
Do Cross-Latitude and Local Studies Give Similar Predictions of Phytoplankton Responses to Warming? An Analysis of Monitoring Data from 504 Danish Lakes


Abstract
:1. Introduction
2. Research Methods and Data
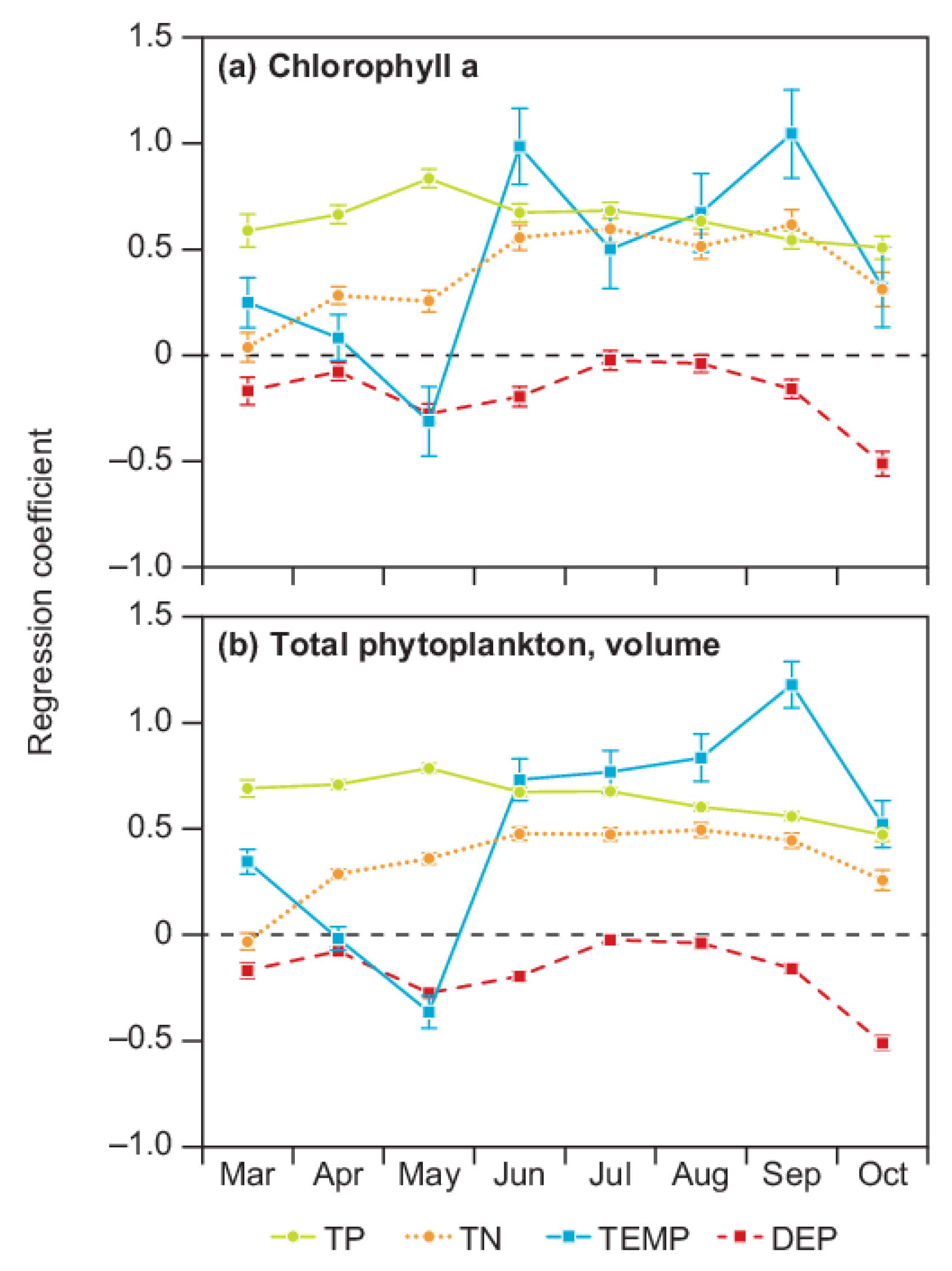
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| March | April | May | June | July | August | September | October | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Int/ Regression Coefficient | SE | p | Int/ Regression Coefficient | SE | p | Int/ Regression Coefficient | SE | p | Int/ Regression Coefficient | SE | p | Regression Coefficient | SE | p | Regression Coefficient | SE | p | Regression Coefficient | SE | p | Regression Coefficient | SE | p | |
| Cyanobacteria (mm3 L−1) | ||||||||||||||||||||||||
| INT | 0.529 | 0.09 | **** | 0.454 | 0.126 | *** | 0.048 | 0.24 | ns | 0.037 | 0.416 | ns | −0.284 | 0.553 | ns | −1.35 | 0.567 | ** | −1.296 | 0.525 | * | 0.354 | 0.31 | |
| TP | 0.162 | 0.024 | **** | 0.193 | 0.021 | **** | 0.219 | 0.023 | **** | 0.288 | 0.032 | **** | 0.328 | 0.036 | **** | 0.282 | 0.034 | **** | 0.218 | 0.037 | **** | 0.195 | 0.035 | **** |
| TN | −0.03 | 0.021 | * | −0.0487 | 0.02 | * | −0.025 | 0.026 | ns | 0.03 | 0.046 | ns | 0.292 | 0.062 | **** | 0.507 | 0.058 | **** | 0.541 | 0.062 | **** | 0.148 | 0.053 | ** |
| TEMP | 0.061 | 0.037 | ns | 0.142 | 0.05 | ** | 0.313 | 0.084 | ** | 0.453 | 0.139 | ** | 0.651 | 0.178 | *** | 1.136 | 0.185 | **** | 1.04 | 0.184 | **** | 0.323 | 0.123 | ** |
| DEP | −0.015 | 0.02 | ns | −0.005 | 0.019 | Ns | 0.01 | 0.024 | ns | 0.003 | 0.036 | ns | 0.008 | 0.043 | ns | 0.021 | 0.04 | ns | −0.111 | 0.039 | ** | −0.091 | 0.039 | * |
| Cyanobacteria (%) | ||||||||||||||||||||||||
| INT | 1.22 | 0.24 | **** | 0.704 | 0.304 | * | −0.177 | 0.548 | ns | 0.453 | 0.76 | ns | −1.06 | 0.84 | ns | −2.394 | 0.783 | ** | −0.64 | 0.785 | ns | 1.066 | 0.608 | ns |
| TP | 0.219 | 0.063 | *** | 0.278 | 0.05 | **** | 0.39 | 0.052 | **** | 0.442 | 0.059 | **** | 0.298 | 0.055 | **** | 0.191 | 0.047 | **** | 0.162 | 0.056 | ** | 0.211 | 0.069 | ** |
| TN | −0.208 | 0.058 | *** | −0.178 | 0.047 | *** | −0.132 | 0.059 | * | −0.143 | 0.084 | ns | 0.31 | 0.093 | *** | 0.658 | 0.081 | **** | 0.711 | 0.093 | **** | 392 | 0.103 | *** |
| TEMP | 0.093 | 0.098 | ns | 0.421 | 0.122 | *** | 0.877 | 0.192 | **** | 0.762 | 0.254 | ** | 1.265 | 0.271 | **** | 1.696 | 0.255 | **** | 1.134 | 0.274 | **** | 0.366 | 0.241 | ns |
| DEP | 0.065 | 0.054 | ns | −0.052 | 0.047 | Ns | 0.058 | 0.054 | ns | 0.236 | 0.066 | *** | 0.301 | 0.066 | **** | 0.348 | 0.056 | **** | 0.303 | 0.058 | **** | 0.469 | 0.073 | **** |
| Dinophytes (mm3 L−1) | ||||||||||||||||||||||||
| INT | 0.36 | 0.08 | **** | 0.23 | 0.06 | *** | −0.01 | 0.08 | ns | −0.60 | 0.20 | ** | −0.78 | 0.36 | * | −0.95 | 0.41 | * | −1.51 | 0.34 | **** | −0.01 | 0.12 | ns |
| TP | 0.02 | 0.02 | ns | 0.01 | 0.01 | Ns | −0.01 | 0.01 | ns | −0.06 | 0.02 | **** | 0.02 | 0.02 | ns | 0.09 | 0.02 | *** | 0.09 | 0.02 | *** | 0.01 | 0.01 | ns |
| TN | −0.04 | 0.02 | ** | −0.03 | 0.01 | ** | −0.02 | 0.01 | * | 0.06 | 0.02 | ** | −0.01 | 0.04 | ns | −0.05 | 0.04 | ns | −0.12 | 0.04 | ** | −0.03 | 0.02 | ns |
| TEMP | −0.05 | 0.03 | ns | −0.05 | 0.03 | Ns | 0.01 | 0.03 | ns | 0.17 | 0.07 | ** | 0.31 | 0.11 | ** | 0.44 | 0.13 | *** | 0.67 | 0.12 | **** | 0.05 | 0.05 | ns |
| DEP | −0.09 | 0.02 | **** | −0.02 | 0.01 | * | 0.02 | 0.01 | * | 0.18 | 0.02 | **** | 0.38 | 0.03 | **** | 0.42 | 0.03 | **** | 0.28 | 0.03 | **** | 0.02 | 0.01 | ns |
| Dinophytes (%) | ||||||||||||||||||||||||
| INT | 0.99 | 0.25 | **** | 1.035 | 0.267 | **** | −0.239 | 0.446 | ns | −0.738 | 0.632 | ns | 0.665 | 0.736 | ns | −0.295 | 0.732 | ns | −2 | 0.681 | ** | −0.972 | 0.377 | * |
| TP | −0.165 | 0.067 | * | −0.171 | 0.044 | **** | −0.325 | 0.042 | **** | −0.484 | 0.0491 | **** | −0.321 | 0.048 | **** | −0.195 | 0.043 | **** | −0.088 | 0.0483 | ns | −0.143 | 0.042 | *** |
| TN | −0.138 | 0.06 | * | −0.129 | 0.041 | ** | −0.119 | 0.048 | * | −0.009 | 0.0699 | ns | −0.319 | 0.081 | **** | −0.229 | 0.075 | ** | −0.405 | 0.08 | **** | −0.195 | 0.064 | ** |
| TEMP | −0.214 | 0.102 | * | −0.362 | 0.107 | *** | −0.0258 | 0.156 | ns | 0.062 | 0.211 | ns | −0.192 | 0.237 | ns | 0.246 | 0.239 | ns | 0.908 | 0.238 | **** | 0.479 | 0.149 | ** |
| DEP | −0.367 | 0.056 | **** | −0.069 | 0.041 | Ns | 0.213 | 0.044 | **** | 0.663 | 0.054 | **** | 0.831 | 0.057 | **** | 0.791 | 0.052 | **** | 0.699 | 0.05 | **** | 0.117 | 0.045 | ** |
| Diatoms (mm3 L−1) | ||||||||||||||||||||||||
| INT | 0.423 | 0.197 | * | 1.385 | 0.213 | **** | 2.32 | 0.305 | **** | 0.449 | 0.341 | ns | 2.589 | 0.428 | **** | 2.913 | 0.395 | **** | 1.922 | 0.375 | **** | 1.322 | 0.265 | **** |
| TP | 0.09 | 0.05 | ns | 0.22 | 0.04 | **** | 0.19 | 0.03 | **** | 0.14 | 0.03 | **** | 0.18 | 0.03 | **** | 0.21 | 0.02 | **** | 0.19 | 0.03 | **** | 0.20 | 0.03 | **** |
| TN | 0.25 | 0.05 | **** | 0.32 | 0.03 | **** | 0.20 | 0.03 | **** | 0.15 | 0.04 | **** | 0.10 | 0.05 | * | −0.01 | 0.04 | ns | 0.03 | 0.04 | ns | 0.04 | 0.05 | ns |
| TEMP | 0.26 | 0.08 | *** | −0.06 | 0.09 | Ns | −0.45 | 0.11 | **** | 0.16 | 0.11 | ss | −0.50 | 0.14 | *** | −0.56 | 0.13 | **** | −0.26 | 0.13 | * | −0.01 | 0.11 | ns |
| DEP | 0.11 | 0.04 | * | 0.14 | 0.03 | **** | −0.14 | 0.03 | **** | −0.08 | 0.03 | ** | −0.14 | 0.03 | **** | −0.26 | 0.03 | **** | −0.22 | 0.03 | **** | −0.25 | 0.03 | **** |
| Chlorophytes (mm3 L−1) | ||||||||||||||||||||||||
| INT | 0.11 | 0.13 | ns | 0.14 | 0.13 | Ns | 1.38 | 0.23 | **** | 1.41 | 0.30 | **** | 1.85 | 0.35 | **** | 1.90 | 0.32 | **** | 1.77 | 0.30 | **** | 1.32 | 0.21 | **** |
| TP | 0.39 | 0.03 | **** | 0.37 | 0.02 | **** | 0.42 | 0.02 | **** | 0.38 | 0.02 | **** | 0.28 | 0.02 | **** | 0.20 | 0.02 | **** | 0.17 | 0.02 | **** | 0.20 | 0.02 | **** |
| TN | −0.11 | 0.03 | **** | −0.05 | 0.02 | * | 0.06 | 0.03 | * | 0.17 | 0.03 | **** | 0.23 | 0.04 | **** | 0.15 | 0.03 | **** | 0.15 | 0.04 | **** | 0.04 | 0.04 | ns |
| TEMP | 0.01 | 0.05 | ns | 0.06 | 0.05 | Ns | 0.15 | 0.08 | ns | 0.15 | 0.10 | ns | −0.13 | 0.11 | ns | −0.24 | 0.10 | * | −0.24 | 0.10 | * | −0.08 | 0.08 | ns |
| DEP | −0.12 | 0.02 | **** | −0.11 | 0.02 | **** | −0.19 | 0.02 | **** | −0.30 | 0.03 | **** | −0.31 | 0.03 | **** | −0.31 | 0.02 | **** | −0.36 | 0.02 | **** | −0.33 | 0.02 | **** |
References
- IPCC. Climate Change: The Physical Science Basis, Summary f1 1or Policymakers. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. 2007. Available online: http://www.ipcc.ch (accessed on 1 November 2021).
- IPCC. The Physical Science Basis: Working Group 464 I Contribution to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013. [Google Scholar]
- Huisman, J.; Matthijs, H.C.P.; Visser, P.M. Harmful Cyanobacteria; Springer: Dordrecht, The Netherlands, 2005. [Google Scholar]
- Jöhnk, K.D.; Huisman, J.; Sharples, J.; Sommeijer, B.; Visser, P.M.; Stroom, J.M. Summer heatwaves promote blooms of harmful cyanobacteria. Glob. Change Biol. 2008, 14, 495–512. [Google Scholar] [CrossRef] [Green Version]
- Jeppesen, E.; Kronvang, B.; Meerhoff, M.; Søndergaard, M.; Hansen, K.M.; Andersen, H.E.; Lauridsen, T.L.; Liboriussen, L.; Beklioglu, M.; Özen, A.; et al. Climate change effects on runoff, catchment phosphorus loading and lake ecological state; potential adaptations. J. Environ. Qual. 2009, 38, 1930–1941. [Google Scholar] [CrossRef]
- Kosten, S.; Huszar, V.L.M.; Bécares, E.; Costa, L.S.; van Donk, E.; Hansson, L.-A.; Jeppesen, E.; Kruk, C.; Lacerot, G.; Mazzeo, N.; et al. Warmer climate boosts cyanobacterial dominance in lakes. Glob. Change Biol. 2012, 18, 118–126. [Google Scholar] [CrossRef]
- Kakouei, K.; Kraemer, B.M.; Anneville, O.; Carvalho, L.; Feuchtmayr, H.; Graham, J.L.; Higgins, S.; Pomati, F.; Rudstam, L.G.; Stockwell, J.D.; et al. Phytoplankton and cyanobacteria abundances in mid-21st century lakes depend strongly on future land use and climate projections. Glob. Change Biol. 2021, 27, 6409–6422. [Google Scholar] [CrossRef]
- Meerhoff, M.; Teixeira-de Mello, F.; Kruk, C.; Alonso, C.; González-Bergonzoni, I.; Pacheco, J.P.; Lacerot, G.; Arim, M.; Beklioğlu, M.; Brucet, S.; et al. Environmental warming in shallow lakes: A review of potential changes in community structure as evidenced from space-for-time-substitution approaches. Adv. Ecol. Res. 2012, 46, 259–349. [Google Scholar]
- Stockenreiter, M.; Navarro, J.I.; Buchberger, F.; Stibor, H. Community shifts from eukaryote to cyanobacteria dominated phytoplankton: The role of mixing depth and light quality. Freshwat. Biol. 2021, 66, 2145–2157. [Google Scholar] [CrossRef]
- Carey, C.C.; Ibelings, B.W.; Hoffmann, E.P.; Hamilton, D.P.; Brookes, J.D. Eco-physiological adaptations that favour freshwater cyanobacteria in a changing climate. Wat. Res. 2021, 46, 1394–1407. [Google Scholar] [CrossRef]
- Deng, J.; Paerl, H.W.; Qin, B.; Zhang, Y.; Jeppesen, E.; Cai, Y.; Xu, H. Climatically-modulated decline in wind speed may strongly affect eutrophication in shallow lakes. Sci. Tot. Environ. 2018, 645, 1361–1370. [Google Scholar] [CrossRef]
- Richardson, J.; Miller, C.; Maberly, S.C.; Taylor, P.; Globevnik, L.; Hunter, P.; Jeppesen, E.; Mischke, E.; Moe, J.; Pasztaleniec, A.; et al. Effects of multiple stressors on cyanobacteria biovolume varies with lake type. Glob. Change Biol. 2018, 24, 5044–5055. [Google Scholar] [CrossRef] [Green Version]
- Jeppesen, E.; Canfield, D.E., Jr.; Bachmann, R.W.; Søndergaard, M.; Havens, K.E.; Johansson, L.S.; Lauridsen, T.L.; Sh, T.; Rutter, R.P.; Warren, G.; et al. Towards predicting climate change effects on lakes: A comparative study of 1656 shallow lakes from subtropical Florida and temperate Denmark reveals substantial differences in nutrient dynamics, metabolism, trophic structure and top-down control. Inl. Wat. 2020, 10, 197–211. [Google Scholar] [CrossRef]
- Jeppesen, E.; Mehner, T.; Winfield, I.J.; Kangur, K.; Sarvala, J.; Gerdeaux, D.; Rask, M.; Malmquist, H.J.; Holmgren, K.; Volta, P.; et al. Impacts of climate warming on lake fish assemblages: Evidence from 24 European long term data series. Hydrobiologia 2012, 694, 1–39. [Google Scholar] [CrossRef] [Green Version]
- Jeppesen, E.; Søndergaard, M.; Lauridsen, T.L.; Liboriussen, L.; Bjerring, R.; Johansson, L.S.; Landkildehus, F.; Kronvang, B.; Andersen, H.E.; Trolle, D.; et al. Recent climate induced changes in freshwaters in Denmark. In Climatic Change and Global Warming of Inland Waters: Impacts and Mitigation for Ecosystems and Societies; Goldman, C.R., Kumagari, M., Robarts, R.D., Eds.; John Wiley & Son Ltd.: Hoboken, NJ, USA, 2012; pp. 156–171. [Google Scholar]
- Teixeira-de Mello, F.; Meerhoff, M.; Pekcan-Hekim, Z.; Jeppesen, E. Substantial differences in littoral fish community structure and dynamics in subtropical and temperate shallow lakes. Freshwat. Biol. 2009, 54, 1202–1215. [Google Scholar] [CrossRef]
- Brucet, S.; Pédron, S.; Lauridsen, T.L.; Mehner, T.; Argillier, C.; Winfield, I.J.; Volta, P.; Emmrich, M.; Holmgreen, K.; Rask, M.; et al. Fish community structure in European lakes: The role of eutrophication, climate and hydromorphology alterations. Freshwat. Biol. 2013, 58, 1779–1793. [Google Scholar] [CrossRef] [Green Version]
- Moss, B. Climate change, nutrient pollution and the bargain of Dr Faustus. Freshwat. Biol. 2010, 55, 175–187. [Google Scholar] [CrossRef]
- Gonzalez-Bergonzoni, I.; Meerhoff, M.; Davidson, T.A.; Teixeira-de Mello, F.; Baattrup-Pedersen, A.; Jeppesen, E. Meta-analysis shows a consistent and strong latitudinal pattern in fish omnivory across ecosystems. Ecosystems 2012, 15, 492–503. [Google Scholar] [CrossRef]
- Iglesias, C.; Mazzeo, N.; Meerhoff, M.; Lacerot, G.; Clemente, J.; Scasso, F.; Kruk, C.; Goyenola, G.; Garcia, J.; Amsinck, S.L.; et al. High predation is the key factor for dominance of small-bodied zooplankton in warm lakes—Evidence from lakes, fish exclosures and surface sediment. Hydrobiologia 2011, 667, 133–147. [Google Scholar] [CrossRef]
- Gyllström, M.; Hansson, L.A.; Jeppesen, E.; García-Criado, F.; Gross, E.; Irvine, K.; Kairesalo, T.; Kornijow, R.; Miracle, M.R.; Nykänen, M.; et al. The role of climate in shaping zooplankton communities of shallow lakes. Limnol. Oceanogr. 2005, 50, 2008–2021. [Google Scholar] [CrossRef] [Green Version]
- Havens, K.E.; Beaver, J.R. Zooplankton to phytoplankton bbiomass ratios in shallow Florida lakes: An evaluation of seasonality and hypotheses about factors controlling variability. Hydrobiologia 2013, 703, 177–187. [Google Scholar] [CrossRef]
- Vidal, N.; Amsinck, S.L.; Gonçalves, V.; Azevedo, J.M.N.; Johansson, L.S.; Christoffersen, K.S.; Lauridsen, T.L.; Søndergaard, M.; Bjerring, R.; Landkildehus, F.; et al. Food web patterns in species-poor insular lakes resemble climate-related patterns in continental lakes. Water 2021, 13, 1380. [Google Scholar] [CrossRef]
- Trolle, D.; Hamilton, D.P.; Pilditch, C.A.; Duggan, I.C.; Jeppesen, E. Predicting the effects of climate change on trophic status of three morphologically varying lakes: Implications for lake restoration and management. Environ. Model. Softw. 2011, 26, 354–370. [Google Scholar] [CrossRef]
- Chen, W.; Nielsen, A.; Kuhlmann, T.; Hu, F.; Chou, Q.; Søndergaard, M.; Jeppesen, E.; Trolle, D. Modelling the ecological response of a temporarily summer-stratified lake to extreme heatwaves. Water 2020, 12, 94. [Google Scholar] [CrossRef] [Green Version]
- Feuchtmayr, H.; Moran, R.; Hatton, K.; Connor, L.; Heyes, T.; Moss, B.; Harvey, I.; Atkinson, D. Global warming and eutrophication: Effects on water chemistry and autotrophic communities in experimental hypertrophic shallow lake mesocosms. J. Appl. Ecol. 2009, 46, 713–723. [Google Scholar] [CrossRef]
- Kratina, P.; Greig, H.S.; Thompson, P.L.; Carvalho-Pereira, T.S.; Shurin, J.B. Warming modifies trophic cascades and eutrophication in experimental freshwater communities. Ecology 2012, 93, 1421–1430. [Google Scholar] [CrossRef] [PubMed]
- Hansson, L.-A.; Nicolle, A.; Granéli, W.; Hallgren, P.; Kritzberg, E.; Persson, A.; Björk, J.; Nilsson, P.A.; Brönmark, C. Food-chain length alters community responses to global change in aquatic systems. Nat. Clim. Change 2013, 3, 228–233. [Google Scholar] [CrossRef]
- Svendsen, L.M.; van der Bijl, L.; Boutrup, S.; Norup, B. (Eds.) NOVANA. National Monitoring and Assessment Programme for the Aquatic and Terrestrial Environments. Programme Description, Part 2; NERI Technical Report No. 537; National Environmental Research Institute: Aarhus, Denmark, 2005. [Google Scholar]
- Johansson, L.S.; Lauridsen, T.L. Feltmålinger, Profilmålinger Samt Udtagning af Prøver til Analyse af Vandkemiske Parametre i Søer—DCE National Cor Miljø og Energi. 2017. Available online: https://ecos.au.dk/forskningraadgivning/fagdatacentre/ferskvand (accessed on 1 November 2021).
- Olrik, K. Planteplankton Metoder—Prøvetagning, Bearbejdning og Rapportering ved Undersøgelser af Planteplankton i søer og Marine områder [Phytoplankton Methods—Sampling, Processing and Reporting at Phytoplankton Investigations in Lakes and Marine Environments]. Miljøprojekt nr. 187; Miljøministeriet/Miljøstyrelsen: Copenhagen, Denmark, 1991; ISBN 87-503-9411-8. 108p. (In Danish) [Google Scholar]
- Romo, S.; Villena, M.J.; Sahuquillo, M.; Soria, J.-M.; Giménez, M.; Alfonso, T.; Vicente, E.; Miracle, M.R. Response of a shallow Mediterranean lake to nutrient diversion: Does it follow similar patterns as in northern shallow lakes? Freshwat. Biol. 2005, 50, 1706–1717. [Google Scholar] [CrossRef]
- Blenckner, T.; Adrian, R.; Livingstone, D.M.; Jennings, E.; Weyhenmeyer, G.A.; George, D.G.; Jankowski, T.; Jarvinen, M.; Aonghusa, C.N.; Noges, T.; et al. Large-scale climatic signatures in lakes across Europe: A meta-analysis. Glob. Change Biol. 2007, 13, 1314–1326. [Google Scholar] [CrossRef] [Green Version]
- Jeppesen, E.; Moss, B.; Bennion, H.; Carvalho, L.; DeMeester, L.; Feuchtmayr, H.; Friberg, N.; Gessner, M.O.; Hefting, M.; Lauridsen, T.L.; et al. Interaction of climate and eutrophication. In Changing Climate and Changing Freshwaters: A European Perspective; Kernan, M., Battarbee, R., Moss, B., Eds.; Blackwell: Oxford, UK, 2010; pp. 119–151. [Google Scholar]
- Søndergaard, M.; Lauridsen, T.L.; Johansson, L.S.; Jeppesen, E. Nitrogen or phosphorus limitation in lakes and its impact on phytoplankton bbiomass and submerged macrophyte covers. Hydrobiologia 2017, 795, 35–48. [Google Scholar] [CrossRef]
- Jeppesen, E.; Jensen, J.P.; Søndergaard, M.; Lauridsen, T.L.; Pedersen, L.J.; Jensen, L. Top-down control in freshwater lakes: The role of nutrient state, submerged macrophytes and water depth. Hydrobiologia 1997, 342/343, 151–164. [Google Scholar] [CrossRef]
- Salk, K.R.; Venkiteswaran, J.J.; Couture, R.M.; Higgins, S.N.; Paterson, M.J.; Schiff, S.L. Warming combined with experimental eutrophication intensifies lake phytoplankton blooms. Limnol. Oceanogr. 2021. [Google Scholar] [CrossRef]
- Markensten, H.; Moore, K.; Persson, I. Simulated lake phytoplankton composition shifts toward cyanobacteria dominance in a future warmer climate. Ecol. Appl. 2010, 20, 752–767. [Google Scholar] [CrossRef] [PubMed]
- Bertani, I.; Steger, C.E.; Obenour, D.R.; Fahnenstiel, G.L.; Bridgeman, T.B.; Johengen, T.H.; Sayers, M.J.; Schuman, R.A.; Scavia, D. Tracking cyanobacteria blooms: Do different monitoring approaches tell the same story? Sci. Tot. Environ. 2017, 575, 294–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reynolds, C.S. The Ecology of Phytoplankton; Cambridge University Press: Cambridge, UK, 2006. [Google Scholar]
- Lürling, M.; Eshetu, F.; Faassen, E.J.; Kosten, S.; Huszar, V.L.M. Comparison of cyanobacterial and green algal growth rates at different temperatures. Freshwat. Biol. 2013, 58, 552–559. [Google Scholar] [CrossRef]
- Paerl, H.W.; Huisman, J. Climate change: A catalyst for global expansion of harmful cyanobacterial blooms. Environ. Microbiol. Reps. 2009, 1, 27–37. [Google Scholar] [CrossRef] [PubMed]








| Mean | Minimum | Maximum | |
|---|---|---|---|
| Total phosphorus (mg L−1) | 0.16 | 0.004 | 3.84 |
| Total nitrogen (mg L−1) | 1.83 | 0.26 | 8.10 |
| Chlorophyll a (µg L−1) | 45 | 1 | 266 |
| Mean depth (m) | 2.4 | 0.2 | 16.4 |
| Lake area (km2) | 2.0 | 0.05 | 39.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeppesen, E.; Johansson, L.S.; Tserenpil, S.; Søndergaard, M.; Lauridsen, T.L.; Andersen, P. Do Cross-Latitude and Local Studies Give Similar Predictions of Phytoplankton Responses to Warming? An Analysis of Monitoring Data from 504 Danish Lakes. Sustainability 2021, 13, 14049. https://doi.org/10.3390/su132414049
Jeppesen E, Johansson LS, Tserenpil S, Søndergaard M, Lauridsen TL, Andersen P. Do Cross-Latitude and Local Studies Give Similar Predictions of Phytoplankton Responses to Warming? An Analysis of Monitoring Data from 504 Danish Lakes. Sustainability. 2021; 13(24):14049. https://doi.org/10.3390/su132414049
Chicago/Turabian StyleJeppesen, Erik, Liselotte S. Johansson, Sh Tserenpil, Martin Søndergaard, Torben L. Lauridsen, and Per Andersen. 2021. "Do Cross-Latitude and Local Studies Give Similar Predictions of Phytoplankton Responses to Warming? An Analysis of Monitoring Data from 504 Danish Lakes" Sustainability 13, no. 24: 14049. https://doi.org/10.3390/su132414049
APA StyleJeppesen, E., Johansson, L. S., Tserenpil, S., Søndergaard, M., Lauridsen, T. L., & Andersen, P. (2021). Do Cross-Latitude and Local Studies Give Similar Predictions of Phytoplankton Responses to Warming? An Analysis of Monitoring Data from 504 Danish Lakes. Sustainability, 13(24), 14049. https://doi.org/10.3390/su132414049