
Long-Term Influence of Stump-Removal on Components of Hemiboreal Pine Forest Ecosystem

 ,
,  ,
, 
Abstract
:1. Introduction
2. Methods and Materials
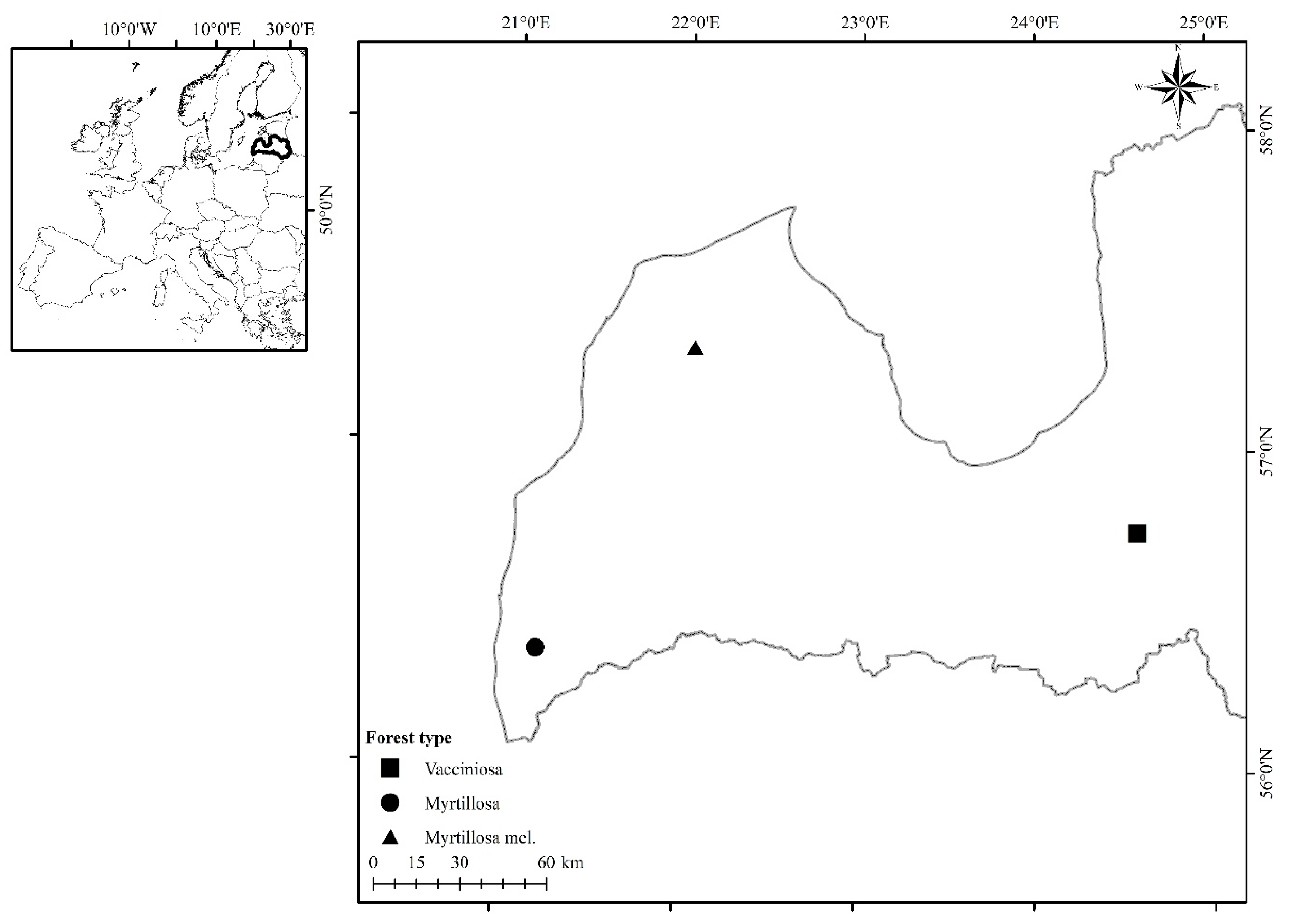
2.1. Study Site
2.2. Data Analysis
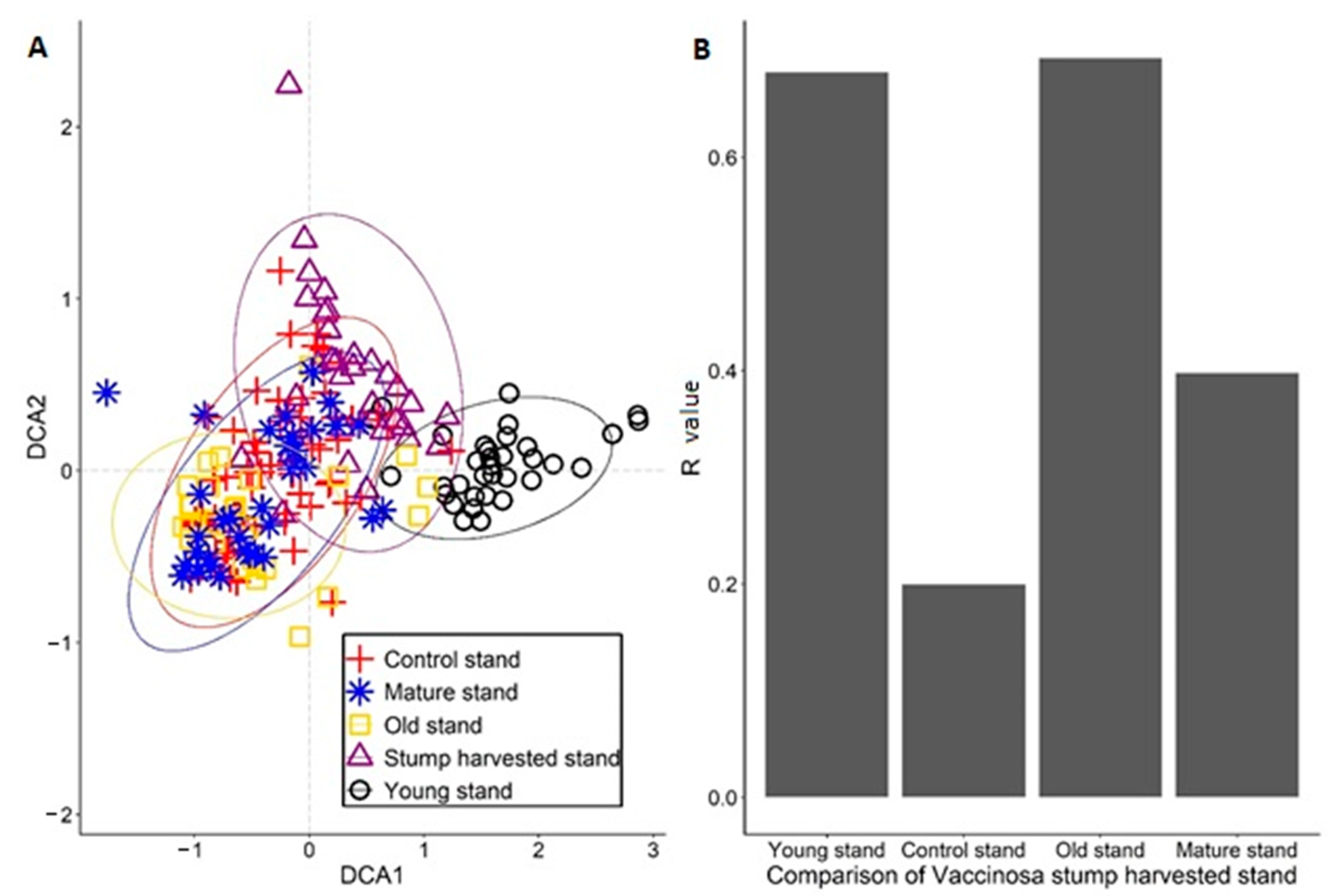
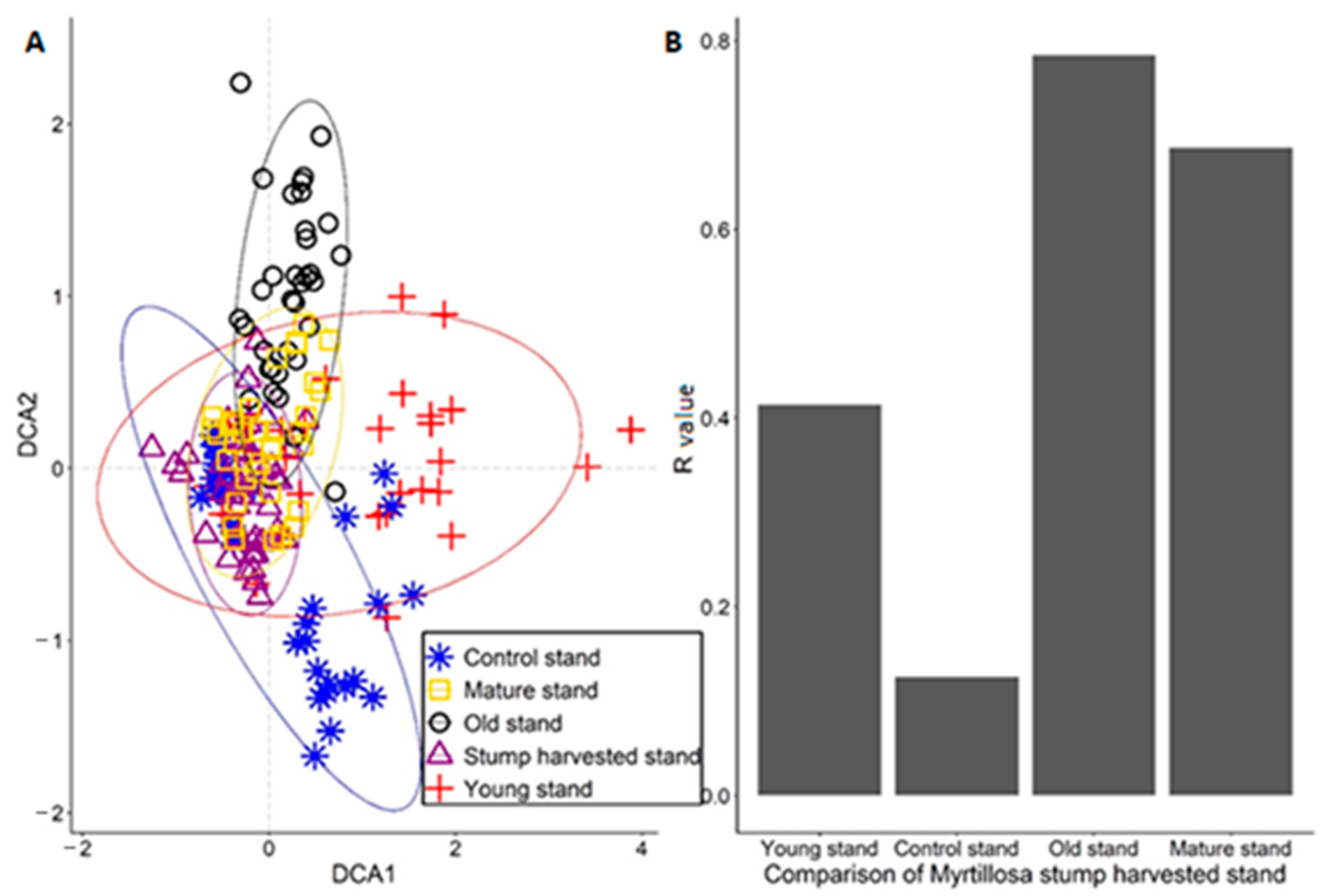
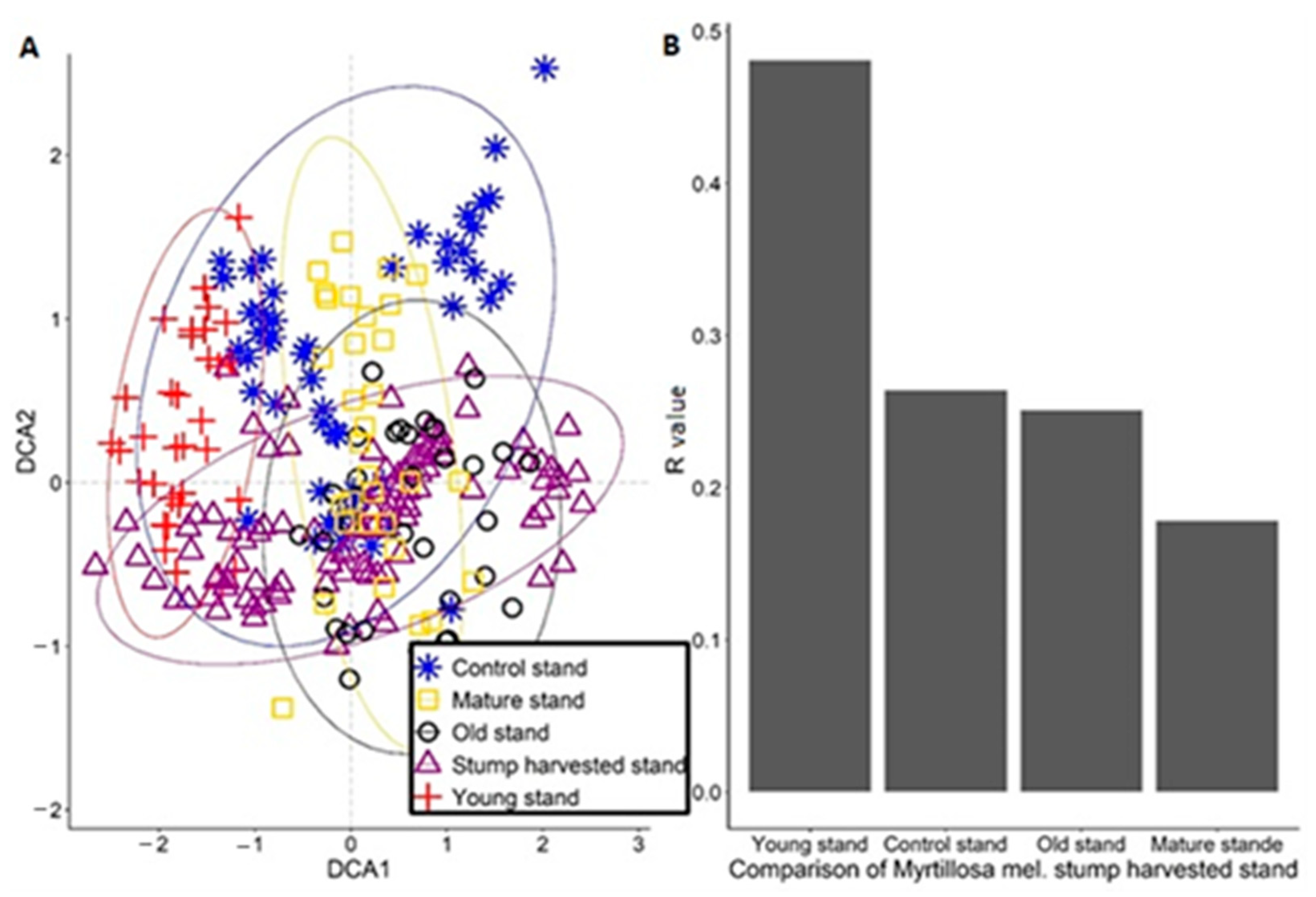
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Šreinerts, P. Kurzemes hercogistes mežsaimniecība un meža likumi 16. un 17. gadu simtenī. Lat. Uni. Rak Lauksai. Fakul. Ser. 1939, 4, 381–411. [Google Scholar]
- Kaye, T.P. Pine Tar; History and Uses; San Francisco Maritime Park Association: San Francisco, CA, USA, 1997; Available online: http://www.maritime.org/conf/conf-kaye-tar.htm (accessed on 5 February 2021).
- Morozovs, A.; Irbe, I.; Bukšāns, E. Koksnes Ķīmiskā Pārstrāde un Aizsardzība.; Avots: Riga, Latvia, 2018; 172p. [Google Scholar]
- Helmisaari, H.S.; Makkonen, K.; Kellomäki, S.; Valtonen, E.; Mälkönen, E. Below- and above-ground biomass, production and nitrogen use in Scots pine stands in eastern Finland. For. Ecol. Manag. 2002, 165, 317–326. [Google Scholar] [CrossRef]
- Iivonen, S.; Kaakinen, S.; Jolkkonen, A.; Vapaavuori, E.; Linder, S. Influence of long-term nutrient optimization on biomass, carbon, and nitrogen acquisition and allocation in Norway spruce. Can. J. For. Res. 2006, 36, 1563–1571. [Google Scholar] [CrossRef]
- Liepiņš, J. Kokaudžu biomasas noteikšanas metodikas izstrāde un oglekļa uzkrājuma aprēķini Latvijā. Ph.D. Thesis, Latvia University of Life Sciences and Technologies, Jelgava, Latvia, 27 August 2019. [Google Scholar]
- Urban, J.; Čermák, J.; Ceulemans, R. Above- and below-ground biomass, surface and volume, and stored water in a mature Scots pine stand. Eur. J. For. Res. 2015, 134, 61–74. [Google Scholar] [CrossRef] [Green Version]
- Cienciala, E.; Černý, M.; Tatarinov, F.; Apltauer, J.; Exnerová, Z. Biomass functions applicable to Scots pine. Trees Struct. Funct. 2006, 20, 483–495. [Google Scholar] [CrossRef]
- LUKE. Natural Resources Institute Finland—Statistics Database. Available online: https://stat.luke.fi/en/energy-2016_en-0 (accessed on 1 September 2020).
- Carey, M.L. Whole Tree Harvesting in Sitka Spruce. Possibilities and Implications. Ir. For. 1980, 37, 48–63. [Google Scholar]
- Nisbet, T.; Dutch, J.; Moffat, A. Harvesting a Guide to Whole—Tree Harvesting; Forestry Practice: Edinburgh, UK, 1997; pp. 1–11.
- Kataja-aho, S.; Fritze, H.; Haimi, J. Short-term responses of soil decomposer and plant communities to stump harvesting in boreal forests. For. Ecol. Manag. 2011, 262, 379–388. [Google Scholar] [CrossRef]
- Mitchell, D. Stump Harvesting. In Proceedings of the 2009 COFE: Environmentally Sound Forest Operations 32nd Annual Meeting of the Council on Forest Engineering, North Tahoe Conference Center, Kings Beach, CA, USA, 15–18 June 2009; p. 6. [Google Scholar]
- Heninger, R.; Scott, W.; Dobkowski, A.; Miller, R.; Anderson, H.; Duke, S. Soil disturbance and 10-year growth response of coast Douglas-fir on nontilled and tilled skid trails in the Oregon Cascades. Can. J. For. Res. 2002, 32, 233–246. [Google Scholar] [CrossRef]
- Thiffault, E.; Béchard, A.; Paré, D.; Allen, D. Recovery rate of harvest residues for bioenergy in boreal and temperate forests: A review. Wiley Interdiscip. Rev. Energy Environ. 2015, 4, 429–451. [Google Scholar] [CrossRef]
- Vanguelova, E.I.; Pitman, R.; Benham, S.; Perks, M.; Morison, J.I. Impact of tree stump harvesting on soil carbon and nutrients and second rotation tree growth in Mid-Wales, UK. Open J. For. 2017, 7, 58–77. [Google Scholar] [CrossRef] [Green Version]
- Karlsson, K.; Tamminen, P. Long-term effects of stump harvesting on soil properties and tree growth in Scots pine and Norway spruce stands. Scand. J. For. Res. 2013, 28, 550–558. [Google Scholar] [CrossRef]
- Palviainen, M.; Finér, L.; Laiho, R.; Shorohova, E.; Kapitsa, E.; Vanha-Majamaa, I. Carbon and nitrogen release from decomposing Scots pine, Norway spruce and silver birch stumps. For. Ecol. Manag. 2010, 259, 390–398. [Google Scholar] [CrossRef]
- Palviainen, M.; Finér, L.; Kurka, A.M.; Mannerkoski, H.; Piirainen, S.; Starr, M. Decomposition and nutrient release from logging residues after clear-cutting of mixed boreal forest. Plant Soil 2004, 263, 53–67. [Google Scholar] [CrossRef]
- Novák, J.; Dušek, D.; Kacálek, D.; Slodičák, M. Analysis of biomass in young Scots pine stands as a basis for sustainable forest management in Czech lowlands. J. For. Sci. 2017, 63, 555–561. [Google Scholar] [CrossRef] [Green Version]
- Bílek, L.; Remeš, J.; Fulín, M.; Chalupová, T.; Procházka, J. Množství a distribuce nadzemní biomasy borovice lesní v oblasti přirozených borů. The amount and distribution of above-ground biomass of Scots pine on natural pine sites. Zprávy Lesn. Výzk. 2016, 61, 108–114. [Google Scholar]
- Saarsalmi, A.; Tamminen, P.; Kukkola, M.; Hautajärvi, R. Whole-tree harvesting at clear-felling: Impact on soil chemistry, needle nutrient concentrations and growth of Scots pine. Scand. J. For. Res. 2010, 25, 148–156. [Google Scholar] [CrossRef]
- Kataja-Aho, S.; Saari, E.; Fritze, H.; Haimi, J. Effects of stump removal on soil decomposer communities in undisturbed patches of the forest floor. Scand. J. For. Res. 2011, 26, 221–231. [Google Scholar] [CrossRef]
- Olsson, B.A.; Staaf, H. Influence of Harvesting Intensity of Logging Residues on Ground Vegetation in Coniferous Forests. J. Appl. Ecol. 1995, 32, 640–654. [Google Scholar] [CrossRef]
- Berg, S. Technology and systems for stump harvesting with low ground disturbance. Ph.D. Thesis, Swedish University of Agricultural Sciences, Umea, Sweden, 2 December 2014. [Google Scholar]
- Pykälä, J. Immediate increase in plant species richness after clear-cutting of boreal herb-rich forests. Appl. Veg. Sci. 2004, 7, 29–34. [Google Scholar] [CrossRef]
- Hyvönen, R.; Kaarakka, L.; Leppälammi-Kujansuu, J.; Olsson, B.A.; Palviainen, M.; Vegerfors-Persson, B.; Helmisaari, H.S. Effects of stump harvesting on soil C and N stocks and vegetation 8–13 years after clear-cutting. For. Eco. Manag. 2016, 371, 23–32. [Google Scholar] [CrossRef]
- Strömgren, M.; Mjöfors, K.; Olsson, B.A. Soil-surface CO2 flux during the first 2 years after stump harvesting and site preparation in 14 Swedish forests. Scand. J. For. Res. 2016, 32, 213–221. [Google Scholar] [CrossRef]
- Persson, T.; Lenoir, L.; Vegerfors, B. Long-term effects of stump harvesting and site preparation on pools and fluxes of soil carbon and nitrogen in central Sweden. Scand. J. For. Res. 2016, 32, 222–229. [Google Scholar] [CrossRef]
- Nikodemums, O.; Kļaviņš, M.; Krišjāne, Z.; Zelčs, V. Latvija. Zeme, Daba, Tauta; Latvijas Universitātes Akadēmiskais Apgāds: Riga, Latvia, 2018; pp. 225–271. [Google Scholar]
- Braun-Blanquet, J. Pflanzensoziologie, Grundzüge der Vegetationskunde, 3rd ed.; Springer: Wien, Austria; New York, NY, USA, 1964; p. 865. [Google Scholar]
- LVS—Latvian Standard—ISO. Augsnes Kvalitāte. Organiskā un Kopējā Oglekļa Noteikšana Pēc Sausās Sadedzināšanas (Elementanalīze); LVS ISO 10694:2006; VSIA Latvijas Standarts: Riga, Latvia, 2006; p. 12. [Google Scholar]
- Bradstreet, R.B. Kjeldahl method for organic nitrogen. Anal. Chem. 1954, 26, 185–187. [Google Scholar] [CrossRef]
- LVS—Latvian Standard—ISO. Augsnes Kvalitāte: Kopējā Slāpekļa Noteikšana. Modificēta Kjeldāla Metode; LVS ISO 11261:2002 L; VSIA Latvijas Standarts: Riga, Latvia, 2002; p. 8. [Google Scholar]
- Thomas, S.C.; Martin, A.R. Carbon content of tree tissues: A synthesis. Forests 2012, 3, 332–352. [Google Scholar] [CrossRef] [Green Version]
- Spellerberg, I.F.; Fedor, P.J. A tribute to Claude-Shannon (1916–2001) and a plea for more rigorous use of species richness, species diversity and the ‘Shannon-Wiener’ Index. Glob. Ecol. Biogeol. 2003, 12, 177–179. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing. Available online: https://www.R-project.org/ (accessed on 20 May 2019).
- Clarck, K.R. Non-parametric multivariate analyses of changes in community structure. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Henry, M.H.; Wagner, H.; et al. Vegan: Community Ecology Package. R Package Version 2.0-10. 2013. Available online: http://CRAN.R-project.org/package=vegan (accessed on 5 February 2021).
- Correa-Metrio, A.; Dechnik, Y.; Lozano-García, S.; Caballero, M. Detrended correspondence analysis: A useful tool to quantify ecological changes from fossil data sets. Bol. Soc. Geol. Mex. 2014, 66, 135–143. [Google Scholar] [CrossRef]
- Ellenberg, H. Zeigerwerte der Gefässpflanzen Mitteleuropas; Scripta Geobotanica: Göttingen, Germany, 1974; 197p. [Google Scholar]
- Ellenberg, H.; Weber, H.E.; Düll, R.; Wirth, V.; Werner, W.; Paulissen, D. Zeigerwerte von Pflanzen in Mitteleuropa, 2nd ed.; Scripta Geobotanica: Göttingen, Germany, 1992; pp. 1–248. [Google Scholar]
- Frego, K.A. Regeneration of four boreal bryophytes: Colonization of experimental gaps by naturally occurring propagules. Can. J. Bot. 1996, 74, 1937–1942. [Google Scholar] [CrossRef]
- Saliņš, Z. Meža Izmantošana Latvijā; Jelgavas Tipogrāfija: Jelgava, Latvia, 1999; pp. 46–190. [Google Scholar]
- Schofield, W.B. Introduction to Bryology; The Blackburn Press: Caldwell, NJ, USA, 2001; 432p. [Google Scholar]
- Auld, B.A.; Coote, B.G. A Model of a Spreading Plant Population. Oikos 1980, 34, 287. [Google Scholar] [CrossRef]
- Schupp, E.W. Quantity, quality and the effectiveness of seed dispersal by animals. In Frugivory and seed dispersal: Ecological and evolutionary aspects. Vegetatio 1993, 107, 15–29. [Google Scholar]
- Watson, A.; Welch, D.; Heslop, R.E.F. Deschampsia flexuosa snowbed grassland on granitic mountains in the cairngorms. Plant Ecol. Divers 2010, 3, 95–99. [Google Scholar] [CrossRef]
- Porley, R.; Hodgetts, N. Mosses and Liverworts; HarperCollins: London, UK, 2005; 495p. [Google Scholar]
- Christy, M. The hornbeam (Carpinus betulus L.). Br. J. Ecol. 1924, 12, 39–94. [Google Scholar] [CrossRef]
- Ritchie, J.C. Vaccinium myrtillus L. J. Ecol. 1956, 44, 291–299. [Google Scholar] [CrossRef]
- Jägerbrand, A.K.; During, H.J. Effects of simulated shade on growth, number of branches and biomass in Hylocomium splendens and Racomitrium lanuginosum. Lindbergia 2005, 117–124. [Google Scholar] [CrossRef]
- Taylor, K.; Rowland, A.P.; Jones, H.E. Molinia caerulea (L.) Moench. J. Ecol. 2001, 89, 126–144. [Google Scholar] [CrossRef]
- Hotanen, J.P.; Saarinen, M.; Nousiainen, H. Purple Moor Grass (Molinia caerulea) on peatlands drained for forestry in Finland. Suo 2016, 67, 81–90. [Google Scholar]
- Jefferies, T.A. Ecology of the Purple Heath Grass (Molinia Caerulea). J. Ecol. 1915, 3, 93–109. [Google Scholar] [CrossRef]
- Ross-Davis, A.L.; Frego, K.A. Comparison of plantations and naturally regenerated clearcuts in the Acadian Forest: Forest floor bryophyte community and habitat features. Can. J. Bot. 2002, 80, 21–33. [Google Scholar] [CrossRef]
- Ellenberg, H. Vegetation Ecology of Central Europe, 4th ed.; Cambridge University Press: Cambridge, UK, 1988; 731p. [Google Scholar]
- Priedītis, N. Latvijas Mežs: Daba un Daudzveidība; WWF—Pasaules Dabas Fonds: Riga, Latvia, 1999; 93p. [Google Scholar]
- Berķis, A.; Meijere, A.; Sedlenieks, A.; Vanags, A.; Ansons, G.; Rove, I.; Brauns, J.; Grīslis, J.; Gaigals, J. Rokasgrāmata Meža Tipu Noteikšanai; LVM—Latvijas Valsts Meži: Riga, Latvia, 2013; 69p. [Google Scholar]
- Ikauniece, S. Aizsargājamo Biotopu Saglabāšanas Vadlīnijas Latvijā. 6. Sējums. Meži; Dabas Aizsardzības Pārvalde: Sigulda, Latvia, 2017; 167p.
- Pearson, G.A. Light and Moisture in Forestry. Ecology 1930, 11, 145–160. [Google Scholar] [CrossRef]
- North, M.; Oakley, B.; Fiegener, R.; Gray, A.; Barbour, M. Influence of light and soil moisture on Sierran mixed-conifer understory communities. Plant Ecol. 2005, 177, 13–24. [Google Scholar] [CrossRef]
- Strazdiņa, L.; Strazdiņa, L. Alpine flora of South Norway. Experience of Latvian bryologists and botanists. Lat. Veģ. 2015, 24, 87–92. [Google Scholar]
- Zālītis, P.; Jansons, J. Latvijas Meža Tipoloģija un Tās Sākotne; LVMI „Silava”: Salaspils, Latvia, 2013; pp. 71–91. [Google Scholar]
- Strazdiņa, L.; Madžule, L.; Brumelis, G. A contribution to the bryoflora of Moricsala island nature reserve, Latvia. Folia Cryptogam. Est. 2011, 48, 107–117. [Google Scholar]
- Horn, H.S. Markovian Properties of Forest Succession. In The Ecology and Evolution of Communities; Harvard University Press: Cambridge, MA, USA, 1975. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forest Type | Type | Composition Ratio | Age | Stand | H1, m | DBH2, cm | Basal Area, m2 ha−1 | Yield, m3 ha−1 |
|---|---|---|---|---|---|---|---|---|
| Vacciniosa | FBR stand | 10P3 | 50 | 504-66-11 | 19 | 22.6 | 21.7 | 198.9 |
| Vacciniosa | Control stand Control stand Control stand | 10P | 53 | 508-110-4 | 18.6 | 18.4 | 19.6 | 179.5 |
| Vacciniosa | 10P | 48 | 508-109-16 | 17.9 | 20.23 | 14.6 | 128.3 | |
| Vacciniosa | 10P | 51 | 504-66-17 | 18.1 | 18.2 | 35.4 | 311.6 | |
| Myrtillosa | FBR stand | 10P | 40 | 705-246-2 | 17.5 | 20.2 | 22.4 | 194.1 |
| Myrtillosa | Control stand | 10P | 41 | 710-300-36 | 19.5 | 24.2 | 25.3 | 240.0 |
| Myrtillosa | Control stand | 10P | 35 | 713-300-14 | 17.7 | 22.1 | 28.1 | 248.0 |
| Myrtillosa | Control stand | 8P 2E4 | 40 | 713-205-19 | 16.3 | 22 | 15.8 | 127.4 |
| Myrtillosa | Control stand | 9P1B5 | 40 | 705-195-33 | 15.3 | 15.3 | 29.8 | 229.1 |
| Myrtillosa mel. | FBR stand | 10P | 41 | 209-418-10 | 21.2 | 24.6 | 30.3 | 302.2 |
| Myrtillosa mel. | FBR stand | 10P | 35 | 209-419-8 | 17.8 | 22.9 | 24.8 | 218 |
| Myrtillosa mel. | Control stand | 9P1E | 41 | 209-448-17 | 13.9 | 18.5 | 21.8 | 159.8 |
| Myrtillosa mel. | Control stand | 9P1B | 43 | 709-216-1 | 17.6 | 19.9 | 23.5 | 200.4 |
| Species | Vacciniosa | Myrtilliosa | Myrtilliosa Mel. |
|---|---|---|---|
| Acer platinoides | - | × | × |
| Amelanchier spicata | - | × | - |
| Betula pendula | × | × | × |
| Betula pubescens | - | - | × |
| Cerasus avium | - | × | - |
| Corylus avellana | - | × | - |
| Frangula alnus | × | × | × |
| Picea abies | × | × | × |
| Pinus sylvestris | × | × | × |
| Populus tremula | × | × | - |
| Prunus domestica | - | - | × |
| Quercus robur | - | × | - |
| Salix sp. | - | - | × |
| Sorbus aucuparia | - | × | × |
| Tilia cordata | × | - | × |
| Viburnum opulus | - | × | - |
| Variable | Forest Type | FBR 5 | Control Stand 1 | Young Stand 2 | Mature Stand 3 | Old Stand 4 |
|---|---|---|---|---|---|---|
| Number of species | Vacciniosa | 22 | 24 | 17 | 14 | 20 |
| Myrtillosa | 41 | 35 | 49 | 20 | 35 | |
| Myrtillosa mel. | 78 | 46 | 20 | 51 | 43 | |
| Shannon- Wiener index | Vacciniosa | 1.05 b | 1.22 bc | 1.57 a | 1.09 bc | 1.32 c |
| Myrtillosa | 1.59 a | 1.59 a | 1.70 a | 2.08 b | 1.95 b | |
| Myrtillosa mel. | 1.80 bc | 1.54 a | 1.63 ab | 1.62 ab | 1.89 c |
| Forest Type | Territories | Light | Temperature | Moisture | Soil pH |
|---|---|---|---|---|---|
| Vaccinosa | FBR stand | 6.79 a | 4.60 a | 4.71 b | 1.97 ab |
| Control stand | 5.66 b | 4.60 a | 4.97 a | 2.09 b | |
| Young stand | 6.76 a | 4.66 a | 4.96 a | 1.67 a | |
| Mature stand | 5.31 b | 4.24 b | 4.97 a | 1.93 ab | |
| Old stand | 5.38 b | 4.63 a | 4.94 a | 2.04 ab | |
| Myrtillosa | FBR stand | 7.37 b | 5.55 b | 3.21 b | 3.92 c |
| Control stand | 6.50 a | 5.29 a | 4.57 a | 3.66 b | |
| Young stand | 6.97 ab | 5.24 a | 4.58 a | 4.38 a | |
| Mature stand | 5.85 d | 5.24 a | 4.77 a | 3.53 b | |
| Old stand | 4.39 c | 5.17 a | 5.16 a | 4.23 a | |
| Myrtillosa mel. | FBR stand | 6.07 ab | 5.15 c | 4.39 c | 3.73 c |
| Control stand | 5.90 a | 4.91 ab | 5.38 ab | 4.10 b | |
| Young stand | 5.59 a | 4.78 a | 5.87 a | 3.33 a | |
| Mature stand | 6.53 b | 5.08 c | 4.76 cd | 4.23 b | |
| Old stand | 4.86 c | 4.94 bc | 5.04 bd | 3.84 bc |
| Forest type | Territories | Light | Temperature | Moisture | Soil pH |
|---|---|---|---|---|---|
| Vaccinosa | FBR stand | 5.75 b | 2.86 b | 3.95 b | 2.61 b |
| Control stand | 5.98 b | 3.00 b | 4.05 bc | 3.51 a | |
| Young stand | 5.22 a | 2.36 a | 3.63 a | 3.33 a | |
| Mature stand | 5.93 b | 3.01 b | 4.21 c | 3.88 ac | |
| Old stand | 5.90 b | 2.86 b | 4.11 bc | 4.18 c | |
| Myrtillosa | FBR stand | 6.05 a | 2.99 a | 4.33 a | 3.56 a |
| Control stand | 6.02 a | 3.01 a | 4.34 a | 3.47 a | |
| Young stand | 5.94 a | 3.00 a | 4.28 a | 3.58 a | |
| Mature stand | 5.47 b | 2.91 a | 4.99 b | 3.37 a | |
| Old stand | 5.60 b | 3.04 a | 4.78 b | 3.44 a | |
| Myrtillosa mel. | FBR stand | 3.72 b | 2.16 ab | 3.00 b | 3.52 c |
| Control stand | 4.67 bc | 2.71 bc | 3.89 a | 4.57 b | |
| Young stand | 2.32 a | 1.66 a | 3.92 ab | 1.72 a | |
| Mature stand | 5.58 c | 3.36 c | 4.15 a | 5.43 b | |
| Old stand | 4.43 bc | 2.65 bc | 3.60 ab | 4.14 b |
| Forest Type | FBR Stand / Control Stand | Total C | Soil Density | pH |
|---|---|---|---|---|
| Vacciniosa | FBR stand | 112.19 | 1164.62 | 5.47 |
| Vacciniosa | Control stand | 140.75 | 754.00 | 4.85 |
| Myrtillosa | FBR stand | 112.96 | 1597.53 | 4.64 |
| Myrtillosa | Control stand | 120.93 | 1445.27 | 3.82 |
| Myrtillosa mel. | FBR stand | 142.82 | 1397.00 | 4.30 |
| Myrtillosa mel. | Control stand | 113.24 | 1343.88 | 4.10 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Čakšs, R.; Čakša, L.; Desaine, I.; Lībiete, Z.; Elferts, D.; Butlers, A.; Jansons, Ā. Long-Term Influence of Stump-Removal on Components of Hemiboreal Pine Forest Ecosystem. Sustainability 2021, 13, 2095. https://doi.org/10.3390/su13042095
Čakšs R, Čakša L, Desaine I, Lībiete Z, Elferts D, Butlers A, Jansons Ā. Long-Term Influence of Stump-Removal on Components of Hemiboreal Pine Forest Ecosystem. Sustainability. 2021; 13(4):2095. https://doi.org/10.3390/su13042095
Chicago/Turabian StyleČakšs, Roberts, Linda Čakša, Iveta Desaine, Zane Lībiete, Didzis Elferts, Aldis Butlers, and Āris Jansons. 2021. "Long-Term Influence of Stump-Removal on Components of Hemiboreal Pine Forest Ecosystem" Sustainability 13, no. 4: 2095. https://doi.org/10.3390/su13042095
APA StyleČakšs, R., Čakša, L., Desaine, I., Lībiete, Z., Elferts, D., Butlers, A., & Jansons, Ā. (2021). Long-Term Influence of Stump-Removal on Components of Hemiboreal Pine Forest Ecosystem. Sustainability, 13(4), 2095. https://doi.org/10.3390/su13042095