
Characteristics of Selected Silphium Species as Alternative Plants for Cultivation and Industry with Particular Emphasis on Research Conducted in Poland: A Review


Abstract
:1. Introduction
2. Methods
- anatomical and morphological features of selected Silphium species,
- biomass productivity in various geographic locations,
- chemical composition with division into primary metabolites, secondary metabolites, and minerals,
- multidirectional utility values of Silphium species biomass.
3. Results and Discussion
3.1. Genus Silphium L.
3.1.1. Description of Selected Silphium Species
Blooming
Seeds
3.1.2. Anatomy
3.1.3. Cultivation
3.1.4. Yields and Dry Matter (DM) Content
3.2. Chemical Composition
3.2.1. Carbohydrates
3.2.2. Protein and Amino Acids
3.2.3. Fat
3.2.4. L-Ascorbic Acid
3.2.5. Chlorophyll
3.2.6. Mineral Substances: Ash
3.2.7. Phenolic Acids
3.2.8. Tannins
3.2.9. Flavonoids
3.2.10. Carotenoids
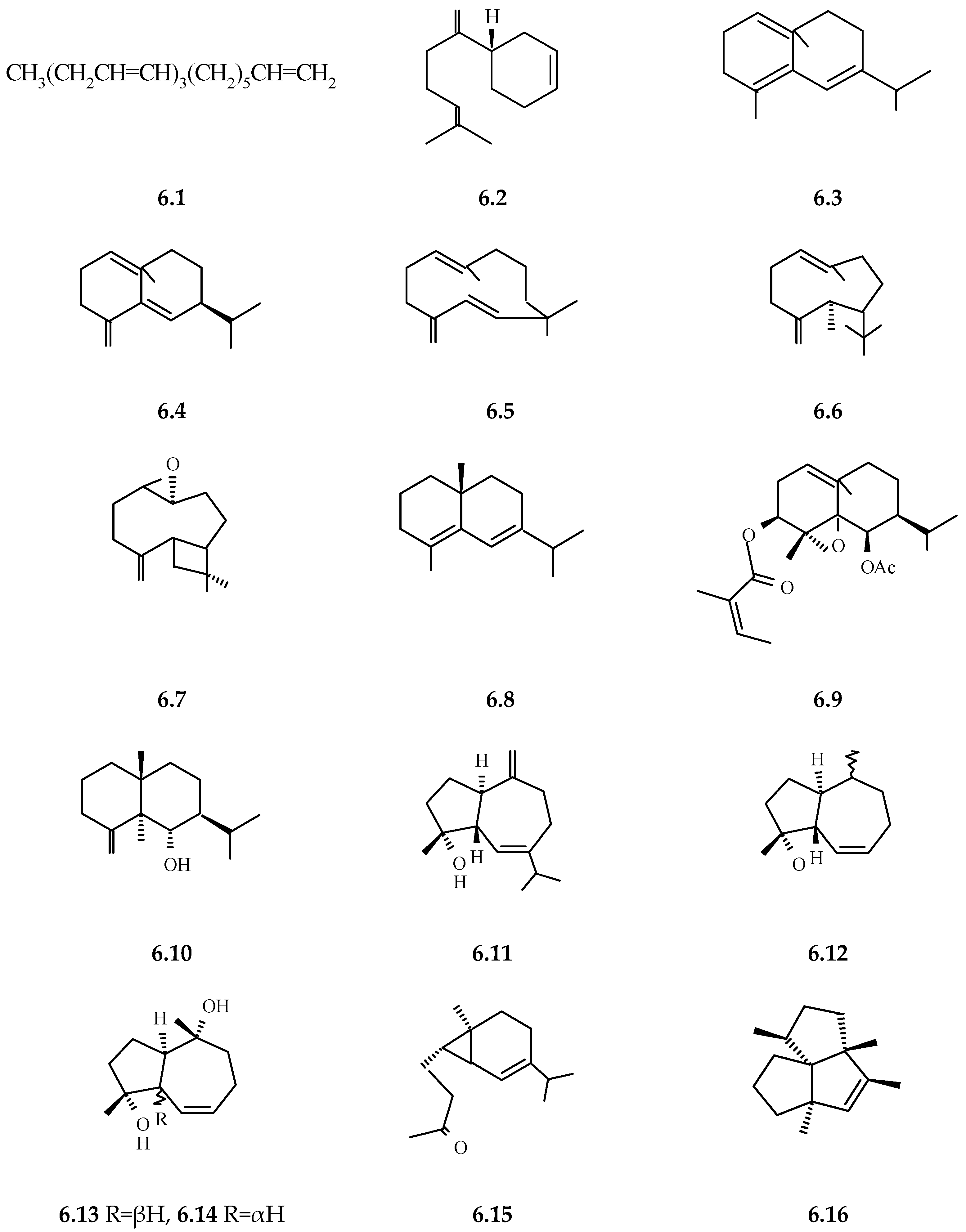
3.2.11. Volatile Components (Essential Oil, Volatile Components of Extracts)
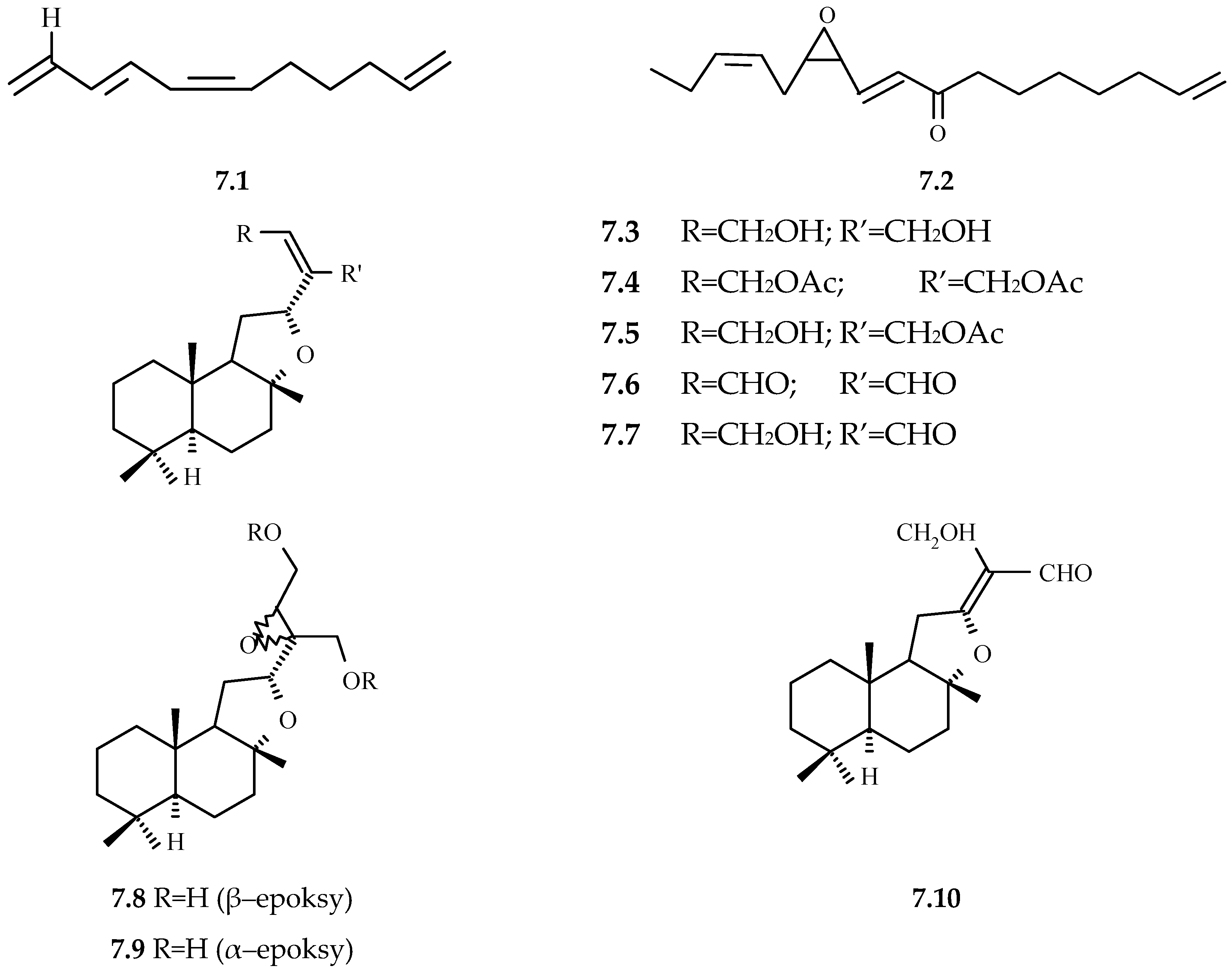
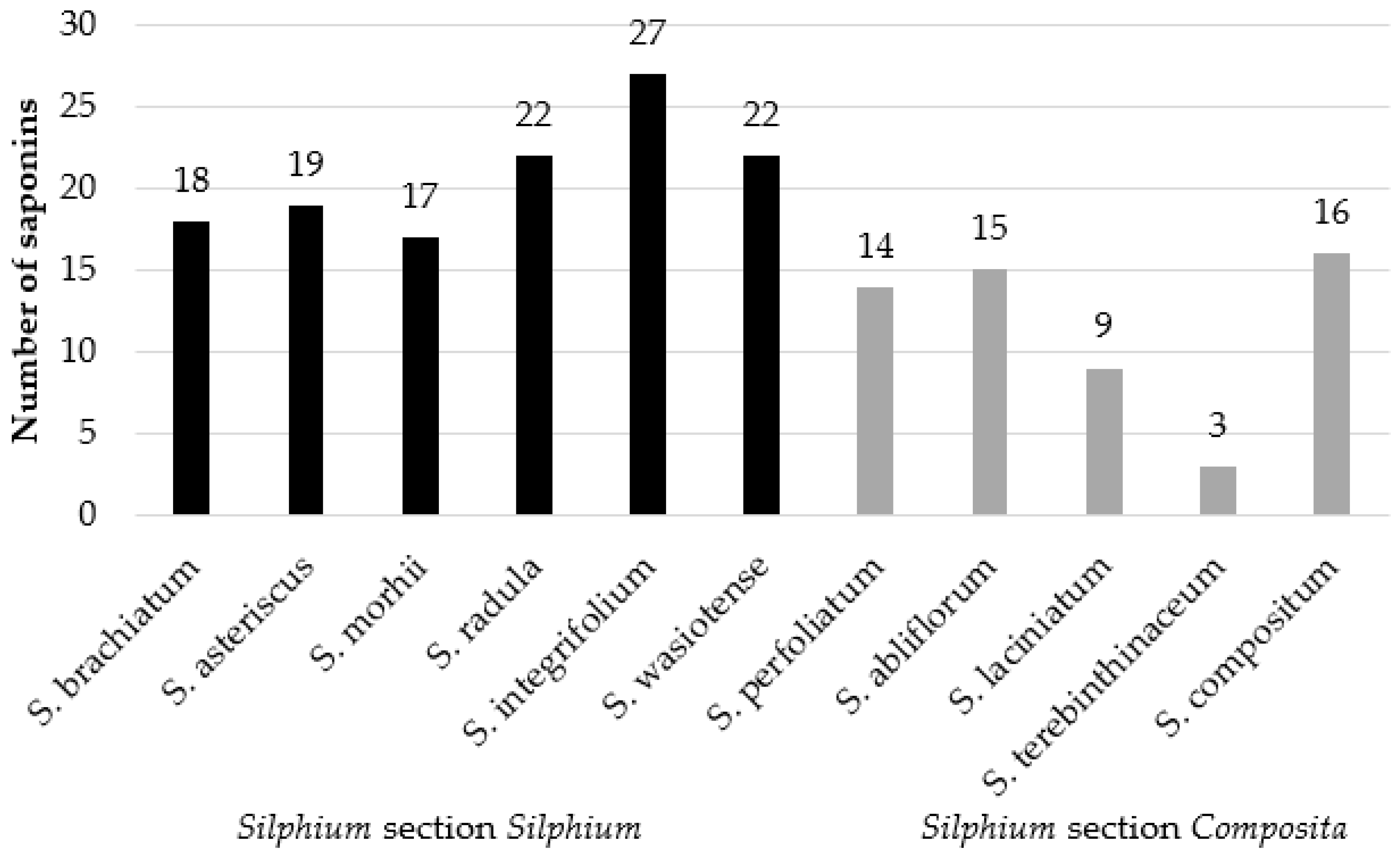
3.2.12. Saponins
3.3. Biological Activity
3.3.1. Historical Medicinal Applications
3.3.2. Research on Biological Activity In Vivo
3.3.3. Research on Biological Activity In Vitro
Experiments on Cancer Cells, against the HIV Virus, and Immunosuppressant Activity
Research in Relation to Fungus- and Bacterium-Caused Diseases of Humans and Animals
Research in Relation to Fungus-Caused Plant Diseases
3.3.4. The Effect of Triterpene Glycosides on Seed Germination and Catalase Activity
3.3.5. Extracts from Silphium as Additives Inhibiting Unfavorable Changes in Fats
3.3.6. Antioxidant Properties of Extracts from Silphium
3.3.7. Application in Animal Feeding
3.3.8. Ornamental Plant and Honeybee Forage
3.3.9. Application for Soil Remediation
3.3.10. Application as an Energy Crop
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAO. Report of the FAO Council, 94th Session; FAO: Rome, Italy, 1988. [Google Scholar]
- Batmunkh, A.; Nugroho, A.D.; Fekete-Farkas, M.; Lakner, Z. Global challenges and responses: Agriculture, economic globalization, and environmental sustainability in Central Asia. Sustainability 2022, 14, 2455. [Google Scholar] [CrossRef]
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S.; et al. Crop production under drought and heat stress: Plant responses and management options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yıldız, M.; Tansı, S.; Sezen, M. New Plants with Commercial Potent. Available online: https://dergipark.org.tr/tr/download/article-file/142220 (accessed on 25 February 2022).
- Kowalski, R.; Kędzia, B. Antibacterial activity of Silphium perfoliatum extracts. Pharm. Biol. 2007, 45, 494–500. [Google Scholar] [CrossRef]
- Steyermark, J.A. Flora of Missouri; The Iova State University Press: Ames, IA, USA, 1962. [Google Scholar]
- Kowalski, R.; Wolski, T. Charakterystyka wzrostu i rozwoju rożnika przerośniętego Silphium perfoliatum L. w pierwszych latach uprawy. Ann. Univ. Mariae Curie-Sklodowska Sect. E 2001, 9, 311–317. [Google Scholar]
- Peni, D.; Stolarski, M.J.; Bordiean, A.; Krzyżaniak, M.; Dębowski, M. Silphium perfoliatum—A Herbaceous Crop with Increased Interest in Recent Years for Multi-Purpose Use. Agriculture 2020, 10, 640. [Google Scholar] [CrossRef]
- Worth, F.; Headwater, P.; Ecology, S.; Population, W.P.; Lizard, T.H. Viability of Silphium albiflorum in Tarrant County. Post Oak Prairie J. 2015, 1, 1–35. [Google Scholar]
- Brickell, C. Wielka Encyklopedia Roślin Ogrodowych od a do z; Muza SA: Warszaw, Poland, 1999. [Google Scholar]
- Rickett, H.W. Wild Flowers of the United States. Volume One. The Northeastern States. From the Atlantic to Minnesota and Missouri and From the Canadian Border to Virginia and Missouri, 1st ed.; McGraw-Hill for the The New York Botanical Garden: New York, NY, USA, 1966. [Google Scholar]
- Tutin, T.G.; Heywood, V.; Burges, N.A.; Moore, D.M.; Valentine, D.H.; Walters, S.M.; Webb, D.A. Flora Europea; Cambrige University Press: Cambrige, UK, 1976; Volume 4. [Google Scholar]
- Clevinger, J.A.; Panero, J.L. Phylogenetic analysis of Silphium and subtribe Engelmanniinae (Asteraceae: Heliantheae) based on ITS and ETS sequence data. Am. J. Bot. 2000, 87, 565–572. [Google Scholar] [CrossRef]
- Puia, I.; Szabó, A.T. Culture experimentale d’ une nouvelle espece fourragére—Silphium perfoliatum L.—dans le Jardin Agrobotanique de Cluj-Napoca. Institutum Agronomicum „Dr. Petru Groza”, Cluj-Napoca (Romania). Not. Bot. Horti Agrobot. 1985, 15, 15–20. [Google Scholar]
- Stuessy, T.F. Heliantheae—Systematic review. In The Biology and Chemistry of the Compositae. II; Heywood, V.H., Harborne, J.B., Turner, B.L., Eds.; Academic Press: London, UK, 1977; pp. 621–672. [Google Scholar]
- Davidyants, E.S.; Abubakirov, N.K. Chimičeskij sostav i perspektivy ispol’zowaniya rasteniy r. Silphium L. Rastit. Resur. 1992, 28, 118–128. [Google Scholar]
- Huxley, A. Dictinary of Gardening, 4th ed.; Stockton Press: New York, NY, USA, 1992. [Google Scholar]
- Čubarova, G.V. Sil’fia pronzennolistnaja—odna iz perspektivnych silosnych kultur. Životnovodstvo 1971, 8, 17–18. [Google Scholar]
- Niqueux, M. Une nouvelle plante fourragére: Le silfe (Silphium perfoliatum L.). Fourrages 1981, 119–136. Available online: https://afpf-asso.fr/article/une-nouvelle-plante-fourragere-le-silfe-silphium-perfoliatum-l (accessed on 25 February 2022).
- Duranti, E.; Santilocchi, R.; Casoli, C. Composizione chimica e valore nutritivo di Silphium perfoliatum L. conservato mediante insilamento. Zootec. Nutr. Anim. 1988, 14, 349–356. [Google Scholar]
- Neumerkel, W.; Märtin, B.; Linke, G. Silphium perfoliatum L. eine Nutzpflanze? Wissenschaftliche Zeitschrift der Martin-Luther-Universität Halle 1978, 27, 31–38. [Google Scholar]
- Zhang, X.; Xia, H.; Li, Z.; Zhuang, P.; Gao, B. Potential of four forage grasses in remediation of Cd and Zn contaminated soils. Bioresour. Technol. 2010, 101, 2063–2066. [Google Scholar] [CrossRef]
- Schittenhelm, S.; Kottmann, L.; Schoo, B. Wasser als ertragsbegrenzender Faktor [Water as a limiting factor for crop yield]. J. Fur Kult. 2017, 69, 80–86. [Google Scholar]
- Haag, N.L.; Nägele, H.-J.; Reiss, K.; Biertümpfel, A.; Oechsner, H. Methane formation potential of cup plant (Silphium perfoliatum). Biomass Bioenergy 2015, 75, 126–133. [Google Scholar] [CrossRef]
- Matthews, J.; Beringen, R.; Huijbregts, M.A.J.; Van der Mheen, H.J.; Odé, B.; Trindade, L.; Van Valkenburg, J.L.C.H.; Van der Velde, G.; Leuven, R.S.E.W. Horizon Scanning and Environmental Risk Analyses of Non-Native Biomass Crops in The Netherlands. Available online: https://www.researchgate.net/publication/297136883_Horizon_scanning_and_environmental_risk_analyses_of_non-native_biomass_crops_in_the_Netherlands (accessed on 25 February 2022).
- Kowalski, R. Silphium trifoliatum L.—A new alternative cultivation herbal plant? Acta Agric. Scand. Sect. B Soil Plant Sci. 2007, 57, 155–166. [Google Scholar]
- Kowalski, R. Growth and development of Silphium integrifolium in the first 3 years of cultivation. N. Z. J. Crop Hortic. Sci. 2004, 32, 389–395. [Google Scholar] [CrossRef]
- Pichard, G. Management, production, and nutritional characteristics of cup-plant (Silphium perfoliatum) in temperate climates of southern Chile. Cienc. Investig. Agrar. 2012, 39, 61–77. [Google Scholar] [CrossRef] [Green Version]
- Weryszko-Chmielewska, E.; Kowalski, R.; Wolski, T. Rożnik przerośnięty (Silphium perfoliatum L.) nowa roślina alternatywna. Część I. Badania morfologiczne i anatomiczne. Zesz. Probl. Postępów Nauk Rol. 1999, 468, 497–505. [Google Scholar]
- Woźniak, M.; Góral, S. Rożnik przerośnięty (Silphium perfoliatum) roślina na pastwiska pszczele. In Proceedings of the Materiały 35 Naukowej Konferencji Pszczelarskiej, Puławy, Poland, 11–12 March 1998; p. 90. [Google Scholar]
- Mueller, A.L.; Biertümpfel, A.; Friedritz, L.; Power, E.F.; Wright, G.A.; Dauber, J. Floral resources provided by the new energy crop, Silphium perfoliatum L. (Asteraceae). J. Apic. Res. 2020, 59, 232–245. [Google Scholar] [CrossRef]
- Mueller, A.L.; Dauber, J. Hoverflies (Diptera: Syrphidae) benefit from a cultivation of the bioenergy crop Silphium perfoliatum L. (Asteraceae) depending on larval feeding type, landscape composition and crop management. Agric. For. Entomol. 2016, 18, 419–431. [Google Scholar] [CrossRef]
- Figas, A.; Sawilska, A.K.; Rolbiecki, R.; Tomaszewska-Sowa, M. Morphological characteristics of achenes and fertility plants of cup plant (Silphium perfoliatum L.) Obtained from micropropagation growing under irrigation. Infrastruct. Ecol. Rural Areas 2016, 4, 1363–1372. [Google Scholar]
- Weryszko-Chmielewska, E.; Michońska, M.; Wolski, T.; Kowalski, R. Porównanie cech morfologicznych kwiatów trzech gatunków Silphium z uwzględnieniem prezenterów pyłkowych i ziarn pyłku. Bibl. Fragm. Agron. 1999, 6, 103–112. [Google Scholar]
- Wróblewska, A. Badania wartości pszczelarskiej Silphium perfoliatum L. In Proceedings of the Materiały I Ogólnopolskiej Konferencji Naukowej „Biologia kwitnienia, nektarowania i zapylania roślin”, Lublin, Poland, 13–14 November 1997; pp. 59–65. [Google Scholar]
- Hoves, F.N. Plants and Beekeeping. An Account of Those Plants, Wild and Cultivated, of Value to the Hive Bee, and for Honey Production in the British Isles. London: 197; Frazer Press: London, UK, 2017. [Google Scholar]
- Daniel, P. Silphium perfoliatum „Durchwachsene Silphie”—Eine neue Bienenweidepflanze. Biene 1984, 120, 196–199. [Google Scholar]
- Pelmenev, V.K. Nowyje kormowyje kul’tury. Nowyje Kormowyje kul’Tury; Medonosnyje Rastenija Rosselchozizdat: Moskva, Russia, 1985; pp. 121–122. [Google Scholar]
- Kučerov, E.V.; Siraeva, S.M. Silfia pronzennolistnaya. Pčelovodstvo 1987, 1, 15–16. [Google Scholar]
- Miller, P.D. Maize pollen: Collection and enzymology. In Maize for biological research; Sheridan, W.F., Ed.; University Press: Grand Forks, ND, USA, 1982; pp. 279–282. [Google Scholar]
- Schäfer, A.; Damerow, L.; Lammers Schulze, P. Durchwachsene Silphie: Bestandesetablierung mittels Aussaat Cup plant: Crop establishment by sowing. J. Kult. 2016, 68, 367–371. [Google Scholar]
- Kowalski, R.; Wierciński, J. Evaluation of chemical composition of some Silphium L. species seeds as alternative foodstuff raw materials. Polish J. Food Nutr. Sci. 2004, 13, 349–354. [Google Scholar]
- Schäfer, A.; Leder, A.; Graff, M.; Damerow, L.; Lammers, P.S. Determination and sorting of cup plant seeds to optimize crop establishment. Landtechnik 2018, 73, 97–105. [Google Scholar]
- von Gehren, P.; Gansberger, M.; Mayr, J.; Liebhard, P. The effect of sowing date and seed pretreatments on establishment of the energy plant Silphium perfoliatum by sowing. Seed Sci. Technol. 2016, 44, 310–319. [Google Scholar] [CrossRef]
- Schäfer, A.; Damerow, L.; Lammers, P.S. Determination of the seed geometry of cup plant as requirement for precision seeding. Landtechnik 2017, 72, 122–128. [Google Scholar]
- Köhler, J.; Biertümpfel, A. Wie die Saat, so die Ernte—Erfolgreiche Etablierung Durchwachsener Silphie durch Aussaat As the sowing, so the harvest—successful establishment of Cup Plant by sowing. J. Kult. 2016, 68, 356–362. [Google Scholar]
- Shalyuta, B.V.; Kostitskaya, E.V. The Yield of Silphium perfoliatum L. Depending on the Conditions of Cultivation. Agric. Eng. 2018, 22, 91–97. [Google Scholar] [CrossRef] [Green Version]
- Kowalski, R.; Kowalska, G.; Jankowska, M.; Nawrocka, A.; Kałwa, K.; Pankiewicz, U.; Włodarczyk-Stasiak, M. Secretory structures and essential oil composition of selected industrial species of Lamiaceae. Acta Sci. Pol. Hortorum Cultus 2019, 18, 53–69. [Google Scholar] [CrossRef]
- Sharifi-Rad, J.; Sureda, A.; Tenore, G.; Daglia, M.; Sharifi-Rad, M.; Valussi, M.; Tundis, R.; Sharifi-Rad, M.; Loizzo, M.; Ademiluyi, A.; et al. Biological Activities of Essential Oils: From Plant Chemoecology to Traditional Healing Systems. Molecules 2017, 22, 70. [Google Scholar] [CrossRef]
- Leyva-López, N.; Gutiérrez-Grijalva, E.; Vazquez-Olivo, G.; Heredia, J. Essential Oils of Oregano: Biological Activity beyond Their Antimicrobial Properties. Molecules 2017, 22, 989. [Google Scholar] [CrossRef] [Green Version]
- Morris, L.J. Silphium laciniatum, Lin., Rosin Weed. Am. J. Pharm. 1881, 53, 487–491. [Google Scholar]
- Kowalski, R. The chemical composition of essential oils and lipophilic extracts of Silphium integrifolium Michx. and Silphium trifoliatum L. rhizomes. J. Essent. Oil Res. 2008, 20, 255–259. [Google Scholar] [CrossRef]
- Kowalski, R. The chemical composition of essential oils and lipophilic extracts of Silphium integrifolium Michx. and S. trifoliatum L. leaves. Flavour Fragr. J. 2008, 23, 164–171. [Google Scholar] [CrossRef]
- Van Tassel, D.L.; Albrecht, K.A.; Bever, J.D.; Boe, A.A.; Brandvain, Y.; Crews, T.E.; Gansberger, M.; Gerstberger, P.; González-Paleo, L.; Hulke, B.S.; et al. Accelerating Domestication: An Opportunity to Develop New Crop Ideotypes and Breeding Strategies Informed by Multiple Disciplines. Crop Sci. 2017, 57, 1274. [Google Scholar] [CrossRef] [Green Version]
- Stanford, G. Silphium perfoliatum (cup-plant) as a new forage. In Proceedings of the 12th North American Prairie Conference, Cedar Falls, IA, USA, 5–9 August 1990; Volume 1, pp. 33–37. [Google Scholar]
- Troxler, J.; Daccord, R. Le silfe (Silphium perfoliatum L.): Un fourrage intéressant? Rev. Suisse d’Agric. 1982, 14, 279–281. [Google Scholar]
- Filatov, V.I.; Baklanow, A.M.; Lavrov, B.V.A.; Komjagin, N. Produktivnost’ sil’fii pronzennolistnoy v zavisimosti ot prijemov agrotechniki na mieliorirovannych počvach. Izv. TSCHA 1986, 1, 58–63. [Google Scholar]
- Douglas, J.A.; Follett, J.M.; Halliday, J.R.; Hughes, J.W.; Parr, C.R. Silphium: Preliminary research on a possible new forage crop for New Zealand. Proc. Agron. Soc. N. Z. 1987, 17, 51–53. [Google Scholar]
- Daniel, P.; Rompf, R. Möglichkeiten und Grenzen der Nutzung der Durchwachsenen Silphie (Silphium perfoliatum L.) als Futter-, nachwachsenole Rohstoff- und Landschaftspflegepflanze. Agribiol. Res. 1994, 47, 345–353. [Google Scholar]
- Kolarova, M.; Grigorova, L. Laboratorna k”lnjaemost na silfiya (Silphium perfoliatum L.). Rastenev’dni Nauk. 1986, 23, 76–84. [Google Scholar]
- Procko, R.F.; Sidorenko, T.V. Doslidi po pidwišcennju schozosti nasinnja Silphium perfoliatum L. Ukr. Bot. Zhurnal 1991, 48, 64–67. [Google Scholar]
- Reinert, S.; Money, K.L.; Rockstad, G.B.G.; Kane, N.C.; Van Tassel, D.L.; Hulke, B.S. Two contrasting laboratory methods improve Silphium integrifolium Michx. germination rate to agronomically acceptable levels. Euphytica 2018, 214, 156. [Google Scholar] [CrossRef]
- von Cossel, M.; Amarysti, C.; Wilhelm, H.; Priya, N.; Winkler, B.; Hoerner, L. The replacement of maize (Zea mays L.) by cup plant (Silphium perfoliatum L.) as biogas substrate and its implications for the energy and material flows of a large biogas plant. Biofuels, Bioprod. Biorefin. 2020, 14, 152–179. [Google Scholar] [CrossRef] [Green Version]
- Han, K.J.; Albrecht, K.A.; Mertens, D.R.; Kim, D.A. Comparison of in vitro digestion kinetics of cup-plant and alfalfa. Asian-Australasian J. Anim. Sci. 2000, 13, 641–644. [Google Scholar] [CrossRef]
- Cumplido-Marin, L.; Graves, A.R.; Burgess, P.J.; Morhart, C.; Paris, P.; Jablonowski, N.D.; Facciotto, G.; Bury, M.; Martens, R.; Nahm, M. Two Novel Energy Crops: Sida hermaphrodita (L.) Rusby and Silphium perfoliatum L.—State of Knowledge. Agronomy 2020, 10, 928. [Google Scholar] [CrossRef]
- Pavlov, V.S.; Pachomov, I.J.; Špakov, A.P.; Yakimčik, T.V. Produktivnost sil’fii pronzennolistnoj w zavisimosti ot azotnogo udobreniya. Chim. w Selskim Chozjajstwie 1984, 21, 24–26. [Google Scholar]
- Jemielin, V.A.; Šjeluto, B.V. Produktivnosť silfii pronziennolistnoj v zavisimosti ot posledjejstvija udobrjenij, pieriodičnosti podkormok i doz azota na sieďmoj hod žizni posievov. Viestn. Biełorusskoj Hosudarstviennoj Selskochoziajstviennoj Akadjemii 2021, 636.085.51, 107–112. [Google Scholar]
- Šjeluto, B.V.; Kostickaja, J.V. Fotosintjetičjeskaja djejatjelnosť rastjenij silfii pronziennolistnoj. Ispolz. Mieliorirovannych Zemiel 2021, 2, 55–61. [Google Scholar]
- Kuklina, A.G. Introdukcija silfii pronziennolistnoy (Silphium perfoliatum L.) v glavnom Botaničeskom Sadu RAN. Bjulletin Gl. Bot. Sada 1994, 170, 30–33. [Google Scholar]
- Klečkovskaya, M.S.; Muchin, N.G. Introdukcija sil’fii pronzennolistnoy v voronežskoy oblasti. Bjul. Glawnogo Bot. Sada 1990, 155, 12–16. [Google Scholar]
- Ţîţei, V. Biological Peculirities of cup plant (Silphium perfoliatum L.) and utilization possibilities in the republic of Moldova. Lucr. Ştiinţifice Ser. Agron. 2014, 57, 289–293. [Google Scholar]
- Edelšteyn, M.M.; Soloveva, I.W. Sravnitelnaja ocenka kormovych kačestv novych silosnych kultur v usloviyach moskovskoj oblasti. Dokl. TSCHA 1971, 168, 52–56. [Google Scholar]
- Edelšteyn, M.M.; Soloveva, I.W. Soderžanije sacharov v vegetatiwnych organach novych silosnych kultur. Dokl. TSCHA 1975, 209, 51–53. [Google Scholar]
- Maslinkov, M.; Donev, N. Sravnitelno izpitvane na novi mnogogodišni furažni kulturi pri uslovijata na plovdiv. Rastenev’dni Nauk. 1987, 24, 53–58. [Google Scholar]
- Andrejnski, Ž.; Angelow, L.; Andrejnska, I. Proučwane na nowi furažni kulturi. Rastenev’dni Nauk. 1989, 26, 41–45. [Google Scholar]
- Gansberger, M.; Montgomery, L.F.R.; Liebhard, P. Botanical characteristics, crop management and potential of Silphium perfoliatum L. as a renewable resource for biogas production: A review. Ind. Crop. Prod. 2015, 63, 362–372. [Google Scholar] [CrossRef]
- Bedlan, G. Ascochyta silphii sp. nov.—Eine neue Ascochyta-Art an Silphium perfoliatum [Ascochyta silphii sp. nov.—A new Ascochyta species on Silphium perfoliatum]. J. Kult. 2014, 66, 281–283. [Google Scholar]
- Schoo, B.; Kage, H.; Schittenhelm, S. Radiation use efficiency, chemical composition, and methane yield of biogas crops under rainfed and irrigated conditions. Eur. J. Agron. 2017, 87, 8–18. [Google Scholar] [CrossRef]
- Reinert, S.; Hulke, B.S.; Prasifka, J.R. Pest potential of Neotephritis finalis (Loew) on Silphium integrifolium Michx., Silphium perfoliatum L., and interspecific hybrids. Agron. J. 2020, 112, 1462–1465. [Google Scholar] [CrossRef]
- Stefanov, D.; Dimitrova, M.; Sanbev, S. Proučvane na vzmožnostite nyakoi rastenija da bdat otgleždani kato furažni kulturi. Životnov’dni Nauk. 1984, 21, 50–56. [Google Scholar]
- Siwek, H.; Włodarczyk, M.; Możdżer, E.; Bury, M.; Kitczak, T. Chemical Composition and Biogas Formation potential of Sida hermaphrodita and Silphium perfoliatum. Appl. Sci. 2019, 9, 4016. [Google Scholar] [CrossRef] [Green Version]
- Šiaudinis, G.; Karčauskienė, D.; Aleinikovienė, J. Assessment of a single application of sewage sludge on the biomass yield of Silphium perfoliatum and changes in naturally acid soil properties. Zemdirbyste-Agriculture 2019, 106, 213–218. [Google Scholar] [CrossRef] [Green Version]
- Frączek, J.; Mudryk, K.; Wróbel, M. Rożnik przerośnięty Silphium perfoliatum l. Źródło biomasy do produkcji biopaliw stałych. Inżynieria Rolnicza 2011, 6, 21–28. [Google Scholar]
- Lopushniak, V.; Hrytsuliak, G.; Dzhus, G. Bioenergetic assessment of sewage sludge application under Silphium perfoliatum L. on sod-podzolic soils of Precarpathians. Agroecol. J. 2021, 126–134. [Google Scholar] [CrossRef]
- Schittenhelm, S.; Schoo, B.; Schroetter, S. Ertragsphysiologie von Biogaspflanzen: Vergleich von Durchwachsener Silphie, Mais und Luzernegras. J. Kult. 2016, 68, 378–384. [Google Scholar]
- Bury, M.; Możdżer, E.; Kitczak, T.; Siwek, H.; Włodarczyk, M. Yields, Calorific Value and Chemical Properties of Cup Plant Silphium perfoliatum L. Biomass, Depending on the Method of Establishing the Plantation. Agronomy 2020, 10, 851. [Google Scholar] [CrossRef]
- Šiaudinis, G.; Jasinskas, A.; Šlepetiene, A.; Karčauskiene, D. The evaluation of biomass and energy productivity of common mugwort (Artemisia vulgaris L.) and cup plant (Silphium perfoliatum L.) in albeluvisol. Žemdirbystė 2012, 99, 357–362. [Google Scholar]
- Boe, A.; Albrecht, K.A.; Johnson, P.J.; Wu, J. Biomass Production of Monocultures and Mixtures of Cup Plant and Native Grasses on Prime and Marginal Cropland. Am. J. Plant Sci. 2019, 10, 911–924. [Google Scholar] [CrossRef] [Green Version]
- Facciotto, G.; Bury, M.; Chiocchini, F.; Marín, L.C.; Czyż, H.; Graves, A.; Martens, R.; Morhart, C.; Paris, P.; Nahm, M.; et al. Preliminary results regarding yields of Virginia mallow (Sida hermaphrodita (L.) Rusby) and cup plant (Silphium perfoliatum L.) in different condition of Europe. In Proceedings of the Biomass Crops and Energy Grasses, Proceedings of the 27th European Biomass Conference and Exhibition, Lisbon, Portugal, 27–30 May 2019; pp. 350–352. Available online: https://www.researchgate.net/profile/Pierluigi-Paris/publication/334612043_Preliminary_results_regarding_yields_of_Virginia_mallow_Sida_hermaphrodity_L_Rusby_and_cup_plant_Silphium_perfomiatum_L_in_different_conditions_of_Europe/links/5d35c46fa6fdcc370a55e8f3/Preliminary-results-regarding-yields-of-Virginia-mallow-Sida-hermaphrodity-L-Rusby-and-cup-plant-Silphium-perfomiatum-L-in-different-conditions-of-Europe.pdf (accessed on 25 February 2022).
- Cossel, M.; Steberl, K.; Hartung, J.; Pereira, L.A.; Kiesel, A.; Lewandowski, I. Methane yield and species diversity dynamics of perennial wild plant mixtures established alone, under cover crop maize (Zea mays L.), and after spring barley (Hordeum vulgare L.). GCB Bioenergy 2019, 11, 1376–1391. [Google Scholar] [CrossRef] [Green Version]
- Ruidisch, M.; Nguyen, T.T.; Li, Y.L.; Geyer, R.; Tenhunen, J. Estimation of annual spatial variations in forest production and crop yields at landscape scale in temperate climate regions. Ecol. Res. 2015, 30, 279–292. [Google Scholar] [CrossRef]
- Schorpp, Q.; Schrader, S. Earthworm functional groups respond to the perennial energy cropping system of the cup plant (Silphium perfoliatum L.). Biomass Bioenergy 2016, 87, 61–68. [Google Scholar] [CrossRef]
- Boe, A.; Albrecht, K.A.; Johnson, P.J.; Wu, J. Biomass production of cup plant (Silphium perfoliatum L.) in response to variation in plant population density in the North Central USA. Am. J. Plant Sci. 2019, 10, 904–910. [Google Scholar] [CrossRef] [Green Version]
- Franzaring, J.; Holz, I.; Kauf, Z.; Fangmeier, A. Responses of the novel bioenergy plant species Sida hermaphrodita (L.) Rusby and Silphium perfoliatum L. to CO2 fertilization at different temperatures and water supply. Biomass Bioenergy 2015, 81, 574–583. [Google Scholar] [CrossRef]
- Šiaudinis, G. The investigation of three potential energy crops: Common mugwort, cup plant and virginia mallow on Western Lithuania’s Albeluvisol. Appl. Ecol. Environ. Res. 2017, 15, 611–620. [Google Scholar] [CrossRef]
- Ustak, S.; Munoz, J. Cup-plant potential for biogas production compared to reference maize in relation to the balance needs of nutrients and some microelements for their cultivation. J. Environ. Manag. 2018, 228, 260–266. [Google Scholar] [CrossRef]
- Heneman, P.; Červinka, J. Energy crops and bioenergetics in the Czech Republic. Ann. Warsaw Univ. Life Sci. 2007, 51, 73–78. [Google Scholar]
- Dimitrova, M.; Stefanov, D.; Sanbev, S.; Lazarov, W.; Christov, P. Technologija za schranenie na zelenata masa ot cherakleum i silfium. Životnov’dni Nauk. 1987, 24, 56–61. [Google Scholar]
- Glyaubertene, V.F.; Ivanovskaya, K.M. Biologičeskaja i biochimičeskaja charakteristika perspektivnych silosnych rastenij. (22. Soderžanie pitatel’nych veščestv v nadzemnoj masse sil’fii pronzennolistnoy i maral’ego kornja i ich izmenenie vo vremya vegetacii). Liet. TSR Moksl. Akad. Darbai 1987, 4, 110–118. [Google Scholar]
- Rakhmetov, D.B.; Vergun, O.M.; Stadnichuk, N.O.; Shymanska, O.V.; Rakhmetova, S.O.; Fishchenko, V.V. Biochemical study of plant raw material of Silphium L. spp. in M.M. Gryshko National Botanical Garden of the NAS of Ukraine. Plant Introd. 2019, 2019, 80–86. [Google Scholar]
- Bek, T.W.; Anikeenko, A.P. Chimičeskij sostaw zelenoj massy sil’fii pronzennolistnoj i sidy mnogoletnej. Naucz. Tech. Bjulleten Vsesojuznogo Instituta Rastenievod. 1983, 136, 50–52. [Google Scholar]
- Kowalska, G.; Pankiewicz, U.; Kowalski, R. Evaluation of Chemical Composition of Some Silphium L. Species as Alternative Raw Materials. Agriculture 2020, 10, 132. [Google Scholar] [CrossRef] [Green Version]
- Dudkin, M.S.; Černo, N.K.; Škantova, N.G. Charakteristika vodorastvorimych polisacharidov listiev Silphium perfoliatum. Khimiya Prir. Soedin. 1979, 6, 771–774. [Google Scholar]
- Shang, H.-M.; Zhou, H.-Z.; Li, R.; Duan, M.-Y.; Wu, H.-X.; Lou, Y.-J. Extraction optimization and influences of drying methods on antioxidant activities of polysaccharide from cup plant (Silphium perfoliatum L.). PLoS ONE 2017, 12, e0183001. [Google Scholar]
- Witaszek, K.; Herkowiak, M.; Pilarska, A.A.; Czekała, W. Methods of Handling the Cup Plant (Silphium perfoliatum L.) for Energy Production. Energies 2022, 15, 1897. [Google Scholar] [CrossRef]
- Kowalski, R.; Wierciński, J. Ocena niektórych gatunków Silphium jako surowców inulinowych. Ann. Univ. Mariae Curie-Sklodowska Sect. E 2004, 59, 189–195. [Google Scholar]
- Utejš, J.A. Introdukcija i vprovadžennja v kul’turu sil’fii pronizanolistoy. Vestn. AN USSR 1975, 7, 61–65. [Google Scholar]
- Vavilov, P.P.; Edelšteyn, M.M.; Soloveva, I.V. Frakcjonnyj sostav bełka zelenoj massy novych silosnych kultur. Izv. TSCHA 1973, 5, 84–93. [Google Scholar]
- Glyaubertene, V.F. Biologičeskaja i biochimičeskaja charakteristika perspektivnych silosnych rasteniy. (18. Soderžanie i dinamika nakoplinija azotistych veščestv nadzemnoy časti gorca vejricha, okopnika šeršavogo, sil’fii pronzennolistnoy i maral’ego kornja v pieriod vegeta. Liet. TSR Moksl. Akad. Darbai 1986, 2, 101–108. [Google Scholar]
- Nedvaras, A.P.; Marčyuiyonis, V.I. Biologičeskaja i biochimičeskaja charakteristika perspektivnych silosnych rastenij. (20. Rost i razvitie, urožajnost, chimičeskij sostav nadzemnoj časti sil’fii pronzennolistnoy w usloviyach Litovskoy SSR). Liet. TSR Moksl. Akad. Darbai 1987, 3, 37–45. [Google Scholar]
- Lee, M.A. A global comparison of the nutritive values of forage plants grown in contrasting environments. J. Plant Res. 2018, 131, 641–654. [Google Scholar] [CrossRef]
- Glyaubertene, V.F.; Marčyulyonis, V.I. Biologičeskaja i biochimičeskaja charakteristika perspektiwnych silosnych rastenij. (17. Belkowye frakcii nadzemnoj časti gorca Wejricha, okopnika šeršawogo, sil’fii pronzennolistnoj i maral’ego kornja i ich izmenenie wo wremja wegetacii na VI godu wyrašč. Liet. TSR Moksl. Akad. Darbai 1985, 3, 103–109. [Google Scholar]
- Glyaubertene, V.F.; Marčyulyonis, V.I. Biologičeskaja i biochimičeskaja charakteristika perspektivnych silosnych rastenij. (11. Belkovye frakcii nadzemnoj časti gorca vejricha, okopnika šeršawogo, sil’fii pronzennolistnoy i maral’ego kornja i ich izmenenie vo vremja vegetacii). Liet. TSR Moksl. Akad. Darbai 1980, 3, 129–139. [Google Scholar]
- Glyaubertene, V.F.; Marčyulyonis, V.I. Biologičeskaja i biochimičeskaja charakteristika perspektivnych silosnych rastenij. (12. Elektroforez legkorastvorimych belkov nadzemnoj časti gorca vejricha, okopnika šeršavogo, sil’fii pronzennolistnoy i maral’ego kornya v poliakrilamidnom gele na IV go. Liet. TSR Moksl. Akad. Darbai 1981, 1, 115–121. [Google Scholar]
- Glyaubertene, V.F.; Marčyulyonis, V.I. Biologičeskaja i biochimičeskaja charakteristika perspektivnych silosnych rastenij. (16. Elektroforez v poliakrilamidnom gele legkorastvorimych belkov nadzemnoy časti sil’fii pronzennolistnoy i maral’ego kornja na VI godu vyraščivanija). Liet. TSR Moksl. Akad. Darbai 1984, 1, 68–75. [Google Scholar]
- Glyaubertene, V.F.; Ivanovskaya, K.M. Biologičeskaja i biochimičeskaja charakteristika perspektivnych silosnych rasteniy. (21. Soderžanie aminokislot w zelenoj masse gorca vejricha, okopnika šeršawogo, sil’fii pronzennolistnoy i maral’ego kornya w processe ontogeneza). Liet. TSR Moksl. Akad. Darbai 1987, 4, 102–109. [Google Scholar]
- Reinert, S.; Van Tassel, D.L.; Schlautman, B.; Kane, N.C.; Hulke, B.S. Assessment of the biogeographical variation of seed size and seed oil traits in wild Silphium integrifolium Michx. genotypes. Plant Genet. Resour. Charact. Util. 2019, 17, 427–436. [Google Scholar] [CrossRef]
- Edelšteyn, M.M.; Soloveva, I.V. Sodierżanije karotina i askorbinovoj kisloty v silosnych kulturach. Dokl. TSCHA 1974, 204, 51–54. [Google Scholar]
- Glyaubertene, V.F. Biologičeskaja i biochimičeskaja charakteristika perspektivnych silosnych rastenij. (13. Soderžanie karotina i askorbinovoy kisloty w nadzemnoj časti gorca vejricha, okopnika šeršawogo, sil’fii pronzennolistnoy i maral’ego kornja). Liet. TSR Moksl. Akad. Darbai 1982, 1, 81–87. [Google Scholar]
- Wever, C.; Höller, M.; Becker, L.; Biertümpfel, A.; Köhler, J.; van Inghelandt, D.; Westhoff, P.; Pude, R.; Pestsova, E. Towards high-biomass yielding bioenergy crop Silphium perfoliatum L.: Phenotypic and genotypic evaluation of five cultivated populations. Biomass and Bioenergy 2019, 124, 102–113. [Google Scholar] [CrossRef]
- Achakzai, A.K.K.; Masood, A.; Tareen, R.B. Mujeeb-Ur-Rahman Nutrients composition of common plant species of asteraceae in Quetta at two growth stages. J. Chem. Soc. Pakistan 2018, 40, 722–732. [Google Scholar]
- Ignat, I.; Volf, I.; Popa, V.I. A critical review of methods for characterisation of polyphenolic compounds in fruits and vegetables. Food Chem. 2011, 126, 1821–1835. [Google Scholar] [CrossRef]
- Andrade, M.; das Graças Cardoso, M.; de Andrade, J.; Silva, L.; Teixeira, M.; Valério Resende, J.; da Silva Figueiredo, A.; Barroso, J. Chemical Composition and Antioxidant Activity of Essential Oils from Cinnamodendron dinisii Schwacke and Siparuna guianensis Aublet. Antioxidants 2013, 2, 384–397. [Google Scholar] [CrossRef]
- Wojdyło, A.; Oszmiański, J.; Czemerys, R. Antioxidant activity and phenolic compounds in 32 selected herbs. Food Chem. 2007, 105, 940–949. [Google Scholar] [CrossRef]
- Olagaray, K.E.; Bradford, B.J. Plant flavonoids to improve productivity of ruminants—A review. Anim. Feed Sci. Technol. 2019, 251, 21–36. [Google Scholar] [CrossRef]
- Li, A.-N.; Li, S.; Zhang, Y.-J.; Xu, X.-R.; Chen, Y.-M.; Li, H.-B. Resources and Biological Activities of Natural Polyphenols. Nutrients 2014, 6, 6020–6047. [Google Scholar] [CrossRef]
- Şentürk, M.; Gülçin, İ.; Beydemir, Ş.; Küfrevioğlu, Ö.İ.; Supuran, C.T. In vitro inhibition of human carbonic anhydrase I and II isozymes with natural phenolic compounds. Chem. Biol. Drug Des. 2011, 77, 494–499. [Google Scholar] [CrossRef]
- Ostolski, M.; Adamczak, M.; Brzozowski, B.; Stolarski, M.J. Screening of Functional Compounds in Supercritical Carbon Dioxide Extracts from Perennial Herbaceous Crops. Agriculture 2021, 11, 488. [Google Scholar] [CrossRef]
- Wojcińska, M.; Drost-Karbowska, K. Phenolic Acids in Silphium perfoliatum L. flowers (Asteraceae/Compositae). Acta Pol. Pharm. Drug Res. 1998, 55, 413–416. [Google Scholar]
- Kowalski, R. Secondary metabolites in Silphium integrifolium in the first 2 years of cultivation. N. Z. J. Crop Hortic. Sci. 2004, 32, 397–406. [Google Scholar] [CrossRef]
- Kowalski, R.; Wolski, T. Evaluation of phenolic acid content in Silphium perfoliatum L. leaves, inflorescences and rhizomes. Electron. J. Polish Agric. Univ. 2003, 6. Available online: http://www.ejpau.media.pl/volume6/issue1/horticulture/art-03.html (accessed on 25 February 2022).
- Kowalski, R.; Wierciński, J. Phenolic acids in leaves of three Silphium L. species. Polish J. Food Nutr. Sci. 2003, 53, 17–20. [Google Scholar]
- Kowalski, R.; Wolski, T. TLC and HPLC analysis of the phenolic acids in Silphium perfoliatum L. leaves, inflorescences and rhizomes. J. Planar Chromatogr. Mod. TLC 2003, 16, 230–236. [Google Scholar] [CrossRef]
- Robbins, R.J. Phenolic acids in foods: An overview of analytical methodology. J. Agric. Food Chem. 2003, 51, 2866–2887. [Google Scholar] [CrossRef]
- Clifford, M.N. Diet-derived phenols in plasma and tissues and their implications for health. Planta Med. 2004, 70, 1103–1114. [Google Scholar] [CrossRef] [Green Version]
- Tomas-Barberan, F.A.; Clifford, M.N. Dietary hydroxybenzoic acid derivatives—Nature, occurrence and dietary burden. J. Sci. Food Agric. 2000, 80, 1024–1032. [Google Scholar] [CrossRef]
- van Dam, R.M.; Hu, F.B. Coffee consumption and risk of type 2 diabetes. JAMA 2005, 294, 97. [Google Scholar] [CrossRef] [PubMed]
- Scalbert, A.; Manach, C.; Morand, C.; Rémésy, C.; Jiménez, L. Dietary polyphenols and the prevention of diseases. Crit. Rev. Food Sci. Nutr. 2005, 45, 287–306. [Google Scholar] [CrossRef] [PubMed]
- Manach, C.; Mazur, A.; Scalbert, A. Polyphenols and prevention of cardiovascular diseases. Curr. Opin. Lipidol. 2005, 16, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.D.; Wojcińska, M.; Calabria, L.M.; Linse, K.; Clevinger, J.A.; Mabry, T.J. The Flavonoids and Phenolic Acids of the Genus Silphium and Their Chemosystematic Value. Nat. Prod. Commun. 2009, 4, 1934578X0900400. [Google Scholar] [CrossRef] [Green Version]
- Williams, J.D. The Flavonoids and Phenolic Acids of the Genus Silphium and Their Chemosystematic and Medicinal Value; University of Texas at Austin: Austin, TX, USA, 2006. [Google Scholar]
- Zaidi, M.A.; Crow, S.A. Biologically active traditional medicinal herbs from Balochistan, Pakistan. J. Ethnopharmacol. 2005, 96, 331–334. [Google Scholar] [CrossRef]
- Kumar, S.; Pandey, A.K. Chemistry and biological activities of flavonoids: An overview. Sci. World J. 2013, 2013, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Fan, M.; Ding, H.; Zhang, G.; Hu, X.; Gong, D. Relationships of dietary flavonoid structure with its tyrosinase inhibitory activity and affinity. LWT 2019, 107, 25–34. [Google Scholar] [CrossRef]
- Mierziak, J.; Kostyn, K.; Kulma, A. Flavonoids as important molecules of plant interactions with the environment. Molecules 2014, 19, 16240–16265. [Google Scholar] [CrossRef]
- Pérez-Jiménez, J.; Saura-Calixto, F. Fruit peels as sources of non-extractable polyphenols or macromolecular antioxidants: Analysis and nutritional implications. Food Res. Int. 2018, 111, 148–152. [Google Scholar] [CrossRef]
- Ekinci, D.; Karagoz, L.; Ekinci, D.; Senturk, M.; Supuran, C.T. Carbonic anhydrase inhibitors: In vitro inhibition of α isoforms (hCA I, hCA II, bCA III, hCA IV) by flavonoids. J. Enzyme Inhib. Med. Chem. 2013, 28, 283–288. [Google Scholar] [CrossRef]
- El-Sayed, N.H.; Wojcińska, M.; Drost-Karbowska, K.; Matławska, I.; Williams, J.; Mabry, T.J. Kaempferol triosides from Silphium perfoliatum. Phytochemistry 2002, 60, 835–838. [Google Scholar] [CrossRef]
- Feng, W.-S.; Pei, Y.-Y.; Zheng, X.-K.; Li, C.-G.; Ke, Y.-Y.; Lv, Y.-Y.; Zhang, Y.-L. A new kaempferol trioside from Silphium perfoliatum. J. Asian Nat. Prod. Res. 2014, 16, 393–399. [Google Scholar] [CrossRef] [PubMed]
- Kowalski, R. Antimicrobial activity of essential oils and extracts of rosinweed (Silphium trifoliatum and Silphium integrifolium) plants used by the American Indians. Flavour Fragr. J. 2008, 23, 426–433. [Google Scholar] [CrossRef]
- Scur, M.C.; Pinto, F.G.S.; Pandini, J.A.; Costa, W.F.; Leite, C.W.; Temponi, L.G. Antimicrobial and antioxidant activity of essential oil and different plant extracts of Psidium cattleianum Sabine. Brazilian J. Biol. 2016, 76, 101–108. [Google Scholar] [CrossRef] [Green Version]
- Kowalski, R.; Kowalska, G.; Jamroz, J.; Nawrocka, A.; Metyk, D. Effect of the ultrasound-assisted preliminary maceration on the efficiency of the essential oil distillation from selected herbal raw materials. Ultrason. Sonochem. 2015, 24, 214–220. [Google Scholar] [CrossRef]
- Palmieri, S.; Maggio, F.; Pellegrini, M.; Ricci, A.; Serio, A.; Paparella, A.; Lo Sterzo, C. Effect of the Distillation Time on the Chemical Composition, Antioxidant Potential and Antimicrobial Activity of Essential Oils from Different Cannabis sativa L. Cultivars. Molecules 2021, 26, 4770. [Google Scholar] [CrossRef]
- Kowalski, R.; Gagoś, M.; Kowalska, G.; Pankiewicz, U.; Sujka, M.; Mazurek, A.; Nawrocka, A. Effects of ultrasound technique on the composition of different essential oils. J. Anal. Methods Chem. 2019, 2019, 6782495. [Google Scholar] [CrossRef]
- Berka-Zougali, B.; Ferhat, M.-A.; Hassani, A.; Chemat, F.; Allaf, K.S. Comparative Study of Essential Oils Extracted from Algerian Myrtus communis L. Leaves Using Microwaves and Hydrodistillation. Int. J. Mol. Sci. 2012, 13, 4673–4695. [Google Scholar] [CrossRef] [Green Version]
- Guzmán, E.; Lucia, A. Essential Oils and Their Individual Components in Cosmetic Products. Cosmetics 2021, 8, 114. [Google Scholar] [CrossRef]
- Riad, N.; Zahi, M.R.; Bouzidi, N.; Daghbouche, Y.; Touafek, O.; El Hattab, M. Occurrence of Marine Ingredients in Fragrance: Update on the State of Knowledge. Chemistry 2021, 3, 1437–1463. [Google Scholar] [CrossRef]
- Burt, S. Essential oils: Their antibacterial properties and potential applications in foods—A review. Int. J. Food Microbiol. 2004, 94, 223–253. [Google Scholar] [CrossRef] [PubMed]
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological effects of essential oils—A review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef] [PubMed]
- Fung, T.K.H.; Lau, B.W.M.; Ngai, S.P.C.; Tsang, H.W.H. Therapeutic Effect and Mechanisms of Essential Oils in Mood Disorders: Interaction between the Nervous and Respiratory Systems. Int. J. Mol. Sci. 2021, 22, 4844. [Google Scholar] [CrossRef] [PubMed]
- Wolski, T.; Kowalski, R.; Mardarowicz, M. Chromatographic analysis of essential oil occuring in inflorescences, leaves and rhizomes of Silphium perfoliatum L. Herba Pol. 2000, 46, 235–242. [Google Scholar]
- Kowalski, R.; Wierciński, J.; Mardarowicz, M. Essential oil in leaves and inflorescences of Silphium integrifolium michx. J. Essent. Oil Res. 2005, 17, 220–222. [Google Scholar] [CrossRef]
- Kowalski, R.; Wolski, T. The chemical composition of essential oils of Silphium perfoliatum L. Flavour Fragr. J. 2005, 20, 306–310. [Google Scholar] [CrossRef]
- Kowalski, R. Chemical composition of essential oils and lipophilic extracts of Silphium integrifolium and S. trifoliatum inflorescences. Chem. Nat. Compd. 2008, 44, 241–244. [Google Scholar] [CrossRef]
- Lawson, S.K.; Sharp, L.G.; Powers, C.N.; McFeeters, R.L.; Satyal, P.; Setzer, W.N. Volatile Compositions and Antifungal Activities of Native American Medicinal Plants: Focus on the Asteraceae. Plants 2020, 9, 126. [Google Scholar] [CrossRef] [Green Version]
- Kowalski, R.; Wawrzykowski, J. Essential oils analysis in dried materials and granulates obtained from Thymus vulgaris L., Salvia officinalis L., Mentha piperita L. and Chamomilla recutita L. Flavour Fragr. J. 2009, 24, 31–35. [Google Scholar] [CrossRef]
- Tomaino, A.; Cimino, F.; Zimbalatti, V.; Venuti, V.; Sulfaro, V.; De Pasquale, A.; Saija, A. Influence of heating on antioxidant activity and the chemical composition of some spice essential oils. Food Chem. 2005, 89, 549–554. [Google Scholar] [CrossRef]
- Sangwan, N.S.; Farooqi, A.H.A.; Shabih, F.; Sangwan, R.S. Regulation of essential oil production in plants. Plant Growth Regul. 2001, 34, 3–21. [Google Scholar] [CrossRef]
- Filly, A.; Fabiano-Tixier, A.S.; Louis, C.; Fernandez, X.; Chemat, F. Water as a green solvent combined with different techniques for extraction of essential oil from lavender flowers. C. R. Chim. 2016, 19, 707–717. [Google Scholar] [CrossRef]
- El Asbahani, A.; Jilale, A.; Voisin, S.N.; Aït Addi, E.H.; Casabianca, H.; El Mousadik, A.; Hartmann, D.J.; Renaud, F.N.R. Chemical composition and antimicrobial activity of nine essential oils obtained by steam distillation of plants from the Souss-Massa Region (Morocco). J. Essent. Oil Res. 2015, 27, 34–44. [Google Scholar] [CrossRef]
- Hristova, Y.; Wanner, J.; Jirovetz, L.; Stappen, I.; Iliev, I.; Gochev, V. Chemical composition and antifungal activity of essential oil of Hyssopus officinalis L. from Bulgaria against clinical isolates of Candida species. Biotechnol. Biotechnol. Equip. 2015, 29, 592–601. [Google Scholar] [CrossRef] [Green Version]
- Kowalski, R. Analysis of lipophilic fraction from leaves, inflorescences and rhizomes of Silphium perfoliatum L. Acta Soc. Bot. Pol. 2005, 74, 5–10. [Google Scholar] [CrossRef] [Green Version]
- Kowalski, R. Silphium L. extracts–composition and protective effect on fatty acids content in sunflower oil subjected to heating and storage. Food Chem. 2009, 112, 820–830. [Google Scholar] [CrossRef]
- Bohlmann, F.; Jakupovic, J. Neue labdan-derivate und sesquiterpene aus Silphium-arten. Phytochemistry 1979, 18, 1987–1992. [Google Scholar] [CrossRef]
- Bohlmann, F.; Jakupovic, J. Neue Sesquiterpen-Kohlenwasserstoffe mit anomalen Kohlenstoffgerüst aus Silphium-arten. Phytochemistry 1980, 19, 259–265. [Google Scholar] [CrossRef]
- Pcolinski, M.J.; Doskotch, R.W.; Lee, A.Y.; Clardy, J. Chlorosilphanol A and Silphanepoxol, Labdane Diterpenes from Silphium, perfoliatum. J. Nat. Prod. 1994, 57, 776–783. [Google Scholar] [CrossRef]
- Davidyants, E.S.; Putieva, Ż.M.; Bandyukova, W.A.; Abubakirov, N.K. Triterpenovye glikozidy Silphium perfoliatum. Khimiya Prir. Soedin. 1984, 1, 120–121. [Google Scholar]
- Davidyants, E.S.; Putieva, Ż.M.; Bandyukova, W.A.; Abubakirov, N.K. Triterpenovye glikozidy Silphium perfoliatum II. Khimiya Prir. Soedin. 1984, 5, 666–667. [Google Scholar]
- Davidyants, E.S.; Putieva, Ż.M.; Bandyukova, W.A.; Abubakirov, N.K. Triterpenovye glikozidy Silphium perfoliatum III. Strojenie sil’fiozida E. Khimiya Prir. Soedin. 1984, 6, 750–753. [Google Scholar]
- Davidyants, E.S.; Putieva, Ż.M.; Šaškov, A.S.; Bandyukova, W.A.; Abubakirov, N.K. Triterpenovye glikozidy Silphium perfoliatum IV. Chim. Prirod Soed 1985, 4, 519–522. [Google Scholar]
- Davidyants, E.S.; Putieva, Ż.M.; Bandyukova, W.A.; Abubakirov, N.K. Triterpenovye glikozidy Silphium perfoliatum V. Stroenie sil’fiozida A. Khimiya Prir. Soedin. 1986, 1, 63–66. [Google Scholar]
- Rogowska, A.; Szakiel, A. Enhancement of Phytosterol and Triterpenoid Production in Plant Hairy Root Cultures—Simultaneous Stimulation or Competition? Plants 2021, 10, 2028. [Google Scholar] [CrossRef] [PubMed]
- Kowalski, R. Ocena zawartości oleanozydów w organach nadziemnych i podziemnych rożnika przerośniętego Silphium perfoliatum L. Acta Sci. Pol. Hortorum Cultus 2002, 1, 5–15. [Google Scholar]
- Kowalski, R. Studies of selected plant raw materials as alternative sources of triterpenes of oleanolic and ursolic acid types. J. Agric. Food Chem. 2007, 55, 656–662. [Google Scholar] [CrossRef]
- Podolak, I.; Galanty, A.; Sobolewska, D. Saponins as cytotoxic agents: A review. Phytochem. Rev. 2010, 9, 425–474. [Google Scholar] [CrossRef] [Green Version]
- Khakimov, B.; Tseng, L.; Godejohann, M.; Bak, S.; Engelsen, S. Screening for Triterpenoid Saponins in Plants Using Hyphenated Analytical Platforms. Molecules 2016, 21, 1614. [Google Scholar] [CrossRef] [Green Version]
- Calabria, L.M. The Isolation and Characterization of Triterpene Saponins from Silphium and the Chemosystematic and Biological Significance of Saponins in the Asteraceae; University of Texas at Austin: Austin, TX, USA, 2008. [Google Scholar]
- Han, L.-K.; Zheng, Y.-N.; Yoshikawa, M.; Okuda, H.; Kimura, Y. Anti-obesity effects of chikusetsusaponins isolated from Panax japonicus rhizomes. BMC Complement. Altern. Med. 2005, 5, 9. [Google Scholar] [CrossRef] [Green Version]
- Densmore, F. Uses of Plants by the Chippewa Indians; U.S. G.P.O.: Washington, DC, USA, 1928; Volume 44. [Google Scholar]
- Smith, H.H. Etnobotany of the Ojibwe Indians. Bull. Public Museum Milwaukee 1932, 4, 365. [Google Scholar]
- Herrick, J.W. Iroquois Medical Botany; Syracuse University Press: Syracuse, NY, USA, 1977. [Google Scholar]
- Gilmore, M.R. A study in the ethnobotany of the Omaha Indians. Nebraska State Hist. Soc. Collect. 1913, 17, 314–357. [Google Scholar]
- Gilmore, M.R. Uses of Plants by the Indians of the Missouri River Region; Bison Books: Lincoln, NB, USA, 1919; Volume 47. [Google Scholar]
- Grieve, M. A Modern Herbal. Electric Newt.. 1995. Available online: http://www.botanical.com/botanical/mgmh/c/cuppl129.html (accessed on 25 February 2022).
- Kuyanceva, A.M.; Davidyants, E.S. Regenerirujuščaja aktivnost’ ekstrakta Silphium perfoliatum. Farm. Moskwa 1988, 6, 36–37. [Google Scholar]
- Syrov, W.N.; Khushabaktova, Z.A.; Davidyants, E.S. Triterpenowye glikozidy Silphium perfoliatum L. Gipolipidemičeskaja aktiwnost’ sil’fiozida. Khimiko-Farmatsevticheskii Zhurnal 1992, 26, 66–69. [Google Scholar]
- Davidyants, E.S. Triterpene Glycosides of Silphium perfoliatum L: Structure, Biological Activity, Possibility of Use; Stavropol State Agrarian University: Stavropol, Russia, 2017. [Google Scholar]
- Kindscher, K.; Manfredi, K.P.; Britton, M.; Demidova, M.; Phurlburt, D.P. Testing prairie plants with ethnobotanical importance for anti-cancer and anti-aids compounds. J. Ethnobiol. 1998, 18, 229–245. [Google Scholar]
- Calabria, L.M.; Piacente, S.; Kapusta, I.; Dharmawardhane, S.F.; Segarra, F.M.; Pessiki, P.J.; Mabry, T.J. Triterpene saponins from Silphium radula. Phytochemistry 2008, 69, 961–972. [Google Scholar] [CrossRef] [PubMed]
- Blay, G.; García, B.; Molina, E.; Pedro, J.R. Syntheses of (+)-Alismoxide and (+)-4- e pi -Alismoxide. J. Org. Chem. 2006, 71, 7866–7869. [Google Scholar] [CrossRef]
- Davidyants, E.S.; Kartaševa, I.A.; Nešin, I.W. Vliyanie triterpenovych glikozidov Silphium perfoliatum L. na fitopatogennye griby. Rastit. Resur. 1997, 4, 93–98. [Google Scholar]
- Zabka, M.; Pavela, R.; Gabrielova-Slezakova, L. Promising antifungal effect of some Euro-Asiatic plants against dangerous pathogenic and toxinogenic fungi. J. Sci. Food Agric. 2011, 91, 492–497. [Google Scholar] [CrossRef]
- Jamiołkowska, A.; Kowalski, R. In vitro estimate of influence of Silphium perfoliatum L. leaves extract on some fungi colonizing the pepper plants. Acta Sci. Pol. Hortorum Cultus 2012, 11, 43–55. [Google Scholar]
- Davidyants, E.S. The effect of purified amount of triterpene glycosides and their enriched extract from leaves of Silphium perfoliatum L. on the germination of seeds of winter wheat and the activity of catalase in them. Chem. Plant Raw Mater. 2021, 2, 353–360. [Google Scholar] [CrossRef]
- Belyak, W.B. Mnogoletniye silosnye kultury dlya kormoproizvodstwa. Wiestnik Sielkochoziaisttwiennych Nauk 1981, 5, 66–70. [Google Scholar]
- Han, K.J.; Albrecht, K.A.; Muck, R.E.; Kim, D.A. Moisture effect on fermentation characteristics of cup-plant silage. Asian-Australasian J. Anim. Sci. 2000, 13, 636–640. [Google Scholar] [CrossRef]
- Varlamova K., A. Obogaščene kul’turnoj flory novymi vidami kormovych rastieniy. Vestn. Sielkochoziaisttviennych Nauk 1984, 4, 87–96. [Google Scholar]
- Mamedov, T.G.; Golubev, N.I. Vyraščivat’ na silos vygodno. Kormoproizvodstwo 1987, 7, 45–46. [Google Scholar]
- Piłat, P.; Majtkowski, W.; Majtkowska, G.; Mikołajczak, J.; Góralska, A. The usefulness for ensiling of chosen plant forms of species of Silphium genus. J. Cent. Eur. Agric. 2007, 8, 363–368. [Google Scholar]
- Lehmkuhler, J.W.; Ramos, M.H.; Albrecht, K.A. Cupplant Silage as a Replacement for Corn Silage in Growing Beef Cattle Diets. Forage Grazinglands 2007, 5, 1–6. [Google Scholar] [CrossRef]
- Bernas, J.; Bernasová, T.; Gerstberger, P.; Moudrý, J.; Konvalina, P.; Moudrý, J. Cup plant, an alternative to conventional silage from a LCA perspective. Int. J. Life Cycle Assess. 2021, 26, 311–326. [Google Scholar] [CrossRef]
- Han, K.-J.; Albrecht, K.A. Substitution Value of Cup Plant (Silphium perfoliatum L.) Silage in Dairy Cow Diet. J. Agric. Sci. 2021, 13, 1–10. [Google Scholar] [CrossRef]
- Weymar, H. Buch der Korbblüter; Neumann Verlag: Leipzig, Germany, 1969. [Google Scholar]
- Gardiner, M.A.; Tuell, J.K.; Isaacs, R.; Gibbs, J.; Ascher, J.S.; Landis, D.A. Implications of three biofuel crops for beneficial arthropods in agricultural landscapes. BioEnergy Res. 2010, 3, 6–19. [Google Scholar] [CrossRef] [Green Version]
- Werling, B.P.; Dickson, T.L.; Isaacs, R.; Gaines, H.; Gratton, C.; Gross, K.L.; Liere, H.; Malmstrom, C.M.; Meehan, T.D.; Ruan, L.; et al. Perennial grasslands enhance biodiversity and multiple ecosystem services in bioenergy landscapes. Proc. Natl. Acad. Sci. USA 2014, 111, 1652–1657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jabłoński, B.; Kołtowski, Z. Nectar secretion and honey potential of honey-plants growing under Poland’s conditions—Part XV. J. Apic. Sci. 2005, 49, 59–63. [Google Scholar]
- Mueller, A.L.; Berger, C.A.; Schittenhelm, S.; Stever-Schoo, B.; Dauber, J. Water availability affects nectar sugar production and insect visitation of the cup plant Silphium perfoliatum L. (Asteraceae). J. Agron. Crop Sci. 2020, 206, 529–537. [Google Scholar] [CrossRef] [Green Version]
- Radu Sumalan, L.; Muntean, C.; Kostov, A.; Kržanović, D.; Noemi Jucsor, L.; Sorin Ciulca, I.; Renata Sumalan, M.; Gheju, M.; Cernicova-Buca, M. The cup plant (Silphium perfoliatum L.)—A viable solution for bioremediating soils polluted with heavy metals. Not. Bot. Horti Agrobot. Cluj-Napoca 2020, 48, 2095–2113. [Google Scholar] [CrossRef]
- Klimont, K.; Bulińska-Radomska, Z.; Górka, J.; Osińska, A. Badanie przydatności rożnika przerośniętego (Silphium perfoliatum L.) do rekultywacji terenów po otworowej eksploatacji złóż siarki. Probl. Inżynierii Rol. 2015, 23, 61–74. [Google Scholar]
- Antonkiewicz, J.; Jasiewicz, C. Effect of soil contamination with heavy metals on element contents in Silphium perfoliatum L. Chem. Inżynieria Ekol. 2003, 10, 205–210. [Google Scholar]
- Moll, L.; Höller, M.; Hubert, C.; Korte, C.A.C.; Völkering, G.; Wever, C.; Pude, R. Cup Plant (Silphium perfoliatum L.) Biomass as Substitute for Expanded Polystyrene in Bonded Leveling Compounds. Agronomy 2022, 12, 178. [Google Scholar] [CrossRef]
- Höller, M.; Lunze, A.; Wever, C.; Deutschle, A.L.; Stücker, A.; Frase, N.; Pestsova, E.; Spiess, A.C.; Westhoff, P.; Pude, R. Meadow hay, Sida hermaphrodita (L.) Rusby and Silphium perfoliatum L. as potential non-wood raw materials for the pulp and paper industry. Ind. Crops Prod. 2021, 167, 113548. [Google Scholar] [CrossRef]
- Du, J.; Zhang, L.; Ali, A.; Li, R.; Xiao, R.; Guo, D.; Liu, X.; Zhang, Z.; Ren, C.; Zhang, Z. Research on thermal disposal of phytoremediation plant waste: Stability of potentially toxic metals (PTMs) and oxidation resistance of biochars. Process Saf. Environ. Prot. 2019, 125, 260–268. [Google Scholar] [CrossRef]
- Conrad, M.; Biertümpfel, A.; Vetter, A. Durchwachsene Silphie (Silphium perfoliatum L.)—Von der Futterpflanze zum Koferment F.N.R. Gülzower Fachgespräche. In Proceedings of the 2nd Symposium Energiepflanzen, Fachagentur Nachwachsende Rohstoffe; Fachagentur Nachwachsende Rohstoffe, Berlin, Germany, 17–18 November 2009; pp. 281–289. [Google Scholar]
- Aurbacher, J.; Bull, I.; Formowitz, B.; Gurgel, A.; Heiermann, M.; Heilmann, H.; Herrmann, C.; Idler, C.; Jänicke, H.; Kornatz, P.; et al. Energiepflanzen für Biogasanlagen. Available online: https://www.fnr.de/fileadmin/Projekte/2021/Mediathek/fnr_brosch.energiepflanzen-mv_web.pdf (accessed on 25 February 2022).
- Aurbacher, J.; Benke, M.; Formowitz, B.; Glauert, T.; Heiermann, M.; Herrmann, C.; Idler, C.; Kornatz, P.; Nehring, A.; Rieckmann, C.; et al. Energiepflanzen für Biogasanlagen. Available online: https://www.fnr.de/fileadmin/Projekte/2021/Mediathek/brosch.energiepflanzen-niedersachsen-webpdf_1.pdf (accessed on 25 February 2022).
- Stolzenburg, K.; Monkos, A. Erste Versuchsergebnisse mit der Durchwachsenen Silphie (Silphium perfoliatum L.) in Baden-Württemberg; Landwirtschaftliches Technologiezentrum Augustenberg: Karlsruhe, Germany, 2012; Available online: https://www.landwirtschaft-bw.info/pb/site/pbs-bw-mlr/get/documents_E545218469/MLR.LEL/PB5Documents/ltz_ka/Arbeitsfelder/Nachwachsende%20Rohstoffe/Kulturinformationen/Silphie_DL/Erste%20Versuchsergebnisse%20mit%20der%20Durchwachsenen%20Silphie%20in%20Baden-W%C3%BCrttemberg%20aktualisiert%202012-08.pdf?attachment=true (accessed on 25 February 2022).
- Slepetys, J.; Kadziuliene, Z.; Sarunaite, L.; Tilvikiene, V.; Kryzeviciene, A. Biomass potential of plants grown for bioenergy production. In Proceedings of the International Scientific Conference Renewable Energy and Energy Efficiency; Rivža, P., Rivža, S., Eds.; Latvia University of Agriculture: Jelgava, Latvia, 2012; pp. 66–72. [Google Scholar]
- Mast, B.; Lemmer, A.; Oechsner, H.; Reinhardt-Hanisch, A.; Claupein, W.; Graeff-Hönninger, S. Methane yield potential of novel perennial biogas crops influenced by harvest date. Ind. Crops Prod. 2014, 58, 194–203. [Google Scholar] [CrossRef]
- Biertümpfel, A.; Köhler, J.; Müller, R. Neues von der Silphie. Biogas J. 2015, 18, 42–43. [Google Scholar]
- Dauber, J.; Müller, A.L.; Schittenhelm, S.; Schoo, B.; Schorpp, Q.; Schrader, S.; Schroetter, S. Schlussbericht zum Vorhaben: Thema: Agrarökologische Bewertung der Durchwachsenen Silphie (Silphium perfoliatum L.) als eine Biomassepflanze der Zukunft; Teilvorhaben 1: Ober- und Unterirdische Biodiversität in Beständen der Durchwachsenen Silphie; Tei. Available online: https://literatur.thuenen.de/digbib_extern/dn056633.pdf (accessed on 25 February 2022).
- Sanderson, M.; Adler, P. Perennial Forages as Second Generation Bioenergy Crops. Int. J. Mol. Sci. 2008, 9, 768–788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ende, L.M.; Knöllinger, K.; Keil, M.; Fiedler, A.J.; Lauerer, M. Possibly Invasive New Bioenergy Crop Silphium perfoliatum: Growth and Reproduction Are Promoted in Moist Soil. Agriculture 2021, 11, 24. [Google Scholar] [CrossRef]
- Ţîţei, V. The evaluation of biomass of the Sida hermaphrodita and Silphium perfoliatum for renewable energi in Moldova. Sci. Pap. Ser. A Agron. 2017, 60, 534–540. [Google Scholar]
- Majtkowski, W.; Piłat, J.; Szulc, P. Perspektywy uprawy i wykorzystania w Polsce rożnika przerosnietego Silphium perfoliatum L. Biul. Inst. Hod. i Aklim. Roślin 2009, 251, 283–291. [Google Scholar]
- Jasinskas, A.; Kleiza, V.; Streikus, D.; Domeika, R.; Vaiciukevičius, E.; Gramauskas, G.; Valentin, M.T. Assessment of Quality Indicators of Pressed Biofuel Produced from Coarse Herbaceous Plants and Determination of the Influence of Moisture on the Properties of Pellets. Sustainability 2022, 14, 1068. [Google Scholar] [CrossRef]
- Eurostat. Energy, Transport and Environment Indicators; Publications Office of the European Union: Luxembourg, 2014; ISBN 9789279412561. [Google Scholar]
- Arce, M.; Saavedra, Á.; Míguez, J.; Granada, E.; Cacabelos, A. Biomass fuel and combustion conditions selection in a fixed bed combustor. Energies 2013, 6, 5973–5989. [Google Scholar] [CrossRef] [Green Version]
- Mohammed, M.A.A.; Salmiaton, A.; Wan Azlina, W.A.K.G.; Mohammad Amran, M.S.; Fakhru’l-Razi, A.; Taufiq-Yap, Y.H. Hydrogen rich gas from oil palm biomass as a potential source of renewable energy in Malaysia. Renew. Sustain. Energy Rev. 2011, 15, 1258–1270. [Google Scholar] [CrossRef]
- Brendieck-Worm, C. Kurskorrektur in der Biogasbranche möglich? Zeitschrift für Ganzheitliche Tiermedizin 2017, 31, 50–53. [Google Scholar] [CrossRef]
- Grunwald, D.; Panten, K.; Schwarz, A.; Bischoff, W.; Schittenhelm, S. Comparison of maize, permanent cup plant and a perennial grass mixture with regard to soil and water protection. GCB Bioenergy 2020, 12, 694–705. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Species | Botanical Varieties | Synonyms | Common Name | Natural Occurence |
|---|---|---|---|---|---|
| 1 | Silphium albiflorum Gray | – * | – | White Rosinweed |  |
| 2 | Silphium asteriscus L. | var. angustatum Gray var.asteriscus var. laevicaule DC | Silphium angustum (Gray) Small, Silphium dentatum var. angustatum (Gray) Perry Silphium asteriscus var. scabrum Nutt. Silphium asteriscus var. dentata (Ell.) Chapman, Silphium dentatum Ell., Silphium elliottii Small, Silphium incisum Greene, Silphium nodum Small | Starry Rosinweed |  |
| 3 | Silphium brachiatum Gattinger | – | – | Cumberland Rosinweed |  |
| 4 | Silphium compositum Michx. | var. compositum var. ovatifolium Torr. & Gray var. reniforme (Raf. ex Nutt.) Torr. & Gray var. venosum (Small) Kartesz & Gandhi | Silphium orae Small Silphium compositum ssp. ovatifolium (Torr. & Gray) Sweeny & T.R. Fisher, Silphium ovatifolium (Torr. & Gray) Small Silphium compositum ssp. reniforme (Raf. ex Nutt.) Sweeny & T.R. Fisher, Silphium reniforme Raf. ex Nutt., Silphium terebinthinaceum Ell. Silphium compositum ssp. venosum (Small) Sweeny & T.R. Fisher, Silphium lapsuum Small; Silphium venosum Small | Kidney-Leaf Rosinweed |  |
| 5 | Silphium gracile Gray | – | Silphium simpsonii var. wrightii Perry | Slender Rosinweed |  |
| 6 | Silphium integrifolium Michx. | var. deamii Perry var. gattingeri Perry var. integrifolium var. laeve Torr. & Gray var. neglectum Settle & T.R. Fisher | – – Silphium laevigatum Pursh Silphium speciosum Nutt. – – | Entire-Leaf Rosinweed |  |
| 7 | Silphium laciniatum L. | var. laciniatum var. robinsonii Perry | – – | Compassplant Mohr’s Rosinweed |  |
| 8 | Silphium mohrii Small | – | – | * | |
| 9 | Silphium perfoliatum L. | var. connatum (L.) Cronq. var. perfoliatum | Silphium connatum L., Silphium perfoliatum ssp. connatum (L.) Cruden, Silphium scabrum Moench | Cup Plant |  |
| 10 | Silphium pinnatifidum Ell. | – | Silphium chickamaugense Canby, Silphium terebinthinaceum var. pinnatifidum (Ell.) Gray | Tansy Rosinweed | * |
| 11 | Silphium radula Nutt. | – | Silphium asperrimum Hook., Silphium dentatum var. gatesii (C. Mohr) Ahles, Silphium gatesii C. Mohr | Rough-Stem Rosinweed |  |
| 12 | Silphium reverchonii Bush | – | – | Reverchon’s Rosinweed | * |
| 13 | Silphium scaberrimum Ell. | – | – | Rough-Leaf Rosinweed | * |
| 14 | Silphium simpsonii Greene | – | – | Simpson’s Rosinweed | * |
| 15 | Silphium terebinthinaceum Jacq. | var. luciae–brauniae Steyermark var. terebinthinaceum | Silphium rumicifolium Small | Prairie Rosinweed |  |
| 16 | Silphium trifoliatum L. | var. latifolium Gray var. trifoliatum | Silphium confertifolium Small; Silphium glabrum Eggert ex Small Silphium atropurpureum Retz. ex Willd., Silphium laevigatum Ell. | Whorled Rosinweed |  |
| 17 | Silphium wasiotense M. Medley | – | – | Appalachian Rosinweed | * |
| Yields [t DM/ha] | Reference | ||
|---|---|---|---|
| Phase of Spring Re-Growth | Budding Phase | Blooming Phase | |
| S. perfoliatum | |||
| 2.9 | 36.6 | [7] | |
| 5.4–24.7 | 8.0–32.9 | 27.7–36.6 | [59,74,80] |
| 7.7–23.0 * | [19,56,57] | ||
| 9.6–22.3 | [28] | ||
| 19.1–20.6 | [81] | ||
| 8.8–17.6 | [82] | ||
| 14–19 | [83] | ||
| 20.5–22.4 | [84] | ||
| 10.8–16.1 | [85] | ||
| 9.3–18.1 | [86] | ||
| 4.5–8.5 | [87] | ||
| 7.4–10.8 | [88] | ||
| 9.5–26.6 | [89] | ||
| 11.5–22 | [87] | ||
| 17.3–27.8 | [90] | ||
| 12.7–23.3 | [91] | ||
| 13–18 | [92] | ||
| 11.7 –16.8 | [23] | ||
| 13.4–21.7 | [81] | ||
| 11.4–21.9 | [87] | ||
| 10.8–14.3 | [93] | ||
| 11.5 | [94] | ||
| 13.5 | [95] | ||
| 15.4 | [96] | ||
| 11.2 | [97] | ||
| 15.4 | [96] | ||
| S. trifoliatum | |||
| 2.4 | 28.8 | [26] | |
| S. integrifolium | |||
| 2.2 | 19.8 | [27] | |
| Phenolic Acid | Content of Phenolic Acids (mg/100 g DM) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Leaves | Inflorescences | Rhizomes | |||||||
| FPA | AH | BH | FPA | AH | BH | FPA | AH | BH | |
| S. perfoliatum | |||||||||
| Hydroxycinnamic acids (HCAs) | |||||||||
| Caffeic | 2.20–3.09 | 0.18–0.26 | 9.47–12.87 | 2.57 | 0.30 | 13.22 | 4.21 | 1.06 | 12.67 |
| p-Coumaric | 0.35–0.48 | – | 1.01–1.74 | 0.09 | – | 0.17 | 0.48 | – | – |
| Ferulic | – | – | – | 0.33 | – | 0.13 | 0.19 | – | – |
| Σ HCAs | 2.55–3.57 | 0.18–0.26 | 11.21–13.88 | 2.98 | 0.30 | 13.52 | 4.87 | 1.06 | 12.67 |
| Hydroxybenzoic acids (HBAs) | |||||||||
| Protocatechuic | 1.54–2.16 | 0.35–0.44 | 0.53–1.26 | 2.13 | 0.83 | 0.91 | 0.15 | 0.05 | 0.05 |
| p-Hydroxybenzoic | 0.83–1.11 | – | – | 1.35 | 0.17 | 0.17 | 0.14 | – | – |
| Vanilic | 0.77–1.09 | – | – | 1.22 | 0.13 | – | – | – | – |
| Σ HBAs | 3.14–4.36 | 0.35–0.44 | 0.53–1.26 | 4.70 | 1.13 | 1.09 | 0.29 | 0.05 | 0.05 |
| Depsides (Ds) | |||||||||
| Chlorogenic | – | 0.18 | – | – | – | – | – | – | – |
| Σ Ds | – | 0.18 | – | – | – | – | – | – | – |
| Σ (HCAs + HBAs + Ds) | 5.69–7.93 | 0.61–0.79 | 12.47–14.41 | 7.68 | 1.43 | 14.61 | 5.16 | 1.11 | 12.72 |
| S. trifoliatum | |||||||||
| Hydroxycinnamic acids (HCAs) | |||||||||
| Caffeic | 0.97–3.96 | 0.78–1.12 | 2.33–9.44 | 1.21–5.84 | – | 15.99–35.88 | 1.53–1.54 | 0.08–0.54 | 6.46–6.76 |
| p-Coumaric | 0.14–0.37 | – | 0.12–1.35 | 0.15–0.21 | – | 0.13–0.31 | 0.25–1.31 | – | – |
| Ferulic | 0.01–0.05 | – | – | 0.28–0.54 | – | 0.09–0.10 | – | – | – |
| Σ HCAs | 1.17–4.383 | 0.78–1.12 | 2.45–10.79 | 1.97–6.27 | – | 16.21–36.29 | 1.79–2.84 | 0.08–0.54 | 6.46–6.76 |
| Hydroxybenzoic acids (HBAs) | |||||||||
| Protocatechuic | 0.22–1.65 | 0.50–1.61 | 0.01–1.09 | 1.97–6.86 | 1.23–5.27 | 1.23–2.18 | 0.16–039 | 0.01 | – |
| p-Hydroxybenzoic | 0.48–0.89 | – | – | 1.79–2.08 | 0.12–0.20 | – | 0.07–0.09 | – | – |
| Vanilic | – | – | – | 1.86–1.87 | 0.30–0.31 | – | – | – | – |
| Salicylic | 0.82–0.96 | – | – | 1.28–4.81 | – | – | – | – | – |
| Σ HBAs | 1.49–3.09 | 0.50–1.61 | 0.01–1.09 | 7.20–15.32 | 1.74–6.14 | 1.23–2.18 | 0.23–0.48 | 0.01 | – |
| Σ (HCAs + HBAs) | 2.73–7.02 | 1.28–2.47 | 2.54–11.88 | 13.47–17.29 | 1.74–6.14 | 18.39–37.52 | 2.02–3.32 | 0.09–0.55 | 6.46–6.76 |
| S. integrifolium | |||||||||
| Hydroxycinnamic acids (HCAs) | |||||||||
| Caffeic | 0.41–1.95 | 1.21–2.04 | 1.24–6.27 | 2.38–2.49 | 1.52–1.70 | 18.14–23.80 | 1.70–1.96 | – | 3.50–4.43 |
| p-Coumaric | 0.35–0.76 | – | 0.64–1.47 | 0.57–2.00 | – | 1.00–1.24 | 0.09–0.28 | – | – |
| Ferulic | 0.01–0.04 | 0.01–0.04 | 0.01–0.27 | 0.88–1.04 | – | 0.22–1.18 | 0.43–0.59 | – | – |
| Σ HCAs | 0.77–2.72 | 1.21–2.08 | 1.97–8.01 | 3.83–5.53 | 1.52–1.70 | 19.36–26.22 | 2.22–2.83 | – | 3.50–4.43 |
| Hydroxybenzoic acids (HBAs) | |||||||||
| Protocatechuic | 5.44–13.04 | 3.97–9.22 | 1.73–6.56 | 4.31–6.22 | 3.58–4.03 | 2.83–3.09 | 0.17–0.27 | – | 0.12–0.17 |
| p-Hydroxybenzoic | 0.40–0.87 | 0.17–0.40 | 0.08–0.48 | 1.21–2.20 | 0.52–0.87 | 0.65–0.72 | 0.09–0.14 | – | – |
| Vanilic | 0.29–0.87 | – | – | 1.36–3.30 | 1.03–1.41 | – | 0.26 | – | – |
| Σ HBAs | 6.19–14.50 | 4.14–9.62 | 1.81–7.04 | 6.88–11.72 | 5.13–6.31 | 3.55–3.74 | 0.41 | – | 0.12–0.17 |
| Σ (HCAs + HBAs) | 6.96–17.22 | 5.49–11.64 | 3.85–13.30 | 10.71–17.25 | 6.83–7.83 | 23.10–29.77 | 2.48–3.24 | – | 3.62–4.60 |
| Benzoic Acids | |
| p-Hydroxybenzoic Protocatechuic Isovanilic Gallic Ellagic | S. compositum, S. terebinthinaceum, S. laciniatum, S. abliflorum, S. perfoliatum, S. wasiotense, S. integrifolium, S. radula, S. morhii, S. brachiatum, S. asteriscus |
| Vanilic | S. laciniatum, S. abliflorum, S. perfoliatum, S. wasiotense, S. integrifolium, S. radula, S. morhii, S. asteriscus |
| Syringic | S. compositum, S. laciniatum, S. perfoliatum, S. wasiotense, S. integrifolium, S. radula, S. morhii, S. asteriscus |
| Salicylic | S. terebinthinaceum, S. laciniatum, S. abliflorum, S. perfoliatum, S. wasiotense, S. integrifolium, S. radula, S. morhii, S. brachiatum, S. asteriscus |
| Cinnamic acids | |
| Chlorogenic, Rosmarinic | S. compositum, S. terebinthinaceum, S. laciniatum, S. abliflorum, S. perfoliatum, S. wasiotense, S. integrifolium, S. radula, S. morhii, S. brachiatum, S. asteriscus |
| p-Coumaric | S. compositum, S. terebinthinaceum, S. laciniatum, S. abliflorum, S. perfoliatum, S. wasiotense, S. integrifolium, S. morhii, S. brachiatum, S. asteriscus |
| Caffeic | S. compositum, S. terebinthinaceum, S. laciniatum, S. abliflorum, S. wasiotense, S. radula, S. morhii, S. brachiatum, S. asteriscus |
| Hydrocaffeic | S. terebinthinaceum, S. laciniatum, S. abliflorum, S. perfoliatum, S. integrifolium, S. radula, S. brachiatum, S. asteriscus |
| Ferulic | S. laciniatum, S. morhii |
| Isoferulic | S. laciniatum, S. abliflorum |
| m-Coumaric | S. terebinthinaceum, S. laciniatum, S. abliflorum, S. wasiotense, S. integrifolium, S. morhii |
| Flavonoids Detected in Leaf Extracts | COM * | TER | LAC | ALB | PER | WAS | INT | RAD | MOR | BRA | AST |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Silphium Section Composita | Silphium Section Silphium | ||||||||||
| Kaempferol 3–O–β–D–glucopyranoside | + | + | + | + | + | ||||||
| Kaempferol 3–O–β–D–galactopyranoside | + | + | + | ||||||||
| Kaempferol 3–O–robinobioside | + | + | + | + | + | ||||||
| Kaempferol 3–O–rutinoside | + | + | + | + | + | + | + | ||||
| Kaempferol 3–O–β–D–apioside | + | + | |||||||||
| Kaempferol 3–O–β–L–apiosyl–(1″″→6″′)–O–β–D–glucopyranoside | + | ||||||||||
| Kaempferol 3–O–β–D–apiofuranoside 7–O–α–L–rhamnosyl (1″″→6″′)–O–β–D–galactopyranoside | + | + | + | + | |||||||
| Kaempferol 3–O–β–D–apiofuranoside 7–O–α–L–rhamnosyl (1″″→6″′)–O–β–D (2″′–O–E–caffeoylgalactopyranoside) | + | + | + | ||||||||
| Quercetin 3–O–β–D–glucopyranoside | + | + | + | + | + | + | + | ||||
| Quercetin 3–O–β–D–galactopyranoside | + | + | + | + | + | + | + | + | + | + | + |
| Quercetin 3–O–robinobioside | + | + | + | + | |||||||
| Quercetin 3–O–rutinoside | + | + | + | + | + | + | |||||
| Quercetin 3–O–β–D–apioside | + | ||||||||||
| Quercetin 3–O–α–L–rhamnosyl (1′″→6″)–O–β–D–glucoside | + | + | |||||||||
| Quercetin 3–O–α–L–rhamnosyl (1′″→6″)–O–β–D–galactoside | + | + | |||||||||
| Quercetin 3–O–α–D–rhamnosyl 7–O–β–L–apiofuranoside | + | ||||||||||
| Quercetin 3–O–α–L–rhamnosyl (1′″→6″)–O–β–D–galactopyranoside 7–O–β–L–apiofuranoside | + | + | + | + | + | + | |||||
| Quercetin 3–O–β–L–galactosyl (1′″→6″)–O–β–D–rhamnopyranoside 7–O–α–L–apiofuranoside | + | ||||||||||
| Isorhamnetin 3–O–β–D–glucopyranoside | + | + | + | + | |||||||
| Isorhamnetin 3–O–β–D–galactopyranoside | + | + | + | ||||||||
| Isorhamnetin 3–O–robinobioside | + | + | + | ||||||||
| Isorhamnetin 3–O–rutinoside | + | + | |||||||||
| Isorhamnetin 3–O–α–L–rha (1′″→6″)–O–β–D–galactoside | + | + | + | ||||||||
| Isorhamnetin 3–O–α–L–rha (1′″→6″)–O–β–D–glucoside | + | + | |||||||||
| Isorhamnetin 3–O–α–L–rhamnosyl (1′″→6″)–O–β–D–galactopyranoside 7–O–β–L–apiofuranoside | + | + | + | + | + | + | + | ||||
| Total number of flavonoids detected in leaf extracts | 5 | 9 | 8 | 10 | 9 | 7 | 9 | 10 | 10 | 4 | 13 |
| Flavonoids Content (% DM) | |||
|---|---|---|---|
| Plant development stage | |||
| Leaves of 1-year-old plants | |||
| S. perfoliatum | S. trifoliatum | S. integrifolium | |
| Leaf rosette | 0.58 | 0.64 | 0.80 |
| Leaves of 2-year-old plants | |||
| Spring re-growth | 0.89 | 0.87 | 1.19 |
| Flower buds | 0.94 | 0.88 | 1.04 |
| Beginning of flowering | 0.85 | 0.79 | 1.02 |
| Full flowering and fructification | 0.80 | 0.72 | 0.95 |
| Inflorescences of 2-year-old plants | |||
| Beginning of flowering | 0.15 | 0.13 | 0.32 |
| Full flowering and fructification | 0.15 | 0.12 | 0.32 |
| Rhizomes of 2-year-old plants | |||
| Fructification | tr. | tr. | tr. |
| Oleanolic Acid/Oleanosides Content (% DM) | |||
|---|---|---|---|
| Plant development stage | |||
| Leaves of 1-year-old plants | |||
| S. perfoliatum | S. trifoliatum | S. integrifolium | |
| Leaf rosette | 1.27/2.46 | 1.21/2.34 | 1.18/2.28 |
| Leaves of 2-year-old plants | |||
| Spring re-growth | 3.02/5.82 | 3.10/6.02 | 2.06/3.99 |
| Flower buds | 1.58/3.04 | 1.82/3.52 | 1.80/3.49 |
| Beginning of flowering | 1.80/3.48 | 1.54/2.99 | 1.76/3.4 |
| Full flowering and fructification | 1.61/3.11 | 1.39/2.69 | 1.32/2.56 |
| Inflorescences of 2-year-old plants | |||
| Beginning of flowering | 1.94/3.75 | 0.95/1.84 | 2.39/4.64 |
| Full flowering and fructification | 1.98/3.82 | 0.93/1.81 | 2.34/4.54 |
| Rhizomes of 2-year-old plants | |||
| Fructification | 0.91/1.75 | 0.97/1.88 | 1.25/2.42 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kowalska, G.; Baj, T.; Kowalski, R.; Hanif, M.A. Characteristics of Selected Silphium Species as Alternative Plants for Cultivation and Industry with Particular Emphasis on Research Conducted in Poland: A Review. Sustainability 2022, 14, 5092. https://doi.org/10.3390/su14095092
Kowalska G, Baj T, Kowalski R, Hanif MA. Characteristics of Selected Silphium Species as Alternative Plants for Cultivation and Industry with Particular Emphasis on Research Conducted in Poland: A Review. Sustainability. 2022; 14(9):5092. https://doi.org/10.3390/su14095092
Chicago/Turabian StyleKowalska, Grażyna, Tomasz Baj, Radosław Kowalski, and Muhammad Asif Hanif. 2022. "Characteristics of Selected Silphium Species as Alternative Plants for Cultivation and Industry with Particular Emphasis on Research Conducted in Poland: A Review" Sustainability 14, no. 9: 5092. https://doi.org/10.3390/su14095092
APA StyleKowalska, G., Baj, T., Kowalski, R., & Hanif, M. A. (2022). Characteristics of Selected Silphium Species as Alternative Plants for Cultivation and Industry with Particular Emphasis on Research Conducted in Poland: A Review. Sustainability, 14(9), 5092. https://doi.org/10.3390/su14095092