
Impact of Bioaugmentation on the Bioremediation of Saline-Produced Waters Supplemented with Anaerobic Digestate
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
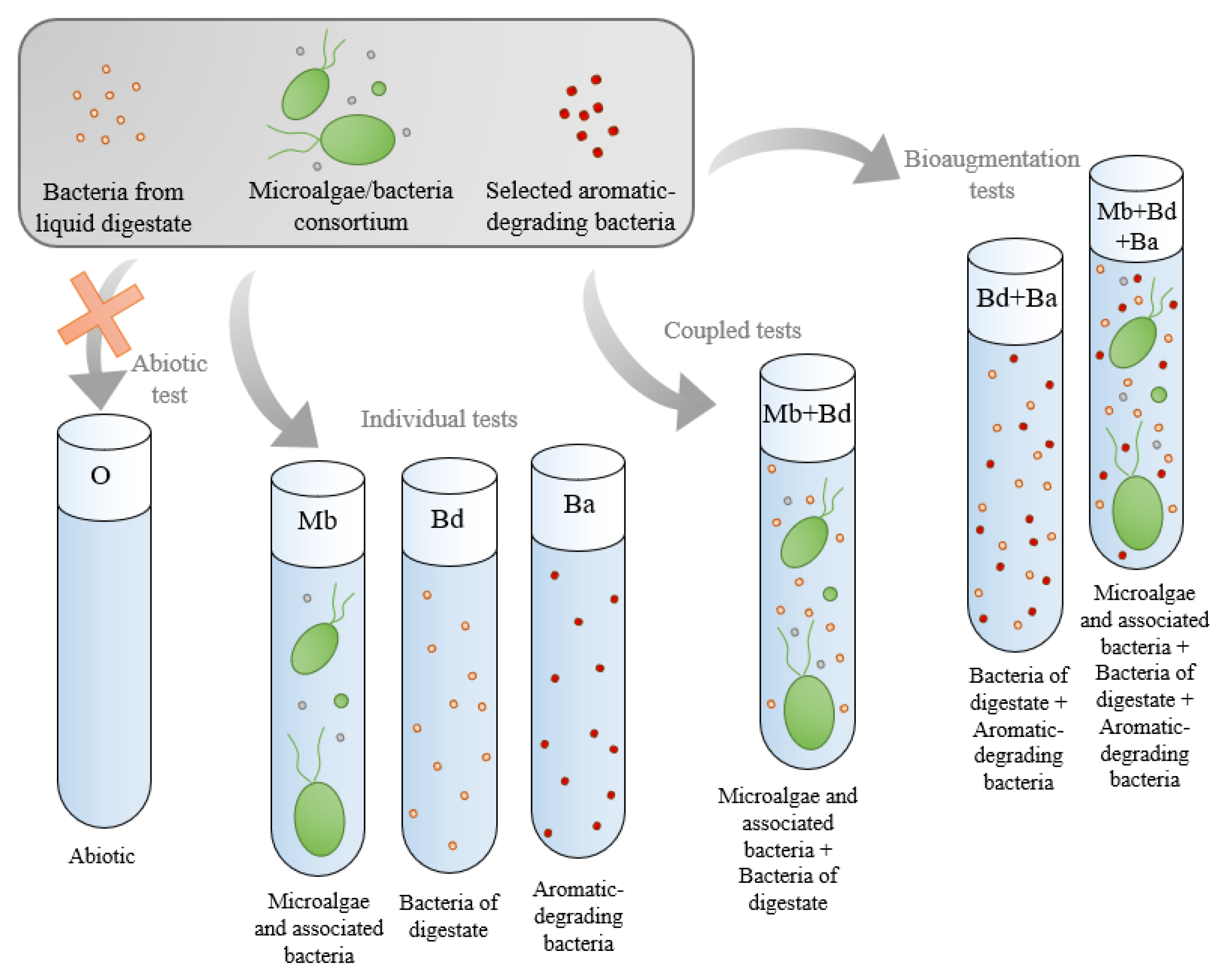
2. Material and Methods
2.1. Artificial-Produced Water Supplemented with Liquid Digestate as Culture Medium
2.2. Strains and Cultivation for Inoculum Preparation
2.3. Growth Test and Monitoring
2.4. Microbial Diversity Monitoring
2.4.1. Sampling, Nucleic Acid Extraction, and RNA Reverse Transcription
2.4.2. PCR Amplification
2.4.3. Sequencing and Bioinformatics Analyses
2.5. Dissolved Compounds Monitoring
3. Results and Discussion
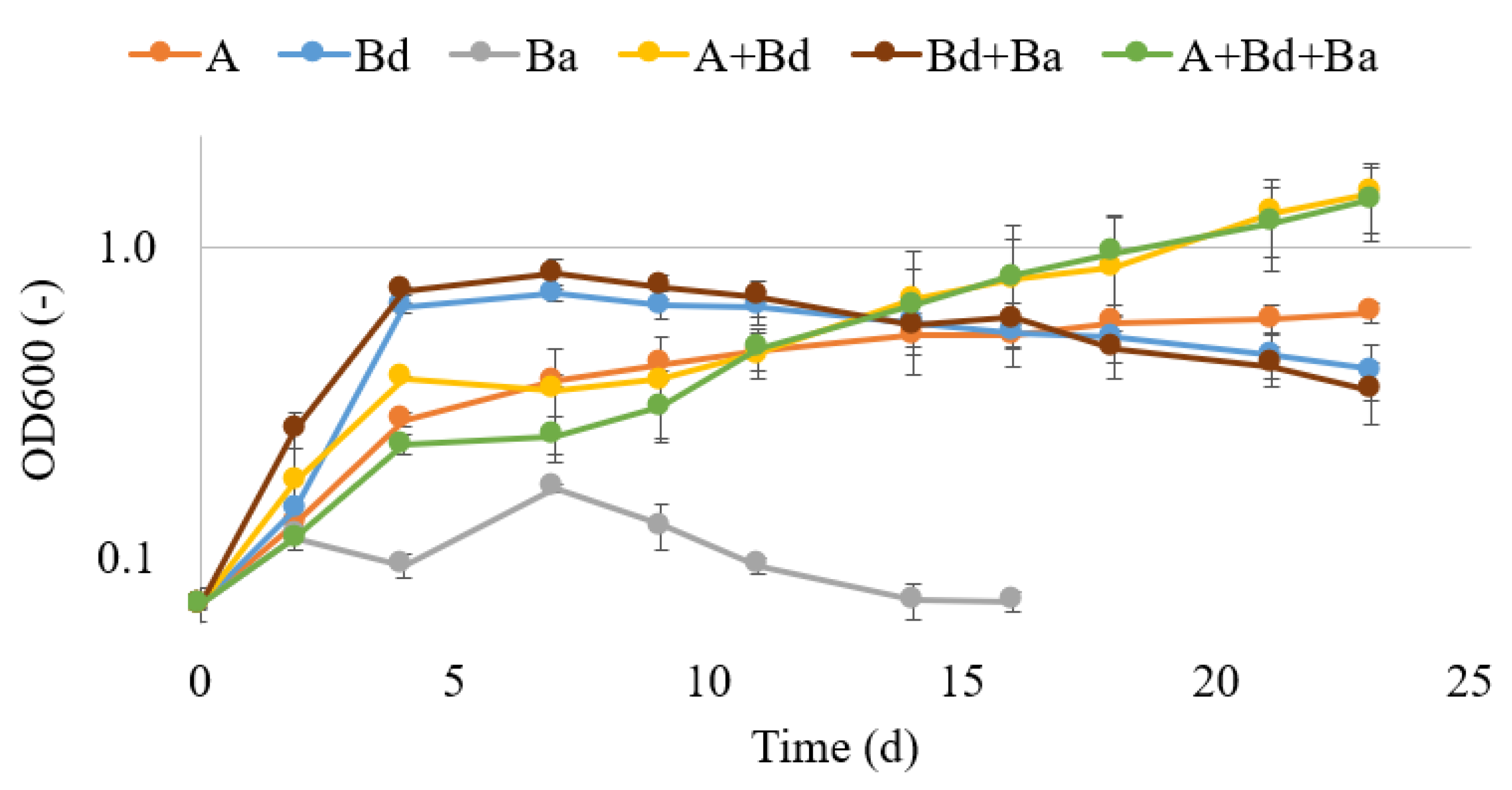
3.1. Microbial Growth
3.2. Microbial Diversity
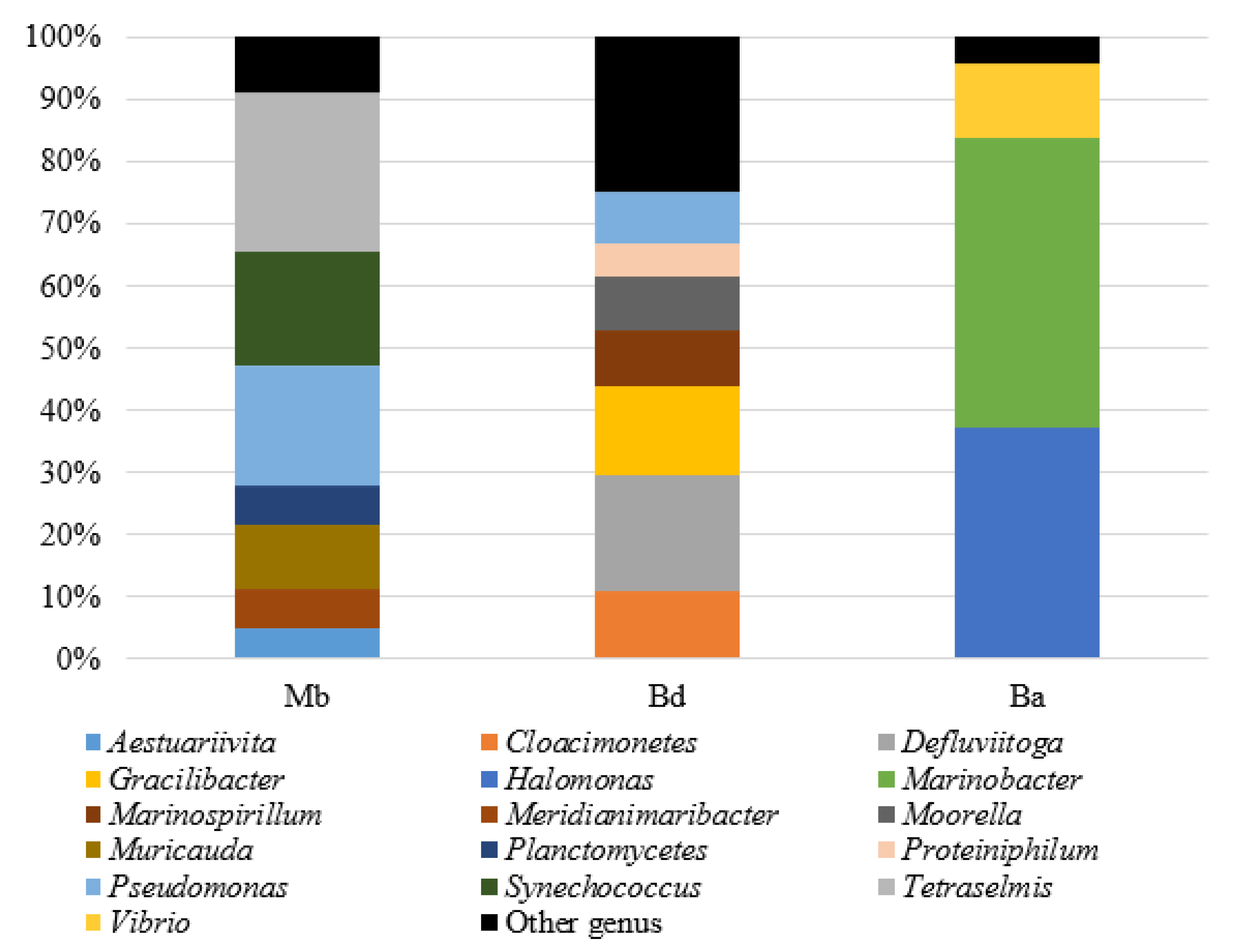
3.2.1. Initial Populations
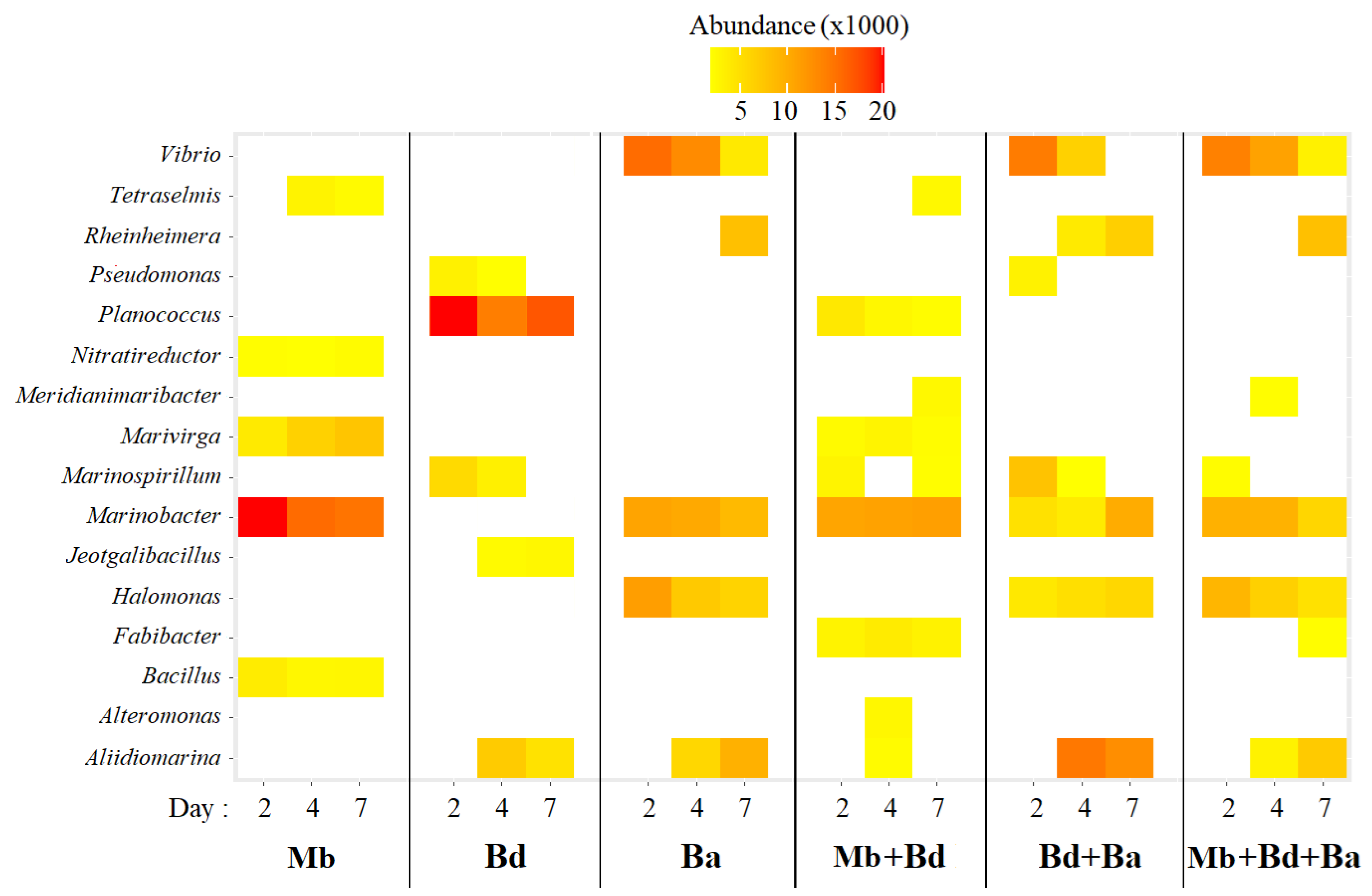
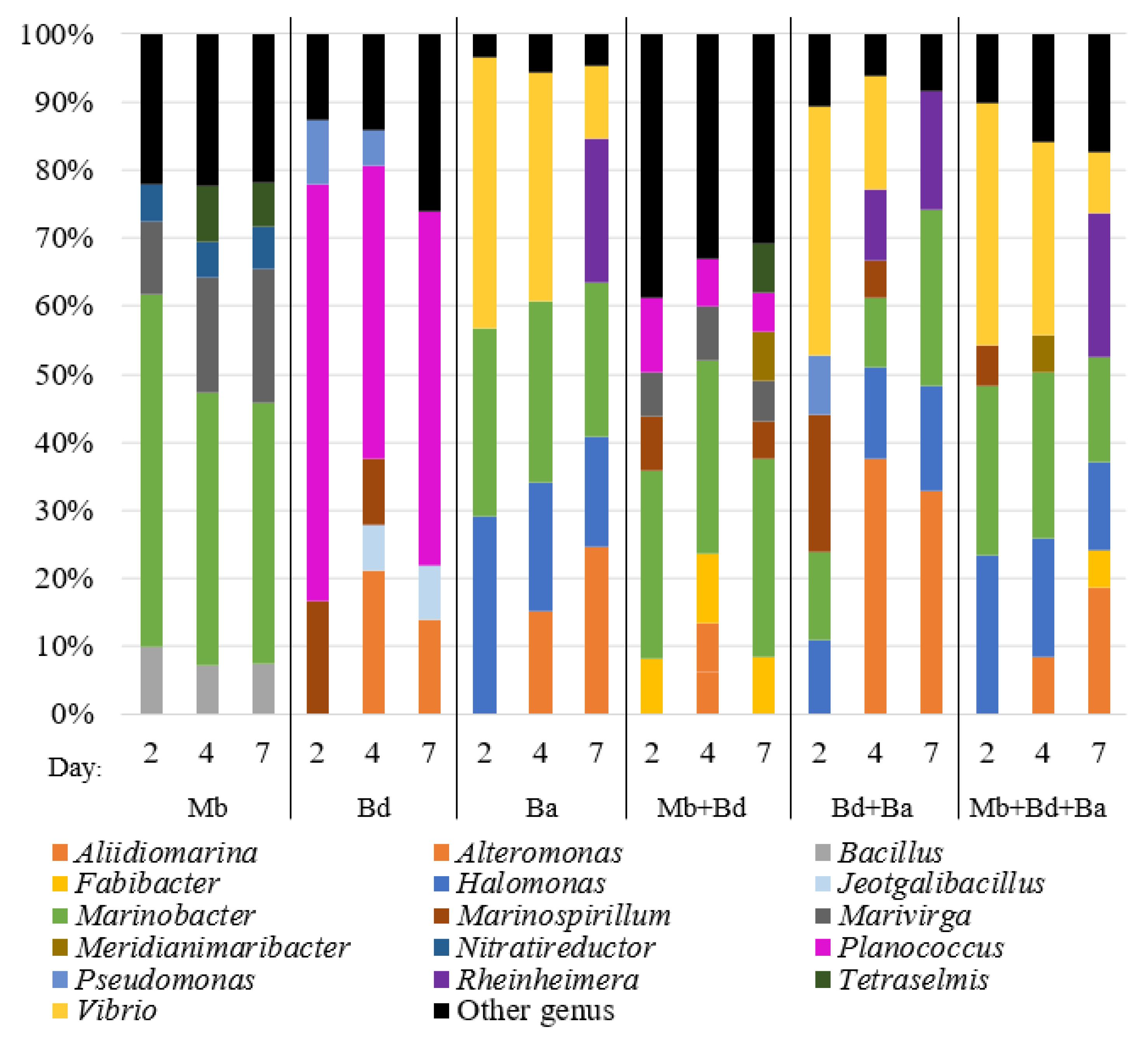
3.2.2. Active Populations
3.3. Bioremediation of Artificial-Produced Water Supplemented with Liquid Digestate
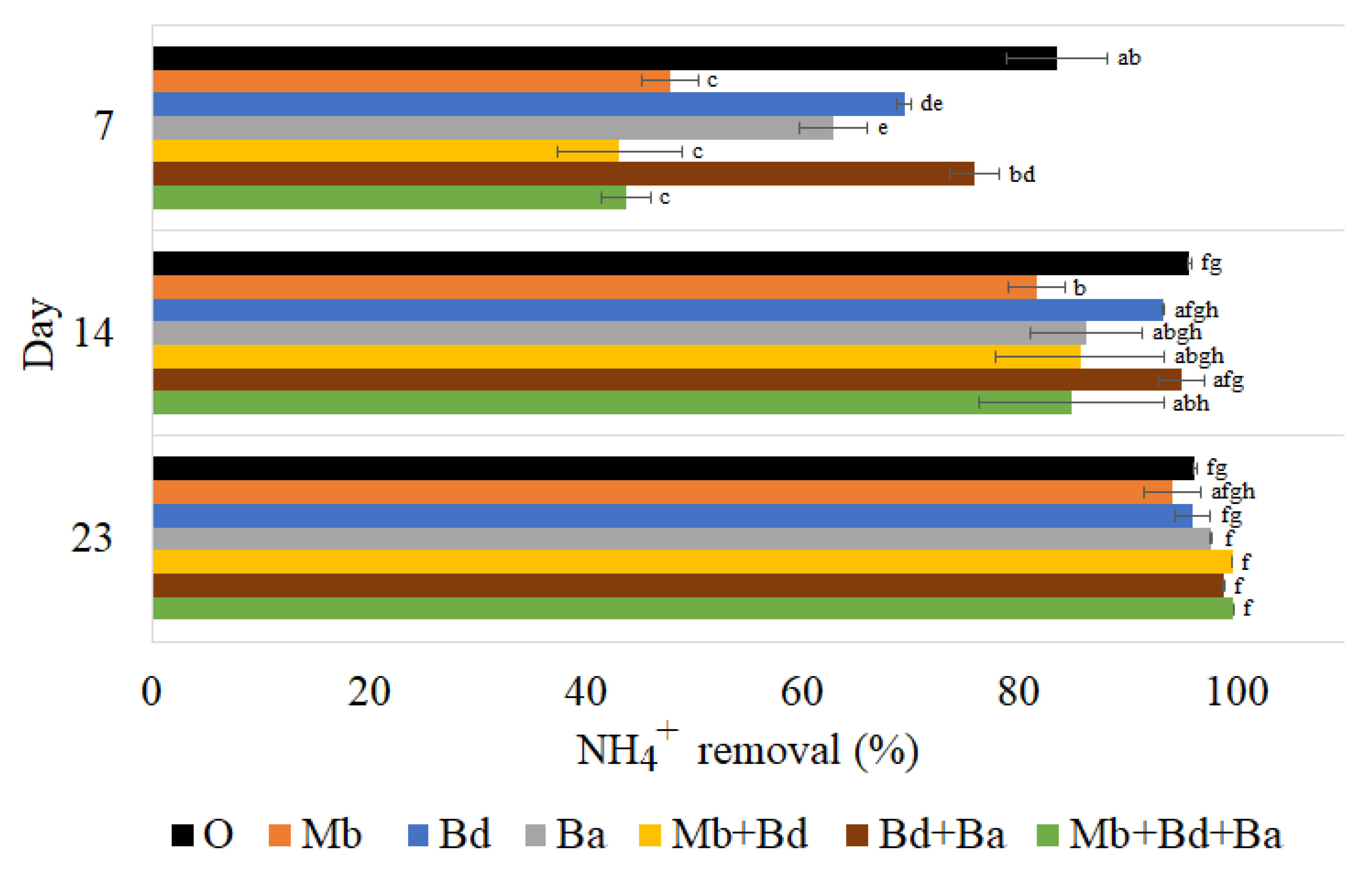
3.3.1. Ammonium Removal
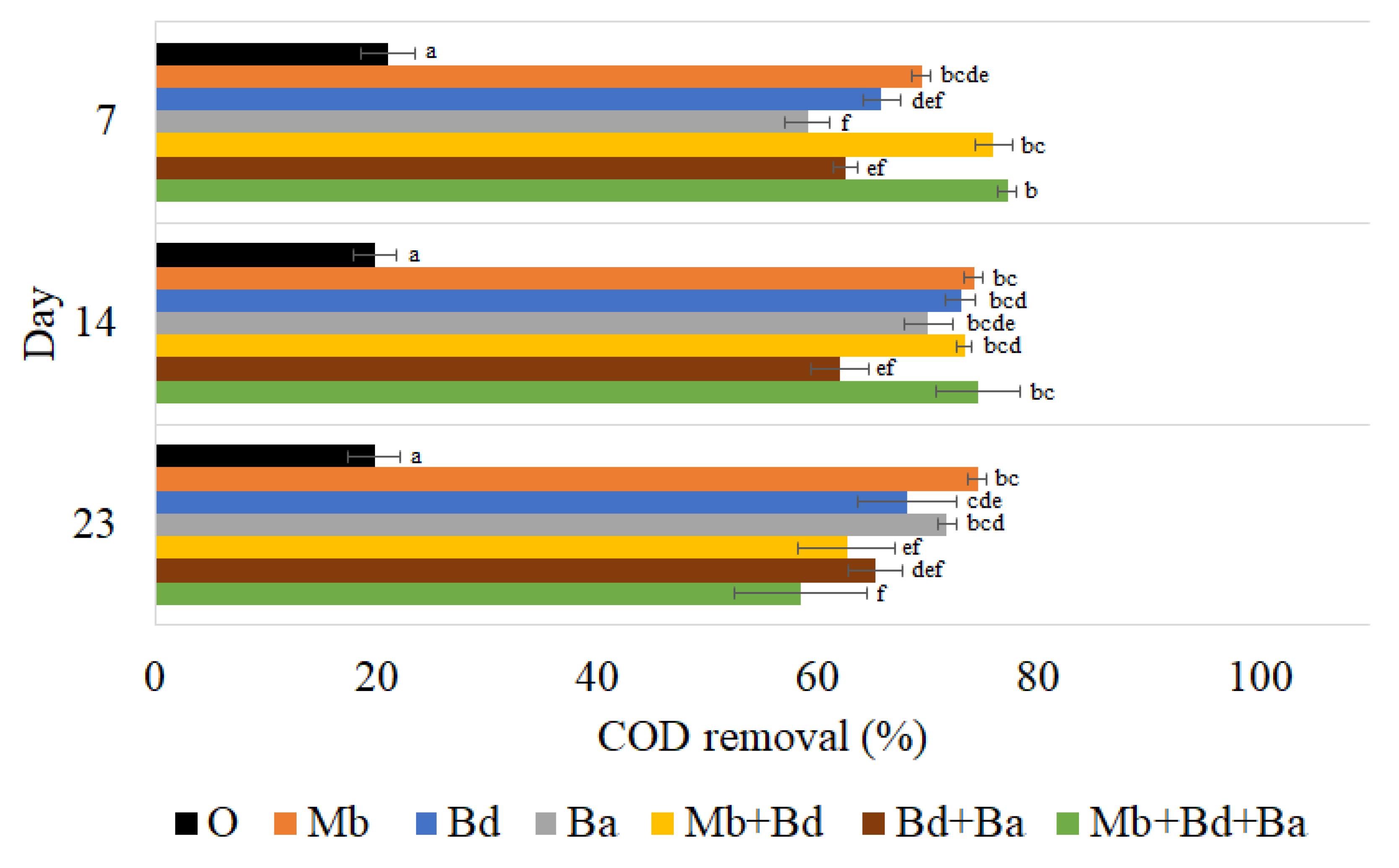
3.3.2. Chemical Oxygen Demand Removal
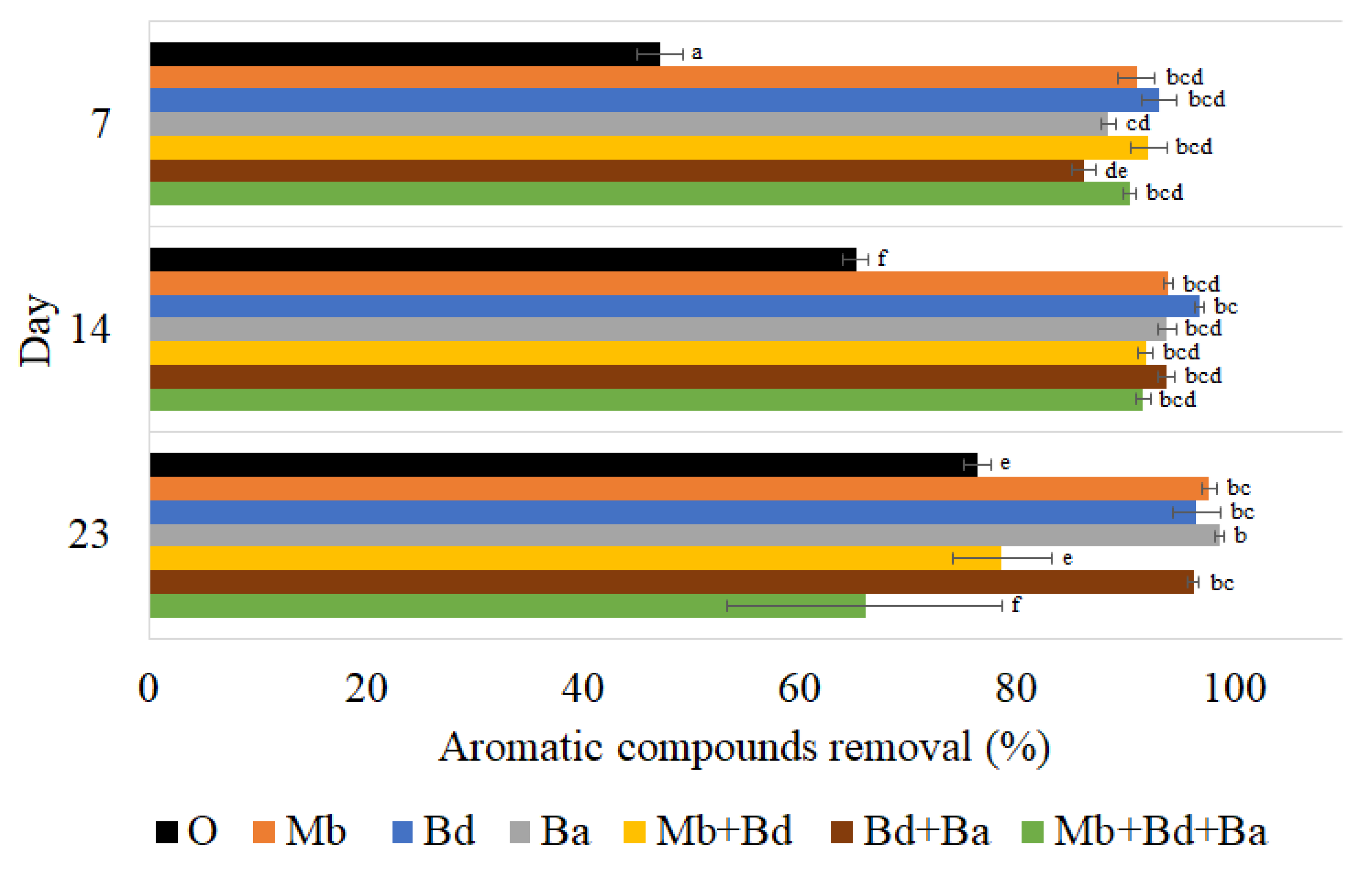
3.3.3. Aromatic Compounds Removal
3.3.4. Bioremediation of Saline-Produced Water Supplemented with Anaerobic Digestate
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- Al-Ghouti, M.A.; Al-Kaabi, M.A.; Ashfaq, M.Y.; Da’Na, D.A. Produced water characteristics, treatment and reuse: A review. J. Water Process. Eng. 2019, 28, 222–239. [Google Scholar] [CrossRef]
- Veil, J.A.; Puder, M.G.; Elcock, D.; Redweik, R.J., Jr. A White Paper Describing Produced Water from Production of Crude Oil, Natural Gas, and Coal Bed Methane; Argonne National Laboratory: Lemont, IL, USA, 2004. [Google Scholar]
- Lusinier, N.; Seyssiecq, I.; Sambusiti, C.; Jacob, M.; Lesage, N.; Roche, N. Biological Treatments of Oilfield Produced Water: A Comprehensive Review. SPE J. 2019, 24, 2135–2147. [Google Scholar] [CrossRef]
- Berberoglu, H.; Gomez, P.S.; Pilon, L. Radiation characteristics of Botryococcus braunii, Chlorococcum littorale, and Chlorella sp. used for CO2 fixation and biofuel production. J. Quant. Spectrosc. Radiat. Transf. 2009, 110, 1879–1893. [Google Scholar] [CrossRef]
- Gerbens-Leenes, W.; de Vries, G.J.; Xu, L. The water footprint of biofuels from microalgae. In Bioenergy and Water 2013, 191–200. [Google Scholar] [CrossRef]
- Linares, L.C.F.; Falfán, K.G.; Ramírez-López, C. Microalgal Biomass: A Biorefinery Approach. In Biomass Volume Estimation and Valorization for Energy; IntechOpen: London, UK, 2017. [Google Scholar] [CrossRef] [Green Version]
- Duran, S.K.; Kumar, P.; Sandhu, S.S. A review on microalgae strains, cultivation, harvesting, biodiesel conversion and engine implementation. Biofuels 2021, 12, 91–102. [Google Scholar] [CrossRef]
- Xia, A.; Murphy, J.D. Microalgal Cultivation in Treating Liquid Digestate from Biogas Systems. Trends Biotechnol. 2016, 34, 264–275. [Google Scholar] [CrossRef]
- Monlau, F.; Sambusiti, C.; Ficara, E.; Aboulkas, A.; Barakat, A.; Carrère, H. New opportunities for agricultural digestate valorization: Current situation and perspectives. Energy Environ. Sci. 2015, 8, 2600–2621. [Google Scholar] [CrossRef]
- Godfrey, V. Production of Biodiesel from Oleaginous Organisms Using Underutilized Wastewaters. All Graduate Theses and Dissertations. Master’s Thesis, Utah State University, Logan, UT, USA, 2012. [Google Scholar] [CrossRef]
- Hakim, M.A.A.; Al-Ghouti, M.A.; Das, P.; Abu-Dieyeh, M.; Ahmed, T.A.; Aljabri, H.M.S. Potential application of microalgae in produced water treatment. Desalination Water Treat. 2018, 135, 47–58. [Google Scholar] [CrossRef]
- Lutzu, G.A.; Turgut, N.D. Growing Algae in Produced Water Generated During Oil and Gas Production Using Hydraulic Frac-turing Technique. Chem. Eng. Trans. 2019, 74, 1261–1266. [Google Scholar] [CrossRef]
- Parsy, A.; Sambusiti, C.; Baldoni-Andrey, P.; Elan, T.; Périé, F. Cultivation of Nannochloropsis oculata in saline oil & gas wastewater supplemented with anaerobic digestion effluent as nutrient source. Algal Res. 2020, 50, 101966. [Google Scholar] [CrossRef]
- Concas, A.; Lutzu, G.A.; Dunford, N.T. Experiments and modeling of Komvophoron sp. Growth in hydraulic fracturing wastewater. Chem. Eng. J. 2021, 426, 131299. [Google Scholar] [CrossRef]
- Ammar, S.H.; Khadim, H.J.; Mohamed, A.I. Cultivation of Nannochloropsis oculata and Isochrysis galbana microalgae in produced water for bioremediation and biomass production. Environ. Technol. Innov. 2018, 10, 132–142. [Google Scholar] [CrossRef]
- Das, P.; Abdulquadir, M.; Thaher, M.; Khan, S.; Chaudhary, A.K.; Alghasal, G.; Al-Jabri, H.M.S.J. Microalgal bioremediation of petroleum-derived low salinity and low pH produced water. J. Appl. Phycol. 2018, 31, 435–444. [Google Scholar] [CrossRef]
- Marques, I.M.; Oliveira, A.C.V.; de Oliveira, O.M.C.; Sales, E.A.; Moreira, T.A. A photobioreactor using Nannochloropsis oculata marine microalgae for removal of polycyclic aromatic hydrocarbons and sorption of metals in produced water. Chemosphere 2021, 281, 130775. [Google Scholar] [CrossRef] [PubMed]
- Cinq-Mars, M.; Bourdeau, N.; Marchand, P.; Desgagné-Penix, I.; Barnabé, S. Characterization of two microalgae consortia grown in industrial wastewater for biomass valorization. Algal Res. 2022, 62, 102628. [Google Scholar] [CrossRef]
- Babatsouli, P.; Fodelianakis, S.; Paranychianakis, N.; Venieri, D.; Dialynas, M.; Kalogerakis, N. Single stage treatment of saline wastewater with marine bacterial–microalgae consortia in a fixed-bed photobioreactor. J. Hazard. Mater. 2015, 292, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Parsy, A.; Guyoneaud, R.; Lot, M.-C.; Baldoni-Andrey, P.; Périé, F.; Sambusiti, C. Impact of salinities, metals and organic compounds found in saline oil & gas produced water on microalgae and cyanobacteria. Ecotoxicol. Environ. Saf. 2022, 234, 113351. [Google Scholar] [CrossRef]
- Díez, B.; Pedrós-Alió, C.; Marsh, T.; Massana, R. Application of Denaturing Gradient Gel Electrophoresis (DGGE) To Study the Diversity of Marine Picoeukaryotic Assemblages and Comparison of DGGE with Other Molecular Techniques. Appl. Environ. Microbiol. 2001, 67, 2942–2951. [Google Scholar] [CrossRef]
- Zimmermann, J.; Jahn, R.; Gemeinholzer, B. Barcoding diatoms: Evaluation of the V4 subregion on the 18S rRNA gene, including new primers and protocols. Org. Divers. Evol. 2011, 11, 173–192. [Google Scholar] [CrossRef]
- Guillard, R.R.; Ryther, J.H. Studies of marine planktonic diatoms: I. Cyclotella nana hustedt, and detonula Confervacea (cleve) Gran. Can. J. Microbiol. 1962, 8, 229–239. [Google Scholar] [CrossRef]
- García, M.T.; Mellado, E.; Ostos, J.C.; Ventosa, A. Halomonas organivorans sp. nov., a moderate halophile able to degrade aromatic compounds. Int. J. Syst. Evol. Microbiol. 2004, 54, 1723–1728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hedlund, B.P.; Staley, J.T. Vibrio cyclotrophicus sp. nov., a polycyclic aromatic hydrocarbon (PAH)-degrading marine bacterium. Int. J. Syst. Evol. Microbiol. 2001, 51, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Berlendis, S.; Cayol, J.-L.; Verhé, F.; Laveau, S.; Tholozan, J.-L.; Ollivier, B.; Auria, R. First Evidence of Aerobic Biodegradation of BTEX Compounds by Pure Cultures of Marinobacter. Appl. Biochem. Biotechnol. 2009, 160, 1992–1999. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Qian, P.Y. Conservative Fragments in Bacterial 16S rRNA Genes and Primer Design for 16S Ribosomal DNA Amplicons in Metagenomic Studies. PLOS ONE 2009, 4, e7401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escudié, F.; Auer, L.; Bernard, M.; Mariadassou, M.; Cauquil, L.; Vidal, K.; Maman, S.; Hernandez-Raquet, G.; Combes, S.; Pascal, G. FROGS: Find, Rapidly, OTUs with Galaxy Solution. Bioinformatics 2018, 34, 1287–1294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afgan, E.; Baker, D.; Batut, B.; van den Beek, M.; Bouvier, D.; Čech, M.; Chilton, J.; Clements, D.; Coraor, N.; Grüning, B.A.; et al. The Galaxy platform for accessible, reproducible and collaborative biomedical analyses: 2018 update. Nucleic Acids Res. 2018, 46, W537–W544. [Google Scholar] [CrossRef] [Green Version]
- Pruesse, E.; Quast, C.; Knittel, K.; Fuchs, B.M.; Ludwig, W.; Peplies, J.; Glöckner, F.O. SILVA: A comprehensive online resource for quality checked and aligned ribosomal RNA sequence data compatible with ARB. Nucleic Acids Res. 2007, 35, 7188–7196. [Google Scholar] [CrossRef] [Green Version]
- Measuring Aromatic Hydrocarbons (BTX) n.d. Available online: https://aai.solutions/application-notes/measuring-aromatic-hydrocarbons-BTX (accessed on 10 October 2022).
- Ouyang, B.; Fang, H.; Dong, W.; Hou, H. Different mechanisms both lead to the production of the naphthalene–OH adduct in the 355nm and 266nm laser flash photolysis of the mixed aqueous solution of naphthalene and nitrous acid. J. Photochem. Photobiol. A: Chem. 2006, 181, 348–356. [Google Scholar] [CrossRef]
- Lima, A.L.C.; Farrington, J.; Reddy, C.M. Combustion-Derived Polycyclic Aromatic Hydrocarbons in the Environment—A Review. Environ. Forensics 2005, 6, 109–131. [Google Scholar] [CrossRef]
- De Souza, K.V.; Peralta-Zamora, P. Spectrophotometric determination of phenol in the presence of congeners by multivariated calibration. An. Acad. Bras. Cienc. 2001, 73, 519–524. [Google Scholar] [CrossRef] [Green Version]
- Jiang, F.; Wang, S.; Zhang, Y.; Ma, S.; Huang, Y.; Fan, H.; Li, Q.; Wang, H.; Wang, A.; Liu, H.; et al. Variation of Metagenome From Feedstock to Digestate in Full-Scale Biogas Plants. Front. Microbiol. 2021, 12. [Google Scholar] [CrossRef]
- Ma, S.; Jiang, F.; Huang, Y.; Zhang, Y.; Wang, S.; Fan, H.; Liu, B.; Li, Q.; Yin, L.; Wang, H.; et al. A microbial gene catalog of anaerobic digestion from full-scale biogas plants. Gigascience 2021, 10, giaa164. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Sun, J.-Q.; Wang, L.-J.; Liu, X.-Z.; Ji, Y.-Y.; Shao, Z.-Q.; Wu, X.-L. Aliidiomarina soli sp. nov., isolated from saline–alkaline soil. Int. J. Syst. Evol. Microbiol. 2017, 67, 724–728. [Google Scholar] [CrossRef] [PubMed]
- Grabowski, A.; Nercessian, O.; Fayolle, F.; Blanchet, D.; Jeanthon, C. Microbial diversity in production waters of a low-temperature biodegraded oil reservoir. FEMS Microbiol. Ecol. 2005, 54, 427–443. [Google Scholar] [CrossRef] [PubMed]
- Lipus, D.; Roy, D.; Khan, E.; Ross, D.; Vikram, A.; Gulliver, D.; Hammack, R.; Bibby, K. Microbial communities in Bakken region produced water. FEMS Microbiol. Lett. 2018, 365, fny107. [Google Scholar] [CrossRef]
- Schwitalla, P.; Mennerich, A.; Austermann-Haun, U.; Müller, A.; Dorninger, C.; Daims, H.; Holm, N.C.; Rönner-Holm, S.G.E. NH4+ ad-/desorption in sequencing batch reactors: Simulation, laboratory and full-scale studies. Water Sci. Technol. J. Int. Assoc. Water Pollut. Res. 2008, 58, 345–350. [Google Scholar] [CrossRef]
- Bassin, J.; Pronk, M.; Kraan, R.; Kleerebezem, R.; van Loosdrecht, M. Ammonium adsorption in aerobic granular sludge, activated sludge and anammox granules. Water Res. 2011, 45, 5257–5265. [Google Scholar] [CrossRef]
- Nadersha, S.; Hassan, A.A. Biodesalination and treatment of raw hypersaline produced water samples using indigenous wastewater algal consortia. Desalination 2022, 528, 115638. [Google Scholar] [CrossRef]
- Mahmoodlu, M.G.; Pontedeiro, E.M.; Guerrero, J.S.P.; Raoof, A.; Hassanizadeh, S.M.; van Genuchten, M.T. Dissolution kinetics of volatile organic compound vapors in water: An integrated experimental and computational study. J. Contam. Hydrol. 2017, 196, 43–51. [Google Scholar] [CrossRef]
- Takáčová, A.; Smolinská, M.; Semerád, M.; Matúš, P. Degradation of btex by microalgae Parachlorella kessleri. Pet. Coal 2015, 57, 101–107. [Google Scholar]
- Ghosal, D.; Ghosh, S.; Dutta, T.K.; Ahn, Y. Current State of Knowledge in Microbial Degradation of Polycyclic Aromatic Hydrocarbons (PAHs): A Review. Front. Microbiol. 2016, 7, 1369. [Google Scholar] [CrossRef] [PubMed]
- Directive 91/271/EEC on Urban Waste Water Treatment. J Off N° 135 30051991 P 0040-0052 n.d. Available online: https://eur-lex.europa.eu/legal-content/FR/TXT/HTML/?uri=CELEX%3A31991L0271 (accessed on 13 January 2023).







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parsy, A.; Sambusiti, C.; Gassie, C.; Baldoni-Andrey, P.; Périé, F.; Guyoneaud, R. Impact of Bioaugmentation on the Bioremediation of Saline-Produced Waters Supplemented with Anaerobic Digestate. Sustainability 2023, 15, 2166. https://doi.org/10.3390/su15032166
Parsy A, Sambusiti C, Gassie C, Baldoni-Andrey P, Périé F, Guyoneaud R. Impact of Bioaugmentation on the Bioremediation of Saline-Produced Waters Supplemented with Anaerobic Digestate. Sustainability. 2023; 15(3):2166. https://doi.org/10.3390/su15032166
Chicago/Turabian StyleParsy, Aurélien, Cecilia Sambusiti, Claire Gassie, Patrick Baldoni-Andrey, Frédéric Périé, and Rémy Guyoneaud. 2023. "Impact of Bioaugmentation on the Bioremediation of Saline-Produced Waters Supplemented with Anaerobic Digestate" Sustainability 15, no. 3: 2166. https://doi.org/10.3390/su15032166
APA StyleParsy, A., Sambusiti, C., Gassie, C., Baldoni-Andrey, P., Périé, F., & Guyoneaud, R. (2023). Impact of Bioaugmentation on the Bioremediation of Saline-Produced Waters Supplemented with Anaerobic Digestate. Sustainability, 15(3), 2166. https://doi.org/10.3390/su15032166