Alternative Testing Methods for Predicting Health Risk from Environmental Exposures

Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells
2.2. Air Samples

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Description | Sampling Period | PM2.5 (µg/m3) | PM1 (µg/m3) | Toxicological Assay |
|---|---|---|---|---|---|
| Supersite | urban background site located in the northern area of the City of Bologna | Autumn | 27.27 | 19.86 | Cytotoxicity assay (CTA) |
| MXW | site located in the surroundings of Bologna and impacted by the waste-to-energy plant | Summer | 23.33 | ND | Microarray experiments |
| Winter | 35.42 | ND | |||
| CTW | site located in the surroundings of Bologna and impacted by all of the same sources as MXW, except the waste-to-energy plant | Summer | 21.11 | ND | |
| Winter | 34.47 | ND | |||
| GMA | Urban background site located in the southern area of Bologna (public park) | Summer | 18.74 | ND | |
| Winter | 29.56 | ND |
2.3. Cell Transformation Assay
2.3.1. Morphological Transformation Assay
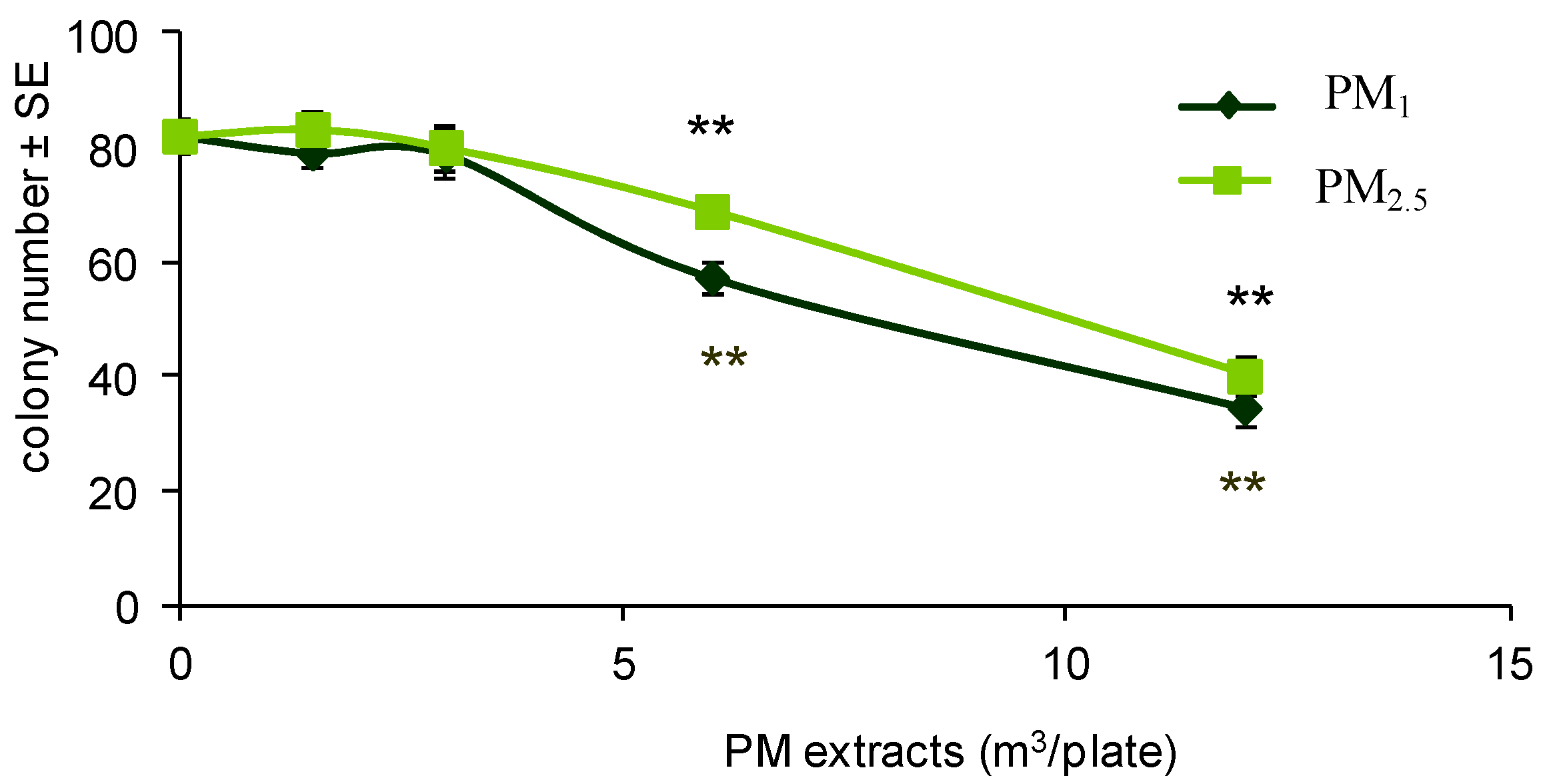
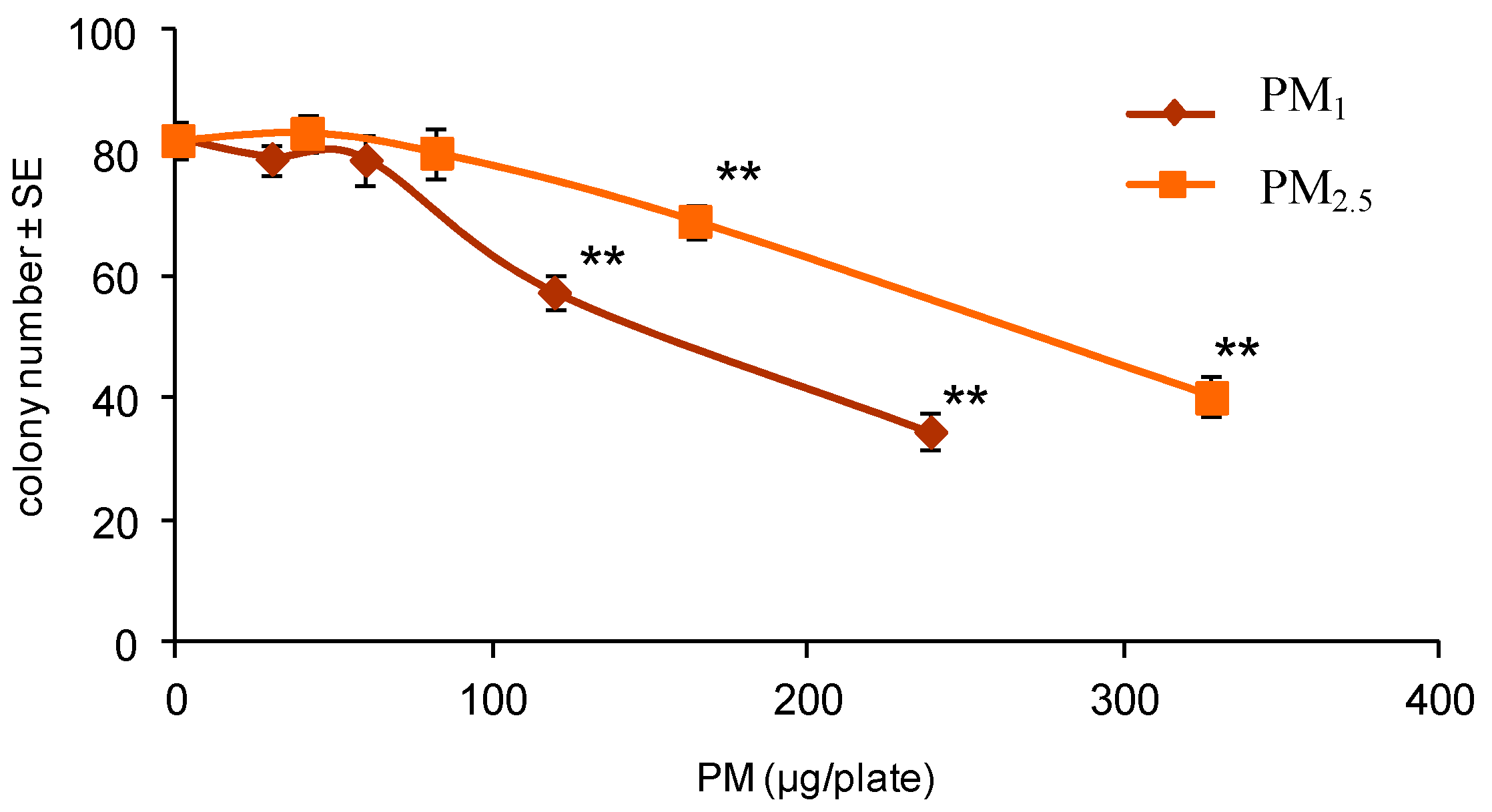
2.3.2. Colony Forming Efficiency Assay
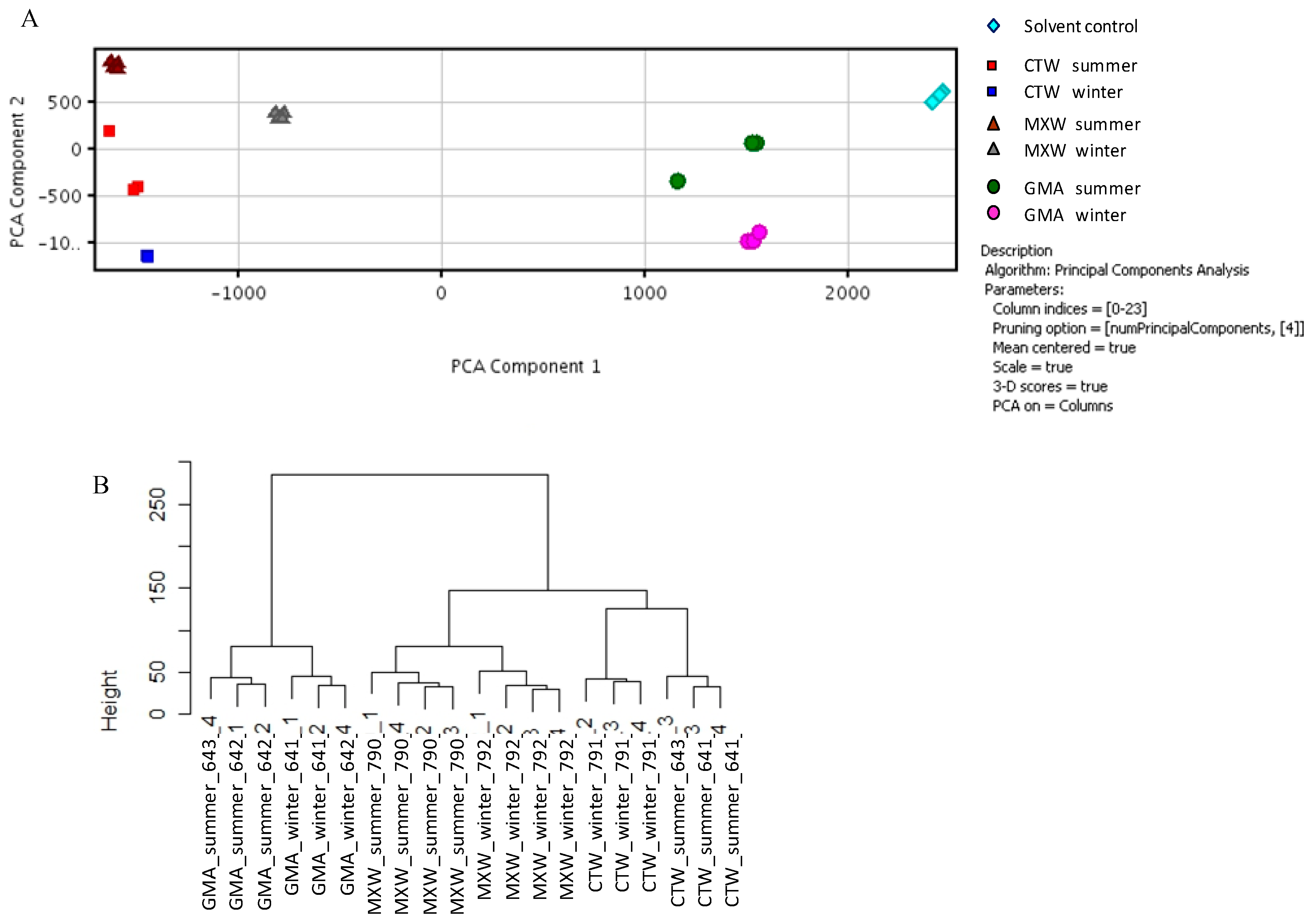
2.4. Microarray Experiments
3. Results and Discussion
3.1. Cell Transformation Assay
| Treatment | Plates with Foci/Plates Scored | Mean No. of Transformed Foci/Plate ± SE | RCE (%) | TF (×10−4) |
|---|---|---|---|---|
| Untreated cells | 0/10 | 0 | 96 | 0 |
| DMSO 0.5% | 0/10 | 0 | 100 | 0 |
| 3-MCA 4.0 µg/mL | 10/10 a | 7.50 ± 0.78 b | 59 c | 12.91 d |
| PM1 (m3) | 0 | |||
| 1.5 | 2/10 | 0.20 ± 0.13 | 96 | 0.21 |
| 3 | 0/10 | 0 | 96 | 0 |
| 6 | 0/10 | 0 | 70 c | 0 |
| 12 | 0/9 | 0 | 42 c | 0 |
| PM2.5 (m3) | ||||
| 1.5 | 1/10 | 0.20 ± 0.20 | 101 | 0.20 |
| 3 | 0/10 | 0 | 98 | 0 |
| 6 | 0/10 | 0 | 84 c | 0 |
| 12 | 1/10 | 0.10 ± 0.10 | 49 c | 0.21 |


3.2. Gene Expression
3.2.1. Experimental Design and Data Analysis

| Title | A549 (t-test, FDR < 0.01, Fold-Change > 1.5) | ||
|---|---|---|---|
| Site | |||
| All | Up | Down | |
| MXW vs. GMA | 1530 | 520 | 1010 |
| CTW vs. GMA | 1305 | 391 | 914 |
| MXW vs. CTW | 279 | 142 | 137 |
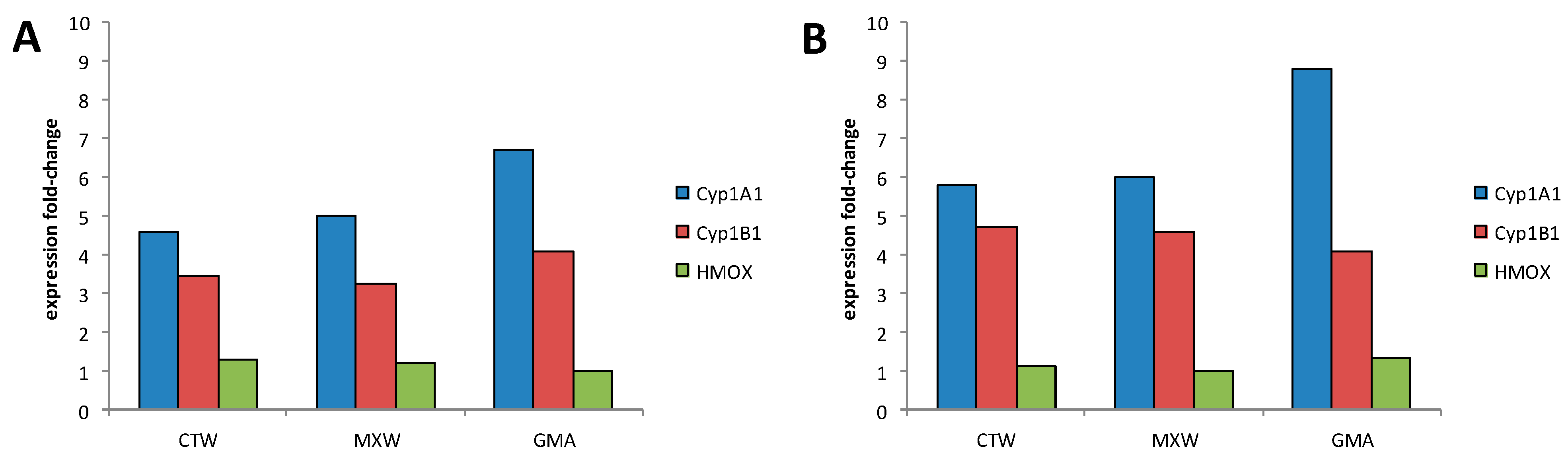
3.2.2. Biomarkers of Exposure

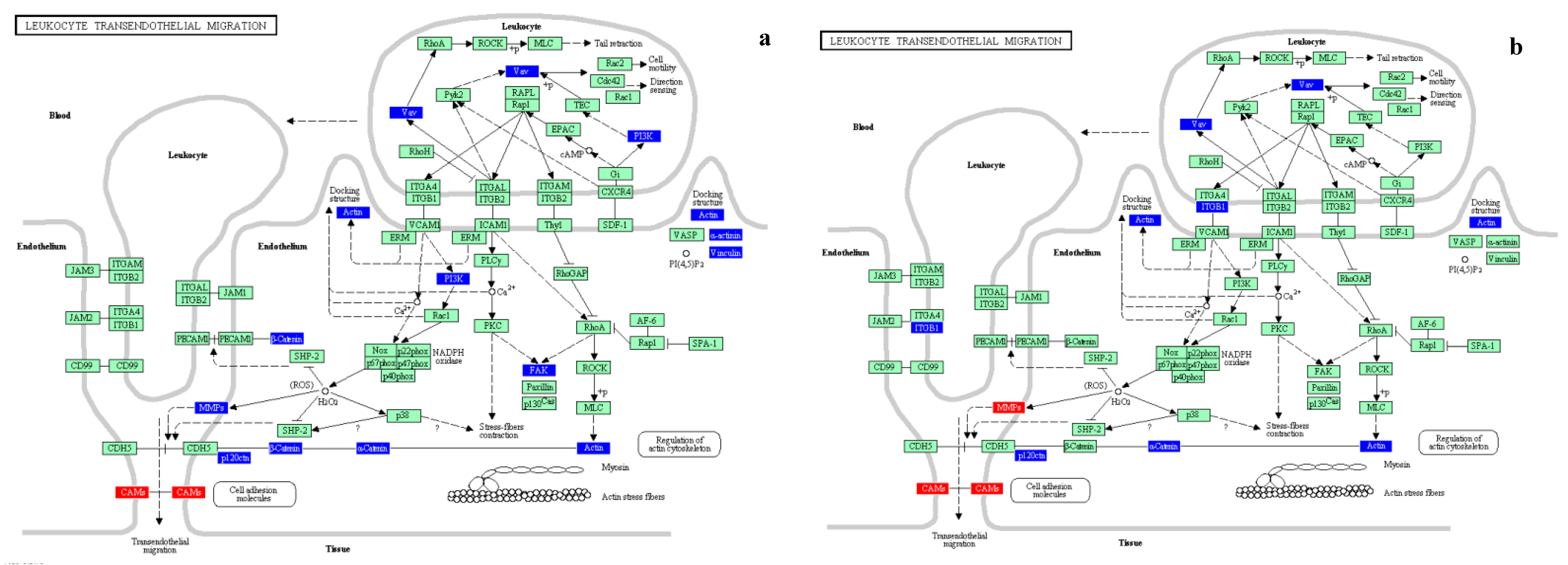
3.2.3. Biological Interpretation
| A549 Pathway-Express Analysis | ||
|---|---|---|
| MXW vs. GMA | CTW vs. GMA | MXW vs. CTW |
| Cell adhesion molecules (CAMs) | Cell adhesion molecules (CAMs) | Antigen processing and presentation |
| Leukocyte transendothelial migration | Leukocyte transendothelial migration | Phosphatidylinositol signaling system |
| Adherens junction | Adherens junction | |
| Phosphatidylinositol signaling system | Phosphatidylinositol signaling system | |
| Antigen processing and presentation | Circadian rhythm | |
| Tight junction | Pathways in cancer | |
| Focal adhesion | Notch signaling pathway | |
| Circadian rhythm | Non-small cell lung cancer | |
| Pathways in cancer | Endometrial cancer | |
| Insulin signaling pathway | Ubiquitin mediated proteolysis | |
| ErbB signaling pathway | Wnt signaling pathway | |
| Regulation of actin cytoskeleton | Bladder cancer | |
| Endometrial cancer | Thyroid cancer | |
| Shigellosis | Focal adhesion | |
| Pathogenic Escherichia coli infection | Tight junction | |
| T cell receptor signaling pathway | Vibrio cholerae infection | |
| Wnt signaling pathway | ||
| Alzheimer’s disease | ||
| GnRH signaling pathway | ||
| Type II diabetes mellitus | ||
| Notch signaling pathway | ||
| Colorectal cancer | ||
| MAPK signaling pathway | ||
| Renal cell carcinoma | ||
| Vibrio cholera infection | ||

| A549 | ||||
|---|---|---|---|---|
| GSEA | KEGG Pathways (FDR < 0.25) | |||
| Versus | MXW | CTW | GMA | |
| MXW | / | 0 | 0 | |
| CTW | 16 | / | 1 | |
| GMA | 98 | 86 | / | |
| A549 GSEA Analysis KEGG Pathways | ||
|---|---|---|
| Enriched in MXW vs. GMA | Significantly Enriched in CTW vs. GMA | Significantly Enriched in CTW vs. MXW |
| Oxidative Phosphorylation | Systemic Lupus Erythematosus (significant) | VEGF Signaling Pathway |
| Parkinson’s Disease | Oxidative Phosphorylation | Glycine Serine and Threonine Metabolism |
| Protein Export | Parkinson’s Disease | Maturity Onset Diabetes of The Young |
| Systemic Lupus Erythematosus | Intestinal Immune Network For Iga Production | Colorectal Cancer |
| Biosynthesis of Unsaturated Fatty Acids | Neuroactive Ligand Receptor Interaction | Prostate Cancer |
| Pentose Phosphate Pathway | ||
| Leukocyte Transendothelial Migration | ||
| B-Cell Receptor Signaling Pathway | ||
| Regulation of Actin Cytoskeleton | ||
| Hedgehog Signaling Pathway | ||
| Primary Immunodeficiency | ||
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Jarvis, I.W.; Dreij, K.; Mattsson, A.; Jernstrom, B.; Stenius, U. Interactions between polycyclic aromatic hydrocarbons in complex mixtures and implications for cancer risk assessment. Toxicology 2014, 321, 27–39. [Google Scholar] [CrossRef]
- Pope, C.A., 3rd; Burnett, R.T.; Krewski, D.; Jerrett, M.; Shi, Y.; Calle, E.E.; Thun, M.J. Cardiovascular mortality and exposure to airborne fine particulate matter and cigarette smoke: Shape of the exposure-response relationship. Circulation 2009, 120, 941–948. [Google Scholar]
- Loomis, D.; Grosse, Y.; Lauby-Secretan, B.; Ghissassi, F.E.; Bouvard, V.; Benbrahim-Tallaa, L.; Guha, N.; Baan, R.; Mattock, H.; Straif, K. The carcinogenicity of outdoor air pollution. Lancet Oncol. 2013, 14, 1262–1263. [Google Scholar] [CrossRef]
- Vasseur, P.; Lasne, C. Oecd detailed review paper (drp) number 31 on “Cell transformation assays for detection of chemical carcinogens”: Main results and conclusions. Mutat. Res. 2012, 744, 8–11. [Google Scholar] [CrossRef]
- Lilienblum, W.; Dekant, W.; Foth, H.; Gebel, T.; Hengstler, J.G.; Kahl, R.; Kramer, P.J.; Schweinfurth, H.; Wollin, K.M. Alternative methods to safety studies in experimental animals: Role in the risk assessment of chemicals under the new european chemicals legislation (reach). Arch. Toxicol. 2008, 82, 211–236. [Google Scholar]
- Vanparys, P.; Corvi, R.; Aardema, M.; Gribaldo, L.; Hayashi, M.; Hoffmann, S.; Schechtman, L. Ecvam prevalidation of three cell transformation assays. Altex 2011, 28, 56–59. [Google Scholar] [CrossRef]
- Corvi, R.; Aardema, M.J.; Gribaldo, L.; Hayashi, M.; Hoffmann, S.; Schechtman, L.; Vanparys, P. Ecvam prevalidation study on in vitro cell transformation assays: General outline and conclusions of the study. Mutat. Res. 2012, 744, 12–19. [Google Scholar]
- Kakunaga, T. Critical review of the use of established cell lines for in vitro cell transformation. IARC Sci. Publ. 1985, 67, 55–73. [Google Scholar]
- Sen, B.; Mahadevan, B.; DeMarini, D.M. Transcriptional responses to complex mixtures: A review. Mutat. Res. 2007, 636, 144–177. [Google Scholar]
- Gualtieri, M.; Mantecca, P.; Corvaja, V.; Longhin, E.; Perrone, M.G.; Bolzacchini, E.; Camatini, M. Winter fine particulate matter from milan induces morphological and functional alterations in human pulmonary epithelial cells (a549). Toxicol. Lett. 2009, 188, 52–62. [Google Scholar] [CrossRef]
- Roig, N.; Sierra, J.; Rovira, J.; Schuhmacher, M.; Domingo, J.L.; Nadal, M. In vitro tests to assess toxic effects of airborne pm(10) samples. Correlation with metals and chlorinated dioxins and furans. Sci. Total Environ. 2013, 443, 791–797. [Google Scholar] [CrossRef]
- Foster, K.A.; Oster, C.G.; Mayer, M.M.; Avery, M.L.; Audus, K.L. Characterization of the a549 cell line as a type II pulmonary epithelial cell model for drug metabolism. Exp. Cell Res. 1998, 243, 359–366. [Google Scholar] [CrossRef]
- Mascolo, M.G.; Perdichizzi, S.; Rotondo, F.; Morandi, E.; Guerrini, A.; Silingardi, P.; Vaccari, M.; Grilli, S.; Colacci, A. Balb/c 3t3 cell transformation assay for the prediction of carcinogenic potential of chemicals and environmental mixtures. Toxicol. In Vitro 2010, 24, 1292–1300. [Google Scholar] [CrossRef]
- Colacci, A.; Mascolo, M.G.; Perdichizzi, S.; Quercioli, D.; Gazzilli, A.; Rotondo, F.; Morandi, E.; Guerrini, A.; Silingardi, P.; Grilli, S.; et al. Different sensitivity of balb/c 3t3 cell clones in the response to carcinogens. Toxicol. In Vitro 2011, 25, 1183–1190. [Google Scholar] [CrossRef]
- Sasaki, K.; Bohnenberger, S.; Hayashi, K.; Kunkelmann, T.; Muramatsu, D.; Poth, A.; Sakai, A.; Salovaara, S.; Tanaka, N.; Thomas, B.C.; et al. Photo catalogue for the classification of foci in the balb/c 3t3 cell transformation assay. Mutat. Res. 2012, 744, 42–53. [Google Scholar] [CrossRef]
- Group, I.N.E.W. Cellular and molecular mechanisms of cell transformation and standardization of transformation assays of established cell lines for the prediction of carcinogenic chemicals: Overview and recommended protocols. Cancer Res. 1985, 45, 2395–2399. [Google Scholar]
- Franken, N.A.; Rodermond, H.M.; Stap, J.; Haveman, J.; van Bree, C. Clonogenic assay of cells In Vitro. Nat. Protoc. 2006, 1, 2315–2319. [Google Scholar] [CrossRef]
- Agilent Technologies. Available online: www.chem.agilent.com (accessed on 11 August 2014).
- Billet, S.; Abbas, I.; le Goff, J.; Verdin, A.; Andre, V.; Lafargue, P.E.; Hachimi, A.; Cazier, F.; Sichel, F.; Shirali, P.; et al. Genotoxic potential of polycyclic aromatic hydrocarbons-coated onto airborne particulate matter (pm 2.5) in human lung epithelial a549 cells. Cancer Lett. 2008, 270, 144–155. [Google Scholar] [CrossRef]
- Colacci, A.; Vaccari, M.; Silingardi, P.; Horn, W.; Mascolo, M.G.; Perdichizzi, S.; Rotondo, F.; Poluzzi, V.; Belladonna, V.; Grilli, S. In vitro endpoints to profile risks associated with waste incineration. Toxicol. Lett. 2007, 172, S113–S114. [Google Scholar]
- De Kok, T.M.; Hogervorst, J.G.; Briede, J.J.; van Herwijnen, M.H.; Maas, L.M.; Moonen, E.J.; Driece, H.A.; Kleinjans, J.C. Genotoxicity and physicochemical characteristics of traffic-related ambient particulate matter. Environ. Mol. Mutagen. 2005, 46, 71–80. [Google Scholar] [CrossRef]
- Don Porto Carero, A.; Hoet, P.H.; Verschaeve, L.; Schoeters, G.; Nemery, B. Genotoxic effects of carbon black particles, diesel exhaust particles, and urban air particulates and their extracts on a human alveolar epithelial cell line (a549) and a human monocytic cell line (thp-1). Environ. Mol. Mutagen. 2001, 37, 155–163. [Google Scholar] [CrossRef]
- Ohmori, K.; Sato, Y.; Nakajima, D.; Kageyama, S.; Shiraishi, F.; Fujimaki, T.; Goto, S. Characteristics of the transformation frequency at the tumor promotion stage of airborne particulate and gaseous matter at ten sites in japan. Environ. Sci. Process. Impacts 2013, 15, 1031–1040. [Google Scholar] [CrossRef]
- De Kok, T.M.; Driece, H.A.; Hogervorst, J.G.; Briede, J.J. Toxicological assessment of ambient and traffic-related particulate matter: A review of recent studies. Mutat. Res. 2006, 613, 103–122. [Google Scholar] [CrossRef]
- Englert, N. Fine particles and human health—A review of epidemiological studies. Toxicol. Lett. 2004, 149, 235–242. [Google Scholar] [CrossRef]
- Grahame, T.J.; Schlesinger, R.B. Health effects of airborne particulate matter: Do we know enough to consider regulating specific particle types or sources? Inhal. Toxicol. 2007, 19, 457–481. [Google Scholar] [CrossRef]
- Kampa, M.; Castanas, E. Human health effects of air pollution. Environ. Pollut. 2008, 151, 362–367. [Google Scholar] [CrossRef]
- Novak, J.; Hilscherova, K.; Landlova, L.; Cupr, P.; Kohut, L.; Giesy, J.P.; Klanova, J. Composition and effects of inhalable size fractions of atmospheric aerosols in the polluted atmosphere. Part ii. In vitro biological potencies. Environ. Int. 2014, 63, 64–70. [Google Scholar] [CrossRef]
- De Martinis, B.S.; Kado, N.Y.; de Carvalho, L.R.; Okamoto, R.A.; Gundel, L.A. Genotoxicity of fractionated organic material in airborne particles from sao paulo, brazil. Mutat. Res. 1999, 446, 83–94. [Google Scholar] [CrossRef]
- Zhao, X.; Wan, Z.; Zhu, H.; Chen, R. The carcinogenic potential of extractable organic matter from urban airborne particles in shanghai, china. Mutat. Res. 2003, 540, 107–117. [Google Scholar] [CrossRef]
- McDonald, J.D.; Eide, I.; Seagrave, J.; Zielinska, B.; Whitney, K.; Lawson, D.R.; Mauderly, J.L. Relationship between composition and toxicity of motor vehicle emission samples. Environ. Health Perspect. 2004, 112, 1527–1538. [Google Scholar] [CrossRef]
- Gabelova, A.; Valovicova, Z.; Horvathova, E.; Slamenova, D.; Binkova, B.; Sram, R.J.; Farmer, P.B. Genotoxicity of environmental air pollution in three european cities: Prague, Kosice and Sofia. Mutat. Res. 2004, 563, 49–59. [Google Scholar] [CrossRef]
- Sharma, A.K.; Jensen, K.A.; Rank, J.; White, P.A.; Lundstedt, S.; Gagne, R.; Jacobsen, N.R.; Kristiansen, J.; Vogel, U.; Wallin, H. Genotoxicity, inflammation and physico-chemical properties of fine particle samples from an incineration energy plant and urban air. Mutat. Res. 2007, 633, 95–111. [Google Scholar] [CrossRef]
- Kang, Y.; Cheung, K.C.; Wong, M.H. Polycyclic aromatic hydrocarbons (pahs) in different indoor dusts and their potential cytotoxicity based on two human cell lines. Environ. Int. 2010, 36, 542–547. [Google Scholar] [CrossRef]
- Asada, S.; Sasaki, K.; Tanaka, N.; Takeda, K.; Hayashi, M.; Umeda, M. Detection of initiating as well as promoting activity of chemicals by a novel cell transformation assay using v-ha-ras-transfected balb/c 3t3 cells (bhas 42 cells). Mutat. Res. 2005, 588, 7–21. [Google Scholar] [CrossRef]
- Sasaki, K.; Mizusawa, H.; Ishidate, M.; Tanaka, N. Establishment of a highly reproducible transformation assay of a ras-transfected balb 3t3 clone by treatment with promoters. Basic Life Sci. 1990, 52, 411–416. [Google Scholar]
- International Agency for Research on Cancer. IARC Monographs on the Evaluation of Carcinogenic Risks to Humans; WHO Document Production Services: Geneva, Switzerland, 2010. [Google Scholar]
- Maunders, H.; Patwardhan, S.; Phillips, J.; Clack, A.; Richter, A. Human bronchial epithelial cell transcriptome: Gene expression changes following acute exposure to whole cigarette smoke in vitro. Am. J. Physiol. Lung Cell Mol. Physiol. 2007, 292, L1248–L1256. [Google Scholar]
- Musafia-Jeknic, T.; Mahadevan, B.; Pereira, C.; Baird, W.M. Long-term effects of a standardized complex mixture of urban dust particulate on the metabolic activation of carcinogenic polycyclic aromatic hydrocarbons in human cells in culture. Toxicol. Sci. 2005, 88, 358–366. [Google Scholar] [CrossRef]
- Intelligent Systems and Bioinformatics Laboratory. Available online: http://vortex.cs.wayne.edu/projects.htm (accessed on 11 August 2014).
- Draghici, S.; Khatri, P.; Tarca, A.L.; Amin, K.; Done, A.; Voichita, C.; Georgescu, C.; Romero, R. A systems biology approach for pathway level analysis. Genome Res. 2007, 17, 1537–1545. [Google Scholar]
- KEGG Kyoto Encyclopedia of Genes and Genomes. Available online: http://www.genome.jp/kegg/ (accessed on 11 August 2014).
- Pietsch, E.C.; Sykes, S.M.; McMahon, S.B.; Murphy, M.E. The p53 family and programmed cell death. Oncogene 2008, 27, 6507–6521. [Google Scholar] [CrossRef]
- Bogdanos, D.P.; Smyk, D.S.; Invernizzi, P.; Rigopoulou, E.I.; Blank, M.; Sakkas, L.; Pouria, S.; Shoenfeld, Y. Tracing environmental markers of autoimmunity: Introducing the infectome. Immunol. Res. 2013, 56, 220–240. [Google Scholar] [CrossRef]
- Sun, W.; Jiao, Y.; Cui, B.; Gao, X.; Xia, Y.; Zhao, Y. Immune complexes activate human endothelium involving the cell-signaling hmgb1-rage axis in the pathogenesis of lupus vasculitis. Lab. Investig. 2013, 93, 626–638. [Google Scholar] [CrossRef]
- Huang, Y.C. The role of in vitro gene expression profiling in particulate matter health research. J Toxicol. Environ. Health B Crit. Rev. 2013, 16, 381–394. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Colacci, A.; Vaccari, M.; Mascolo, M.G.; Rotondo, F.; Morandi, E.; Quercioli, D.; Perdichizzi, S.; Zanzi, C.; Serra, S.; Poluzzi, V.; et al. Alternative Testing Methods for Predicting Health Risk from Environmental Exposures. Sustainability 2014, 6, 5265-5283. https://doi.org/10.3390/su6085265
Colacci A, Vaccari M, Mascolo MG, Rotondo F, Morandi E, Quercioli D, Perdichizzi S, Zanzi C, Serra S, Poluzzi V, et al. Alternative Testing Methods for Predicting Health Risk from Environmental Exposures. Sustainability. 2014; 6(8):5265-5283. https://doi.org/10.3390/su6085265
Chicago/Turabian StyleColacci, Annamaria, Monica Vaccari, Maria Grazia Mascolo, Francesca Rotondo, Elena Morandi, Daniele Quercioli, Stefania Perdichizzi, Cristina Zanzi, Stefania Serra, Vanes Poluzzi, and et al. 2014. "Alternative Testing Methods for Predicting Health Risk from Environmental Exposures" Sustainability 6, no. 8: 5265-5283. https://doi.org/10.3390/su6085265
APA StyleColacci, A., Vaccari, M., Mascolo, M. G., Rotondo, F., Morandi, E., Quercioli, D., Perdichizzi, S., Zanzi, C., Serra, S., Poluzzi, V., Angelini, P., Grilli, S., & Zinoni, F. (2014). Alternative Testing Methods for Predicting Health Risk from Environmental Exposures. Sustainability, 6(8), 5265-5283. https://doi.org/10.3390/su6085265