- Rooke, J.B. The invertebrate fauna of four macrophytes in a lotic system. Freshw. Biol. 1984, 14, 507–513. [Google Scholar] [CrossRef]
- Dudley, T.L. The role of plant complexity and epiphyton in colonization of macrophytes by stream insects. Int. Vereinigung Theoretische Angewandte Limnol. 1988, 23, 1153–1158. [Google Scholar] [CrossRef]
- Sand-Jensen, K.; Søndergaard, M. Phytoplankton and epiphyte development and their shading effect on submerged macrophytes in lakes of different nutrient status. Int. Rev. Gesamten Hydrobiol. Hydrogr. 1981, 66, 529–552. [Google Scholar] [CrossRef]
- Van Donk, E.; van de Bund, W.J. Impact of submerged macrophytes including charophytes on phyto- and zooplankton communities: Allelopathy versus other mechanisms. Aquat. Bot. 2002, 72, 261–274. [Google Scholar] [CrossRef]
- Warfe, D.M.; Barmuta, L.A. Habitat structural complexity mediates food web dynamics in a freshwater macrophyte community. Oecologia 2006, 150, 141–154. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.P.; Frost, N.J.; Mosley, M.W.; Roberts, M.F.; Hawkins, S.J. The area-independent effects of habitat complexity on biodiversity vary between regions. Ecol. Lett. 2003, 6, 126–132. [Google Scholar] [CrossRef]
- Lauridsen, T.L.; Lodge, D.M. Avoidance by Daphnia magna of fish and macrophytes: Chemical cues and predator-mediated use of macrophyte habitat. Limnol. Oceanogr. 1996, 41, 794–798. [Google Scholar] [CrossRef]
- Moss, B.; Kornijow, R.; Measey, G. The effect of nymphaeid (Nuphar lutea) density and predation by perch (Perca fluviatilis) on the zooplankton communities in a shallow lake. Freshw. Biol. 1998, 39, 689–697. [Google Scholar] [CrossRef]
- Hooper, D.U.; Chapin, F.S.; Ewel, J.J.; Hector, A.; Inchausti, P.; Lavorel, S.; Lawton, J.H.; Lodge, D.M.; Loreau, M.; Naeem, S.; et al. Effects of biodiversity on ecosystem functioning: A consensus of current knowledge. Ecol. Monogr. 2005, 75, 3–35. [Google Scholar] [CrossRef]
- Okun, N.; Mehner, T. Distribution and feeding of juvenile fish on invertebrates in littoral reed (Phragmites) stands. Ecol. Freshw. Fish 2005, 14, 139–149. [Google Scholar] [CrossRef]
- Lehtiniemi, M. Swim or hide: Predator cues cause species specific reactions in young fish larvae. J. Fish Biol. 2005, 66, 1285–1299. [Google Scholar] [CrossRef]
- Crowder, L.B.; Cooper, W.E. Habitat structural complexity and the interaction between bluegills and their prey. Ecology 1982, 63, 1802–1813. [Google Scholar] [CrossRef]
- Cheruvelil, K.S.; Soranno, P.A.; Madsen, J.D.; Roberson, M.J. Plant architecture and epiphytic macroinvertebrate communities: The role of an exotic dissected macrophyte. J. N. Am. Benthol. Soc. 2002, 21, 261–277. [Google Scholar] [CrossRef]
- Meerhoff, M.; Fosalba, C.; Bruzzone, C.; Mazzeo, N.; Noordoven, W.; Jeppesen, E. An experimental study of habitat choice by Daphnia: Plants signal danger more than refuge in subtropical lakes. Freshw. Biol. 2006, 51, 1320–1330. [Google Scholar] [CrossRef]
- Choi, J.Y.; Jeong, K.S.; Kim, S.K.; La, G.H.; Chang, K.H.; Joo, G.J. Role of macrophytes as microhabitats for zooplankton community in lentic freshwater ecosystems of South Korea. Ecol. Inform. 2014, 24, 177–185. [Google Scholar] [CrossRef]
- Choi, J.Y.; Jeong, K.S.; La, G.H.; Chang, K.H.; Joo, G.J. The influence of aquatic macrophytes on distribution and feeding habit of two Asplanchna species (A. priodonta and A. herrickii) in shallow wetlands, South Korea. J. Limnol. 2015, 74, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Davey, A.J.H.; Hawkins, S.J.; Turner, G.F.; Doncaster, C.P. Size-dependent microhabitat use and intraspecific competition in Cottus gobio. J. Fish Biol. 2005, 67, 428–443. [Google Scholar] [CrossRef]
- Thomaz, S.M.; Dibble, E.D.; Evangelista, L.R.; Higuti, J.; Bini, L.M. Influence of aquatic macrophyte habitat complexity on invertebrate abundance and richness in tropical lagoons. Freshw. Biol. 2008, 53, 358–367. [Google Scholar] [CrossRef]
- Kuczynska-Kippen, N.; Nagengast, B. The influence of the spatial structure of hydromacrophytes and differentiating habitat on the structure of rotifer and cladoceran communities. Hydrobiologia 2006, 559, 203–212. [Google Scholar] [CrossRef]
- Hathaway, E.S. The relation of temperature to the quantity of food consumed by fishes. Ecology 1927, 8, 428–434. [Google Scholar] [CrossRef]
- Araya, H.; Contreras, P.; Alvina, M.; Vera, G.; Pak, N. A comparison between an in vitro method to determine carbohydrate digestion rate and the glycemic response in young men. Eur. J. Clin. Nutr. 2002, 56, 735. [Google Scholar] [CrossRef] [PubMed]
- Beamish, F.W.H. Swimming capacity. Fish Physiol. 1978, 7, 101–187. [Google Scholar]
- Cooke, S.J.; Grant, E.C.; Schreer, J.F.; Philipp, D.P.; Devries, A.L. Low temperature cardiac response to exhaustive exercise in fish with different levels of winter quiescence. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2003, 134, 157–165. [Google Scholar] [CrossRef]
- Pelicice, F.M.; Agostinho, A.A. Fish fauna destruction after the introduction of a non-native predator (Cichla kelberi) in a Neotropical reservoir. Biol. Invasions 2009, 11, 1789–1801. [Google Scholar] [CrossRef]
- Pelicice, F.M.; Latini, J.D.; Agostinho, A.A. Fish fauna disassembly after the introduction of a voracious predator: Main drivers and the role of the invader’s demography. Hydrobiologia 2015, 746, 271–283. [Google Scholar] [CrossRef]
- Mittelbach, G.G. Foraging efficiency and body size: A study of optimal diet and habitat use by bluegills. Ecology 1981, 62, 1370–1386. [Google Scholar] [CrossRef]
- Michelan, T.S.; Thomaz, S.M.; Mormul, R.P.; Carvalho, P. Effects of an exotic invasive macrophyte (tropical signalgrass) on native plant community composition, species richness and functional diversity. Freshw. Biol. 2010, 55, 1315–1326. [Google Scholar] [CrossRef]
- Burkett, V.; Kusler, J. Climate change: Potential impacts and interactions in wetlands of the United States. J. Am. Water Resour. Assoc. 2000, 36, 313–320. [Google Scholar] [CrossRef]
- Peterjohn, W.T.; Correll, D.L. Nutrient dynamics in an agricultural watershed: Observations on the role of a riparian forest. Ecology 1984, 65, 1466–1475. [Google Scholar] [CrossRef]
- Wetzel, R.G.; Likens, G.E. Limnological Analyses; Springer-Verlag: New York, NY, USA, 2000. [Google Scholar]
- Stansfield, J.H.; Perrow, M.R.; Tench, L.D.; Jowitt, A.J.; Taylor, A.A. Submerged macrophytes as refuges for grazing Cladocera against fish predation: Observations on seasonal changes in relation to macrophyte cover and predation pressure. In Shallow Lakes’ 95; Springer: Dordrecht, The Netherlands, 1997; pp. 229–240. [Google Scholar]
- Grenouillet, G.; Pont, D. Juvenile fishes in macrophyte beds: Influence of food resources, habitat structure and body size. J. Fish Biol. 2001, 59, 939–959. [Google Scholar] [CrossRef]
- Hambright, K.D. Experimental analysis of prey selection by largemouth bass: Role of predator mouth width and prey body depth. Trans. Am. Fish. Soc. 1991, 120, 500–508. [Google Scholar] [CrossRef]
- Savino, J.F.; Stein, R.A. Behavior of fish predators and their prey: Habitat choice between open water and dense vegetation. Environ. Biol. Fishes 1989, 24, 287–293. [Google Scholar] [CrossRef] [Green Version]
- Engel, S. The impact of submerged macrophytes on largemouth bass and bluegills. Lake Reserv. Manag. 1987, 3, 227–234. [Google Scholar] [CrossRef]
- Choi, J.Y.; Jeong, K.S.; Kim, S.K.; Joo, G.J. Impact of habitat heterogeneity on the biodiversity and density of the zooplankton community in shallow wetlands (Upo wetlands, South Korea). Oceanol. Hydrobiol. Stud. 2016, 44, 485–492. [Google Scholar] [CrossRef]
- Miranda, L.E.; Hodges, K.B. Role of aquatic vegetation coverage on hypoxia and sunfish abundance in bays of a eutrophic reservoir. Hydrobiologia 2000, 427, 51–57. [Google Scholar] [CrossRef]
- Perrow, M.R.; Jowitt, A.J.D.; Stansfield, J.H.; Phillips, G.L. The practical importance of the interactions between fish, zooplankton and macrophytes in shallow lake restoration. Hydrobiologia 1999, 395, 199–210. [Google Scholar] [CrossRef]
- Castro, B.B.; Goncalves, F. Planktivory in non-indigenous fish and implications for trophic interactions in a Mediterranean shallow lake. Ann. Limnol. Int. J. Limnol. 2011, 47, 269–280. [Google Scholar] [CrossRef] [Green Version]
- Nicolle, A.; Hansson, L.A.; Bronmark, C. Habitat structure and juvenile fish ontogeny shape zooplankton spring dynamics. Hydrobiologia 2010, 652, 119–125. [Google Scholar] [CrossRef]
- Garvey, J.E.; Ostrand, K.G.; Wahl, D.H. Energetics, predation, and ration affect size-dependent growth and mortality of fish during winter. Ecology 2004, 85, 2860–2871. [Google Scholar] [CrossRef] [Green Version]
- Rellstab, C.; Spaak, P. Lake origin determines Daphnia population growth under winter conditions. J. Plankton Res. 2008, 31, 261–271. [Google Scholar] [CrossRef] [Green Version]
- Orcutt, J.D.; Porter, K.G. The synergistic effects of temperature and food concentration of life history parameters of Daphnia. Oecologia 1984, 63, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Odenkirk, J.; Owens, S. Expansion of a Northern Snakehead population in the Potomac River System. Trans. Am. Fish. Soc. 2007, 136, 1633–1639. [Google Scholar] [CrossRef]
- Cudmore, B.; Mandrak, N.E.; Department of Fisheries and Oceans, Ottawa, ON(Canada); Canadian Science Advisory Secretariat, Ottawa, ON(Canada). Risk Assessment for Northern Snakehead (Channa Argus) in Canada; Fisheries and Oceans Canada, Science: Calgary, AB, Canada, 2006.
- Connell, J.H. A predator-prey system in the marine intertidal region. I. Balanus glandula and several predatory species of Thais. Ecol. Monogr. 1970, 40, 49–78. [Google Scholar]
- Lau, H.H.; Huang, J.; Kwan, Y.S.; Lee, W.O.; Won, Y.J. Genetic distribution pattern of bluegill sunfish Lepomis macrochirus in freshwater ecosystems across Korea. Anim. Syst. Evol. Divers. 2009, 25, 325–329. [Google Scholar] [CrossRef]
- Jo, H.; Jang, M.H.; Jeong, K.S.; Joo, G.J.; Yoon, J.D. Long-term changes in fish community and the impact of exotic fish, between the Nakdong River and Upo Wetlands. J. Ecol. Environ. 2011, 34, 59–68. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Factors | Winter | Summer | ||||
|---|---|---|---|---|---|---|
| VB | EVB | OW | VB | EVB | OW | |
| VW (cm) | 50.9 ± 26.9 | - | - | 147.6 ± 72.9 | - | - |
| Depth (m) | 0.6 ± 0.4 | 1.8 ± 2.7 | 3.7 ± 4.8 | 0.9 ± 0.6 | 2.2 ± 1.9 | 3.9 ± 5.7 |
| WT (℃) | 2.7 ± 1.7 | 2.4 ± 1.6 | 2.4 ± 2.1 | 28.4 ± 18.4 | 27.4 ± 15.6 | 27.2 ± 18.7 |
| DO (%) | 71.6 ± 23.7 | 86.2 ± 21.2 | 91.7 ± 26.8 | 17.4 ± 11.4 | 45.2 ± 20.7 | 98.6 ± 22.7 |
| pH | 8.4 ± 1.4 | 7.2 ± 0.9 | 7.1 ± 1.3 | 6.7 ± 2.7 | 7.5 ± 2.8 | 7.6 ± 1.7 |
| Cond. (µS/cm) | 227.8 ± 137 | 234.1 ± 128 | 232.7 ± 134 | 342.8 ± 247 | 357.7 ± 285 | 312.4 ± 257 |
| Chl. a (µg/L) | 5.7 ± 6.3 | 6.4 ± 3.7 | 5.6 ± 4.5 | 34.7 ± 31.7 | 36.4 ± 27.1 | 46.7 ± 26.7 |
| Branchiopods (ind/L) | 0.7 ± 6.4 | - | - | 92.3 ± 28.3 | 32.1 ± 17.1 | 17.8 ± 12.4 |
| Isopods (ind/L) | 6.8 ± 3.7 | 2.4 ± 6.9 | - | 34.7 ± 13.1 | 24.7 ± 10.4 | 16.3 ± 7.4 |
| Dipterans (ind/L) | - | - | - | 12.4 ± 6.8 | 6.7 ± 3.6 | 3.3 ± 2.7 |
| Odonatans (ind/L) | 2.4 ± 1.4 | - | - | 17.4 ± 7.2 | 10.4 ± 5.4 | 2.5 ± 2.7 |
| Fish | Component of Variance | df | Residual df | F | P |
|---|---|---|---|---|---|
| Lepomis macrochirus | Habitat | 2 | 246 | 233.32 | 0.00 |
| Season | 1 | 246 | 3.56 | 0.03 | |
| Habitat x Season | 3 | 246 | 194.2 | 0.00 | |
| Micropterus salmoides | Habitat | 2 | 246 | 87.43 | 0.02 |
| Season | 1 | 246 | 0.77 | 0.91 | |
| Habitat x Season | 3 | 246 | 8.24 | 0.63 |
| Response Variable | Explanatory Variables | Bj | t | p-Value |
|---|---|---|---|---|
| Lepomis macrochirus | Macrophyte biomass (g) | 0.018 | 10.541 | 0.000 |
| Dissolved oxygen (%) | −0.141 | −2.342 | 0.037 | |
| Micropterus salmoides | pH | −0.197 | −2.778 | 0.032 |
| Fish Species | Diet Composition | Winter | Summer | ||||
|---|---|---|---|---|---|---|---|
| VB | EVB | OW | VB | EVB | OW | ||
| Lepomis macrochirus | Branchiopods | - | - | - | 72.8 ± 64.6 | 35.4 ± 47.3 | - |
| Isopods | 8.4 ± 12.4 | - | - | 112.1 ± 82 | 51.4 ± 32.1 | 13.4 ± 22.4 | |
| Dipterans | - | - | - | 26.5 ± 32.1 | 66.7 ± 60.4 | - | |
| Odonatans | - | - | - | 13.5 ± 22.7 | 33.1 ± 32.4 | - | |
| Young fishes | - | - | - | - | - | - | |
| Micropterus salmoides | Branchiopods | - | - | - | 14.7 ± 5.7 | 2.0 ± 0.6 | - |
| Isopods | - | - | - | 26.4 ± 66.4 | 12.4 ± 7.7 | - | |
| Dipterans | - | - | - | 32.4 ± 75.1 | 53.4 ± 35.7 | - | |
| Odonatans | - | - | - | 12.5 ± 44.1 | 44.5 ± 22.8 | 11.4 ± 16.7 | |
| Young fishes | - | 6.4 ± 3.6 | 4.6 ± 4.1 | - | 6.7 ± 3.7 | 9.4 ± 13.5 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, J.-Y.; Kim, S.-K. Effects of Aquatic Macrophytes on Spatial Distribution and Feeding Habits of Exotic Fish Species Lepomis macrochirus and Micropterus salmoides in Shallow Reservoirs in South Korea. Sustainability 2020, 12, 1447. https://doi.org/10.3390/su12041447
Choi J-Y, Kim S-K. Effects of Aquatic Macrophytes on Spatial Distribution and Feeding Habits of Exotic Fish Species Lepomis macrochirus and Micropterus salmoides in Shallow Reservoirs in South Korea. Sustainability. 2020; 12(4):1447. https://doi.org/10.3390/su12041447
Chicago/Turabian StyleChoi, Jong-Yun, and Seong-Ki Kim. 2020. "Effects of Aquatic Macrophytes on Spatial Distribution and Feeding Habits of Exotic Fish Species Lepomis macrochirus and Micropterus salmoides in Shallow Reservoirs in South Korea" Sustainability 12, no. 4: 1447. https://doi.org/10.3390/su12041447
APA StyleChoi, J. -Y., & Kim, S. -K. (2020). Effects of Aquatic Macrophytes on Spatial Distribution and Feeding Habits of Exotic Fish Species Lepomis macrochirus and Micropterus salmoides in Shallow Reservoirs in South Korea. Sustainability, 12(4), 1447. https://doi.org/10.3390/su12041447