1. Introduction
Yellow pitahaya (
Hylocereus megalanthus sinonimus of
Selenicereus megalanthus) is an exotic fruit that is desired worldwide for its flavor, appearance and quality, it possesses nutritional and bioactive components (glucose content, betalains, vitamins, organic acids, dietary soluble fiber, phytoalbumins and minerals) that has allowed the fruit to be considered as a functional food [
1,
2]. This has created an increase demand of this fruit, which has resulted in the expansion of the cultivation area in many countries such as USA, Mexico, Guatemala, El Salvador, Nicaragua, Costa Rica, Venezuela, Panama, Uruguay, Peru, Brazil, Ecuador, Colombia, Thailand, Indonesia and Vietnam [
3].
In Ecuador, there are about 850 hectares of pitahaya [
4]; however, in recent years the cultivation area has been steadily increasing as exports have grown. In 2019, 7498.80 tons of fruit were exported to Hong Kong, USA, Russia, the Netherlands, France, Germany and Spain, generating more than US
$44 million in income for the country [
5]. In the country, it is grown mainly in the provinces of Pichincha, Manabí and in the Ecuadorian Amazon in Morona Santiago, Orellana and Sucumbíos [
6,
7].
Currently, pitahaya is grown commercially as a monoculture with conventional agronomic management (high use of agrochemicals), a production technology that has caused negative impacts on natural resources, such as loss of biodiversity, soil degradation and erosion due to the excessive use of agrochemicals, and total destruction of the ecosystem [
8,
9]. This type of agricultural practices worldwide is causing extreme fluctuations in climatic conditions, such as temperature increase, alteration of rainfall distribution, droughts or floods caused by the concentration of greenhouse gases CO
2, methane and nitrous oxides in the atmosphere [
10,
11]; conditions that threaten food security [
12]. New production alternatives are being investigated to reduce greenhouse gas emissions in order to mitigate the most extreme effects of climate change [
13].
Among these alternatives, there are the agroforestry systems (AFS) that has the potential to face both food security and carbon (C) sequestration mitigation goals [
14,
15]. These systems combine commercial crops (fruit trees), shrubs and trees [
6,
16] in the same area to improve the economic gains of the producers [
17]. In addition, it has been identified as a potential production approach for greenhouse gas mitigation under the Kyoto Protocol [
13].
Several studies have determined that AFSs store more carbon than monocultures, depending on biological, climatic, edaphic and site-specific management conditions [
17,
18,
19]. Ref. [
20] reported that an AFS stored twice as much carbon as a monoculture (34.61 t C ha
−1 and 18.74 t C ha
−1, respectively). This same behavior was found in the Ecuadorian Amazon in chakra-type systems (a traditional and diverse AFS) [
7] with seed-propagated cacao, where C storage was 141.4 t ha
−1 and in a monoculture was from 4.9 to 7.6 t ha
−1 [
21]. In other studies conducted in AFS in the Peruvian and Colombian Amazon, Panama and Costa Rica; the amount of C stored in cocoa AFSs with scattered trees (timber and fruit trees) was 131.18 (65.61 for aerial biomass and 65.57 for the soil component), 61, 43 to 60 and 50 to 100 t ha
−1 per year, respectively. In Mexico and Costa Rica, in coffee with
Erythrina poeppigiana, an accumulation of 115 and 195 t ha
−1 of C per year has been reported; and in Filipinas it was 93 t ha
−1 per year in cocoa with
Gliricidia sepium [
22,
23,
24,
25,
26]. In terms of economic approach, it has been reported that the producers’ income improves 13 times more in systems with fruit species (
Theobroma cacao,
Cocos nucifera and
Coffea sp.) and short cycle species (
Zea mays and
Oryza sativa) [
27]. In addition, [
28] points out that agroforestry systems are 36 to 100% more productive than monocultures, depending on the type of crop, crop arrangement and edaphoclimatic conditions.
Studies of C sequestration in AFSs with fruit trees are scarce, and specifically none or non-existent with the yellow pitahaya crop. Considering this situation, this study was initiated to investigate the hypothesis that pitahaya planted with forest species does not decrease its yield and that leguminous forest species provide nutrients to the crop and store a significant amount of carbon. The main objective of this study was to estimate the agronomic behavior of pitahaya crop in agroforestry systems compared to the monoculture. To reach this purpose, pitahaya yield, carbon storage and nutritional contribution by leguminous species was evaluated in the agroforestry system.
2. Materials and Methods
2.1. Location of the Experiment
This study was conducted from 2016 to 2020 at the Palora Experimental Farm of the Central Experimental Station of the Amazon (EECA) of the National Institute of Agricultural Research (INIAP), located in the canton Palora, province of Morona Santiago. The experimental site was located at 1°40′14.5′′ S latitude and 77°57′50.3′′ W longitude (
Figure 1), with an altitude of 864 masl, corresponding to the Piedemont Periandine [
29]. The climate of the study area is humid subtropical, with an average temperature of 20 °C, relative humidity of 89% and rainfall of 3122 mm year
−1.
According to the Development and Territorial Planning Plan for the year 2015–2025, Palora is located at 920 masl, in the northwest of the province of Morona Santiago. It has 6936 inhabitants, where 80% live in the urban area and 20% in the rural sector, with an annual population growth rate of 1.04%. The 46% of the population belongs to the economically active population which is dedicated to agriculture, livestock, forestry and fishing; while the rest of the population is dedicated to other activities, such as commerce wholesale and retail, public and private administration, construction, education, manufacturing and transportation industries. Many years ago, farmers were mainly dedicated to the cultivation of Bixa orellana, Carica papaya, Manihot esculenta, Musa paradisiaca, Colocasia esculenta, Solanum quitoense, Saccharum officinarum, Camellia sinensis and citrus fruits. However, in recent years, agriculture is based on the production of a non-traditional fruit crops such as pitahaya which provides a relatively high profit margin to producers.
In 2020, the National Planning Secretary of Ecuador points out that 77% of the fruit produced in Palora is marketed nationally and exported to Hong Kong, Asia, the United States, Russia, the Netherlands, France, Germany and Spain which are the main consumers of pitahaya in the world [
6,
30] (Vargas, 2020). Therefore, this fruit crop has generated sources of employment improving the quality of life of at least 80% of the population (National Planning Secretary, 2020). Due the growth in the demand for this fruit and the fact that producers needed to organize, the “Association of Producers and Marketing of Pitahaya and other Palora products” was created, which is currently made up of 672 active members [
6] (Vargas, 2020).
2.2. Experimental Treatments
The experiment was organized in a randomized complete block design with three replications, the treatments being two agroforestry arrangements and monoculture as a control. For the agroforestry arrangements, multipurpose trees of
Erythrina poeppigiana and
Gliricidia sepium were used, each with the yellow pitahaya crop. Additionally, a shrub legume (
Flemingia macrophylla) was planted in the two agroforestry arrangements to provide biomass. The three species used in the agroforestry arrangements are considered to have high agronomic potential as nitrogen (N) fixers and soil improvers (structure) [
31]. Multipurpose trees have two functions: providing shade to the crop and the extraction of nutrients from deeper soil layers that are then deposited on the surface in the form of organic residues (biomass) resulting from frequent pruning of these trees [
32].
F. macrophylla was used because producers in the northern Amazon use it as a cover species to conserve and protect the soil [
33] and conserve macrofauna [
34].
2.3. Crop Management
At the beginning of the trial, the yellow pitahaya plants were two years old. They were planted at 3 m between rows and 2.5 m between plants, using inert concrete stakes 1.80 m high, two posts per plant with two lines of galvanized wire number 10 at 2.5 m. The multipurpose trees (E. poeppigiana and G. sepium) were transplanted 6 m between rows and 6 m between plants and the arbustiva species (F. macrophylla) were planted in double rows in the rows where the multipurpose species were located, with a spacing of 0.50 m between rows × 0.50 m between plants. The gross and net plot sizes were 15.0 × 25 m and 6.0 × 8.0 m, respectively. The net plot area was used to determine the crop yield.
Six months after planting, the basal shoots of the multipurpose trees were removed in order to leave only a single stem. In the first year, the multipurpose trees were pruned, eliminating lower branches and forming the canopy from 4 m [
35].
F. macrophylla were not pruned.
Shade species biomass cutting consisted of eliminating 60% of the aerial biomass. The amount of biomass incorporated from
E. poeppigiana ranged from 26 to 50 kg plant
−1 between the first year and the fifth year of biomass incorporation; while that of
G. sepium ranged from 29 to 10 kg plant
−1 between the first year and the fifth year. Biomass incorporation was carried out every 120 days (3 pruning per year). In contrast
F. macrophylla plants were pruned every 90 days (4 pruning per year) when they presented 50% flowering, adding 1 to 4 kg plant
−1 between the first year and the fifth year of biomass incorporation [
36,
37]; this pruning was done 20 cm above the ground. All organic matter was chopped and left on the soil surface next to the pitahaya plants, as recommended by [
38].
From the second year on, two sanitary pruning were carried out in the pitahaya crop in April and December; removing the unproductive branches, diseased stalks and intertwined branches, allowing good air circulation, reducing the weight of the plants on the training system and reducing the spread of pathogens [
6].
The amount of nutrients applied was calculated according to the procedure described by [
6], which considers the crop requirement, the contribution of nutrients from the soil according to its fertility (
Table 1), and the efficiency of the fertilizer.
The fertilizers used in this experiment were ammonium nitrate (38% N), di-ammonium phosphate (18% N, 48% P), magnesium sulfate (12% Mg, 20% S), and potassium chloride (60% K). A total of 369 to 177 g N plant−1, 322 to 120 g P plant−1, 480 to 292 g K plant−1 and 162 to 300 g S plant−1 were applied, the highest amount of these elements being applied in year one and decreasing in year five, respectively.
In the case of N, it was applied four times a year, 50% was applied in the vegetative growth stage (June and August), and another 50% in the reproductive stage (September and December). In the vegetative stage, 40% of P was applied and two fractions (30% each) were applied in the reproductive stage. K was applied in the vegetative stage 20% and two fractions in the reproductive stage (60% and 20%). Sulfur was applied 33% in the vegetative stage and 67% in the reproductive stage.
Phytosanitary controls were carried out every 15 days, using preventive and curative agrochemicals such as chlorothalonil, cymoxanil, metalaxyl and mancozeb, abamectin and lambdacyhalothrin. Weeding was carried out monthly with a brush cutter. Fruit was harvested manually with pruning shears when the fruit was at ripening stage 4 (
Figure 2).
2.4. Plant Sampling and Analysis
Fresh biomass of legumes. Three shade trees (
E. poeppigiana and
G. sepium) were taken from the net plot to quantify the biomass contribution of the pruning. In situ, the total pruning biomass (leaves and branches) was weighed in kg using a balance. To determine the amount of fresh biomass per hectare, the average biomass was determined and multiplied by the number of trees. To quantify the total biomass of pruned shrub legumes (
F. macrophylla), three rows (232 plants) were taken from the total plot, the average biomass per row was determined and multiplied by the number of rows per hectare [
32]. On the other hand, the biomass of the shade and shrub species was added to determine the total amount of biomass per hectare per year for each agroforestry system (treatments).
Composite samples (leaves and branches) of 250 g were taken from each pruning, deposited in paper bags duly identified and taken to the Soil and Water laboratory of the EECA for dry matter, N, P, K, Ca, Mg and S determination analysis.
Concentration of nutrients in the biomass. For the calculation of nutrient contents, the total biomass produced by each treatment was multiplied by the dry matter produced by each legume species and then the equation for macroelements suggested by [
36] was applied.
Q = Nutrient content in total dry matter (expressed in kg of nutrient ha−1)
MST = Total dry matter
X = Concentration of nutrient in dry matter
The nutrient content obtained in the treatments and replications was extrapolated to estimate the contribution of N, P, K, Ca, Mg and S in kg per hectare per year. For the determination of total N, the Semimicro Kjeldahl method was used, P was determined by the nitric-perchloric digestion extract colorimetric method, while K, Ca, Mg and S were determined by atomic absorption spectrometry [
39].
For the determination of total organic carbon of the system components (multipurpose legumes and shrubs), C present in the biomass was assumed to be 50% [
40,
41]. The C stocks obtained in each treatment were extrapolated to estimate the stocks per hectare.
Soil nutrient concentration. Once a year, composite samples of 1 kg (per treatment and repetition) were taken and deposited in plastic bags properly identified and taken to the Soil and Water laboratory of the EECA to carry out the analyses of organic C, total N, P, K, Ca, Mg and S. The determination of N was carried out with the Semimicro Kjeldahl method. The determination of P, K, Ca, Mg and S was carried out from an extract obtained by the modified Olsen method (pH 8.5) and P was determined by colorimetry, K, Ca and Mg by atomic absorption spectrometry and S by turbidimetry. C was determined by the Walkey and Black method [
39]. With the C and N values, the carbon/nitrogen ratio was determined.
Pitahaya fruit yield. Crop yield was determined in the net plot by weighing all fruits (g plant
−1). The number of fruits per plant and by size (size 20 = fruits ≤ 110 g, size 14 = 111 to 150 g, size 2 = 151 to 250 g, size 9 > 250 g) was counted [
42]. The yield obtained was extrapolated per hectare.
2.5. Data Analysis
2.5.1. Multivariate Statistics
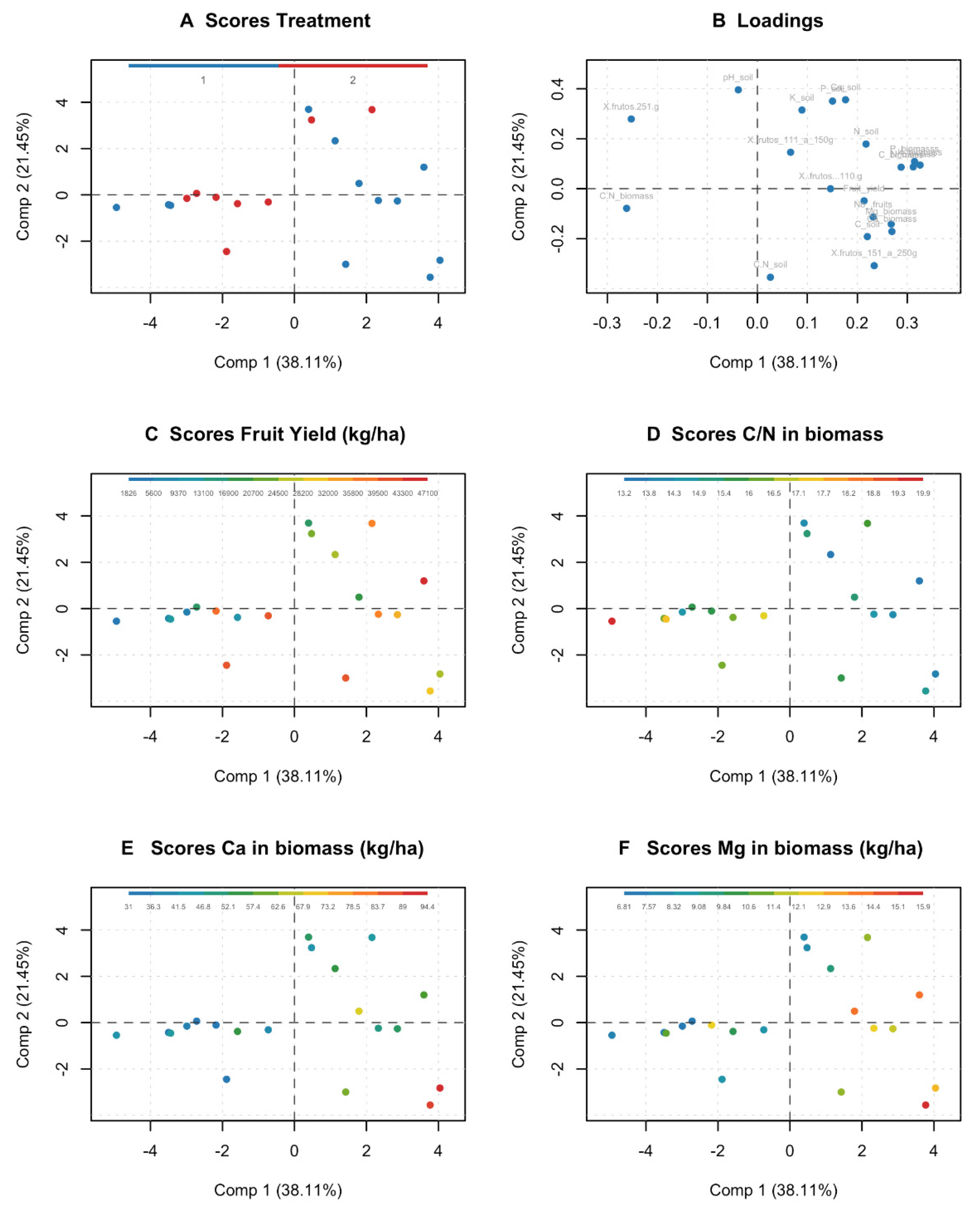
A multivariate analysis was performed using the R (3.2.4) program coupled with the MDA tools package (doi: 10.5281/zenodo.59547). A Principal component analysis model was used to identify the companion crops that affect pitahaya yield. The companion crops were: E. poeppigiana + F. macrophylla, G. sepium + F. macrophylla and monoculture was set as control. This experiment was carried out from 2016 to 2020. Scaling was performed because the variables were expressed in different units.
2.5.2. Response Analysis
The responses identified by PCA due to different years and complaining crops were modeled as:
4 Years = i = (2016, 2017, 2018, 2019, 2020) 3 blocks j = 1, 2, 3 3 Companion crops k = 1 to 3, where:
Yijkl = denotes the observation at the ith year, the jth block for the kth companion crop
μ = grand mean
Ai = random effect of the ith year
Bj = random effect of the jth block
Tk = fixed effect of the kth companion crop
ATik = interaction effect between the ith year and the kth companion crop
ε(ijk)l = Random experimental error (0, σe 2).
The data was analyzed using SAS 9.4 mixed model procedure. A repeated measures approach was used for successive sampling years, using a variance-covariance structure which was selected based on the lowest Akaike’s Information Criterion [
43].
The data was analyzed using SAS 9.3 mixed model procedure. Interactions and main effects were declared significant at
p < 0.1. Variables were checked for assumptions of normality and homogeneity of variances based on plot of residuals vs. predicted values. Transformations were performed as needed to comply with the normality assumption. The transformations were based on the Box-Cox power transformation series [
44]. Least square means were separated using Tukey mean separation (alpha = 0.1) procedure in SAS proc mixed and the means were order using mean separation into groups by letters [
45].
4. Discussion
Pitahaya yield increased from 1.89 t ha
−1 in 2016 to 25.06 t ha
−1 in 2020. The yield variation in the different years of evaluation is possibly due to the fact that the plant is reaching its full maturity (fruiting age), similar yields have been reported by [
46] in pitahaya monocultures where in the first two years a plant of pitahaya produces 1.0 t ha
−1 and between the fifth and sixth years, the yield stabilizes at 18 kg per plant.
In addition, it was determined that the highest yield was obtained in the two agroforestry systems with 12.89 t ha
−1 (
E. poeppigiana +
F. macrophylla) and 17.17 t ha
−1 (
G. sepium +
F. macrophylla) in relation to the monoculture (8.60 t ha
−1). Yields obtained in agroforestry arrangements differ from those reported by [
3] for sustainable pitahaya systems with live
Bursera simaruba tutors where in third year reached 20 t ha
−1 and 17 t ha
−1 for a monoculture.
This same favorable behavior for yield increase with the agroforestry systems, was obtained in an intercropped system of
Zea mays and
Sorghum sp. with
G. sepium, where yield was higher by 42 and 55%, respectively, compared to the monoculture; in addition the yield stabilized over time as observed in our study with pitahaya [
47]. This trend has also been reported in agroforestry systems of coffee with legume shade (
G. sepium +
Leucaena leucocephala +
E. variegata), where yields increased by 20% [
46,
48] also points out that it is very difficult to estimate fruit yield of pitahaya because it depends a lot on the age of the crop, agronomic management, production systems, planting distance, climate and sexual incompatibility.
In 2016 (first year), the number of fruits per plant were 9 and 11 in the agroforestry systems
E. poeppigiana +
F. macrophylla and
G. sepium +
F. macrophylla, respectively, and 2 in the monoculture, While in 2020, it increased to 106 in the
E. poeppigiana +
F. macrophylla system and 56 in the
G. sepium +
F. macrophylla system but it was lower in the monoculture (38). The relationship between number of fruits and yield is mainly due to the fact that this variable is an important component of yield [
49].
In contrast the soluble solids content of the pitahaya fruits from the agroforestry systems and the monoculture was determined in 2020. A value of 21.1° Brix was obtained in the
E. poeppigiana +
F. macrophylla system, whereas the
G. sepium +
F. macrophylla system reached 21.4° Brix; on the another hand, the monoculture got 22° Brix; these values are similar among them thus this variable does not indicate any significant difference. These results were relatively higher than the values reported by [
1], who mentions that the yellow pitahaya harvested at maturity stage 6 in the Palora site was 20.74° Brix.
In agroforestry systems, the strong relationship between yield and nutrients provided by biomass (Ca and Mg) is possibly due to the fact that these elements are absorbed by the deep roots of the trees and deposited through the litter on the most superficial soils, that is, the effect of nutrient pumping occurs [
50]. This direct supply of nutrients from decomposition processes and mulching effects improve crop yield [
51].
Calcium (>39 kg ha
−1) and Mg (>10 kg ha
−1) contained in the biomass of the legumes species used in the agroforestry systems are directly related to crop yield. This is possibly due to the fact that these species store high concentrations of N, P, K, Ca and Mg in their leaves; which can be supported by [
52], who reports that
Tithonia diversifolia accumulates high concentrations of nutrients in its leaves and that the moment they are incorporated into the soil, they improve the nutritional status through the mobilization and return of nutrients, as well as to the contribution of organic matter [
53]. Ca and Mg are the most important mineral nutrients in production [
54,
55]. A higher yield, larger fruit size, higher percentage of marketable fruit [
54] and better shelf life in crops are achieved when Ca is present in crop nutrition [
55]. According to the results, Mg influenced it to improve pitahaya yield in agroforestry systems by 29% with respect to monoculture [
56] who points out that Mg improves fruit yield by 12.5% when fruit crops grown in acidic soils that was the case of this study (Palora soil pH 5.2 to 5.6).
In addition, it was determined that the Ca and Mg content varied from one year to another and that the highest contribution was achieved with the agroforestry system
E. poeppigiana +
F. macrophylla which contributed with amount of biomass of 19,753 kg ha
−1, while the
G. sepium +
F. macrophylla system contributed with 6700 kg ha
−1, consequently the nutrient content basically depends on the amount of biomass produced by the leguminous species for the system. This result is corroborated by [
51] who mentions that the amount of nutrients that is incorporated depends on the amount of biomass that is produced with pruning. For example, [
57] in an agroforestry system study, determined that although
G. sepium contains the highest concentration of nutrients in its leaves, it did not produce abundant biomass like
L. leucocephala, causing this legume to be considered the best source of nutrients because it incorporated 144 kg ha
−1 of Ca and 60 kg ha
−1 of Mg, amounts that would meet the nutritional needs of various crops.
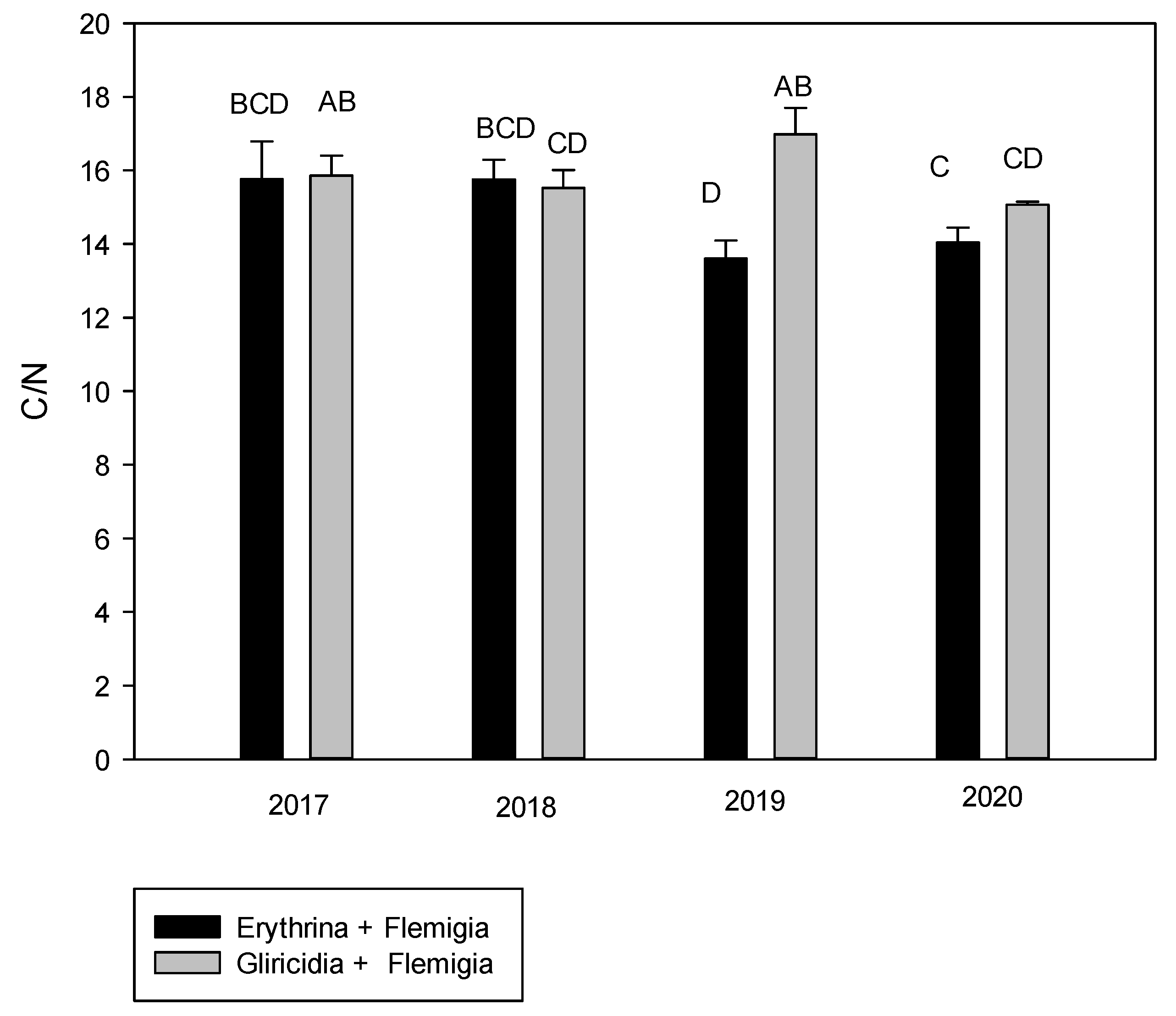
The association of yield with the C:N ratio obtained in this study is possibly due to the fact that legumes produce a lower C:N ratio than tree and grass species in agroforestry systems [
58], which would favor the release of N to the soil, an effect that is reflected in the crop yield. This is corroborated by [
59], who mentions that legume species contribute to natural regeneration due to their association with N-fixing bacteria, helping to increase the yield and fertility of agroforestry soils by producing a high quality leaf litter. Ref. [
60] indicated that an agroforestry system of
Calliandra calothyrsus and
Sesbania sesban with
Zea mays, these species contributed more N and competed favorably with associated crops than when agricultural crops were with non-legume species.
It was determined that the greater contribution of biomass in the agroforestry system of
E. poeppigiana +
F. macrophylla increased the yield when the C:N ratio was around 15. It was supported by [
61], who points out that when the C:N ratio is <20 or 30 there is a mineralization of N and when the ratio is >30 there is a immobilization of N. In addition, it was determined that the C:N ratio in the years of evaluation was decreasing and the lowest ratio was achieved with the
E. poeppigiana +
F. macrophylla system in 2020. Ref. [
58] points out that the decomposition rate should not be determined only by the C:N ratio but also by other characteristics of the plant such as chemical composition, size, distribution, branching of the shoots and roots, as well as the texture of the soil, temporal variations of the gas content and water with time and the atmosphere (evapotranspiration rates for each type of plant over), thus the activity of a series of microorganisms in the soil [
55]. Therefore, the presence of legumes in agroforestry systems improves the quality of litter, due to the fact that their residues rich in N improve the efficiency of the use of C by part of the decomposing microorganisms, which results in a higher microbial biomass [
62] and the eventual stabilization of soil organic C [
59].
In previous studies in coffee agroforestry systems with
E. poeppigiana, it was determined that 50% N is fixed by this legume and it is a species that is recommended to be able to substitute the use of nitrogen fertilizers [
63]. In addition, it is indicated that the mineralization processes of P, K, Ca and Mg of
E. poeppigiana leaves are faster than in other species such as
Inga edulis or
Cajanus cajan, approximately 40% of the initial P and Ca contents and the 75% of the Mg and K content of the leaves of
E. poeppigiana are mineralized in four weeks [
64] which is advantageous for the nutrition of a crop in association with this species.
Finally, this kind of sustainable production systems will strengthen the pitahaya production to encourage integrated fruit production in transition to organic fruit production which can be mainly for exportation [
65]. On the other hand, in the Ecuadorian Amazon, these sustainable agriculture alternatives (agroforestry systems) help improve soil health by reducing erosion rates by 50%, increasing the infiltration rate by 75%, reducing runoff by 35%, increasing soil organic C by 21%, storing organic N by 13% and available N by 46%, and phosphorous is available by 11%. In addition, these systems contribute to reduce the use of external inputs (fertilizers) because the associated species incorporate nutrients into the system, improve crop profitability because the income comes from different sources, guarantee food security because 40% of the production is used for family nutrition, conservation and local use of the agrobiodiversity and provide ecosystem services (regulation of hydrological cycles, recovery of degraded and contaminated areas) [
30] The legumes present in agroforestry systems improve crop yield, reduce production costs by 30% due to the reduction of the use of external inputs such as synthetic fertilizers and improve producers’ income.


 ,
, 






{kind=link}
{kind=link}
{kind=link}
{kind=link}