
Salinity-Induced Extinction of Zostera marina in Lake Grevelingen? How Strong Habitat Modification May Require Introduction of a Suitable Ecotype
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
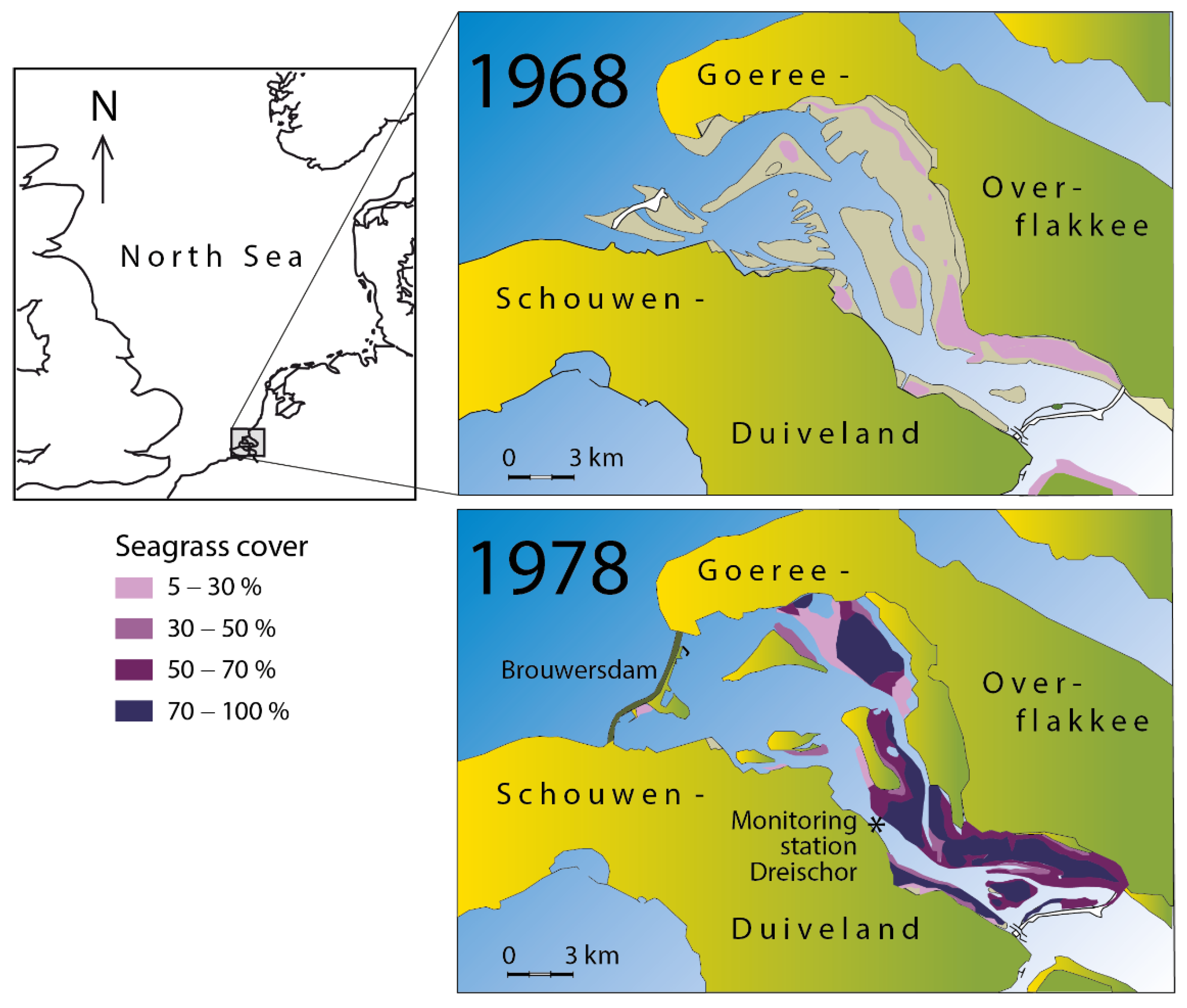
2.1. Study Area
2.2. Correlative Study of Seagrass Cover and Salinity
2.3. Germination Experiments
2.4. Statistical Analyzes
3. Results
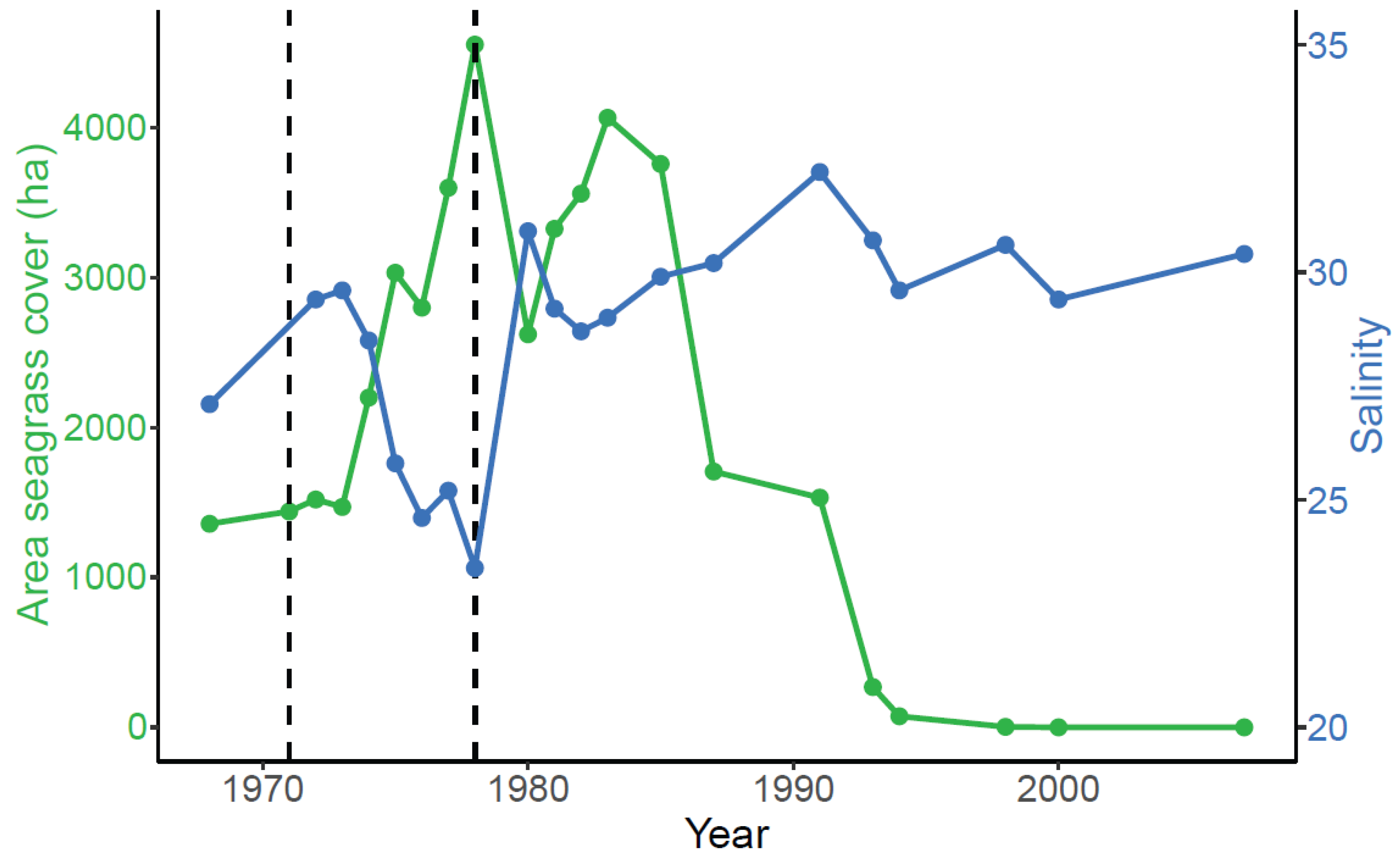
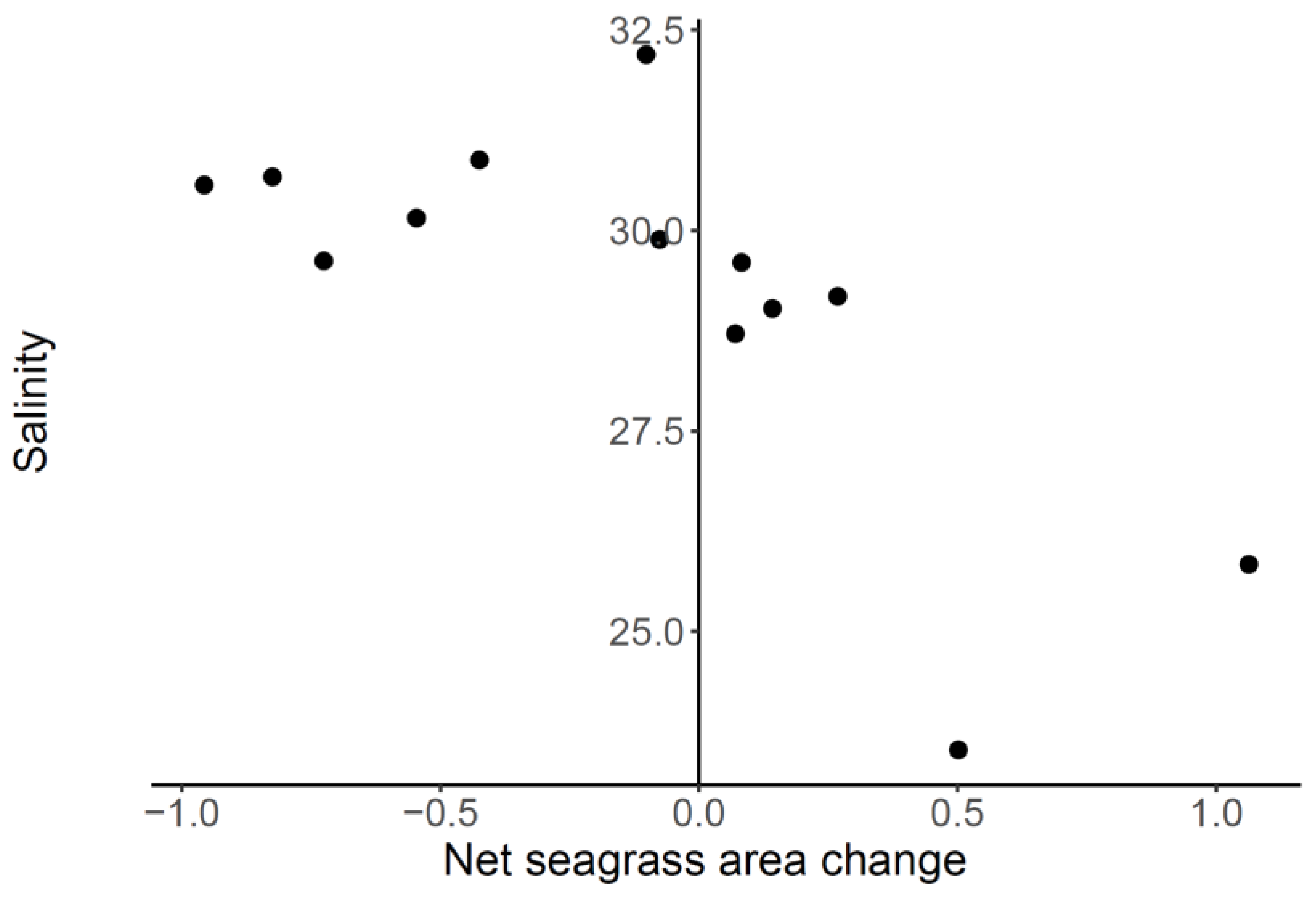
3.1. Correlation between Seagrass and Salinity
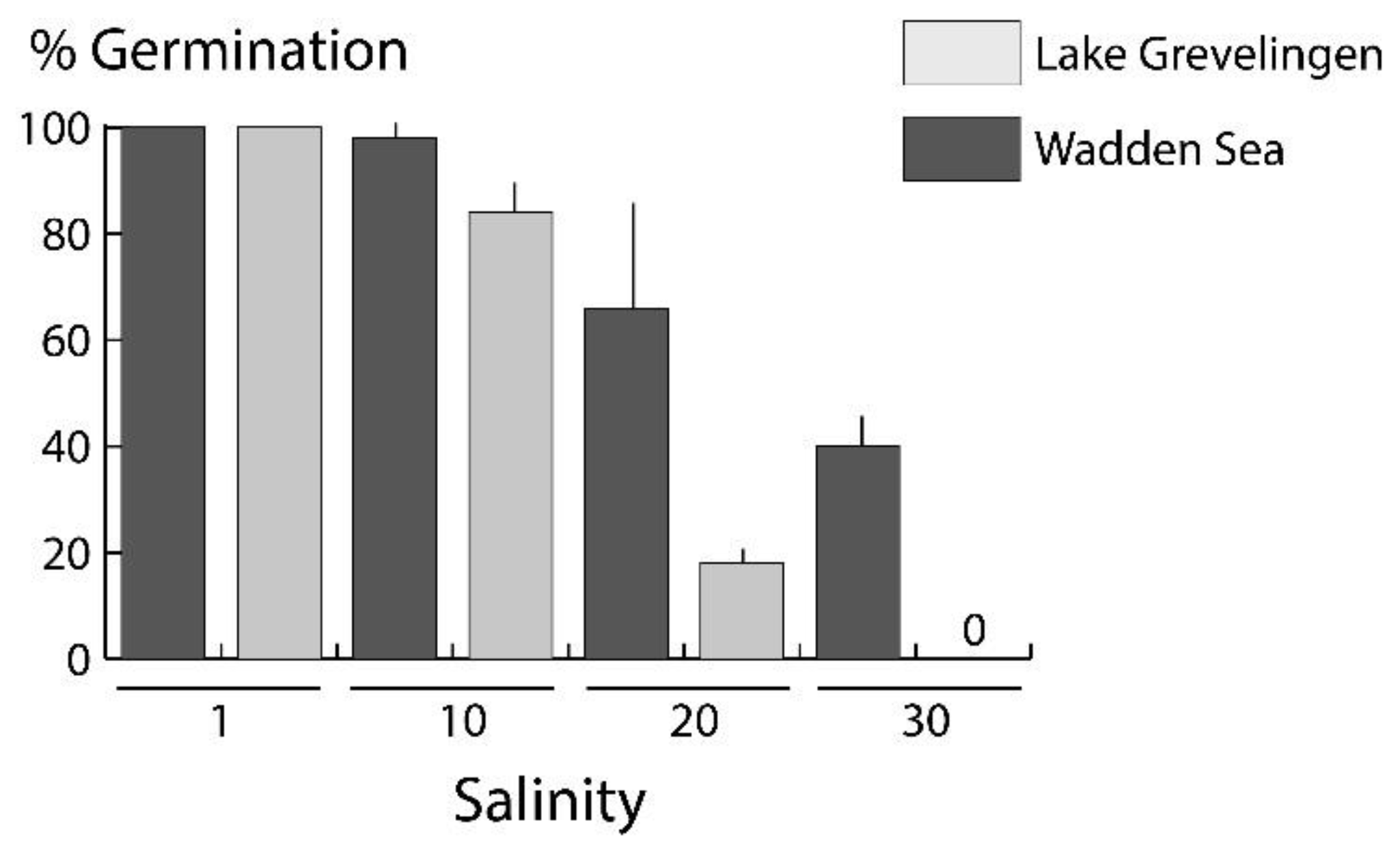
3.2. Germination Experiments
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nordlund, L.M.; Koch, E.W.; Barbier, E.B.; Creed, J.C. Seagrass ecosystem services and their variability across genera and geographical regions. PLoS ONE 2016, 11, e0163091. [Google Scholar] [CrossRef] [PubMed]
- Waycott, M.; Duarte, C.M.; Carruthers, T.J.B.; Orth, R.J.; Dennison, W.C.; Olyarnik, S.; Calladine, A.; Fourqurean, J.W.; Heck, K.L.; Hughes, A.R.; et al. Accelerating loss of seagrasses across the globe threatens coastal ecosystems. Proc. Natl. Acad. Sci. USA 2009, 106, 12377–12381. [Google Scholar] [CrossRef] [PubMed]
- van Katwijk, M.M.; Thorhaug, A.; Marba, N.; Orth, R.J.; Duarte, C.M.; Kendrick, G.A.; Althuizen, I.H.J.; Balestri, E.; Bernard, G.; Cambridge, M.L.; et al. Global analysis of seagrass restoration: The importance of large-scale planting. J. Appl. Ecol. 2016, 53, 567–578. [Google Scholar] [CrossRef]
- Rezek, R.J.; Furman, B.T.; Jung, R.P.; Hall, M.O.; Bell, S.S. Long-term performance of seagrass restoration projects in Florida, USA. Sci. Rep. 2019, 9, 15514. [Google Scholar] [CrossRef] [PubMed]
- Orth, R.J.; Lefcheck, J.S.; McGlathery, K.S.; Aoki, L.; Luckenbach, M.W.; Moore, K.A.; Oreska MP, J.; Snyder, R.; Wilcox, D.J.; Lusk, B. Restoration of seagrass habitat leads to rapid recovery of coastal ecosystem services. Sci. Adv. 2020, 6, eabc6434. [Google Scholar] [CrossRef] [PubMed]
- de los Santos, C.B.; Krause-Jensen, D.; Alcoverro, T.; Marba, N.; Duarte, C.M.; van Katwijk, M.M.; Perez, M.; Romero, J.; Sanchez-Lizaso, J.L.; Roca, G.; et al. Recent trend reversal for declining European seagrass meadows. Nat. Commun. 2019, 10, 3356. [Google Scholar] [CrossRef]
- Nienhuis, P.H.; de Bree, B.H.H.; Herman, P.M.J.; Holland, A.M.B.; Verschuure, J.M.; Wessel, E.G.J. Twenty-five years of changes in the distribution and biomass of eelgrass, Zostera marina, in Grevelingen Lagoon, The Netherlands. Neth. J. Aquat. Ecol. 1996, 30, 107–117. [Google Scholar] [CrossRef]
- van der Heide, T.; Peeters, E.T.H.M.; Hermus, D.C.R.; van Katwijk, M.M.; Roelofs, J.G.M.; Smolders, A.J.P. Predicting habitat suitability in temperate seagrass ecosystems. Limnol. Oceanogr. 2009, 54, 2018–2024. [Google Scholar] [CrossRef]
- Wolff, W.J.; Koeijer, P.; Sandee, A.J.J.; Wolf, L. De verspreiding van Rotganzen in het Deltagebied in relatie tot de verspreiding van hun voedsel. Limosa 1967, 40, 163–174. [Google Scholar]
- Bannink, B.A.; Vandermeulen, J.H.M.; Nienhuis, P.H. Lake Grevelingen—from an estuary to a saline lake—An introduction. Neth. J. Sea Res. 1984, 18, 179–190. [Google Scholar] [CrossRef]
- Beeftink, W.G. De zoutvegetatie van ZW-Nederland beschouwd in Europees verband. Meded. Landbouwhogesch. Wagening 1965, 65–61, 82–85. [Google Scholar]
- Nienhuis, P.H.; de Bree, B.H.H. Production and growth dynamics of eelgrass (Zostera marina) in Brackish Lake Grevelingen (The Netherlands). Neth. J. Sea Res. 1980, 14, 102–118. [Google Scholar] [CrossRef]
- Wielemaker-Van den Dool, A.; Herman, P.M.J. Historical Coverage Seagrasses in Veerse Meer and Grevelingen Meer. 2003. Available online: https://www.rijkswaterstaat.nl/apps/geoservices/geodata/dmc/zeegras/ (accessed on 14 January 2020).
- Verschuure, J.M. Verspreiding van Groot zeegras (Zostera marina L.) in het Grevelingenmeer en het Veerse Meer in 2000 (In Dutch); Nederlands Instituut voor Oecologisch Onderzoek, Centrum voor Estuariene en Mariene Oecologie: Yerseke, The Netherlands, 2000. [Google Scholar]
- Engelberts, A.; van Avesaath, P.H.; Hummel, H. Inventarisatie Groot zeegras in de Grevelingen. Situatie 2007 (In Dutch); Nederland Instituut voor Ecologie, Centrum voor Estuariene en Mariene Ecologie, Monitor Taskforce: Yerseke, The Netherlands, 2007. [Google Scholar]
- Nienhuis, P.H. Temporal and spatial patterns of eelgrass (Zostera marina L.) in a former estuary in The Netherlands, dominated by human activities. Mar. Technol. Soc. J. 1983, 17, 69–77. [Google Scholar]
- Herman, P.M.J.; Hemminga, M.A.; Nienhuis, P.H.; Verschuure, J.M.; Wessel, E.G.J. Wax and wane of eelgrass Zostera marina and water column silicon levels. Mar. Ecol. Prog. Ser. 1996, 144, 303–307. [Google Scholar] [CrossRef]
- van Lent, F.; Verschuure, J.M.; van Veghel, M.L.J. Comparative study on populations of Zostera marina L. (eelgrass): In situ nitrogen enrichment and light manipulation. J. Exp. Mar. Biol. Ecol. 1995, 185, 55–76. [Google Scholar] [CrossRef]
- Wetsteijn, L.P. Grevelingenmeer: Meer Kwetsbaar? Een Beschrijving van de Ecologische Ontwikkelingen voor de Periode 1999 t/m 2008–2010 in Vergelijking met de Periode 1990 t/m 1998. (In Dutch); RWS Waterdienst: Lelystad, The Netherlands, 2011. [Google Scholar]
- Hagens, M.; Slomp, C.P.; Meysman, F.J.R.; Seitaj, D.; Harlay, J.; Borges, A.V.; Middelburg, J.J. Biogeochemical processes and buffering capacity concurrently affect acidification in a seasonally hypoxic coastal marine basin. Biogeosciences 2015, 12, 1561–1583. [Google Scholar] [CrossRef]
- Seitaj, D.; Sulu-Gambari, F.; Burdorf, L.D.W.; Romero-Ramirez, A.; Maire, O.; Malkin, S.Y.; Slomp, C.P.; Meysman, F.J.R. Sedimentary oxygen dynamics in a seasonally hypoxic basin. Limnol. Oceanogr. 2017, 62, 452–473. [Google Scholar] [CrossRef]
- Cronau, R.J.T.; de Fouw, J.; van Katwijk, M.M.; Bouma, T.J.; Heusinkveld, J.H.T.; Hoeijmakers, D.; Lamers, L.P.M.; van der Heide, T. Seed-versus transplant-based eelgrass (Zostera marina L.) restoration success in a temperate marine lake. Restor. Ecol. 2023, 31, e13786. [Google Scholar] [CrossRef]
- Didderen, K.; Lengkeek, W.; Bakker, E.G.R.; Tummers, J.; Gmelig Meyling, A. Vis in de Grevelingen. Bureau Waardenburg Rapportnr. 20-328 (In Dutch); Bureau Waardenburg/RAVON/ANEMOON: Culemborg, The Netherlands, 2021. [Google Scholar]
- van Katwijk, M.M.; Schmitz, G.H.W.; Hanssen, L.S.A.M.; den Hartog, C. Suitability of Zostera marina populations for transplantation to the Wadden Sea as determined by a mesocosm shading experiment. Aquat. Bot. 1998, 60, 283–305. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing, Version 4.1.0; R foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Kelderman, P. Nutrient concentrations in the interstitial water of Lake Grevelingen sediment—Effects of sediment redistribution and benthic primary production processes. Neth. J. Sea Res. 1984, 18, 312–336. [Google Scholar] [CrossRef]
- van Lent, F.; Verschuure, J.M. Intraspecific variability of Zostera marina L. (eelgrass) in the estuaries and lagoons of the Southwester Netherlands. 2. Relation with environmental factors. Aquat. Bot. 1994, 48, 59–75. [Google Scholar] [CrossRef]
- van der Heide, T. Stressors and Feedbacks in Temperate Seagrass Ecosystems. Ph.D. Thesis, Radboud University, Nijmegen, The Netherlands, 2009. [Google Scholar]
- Holland, A.M.B. Waterbeheer Grevelingenmeer 1980–1990; Nota GWWS-91.086; Rijkswaterstaat: Utrecht, The Netherlands, 1991. [Google Scholar]
- Roca, G.; Alcoverro, T.; Krause-Jensen, D.; Balsby, T.J.S.; van Katwijk, M.M.; Marba, N.; Santos, R.; Arthur, R.; Mascaro, O.; Fernandez-Torquemada, Y.; et al. Response of seagrass indicators to shifts in environmental stressors: A global review and management synthesis. Ecol. Indic. 2016, 63, 310–323. [Google Scholar] [CrossRef]
- Touchette, B.W.; Burkholder, J.M. Carbon and nitrogen metabolism in the seagrass, Zostera marina L.: Environmental control of enzymes involved in carbon allocation and nitrogen assimilation. J. Exp. Mar. Biol. Ecol. 2007, 350, 216–233. [Google Scholar] [CrossRef]
- Vonk, J.A.; Stapel, J. Regeneration of nitrogen (15N) from seagrass litter in tropical Indo-Pacific meadows. Mar. Ecol. Prog. Ser. 2008, 368, 165–175. [Google Scholar] [CrossRef]
- Kamermans, P.; Hemminga, M.A.; de Jong, D.J. Significance of salinity and silicon levels for growth of a formerly estuarine eelgrass (Zostera marina) population (Lake Grevelingen, the Netherlands). Mar. Biol. 1999, 133, 527–539. [Google Scholar] [CrossRef]
- Blok, S.E.; Olesen, B.; Krause-Jensen, D. Life history events of eelgrass Zostera marina L. populations across gradients of latitude and temperature. Mar. Ecol. Prog. Ser. 2018, 590, 79–93. [Google Scholar] [CrossRef]
- Zang, Y.; Liu, J.; Tang, X.X.; Zhou, B. Description of a Zostera marina catalase gene involved in responses to temperature stress. Peerj 2018, 6, e4532. [Google Scholar] [CrossRef] [PubMed]
- Olesen, B.; Krause-Jensen, D.; Marba, N.; Christensen, P.B. Eelgrass Zostera marina in subarctic Greenland: Dense meadows with slow biomass turnover in cold waters. Mar. Ecol. Prog. Ser. 2015, 518, 107–121. [Google Scholar] [CrossRef]
- Kaldy, J.E.; Shafer, D.J.; Magoun, A.D. Duration of temperature exposure controls growth of Zostera japonica: Implications for zonation and colonization. J. Exp. Mar. Biol. Ecol. 2015, 464, 68–74. [Google Scholar] [CrossRef]
- de Jong, D.J.; van Katwijk, M.M.; Jager, Z. Zeegras in Nederland (in Dutch). De Levende Nat. 2004, 105, 209–211. [Google Scholar]
- van Katwijk, M.M.; Wijgergangs, L.J.M. Effects of locally varying exposure, sediment type and low-tide water cover on Zostera marina recruitment from seed. Aquat. Bot. 2004, 80, 1–12. [Google Scholar] [CrossRef]
- van Lent, F.; Verschuure, J.M. Comparative study on populations of Zostera marina L. (eelgrass): Experimental germination and growth. J. Exp. Mar. Biol. Ecol. 1995, 185, 77–91. [Google Scholar] [CrossRef]
- Duarte, C.M. Allometric scaling of seagrass form and productivity. Mar. Ecol. Prog. Ser. 1991, 77, 289–300. [Google Scholar] [CrossRef]
- Hootsmans, M.J.M.; Vermaat, J.E.; Van Vierssen, W. Seed-bank development, germination and early seedling survival of two seagrass species from the Netherlands: Zostera marina L. and Z. noltii Hornemann. Aquat. Bot. 1987, 28, 275–285. [Google Scholar] [CrossRef]
- Xu, S.C.; Zhou, Y.; Wang, P.M.; Wang, F.; Zhang, X.M.; Gu, R.T. Salinity and temperature significantly influence seed germination, seedling establishment, and seedling growth of eelgrass Zostera marina L. Peerj 2016, 4, e2697. [Google Scholar] [CrossRef] [PubMed]
- McMillan, C. Seed germination for an annual form of Zostera marina from the Sea of Cortez, Mexico. Aquat. Bot. 1983, 16, 105–110. [Google Scholar] [CrossRef]
- van Katwijk, M.M.; Schmitz, G.H.W.; Gasseling, A.M.; van Avesaath, P.H. The effects of salinity and nutrient load and their interaction on Zostera marina L. Mar. Ecol. Prog. Ser. 1999, 190, 155–165. [Google Scholar] [CrossRef]
- Peelen, R. Isohalines in the delta area of the rivers Rhine, Meuse and Scheldt. Neth. J. Sea Res. 1967, 3, 575–597. [Google Scholar] [CrossRef]
- Olsen, J.L.; Stam, W.T.; Coyer, J.A.; Reusch, T.B.H.; Billingham, M.; Bostrom, C.; Calvert, E.; Christie, H.; Granger, S.; La Lumiere, R.; et al. North Atlantic phylogeography and large-scale population differentiation of the seagrass Zostera marina L. Mol. Ecol. 2004, 13, 1923–1941. [Google Scholar] [CrossRef]
- Frankham, R. Genetics and extinction. Biol. Conserv. 2005, 126, 131–140. [Google Scholar] [CrossRef]
- Larkin, P.D.; Maloney, T.J.; Rubiano-Rincon, S.; Barrett, M.M. A map-based approach to assessing genetic diversity, structure, and connectivity in the seagrass Halodule wrightii. Mar. Ecol. Prog. 2017, 567, 95–107. [Google Scholar] [CrossRef]
- Alotaibi, N.M.; Kenyon, E.J.; Cook, K.J.; Borger, L.; Bull, J.C. Low genotypic diversity and long-term ecological decline in a spatially structured seagrass population. Sci. Rep. 2019, 9, 18387. [Google Scholar] [CrossRef] [PubMed]
- Pulido, C.; Borum, J. Eelgrass (Zostera marina) tolerance to anoxia. J. Exp. Mar. Biol. Ecol. 2010, 385, 8–13. [Google Scholar] [CrossRef]
- Franssen, S.U.; Gu, J.; Winters, G.; Huylmans, A.K.; Wienpahl, I.; Sparwel, M.; Coyer, J.A.; Olsen, J.L.; Reusch, T.B.H.; Bornberg-Bauer, E. Genome-wide transcriptomic responses of the seagrasses Zostera marina and Nanozostera noltii under a simulated heatwave confirm functional types. Mar. Genom. 2014, 15, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; English, M.K.; Tomas, F.; Mueller, R.S. Recovery and community succession of the Zostera marina rhizobiome after transplantation. Appl. Environ. Microbiol. 2021, 87, e02326-20. [Google Scholar] [CrossRef] [PubMed]
- Pazzaglia, J.; Nguyen, H.M.; Santillan-Sarmiento, A.; Ruocco, M.; Dattolo, E.; Marin-Guirao, L.; Procaccini, G. The Genetic Component of Seagrass Restoration: What We Know and the Way Forwards. Water 2021, 13, 829. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Origin | Salinity 1–15 No Si Added | Salinity 22 5 µM Si | Salinity 22 100 µM Si | Salinity 32 5 µM Si | Salinity 32 100 µM Si |
|---|---|---|---|---|---|
| Lake Grevelingen | 35–40% | 0% | 0% | 0% | 0% |
| Lake Veere | 35–40% | 7% | 0% | 0% | 0% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
van Katwijk, M.M.; Cronau, R.J.T.; Lamers, L.P.M.; Kamermans, P.; van Tussenbroek, B.I.; de Jong, D.J. Salinity-Induced Extinction of Zostera marina in Lake Grevelingen? How Strong Habitat Modification May Require Introduction of a Suitable Ecotype. Sustainability 2023, 15, 3472. https://doi.org/10.3390/su15043472
van Katwijk MM, Cronau RJT, Lamers LPM, Kamermans P, van Tussenbroek BI, de Jong DJ. Salinity-Induced Extinction of Zostera marina in Lake Grevelingen? How Strong Habitat Modification May Require Introduction of a Suitable Ecotype. Sustainability. 2023; 15(4):3472. https://doi.org/10.3390/su15043472
Chicago/Turabian Stylevan Katwijk, Marieke M., Rens J. T. Cronau, Leon P. M. Lamers, Pauline Kamermans, Brigitta I. van Tussenbroek, and Dick J. de Jong. 2023. "Salinity-Induced Extinction of Zostera marina in Lake Grevelingen? How Strong Habitat Modification May Require Introduction of a Suitable Ecotype" Sustainability 15, no. 4: 3472. https://doi.org/10.3390/su15043472
APA Stylevan Katwijk, M. M., Cronau, R. J. T., Lamers, L. P. M., Kamermans, P., van Tussenbroek, B. I., & de Jong, D. J. (2023). Salinity-Induced Extinction of Zostera marina in Lake Grevelingen? How Strong Habitat Modification May Require Introduction of a Suitable Ecotype. Sustainability, 15(4), 3472. https://doi.org/10.3390/su15043472