Temporal Variability and Geographical Origins of Airborne Pollen Grains Concentrations from 2015 to 2018 at Saclay, France

 ,
,  and
and
Abstract
:
1. Introduction
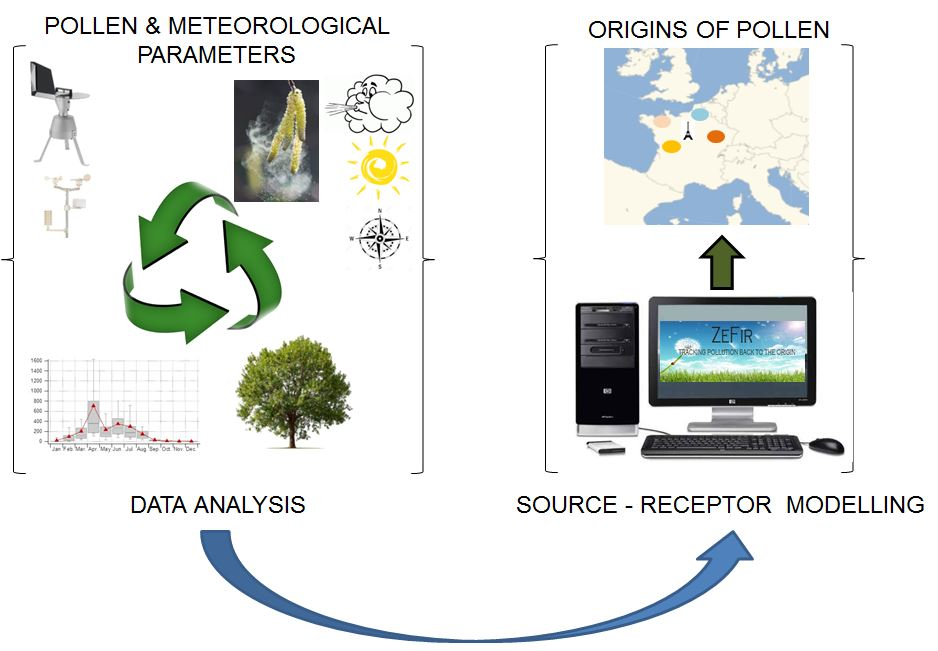
2. Materials and Methods
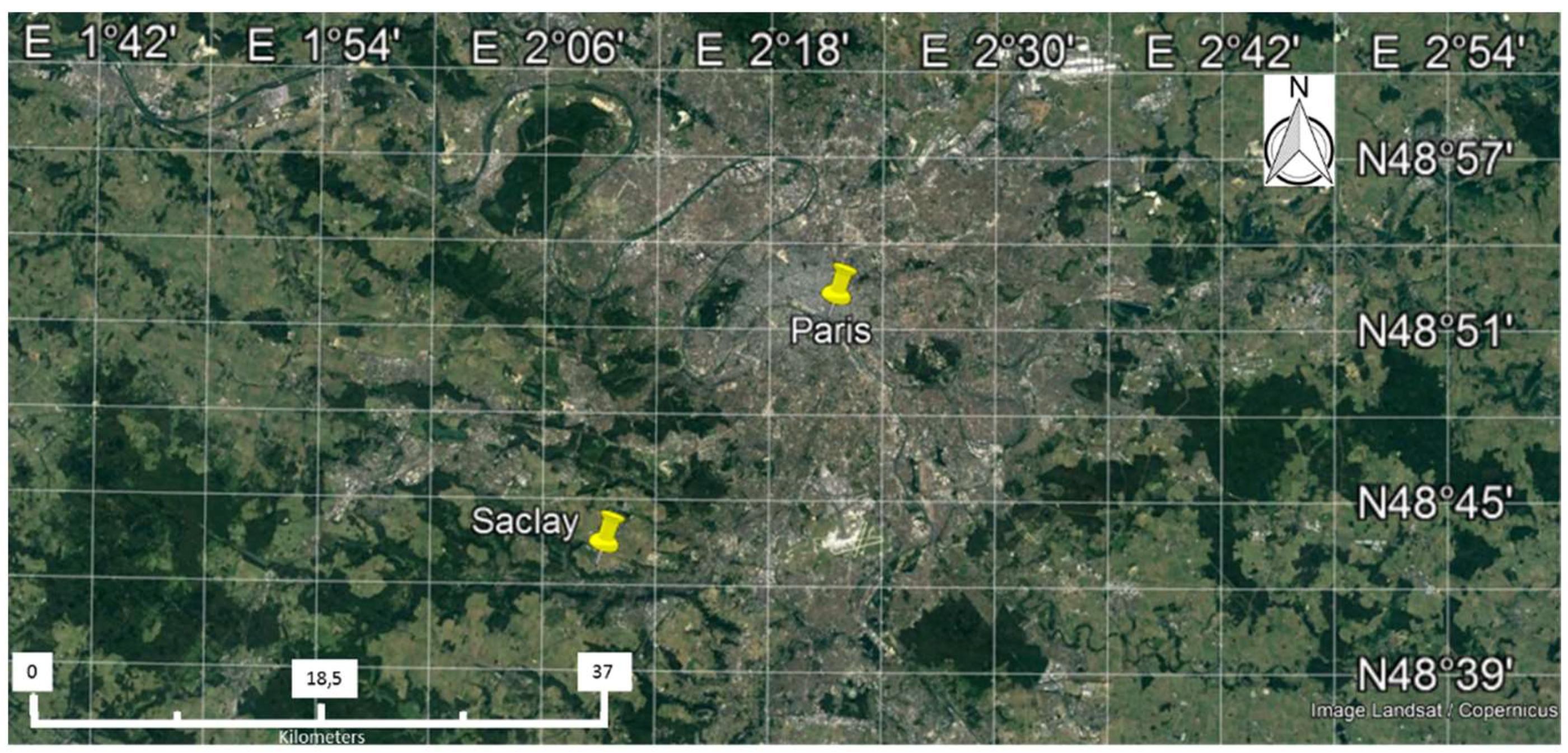
2.1. Experimental Site
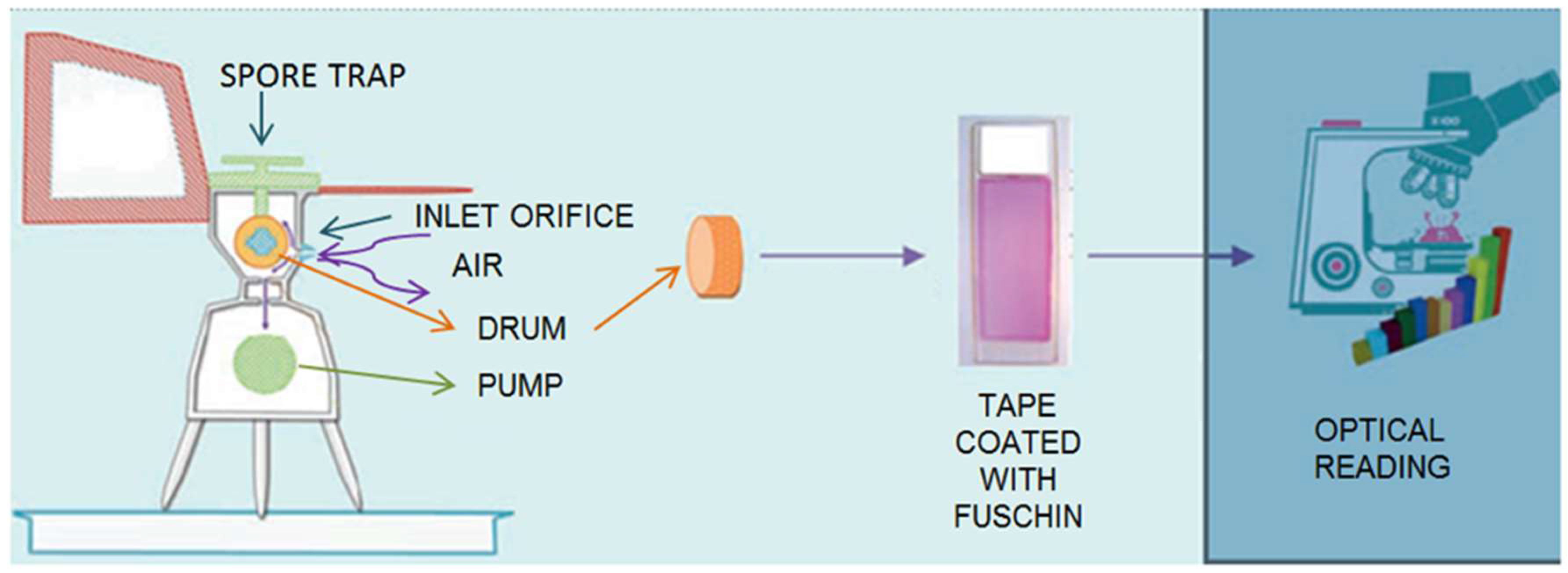
2.2. Pollen Grains Identification and Counting
2.3. Geographical Origins
3. Results
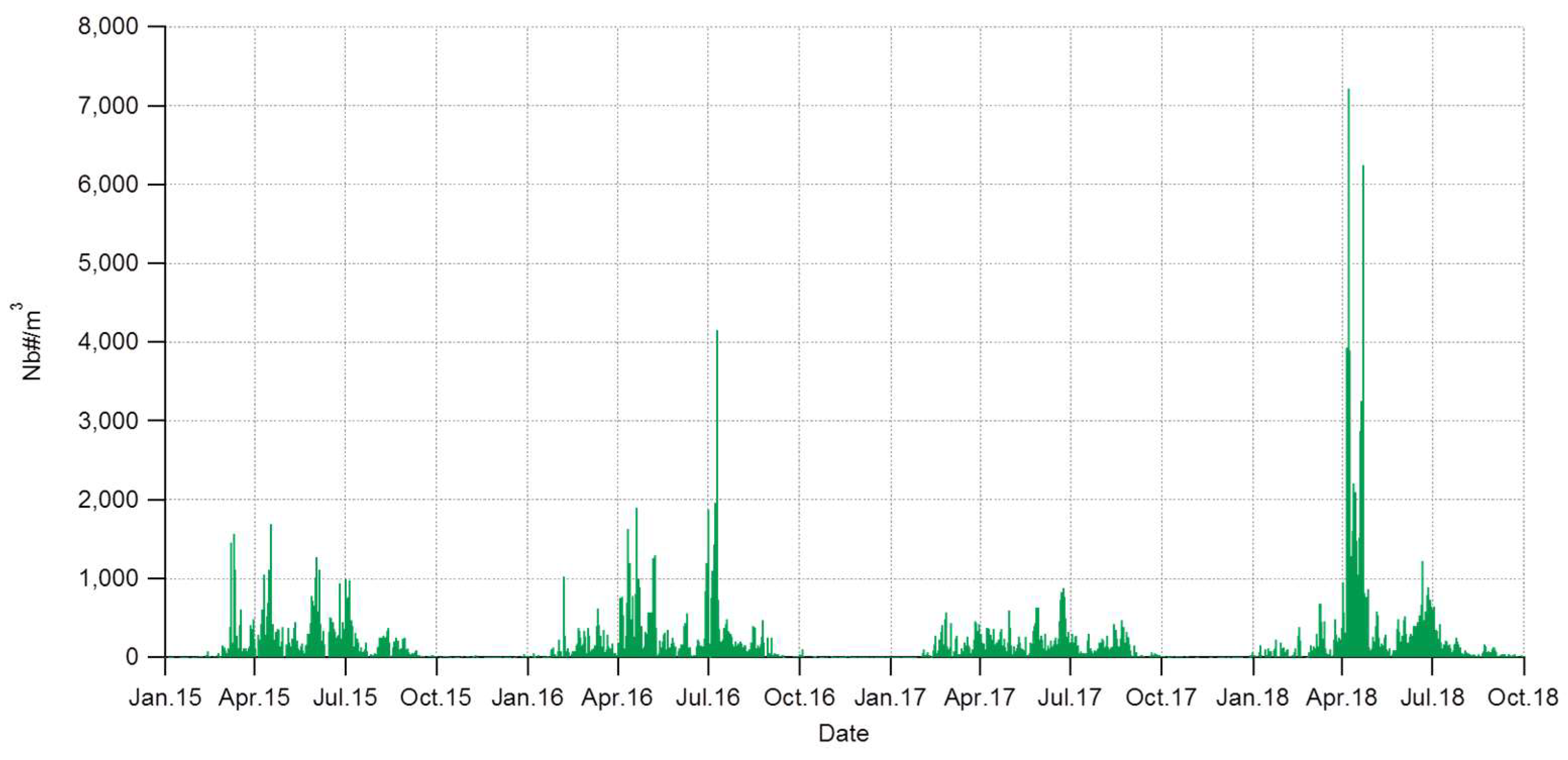
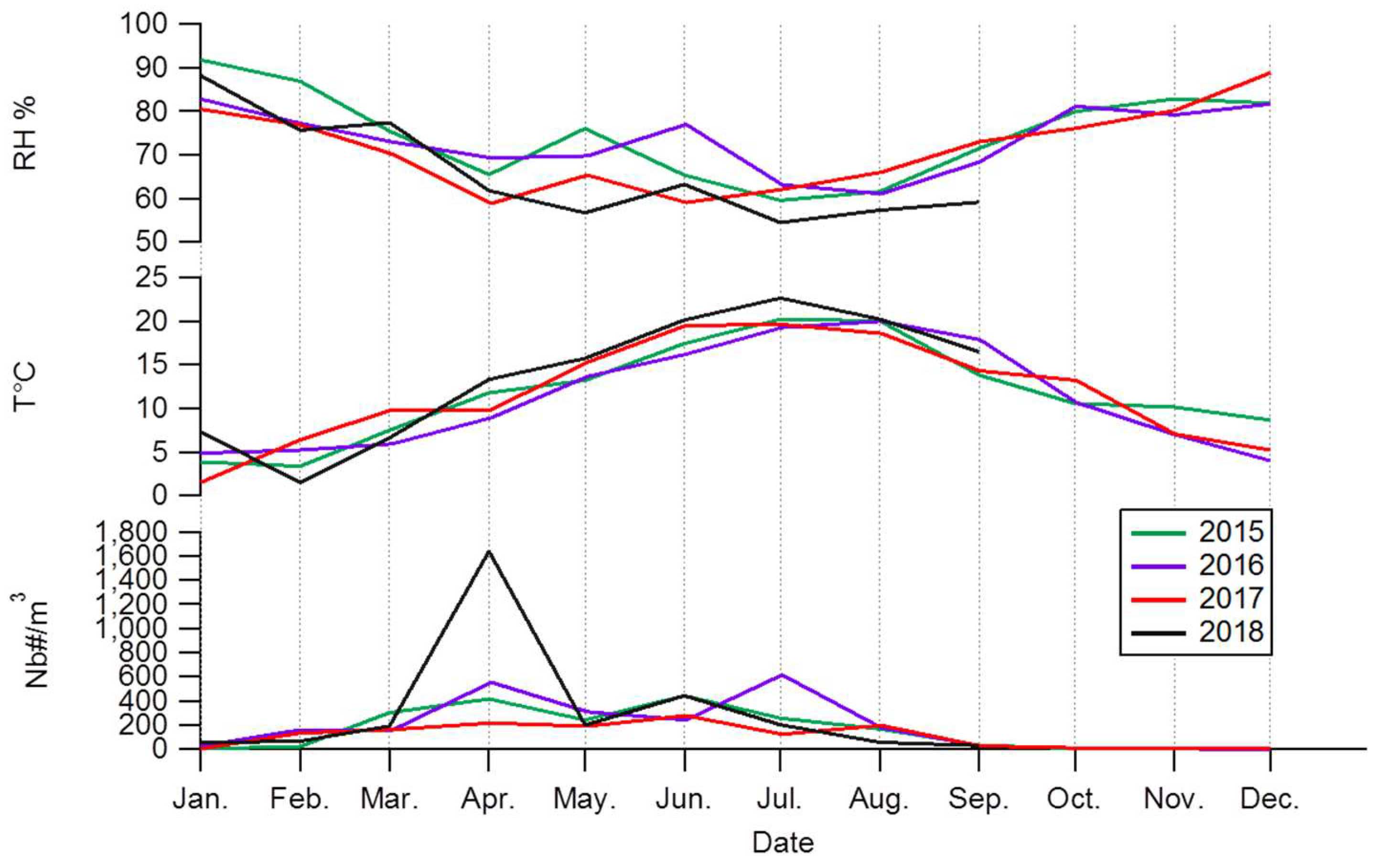
3.1. Interannuality of the Total Pollen Grains
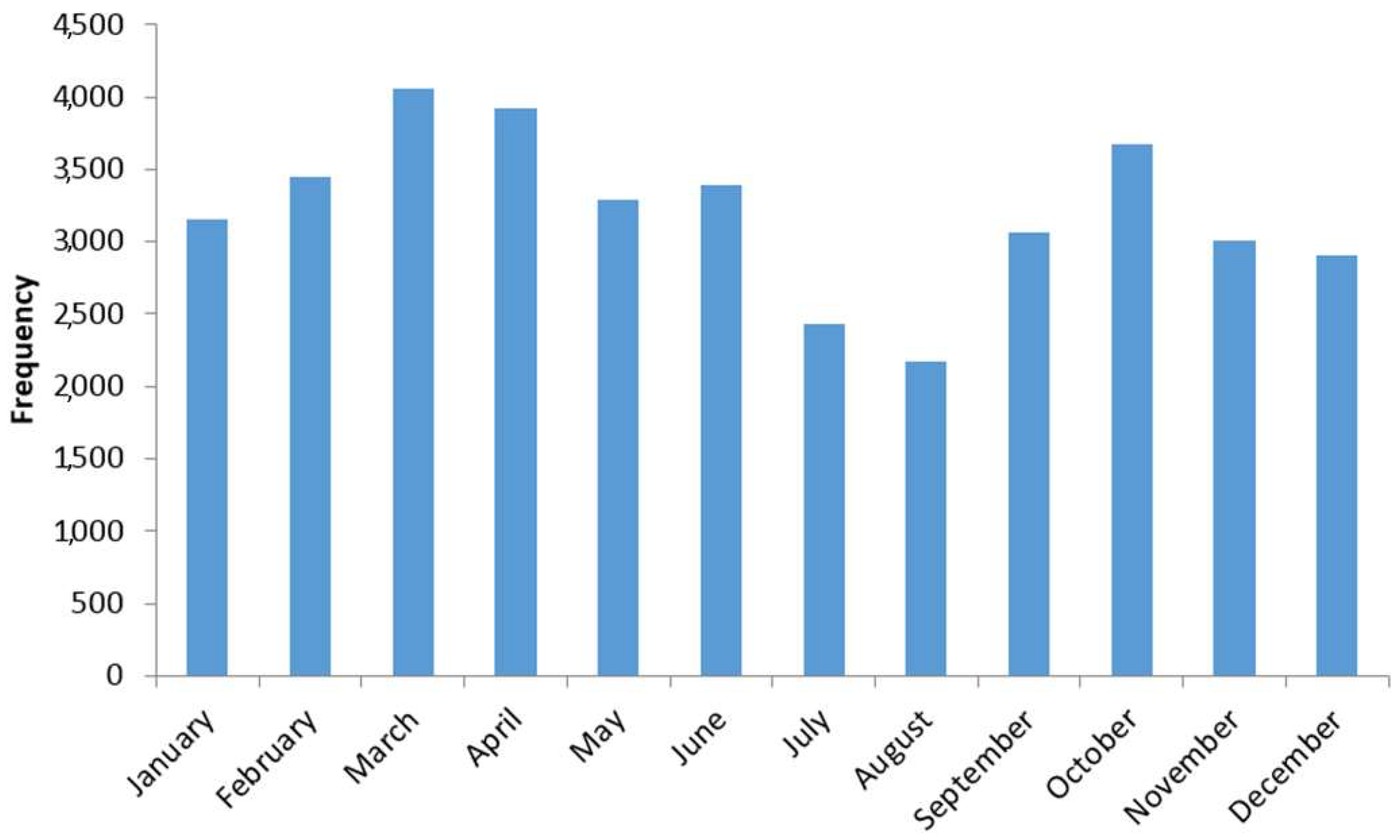
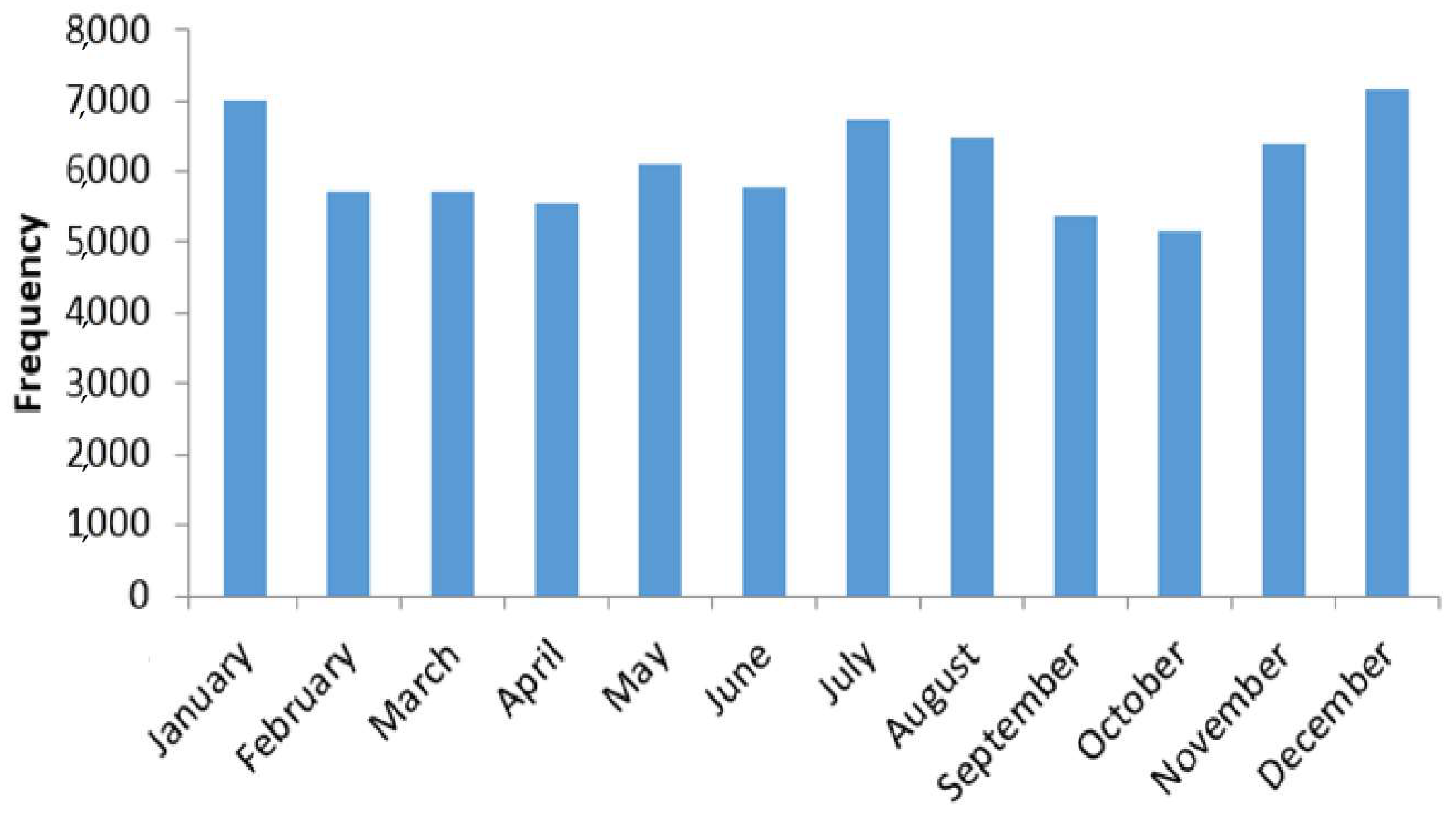
3.2. Seasonality of Total Pollen Grains Concentration at Saclay
3.3. Daily Variability of Total Pollen Grains Concentration at Saclay
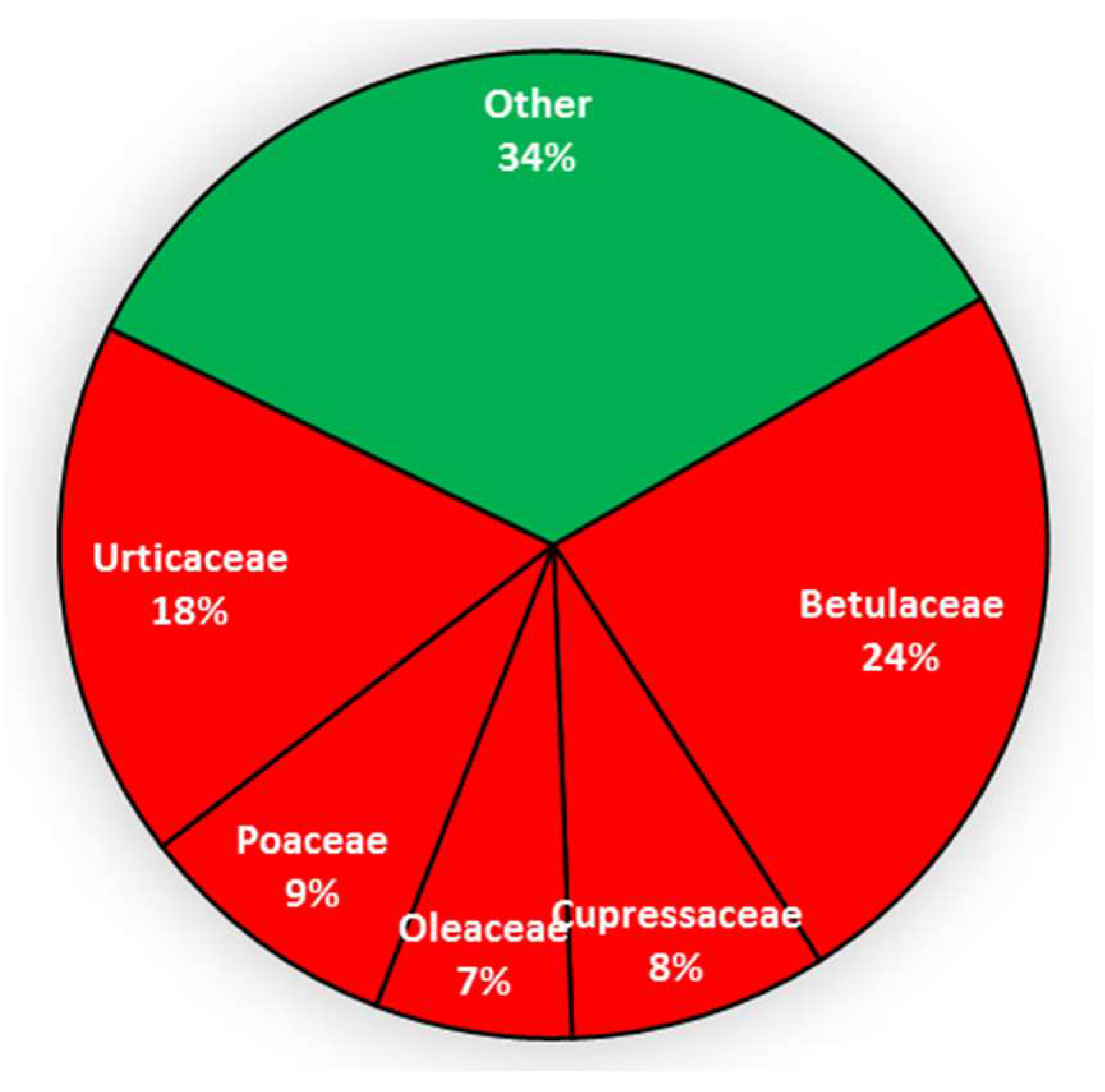
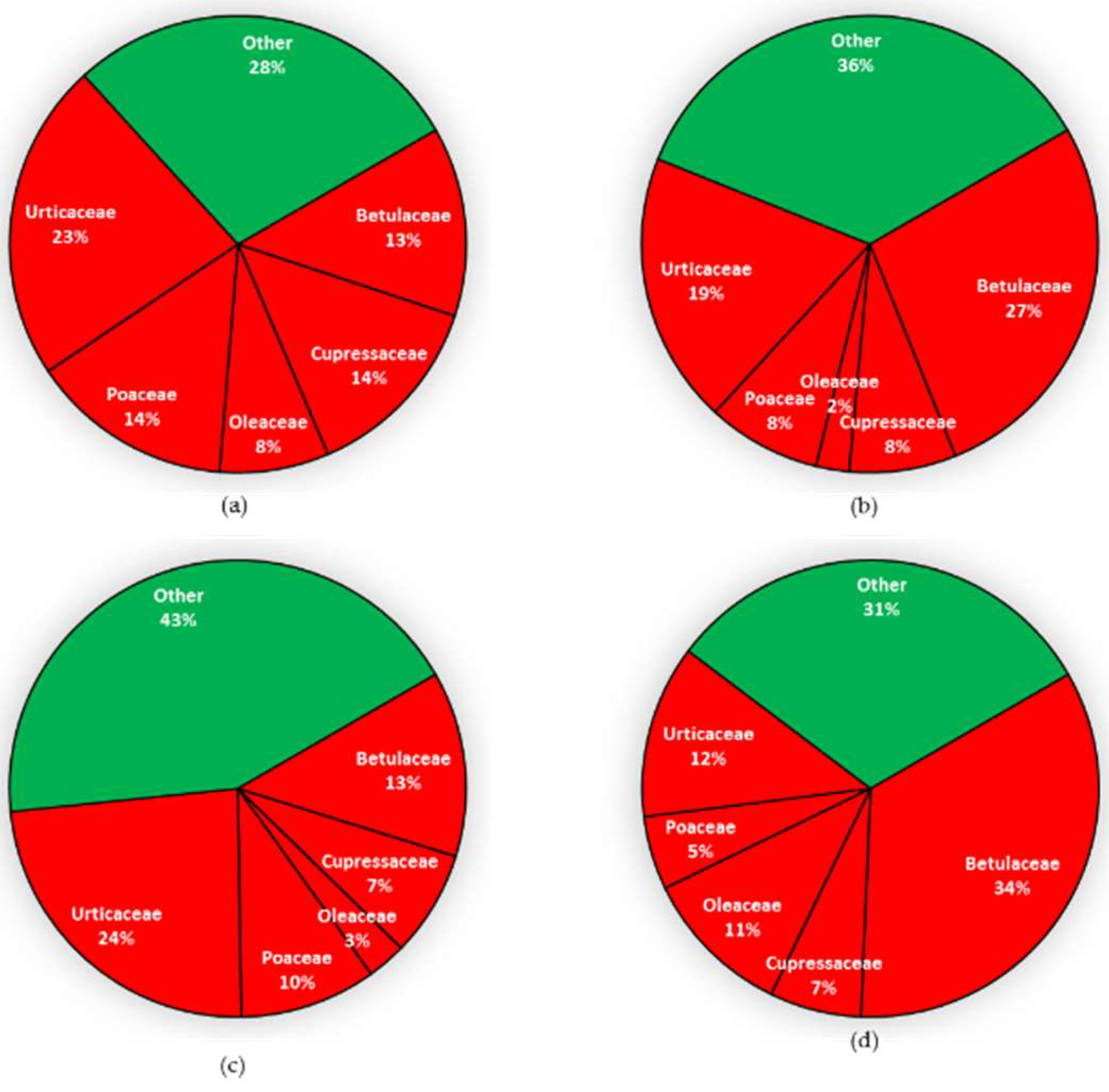
3.4. Allergenic Pollen Grains Abundance and Variability at Saclay
4. Discussion
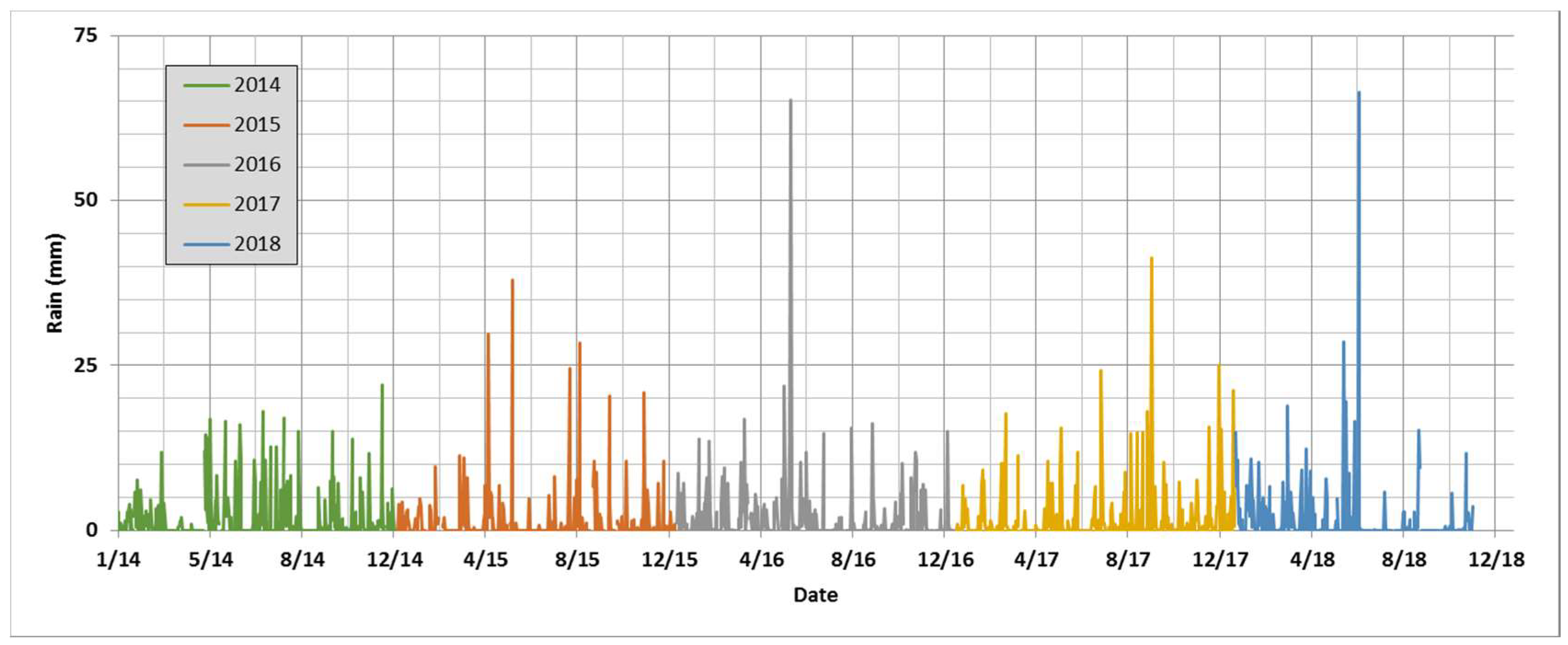
4.1. Interannuality and General Increase of Pollen Concentrations
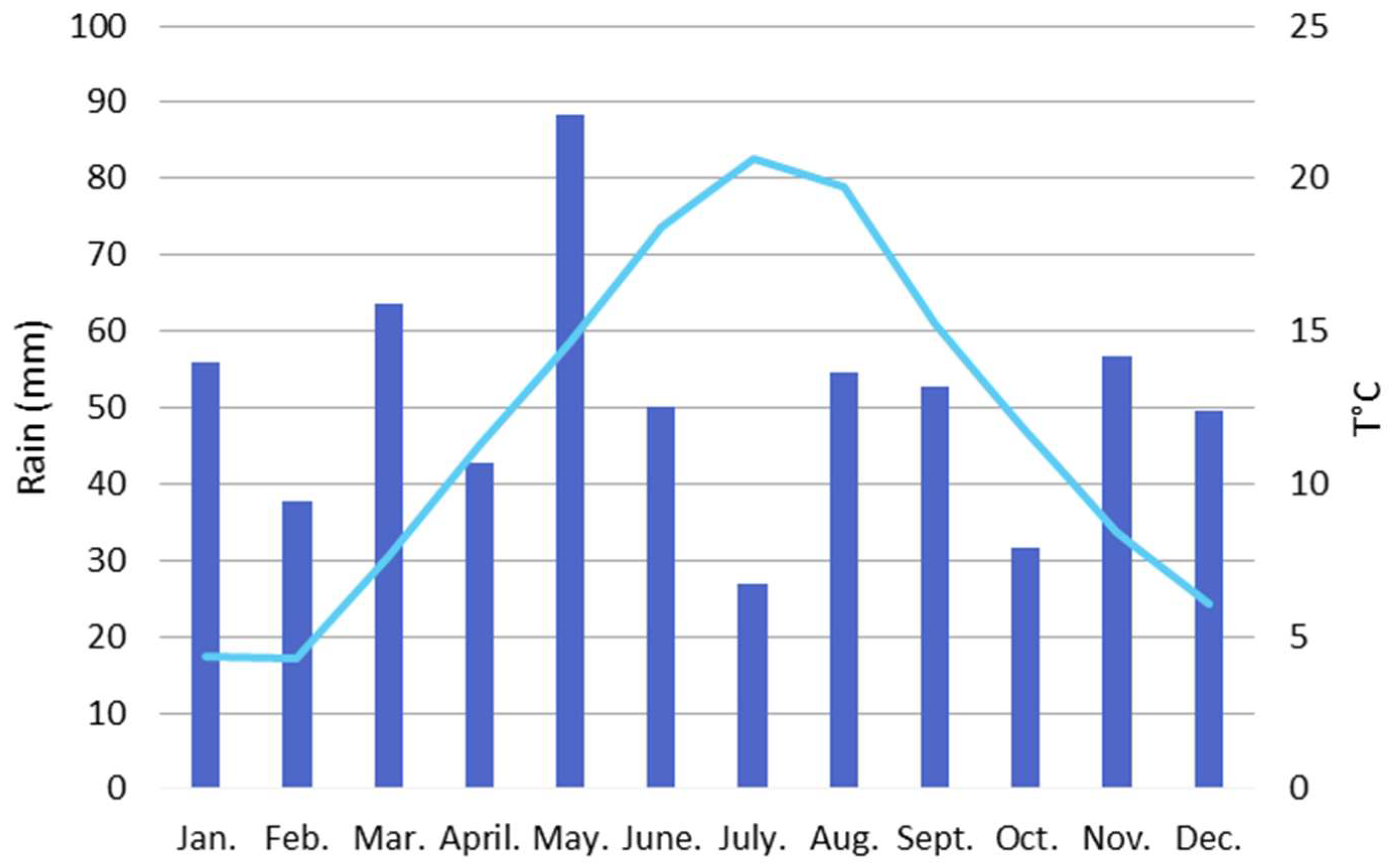
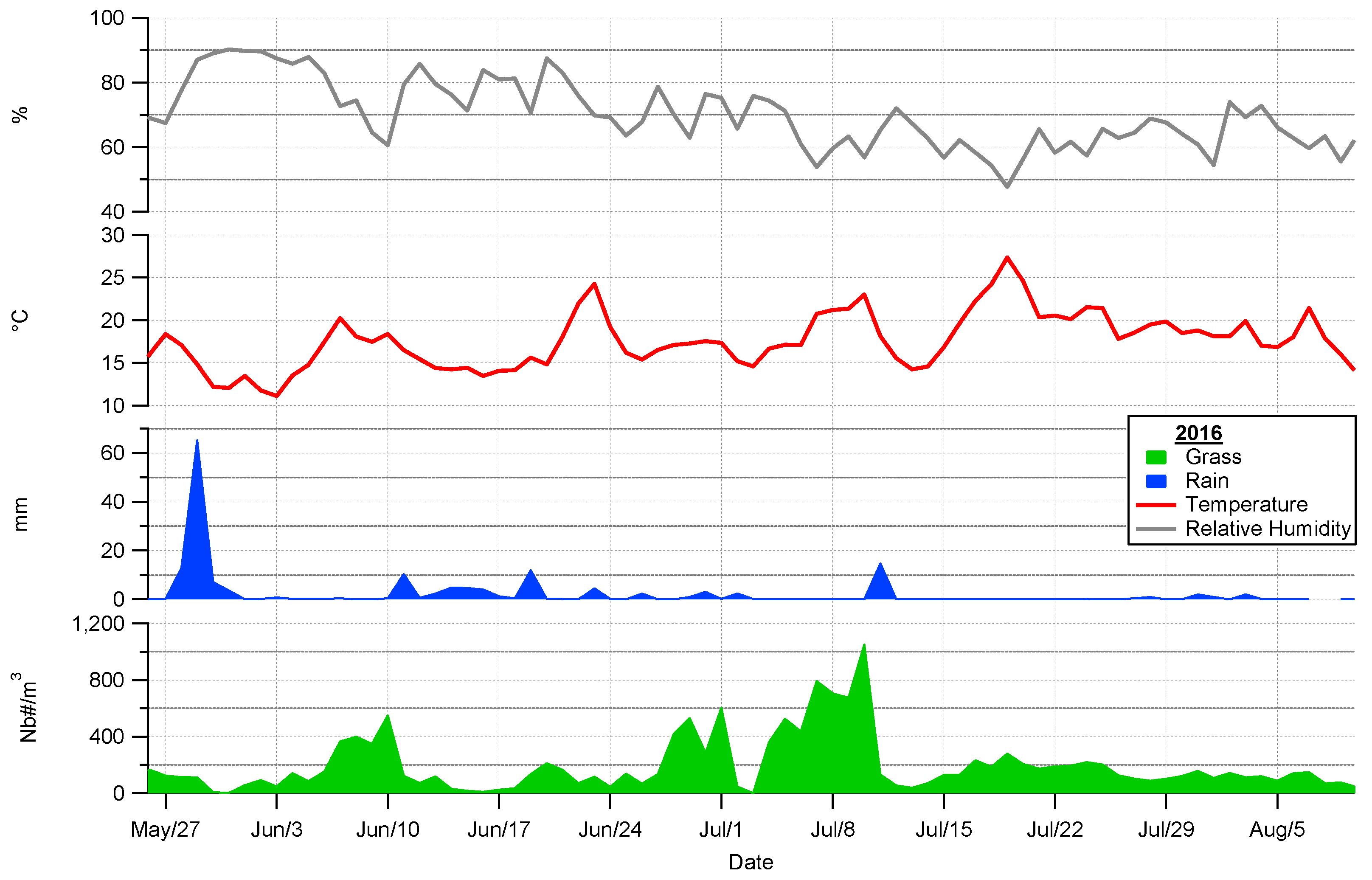
4.2. Pollen Seasonality Related to Air Temperature, Relative Humidity and Rain
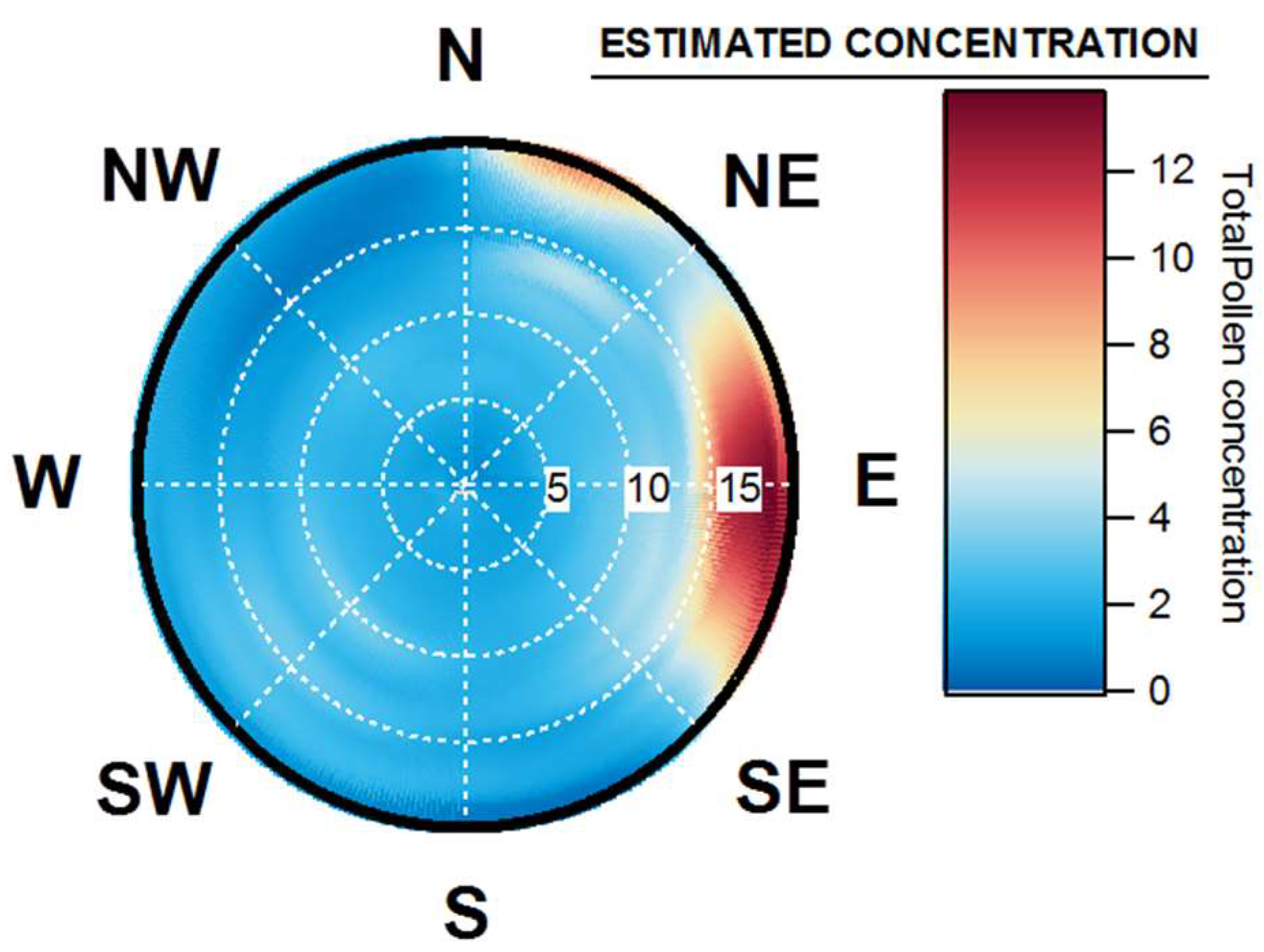
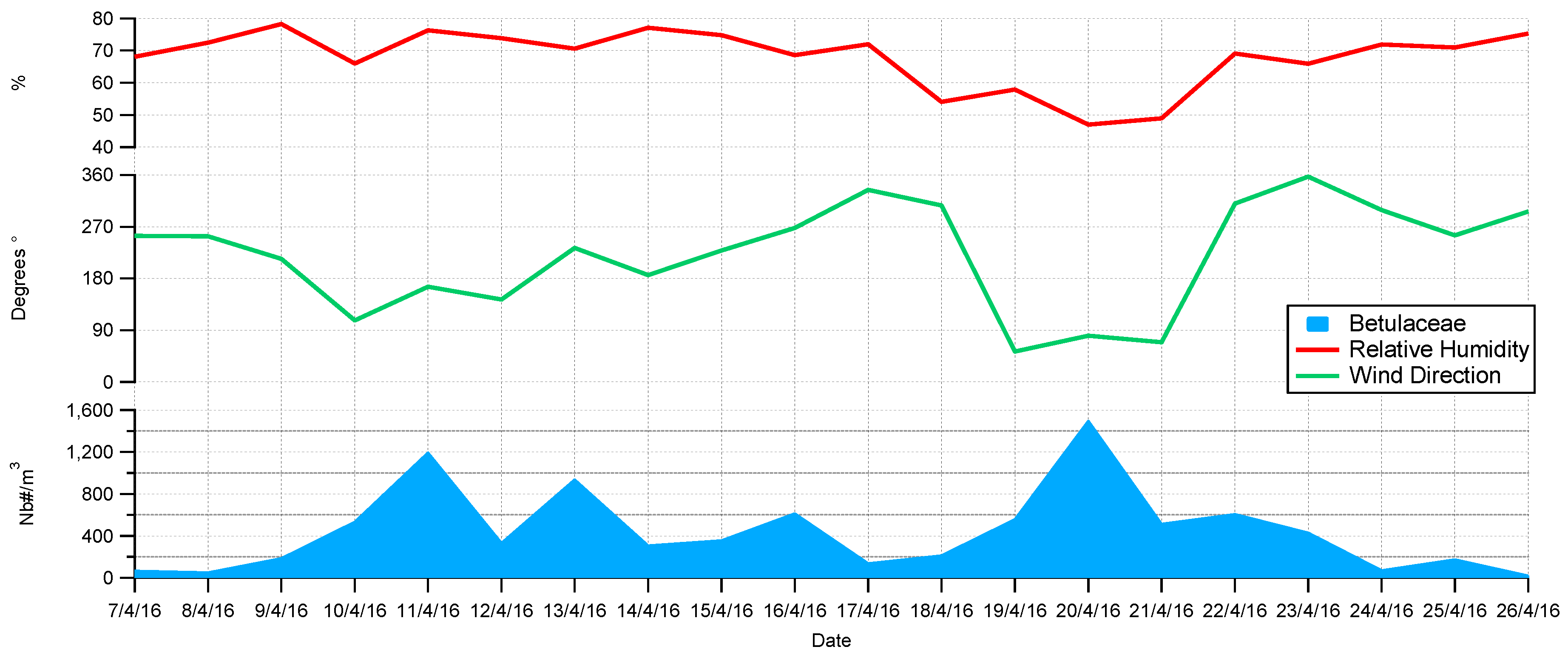
4.3. Wind Prevalence at Saclay
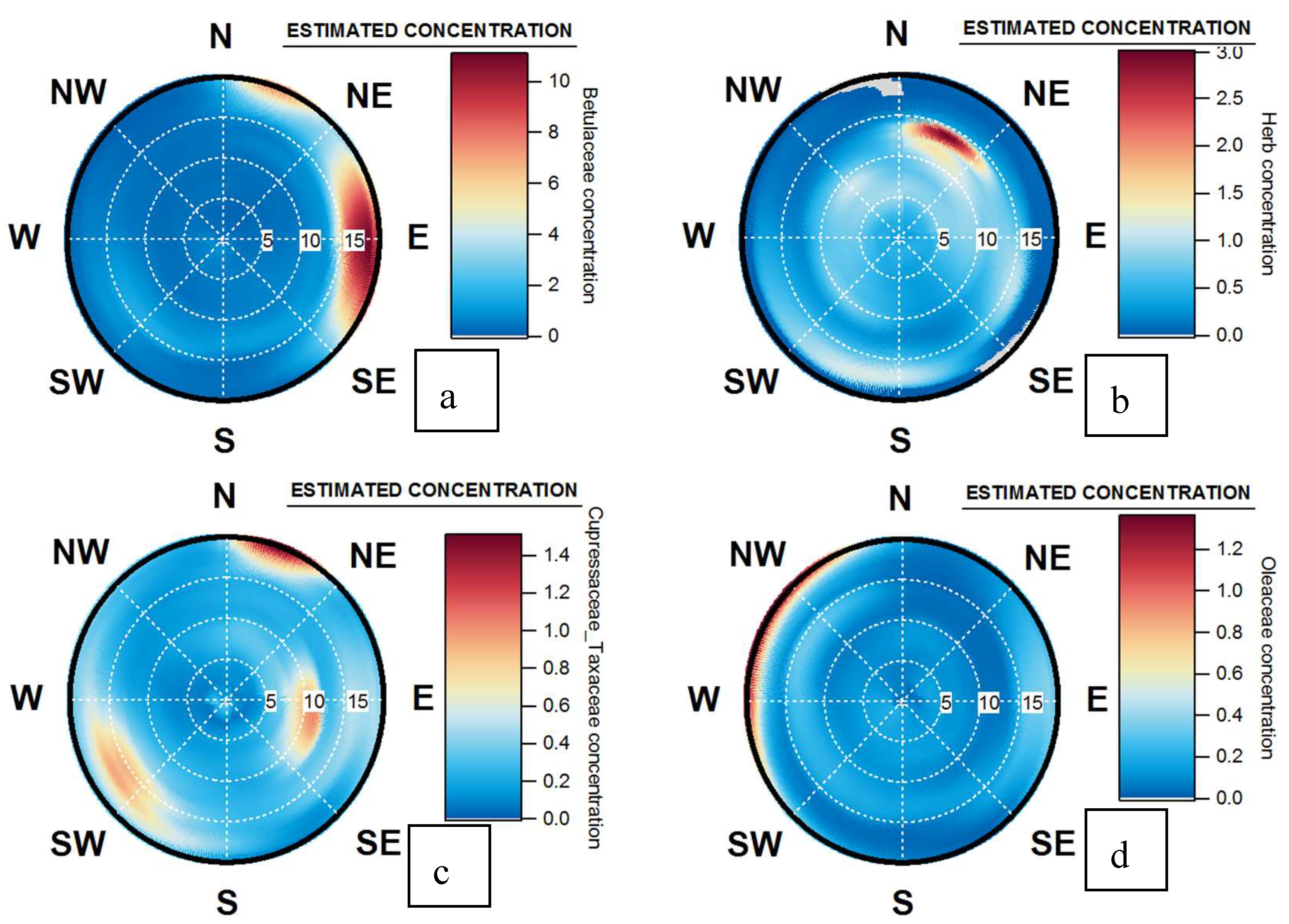
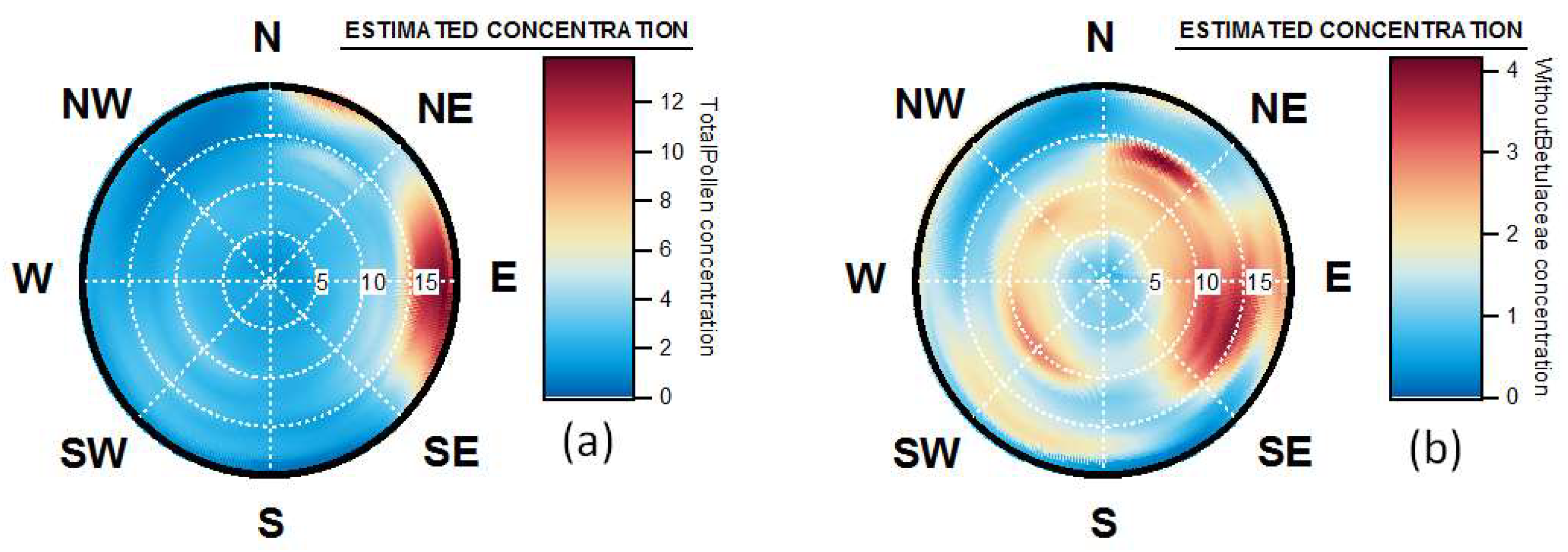
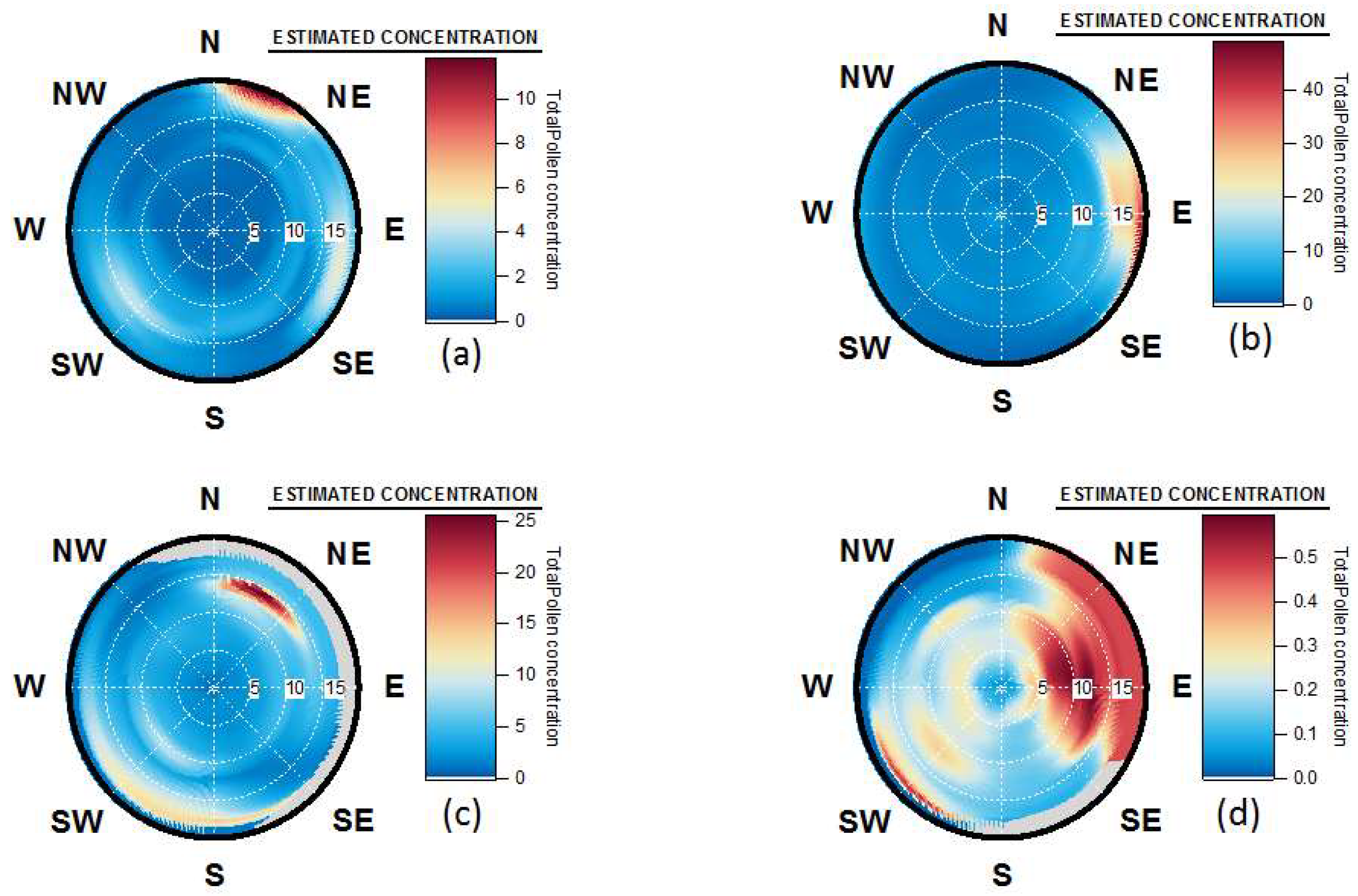
4.4. Origins and Point Sources of Pollen Grains
4.5. Wind Occurrence and Pollen Patterns
4.6. Pollen Point Sources, Long Range Transport and Allergen Transfers during Pollution Events
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 2015 | 2016 | 2017 | 2018 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Month | Nb#/m3 | std | Max | Nb#/m3 | std | Max | Nb#/m3 | std | Max | Nb#/m3 | std | Max |
| Jan. | 3.1 | 3.3 | 57.0 | 22.1 | 34.1 | 361.0 | 1.6 | 2.5 | 13.0 | 54.4 | 59.7 | 978.0 |
| Feb. | 21.1 | 34.0 | 359.0 | 103.3 | 199.4 | 2396.0 | 133.6 | 149.4 | 1426.0 | 67.1 | 93.1 | 771.0 |
| Mar. | 301.7 | 390.9 | 5854.0 | 158.9 | 145.4 | 1975.0 | 162.2 | 141.7 | 1586.0 | 191.3 | 184.1 | 1952.0 |
| Apri. | 400.9 | 373.3 | 5012.0 | 554.3 | 459.0 | 8255.0 | 217.5 | 119.0 | 3306.0 | 1638.6 | 1755.5 | 23317.0 |
| May | 240.0 | 197.5 | 2359.0 | 309.8 | 350.1 | 3898.0 | 189.0 | 163.4 | 1632.0 | 199.2 | 148.9 | 1964.0 |
| June | 384.5 | 321.4 | 4634.0 | 241.6 | 297.1 | 2517.0 | 269.4 | 236.9 | 5116.0 | 443.3 | 252.1 | 5367.0 |
| July | 238.6 | 284.1 | 5137.0 | 615.5 | 826.0 | 10551.0 | 120.3 | 78.9 | 1418.0 | 202.6 | 146.9 | 3689.0 |
| Aug. | 123.9 | 113.4 | 4252.0 | 159.1 | 117.3 | 3942.0 | 197.3 | 109.2 | 5408.0 | 55.8 | 36.2 | 1374.0 |
| Sept. | 32.8 | 27.5 | 648.0 | 26.7 | 45.0 | 477.0 | 28.1 | 30.2 | 438.0 | 28.4 | 26.2 | 485.0 |
| Oct. | 5.4 | 5.1 | 52.0 | 10.6 | 18.7 | 227.0 | 8.7 | 5.4 | 77.0 | |||
| Nov. | 4.7 | 6.6 | 50.0 | 2.6 | 3.7 | 30.0 | 2.8 | 2.0 | 16.0 | |||
| Dec. | 7.1 | 10.2 | 121.0 | 1.1 | 1.4 | 9.0 | 4.6 | 12.5 | 76.0 | |||
| 2015 | 2016 | 2017 | 2018 | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Month | WD | WD_std | WS | WS_std | WD | WD_std | WS | WS_std | WD | WD_std | WS | WS_std | WD | WD_std | WS | WS_std |
| Jan. | 234.5 | 63.1 | 1.8 | 1.5 | 217.9 | 57.7 | 1.9 | 1.3 | 230.4 | 99.6 | 0.3 | 1.0 | 240.6 | 64.2 | 2.0 | 1.5 |
| Feb. | 277.0 | 90.4 | 0.4 | 1.2 | 239.6 | 77.0 | 1.4 | 1.4 | 220.1 | 84.5 | 1.0 | 1.4 | 44.7 | 91.7 | 0.6 | 1.0 |
| Mar. | 297.9 | 86.6 | 0.7 | 1.4 | 312.8 | 85.6 | 0.4 | 1.2 | 233.3 | 88.4 | 0.8 | 1.2 | 195.3 | 83.7 | 0.7 | 1.1 |
| April | 7.7 | 93.9 | 0.3 | 1.1 | 243.7 | 87.7 | 0.5 | 1.1 | 348.0 | 64.2 | 0.8 | 0.8 | 209.9 | 87.0 | 0.8 | 1.1 |
| May | 255.4 | 75.2 | 1.0 | 1.1 | 304.1 | 91.0 | 0.3 | 1.0 | 216.5 | 102.0 | 0.1 | 0.9 | 353.4 | 73.4 | 0.7 | 0.8 |
| June | 323.6 | 78.8 | 0.6 | 1.0 | 266.2 | 67.0 | 1.0 | 0.8 | 253.7 | 84.1 | 0.8 | 1.1 | 13.7 | 63.8 | 0.9 | 0.8 |
| July | 262.3 | 75.2 | 1.0 | 1.0 | 271.1 | 60.9 | 1.0 | 0.9 | 253.3 | 72.9 | 1.0 | 1.0 | 330.4 | 76.7 | 0.5 | 0.8 |
| Aug. | 217.5 | 101.5 | 0.3 | 1.0 | 269.8 | 74.0 | 0.8 | 0.9 | 249.6 | 82.2 | 0.7 | 1.0 | 271.1 | 78.1 | 0.9 | 1.0 |
| Sept. | 292.0 | 93.3 | 0.1 | 1.1 | 228.5 | 89.7 | 0.4 | 0.9 | 237.7 | 65.4 | 1.2 | 1.2 | 309.3 | 81.5 | 0.6 | 0.9 |
| Oct. | 78.6 | 95.2 | 0.2 | 0.8 | 57.4 | 87.6 | 0.5 | 0.9 | 235.8 | 67.7 | 1.2 | 1.1 | ||||
| Nov. | 228.9 | 56.8 | 1.9 | 1.3 | 217.6 | 96.4 | 0.5 | 1.3 | 251.1 | 63.3 | 1.3 | 1.3 | ||||
| Dec. | 201.9 | 33.3 | 2.1 | 0.9 | 157.4 | 84.8 | 0.5 | 0.7 | 244.2 | 50.9 | 1.9 | 1.5 | ||||
| 2015 | 2016 | 2017 | 2018 | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Month | T | T_std | RH | RH_std | T | T_std | RH | RH_std | T | T_std | RH | RH_std | T | T_std | RH | RH_std |
| Jan. | 3.9 | 3.7 | 91.6 | 8.5 | 4.9 | 3.6 | 89.5 | 8.8 | 1.2 | 3.8 | 86.8 | 12.5 | 7.3 | 2.5 | 90.7 | 8.0 |
| Feb. | 3.4 | 2.8 | 86.9 | 13.7 | 5.4 | 3.6 | 84.9 | 13.7 | 6.6 | 3.6 | 83.1 | 10.7 | 1.7 | 3.2 | 79.6 | 18.0 |
| Mar. | 7.5 | 3.4 | 75.4 | 17.8 | 6.3 | 2.8 | 77.5 | 15.8 | 10.2 | 3.6 | 75.6 | 16.6 | 6.6 | 4.1 | 79.7 | 14.2 |
| April | 12.0 | 4.7 | 63.9 | 23.0 | 9.1 | 4.0 | 76.0 | 17.4 | 10.3 | 4.2 | 64.0 | 18.3 | 13.6 | 4.8 | 69.2 | 18.6 |
| May | 13.3 | 3.8 | 75.1 | 17.6 | 13.9 | 4.1 | 76.5 | 21.4 | 15.7 | 5.5 | 76.1 | 17.8 | 15.6 | 5.1 | 68.4 | 19.7 |
| June | 17.3 | 4.6 | 65.6 | 18.0 | 16.3 | 3.7 | 85.9 | 16.0 | 20.0 | 5.3 | 68.0 | 17.3 | 20.0 | 4.2 | 62.2 | 18.0 |
| July | 20.6 | 5.3 | 63.0 | 18.9 | 19.3 | 4.6 | 71.4 | 18.1 | 20.1 | 4.6 | 67.7 | 17.8 | 22.8 | 4.2 | 58.0 | 17.7 |
| Aug. | 20.2 | 4.9 | 65.0 | 21.4 | 19.4 | 4.7 | 68.3 | 19.4 | 19.0 | 4.3 | 72.8 | 18.4 | 20.4 | 5.0 | 57.2 | 17.3 |
| Sept. | 14.0 | 3.0 | 77.1 | 18.1 | 16.2 | 3.3 | 75.3 | 18.5 | 14.5 | 3.5 | 82.0 | 14.2 | 16.4 | 2.8 | 59.2 | 7.8 |
| Oct. | 10.7 | 3.3 | 85.9 | 14.5 | 10.7 | 3.0 | 82.6 | 14.9 | 13.7 | 3.6 | 84.8 | 12.6 | ||||
| Nov. | 10.2 | 4.6 | 89.9 | 10.2 | 7.4 | 3.2 | 85.5 | 12.6 | 7.7 | 3.3 | 86.5 | 11.0 | ||||
| Dec. | 8.8 | 2.9 | 88.5 | 9.7 | 4.4 | 3.2 | 87.5 | 14.4 | 5.0 | 3.3 | 92.4 | 7.1 | ||||
| 2014 | 2015 | 2016 | 2017 | 2018 | |
|---|---|---|---|---|---|
| Jan. | 52.4 | 36.4 | 52.2 | 24.0 | 110.7 |
| Feb. | 47.1 | 32.1 | 49.7 | 36.4 | 33.1 |
| Mar. | 11.4 | 31.3 | 82.2 | 68.9 | 71.9 |
| April | 49.6 | 56.4 | 51.0 | 16.9 | 46.6 |
| May | 80.2 | 77.5 | 153.7 | 53.4 | 69.1 |
| June | 74.7 | 6.2 | 54.2 | 36.4 | 103.6 |
| July | 80.8 | 16.3 | 21.3 | 58.2 | 11.7 |
| Aug. | 81.5 | 89.4 | 27.7 | 64.4 | 36.5 |
| Sept. | 14.3 | 68.0 | 32.8 | 110.5 | 0.0 |
| Oct. | 55.7 | 41.9 | 30.4 | 29.5 | 24.6 |
| Nov. | 53.6 | 51.8 | 70.4 | 47.7 | |
| Dec. | 50.0 | 24.2 | 21.9 | 102.4 | |
| Total | 651.3 | 531.5 | 647.5 | 648.7 | 507.8 |
References
- Núñez, A.; Amo de Paz, G.; Rastrojo, A.; García Ruiz, A.M.; Alcamí, A.; Gutiérrez-Bustillo, A.M.; Moreno Gómez, D.A. Monitoring of airborne biological particles in outdoor atmosphere. Part 2: Metagenomics applied to urban environments. Int. Microbiol. 2016, 19, 69–80. [Google Scholar] [PubMed]
- Clot, B. Trends in airborne pollen: An overview of 21 years of data in Neuchâtel (Switzerland). Aerobiologia 2003, 19, 227–234. [Google Scholar] [CrossRef]
- D’Amato, G.; Cecchi, L.; D’Amato, M.; Liccardi, G. Urban air pollution and climate change as environmental risk factors of respiratory allergy: An update. J. Investig. Allergol. Clin. Immunol. 2010, 20, 95–102. [Google Scholar] [PubMed]
- García-Mozo, H. Poaceae pollen as the leading aeroallergen worldwide: A review. Allergy 2017, 72, 1849–1858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cecchi, L.; D’Amato, G.; Ayres, J.G.; Galan, C.; Forastiere, F.; Forsberg, B.; Gerritsen, J.; Nunes, C.; Behrendt, H.; Akdi, C.; et al. Projections of the effects of climate change on allergic asthma: The contribution of aerobiology. Allergy 2010, 65, 1073–1081. [Google Scholar] [CrossRef] [PubMed]
- D’Amato, G.; Cecchi, L.; Bonini, S.; Nunes, C.; Annesi-Maesano, I.; Behrendt, H.; Van Cauwenberge, P. Allergenic pollen and pollen allergy in Europe. Allergy 2007, 62, 976–990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Mozo, H.; Oteros, J.A.; Galán, C. Impact of land cover changes and climate on the main airborne pollen types in Southern Spain. Sci. Total Environ. 2016, 548, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Tormo-Molina, R.; Rodríguez, A.M.; Palaciso, I.S.; López, F.G. Pollen production in anemophilous trees. Grana 1996, 35, 38–46. [Google Scholar] [CrossRef] [Green Version]
- Vázquez, L.M.; Galán, C.; Domínguez-Vilches, E. Influence of meteorological parameters on olea pollen concentrations in Córdoba (South-western Spain). Int. J. Biometeorol. 2003, 48, 83–90. [Google Scholar] [CrossRef]
- Puc, M.; Bosiacka, B. Effects of meteorological factors and air pollution on urban pollen concentrations. Pol. J. Environ. Stud. 2011, 20, 611–618. [Google Scholar]
- Latałowa, M.; Miętus, M.; Uruska, A. Seasonal variations in the atmospheric Betula pollen count in Gdańsk (southern Baltic coast) in relation to meteorological parameters. Aerobiologia 2002, 18, 33–43. [Google Scholar] [CrossRef]
- Drake, J.E.; Tjoelker, M.G.; Vårhammar, A.; Medlyn, B.E.; Reich, P.B.; Leigh, A.; Pfautsc, S.; Blackman, C.J.; López, R.; et al. Trees tolerate an extreme heatwave via sustained transpirational cooling and increased leaf thermal tolerance. Glob. Chang. Boil. 2018, 24, 2390–2402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berggren, B.; Nilsson, S.; Boëthius, G. Diurnal variation of airborne birch pollen at some sites in Sweden. Grana 1995, 34, 251–259. [Google Scholar] [CrossRef]
- García-Mozo, H.; Galán, C.; Jato, V.; Belmonte, J.; de la Guardia, C.D.; Fernández, D.; Gutiérrez, M.; Aira, M.J.D.M.; Roure, J.M.; Ruiz, L.; et al. Quercus pollen season dynamics in the Iberian Peninsula: Response to meteorological parameters and possible consequences of climate change. Ann. Agric. Environ. Med. 2006, 13, 209. [Google Scholar] [PubMed]
- García-Mozo, H.; Yaezel, L.; Oteros, J.; Galán, C. Statistical approach to the analysis of olive long-term pollen season trends in southern Spain. Sci. Total Environ. 2014, 473, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Grote, M.; Vrtala, S.; Niederberger, V.; Wiermann, R.; Valenta, R.; Reichelt, R. Release of allergen-bearing cytoplasm from hydrated pollen: A mechanism common to a variety of grass (Poaceae) species revealed by electron microscopy. J. Allergy Clin. Immunol. 2001, 108, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Motta, A.C.; Marliere, M.; Peltre, G.; Sterenberg, P.A.; Lacroix, G. Traffic-related air pollutants induce the release of allergen-containing cytoplasmic granules from grass pollen. Int. Arch. Allergy Immunol. 2006, 139, 294–298. [Google Scholar] [CrossRef] [PubMed]
- Damialis, A.; Halley, J.M.; Gioulekas, D.; Vokou, D. Long-term trends in atmospheric pollen levels in the city of Thessaloniki, Greece. Atmos. Environ. 2007, 41, 7011–7021. [Google Scholar] [CrossRef]
- Grundström, M.; Dahl, Å.; Ou, T.; Chen, D.; Pleijel, H. The relationship between birch pollen, air pollution and weather types and their effect on antihistamine purchase in two Swedish cities. Aerobiologia 2017, 33, 457–471. [Google Scholar] [CrossRef] [PubMed]
- Kruczek, A.; Puc, M.; Wolski, T. Airborne pollen from allergenic herbaceous plants in urban and rural areas of Western Pomerania, NW Poland. Grana 2017, 56, 71–80. [Google Scholar] [CrossRef]
- Borycka, K.; Kasprzyk, I. Hourly pattern of allergenic alder and birch pollen concentrations in the air: Spatial differentiation and the effect of meteorological conditions. Atmos. Environ. 2018, 182, 179–192. [Google Scholar] [CrossRef]
- Kubik-Komar, A.; Piotrowska-Weryszko, K.; Weryszko-Chmielewska, E.; Kaszewski, B.M. Analysis of Fraxinus pollen seasons and forecast models based on meteorological factors. Ann. Agric. Environ. Med. 2018, 25, 285–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wozniak, M.C.; Solmon, F.; Steiner, A.L. Pollen rupture and its impact on precipitation in clean continental conditions. Geophys. Res. Lett. 2018, 45, 7156–7164. [Google Scholar] [CrossRef]
- Sofiev, M. On impact of transport conditions on variability of the seasonal pollen index. Aerobiologia 2017, 33, 167–179. [Google Scholar] [CrossRef] [PubMed]
- Rojo, J.; Rapp, A.; Lara, B.; Fernández-González, F.; Pérez-Badia, R. Effect of land uses and wind direction on the contribution of local sources to airborne pollen. Sci. Total Environ. 2015, 538, 672–682. [Google Scholar] [CrossRef] [PubMed]
- Tseng, Y.T.; Kawashima, S.; Kobayashi, S.; Takeuchi, S.; Nakamura, K. Algorithm for forecasting the total amount of airborne birch pollen from meteorological conditions of previous years. Agric. For. Meteorol. 2018, 249, 35–43. [Google Scholar] [CrossRef]
- Mesa, J.A.S.; Smith, M.; Emberlin, J.; Allitt, U.; Caulton, E.; Galan, C. Characteristics of grass pollen seasons in areas of southern Spain and the United Kingdom. Aerobiologia 2003, 19, 243–250. [Google Scholar] [CrossRef]
- Rodríguez-Rajo, F.J.; Fdez-Sevilla, D.; Stach, A.; Jato, V. Assessment between pollen seasons in areas with different urbanization level related to local vegetation sources and differences in allergen exposure. Aerobiologia 2010, 26, 1–14. [Google Scholar] [CrossRef]
- Aguilera, F.; Dhiab, A.B.; Msallem, M.; Orlandi, F.; Bonofiglio, T.; Ruiz-Valenzuela, L.; Galan, C.; Diaz-de la Guardia, C.; Gianelli, A.; et al. Airborne-pollen maps for olive-growing areas throughout the Mediterranean region: Spatio-temporal interpretation. Aerobiologia 2015, 31, 421–434. [Google Scholar] [CrossRef]
- Voukantsis, D.; Niska, H.; Karatzas, K.; Riga, M.; Damialis, A.; Vokou, D. Forecasting daily pollen concentrations using data-driven modeling methods in Thessaloniki, Greece. Atmos. Environ. 2010, 44, 5101–5111. [Google Scholar] [CrossRef]
- Fernández-Rodríguez, S.; Skjøth, C.A.; Tormo-Molina, R.; Brandao, R.; Caeiro, E.; Silva-Palacios, I. Identification of potential sources of airborne Olea pollen in the Southwest Iberian Peninsula. Int. J. Biometeorol. 2014, 58, 337–348. [Google Scholar] [CrossRef] [PubMed]
- Sofiev, M.; Siljamo, P.; Ranta, H.; Linkosalo, T.; Jaeger, S.; Rasmussen, A.; Rantio-Lehtimaki, A.; Severova, E.; Kukkonen, J. A numerical model of birch pollen emission and dispersion in the atmosphere. Description of the emission module. Int. J. Biometeorol. 2013, 57, 45–58. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Ceballos, M.A.; Skjøth, C.A.; García-Mozo, H.; Bolívar, J.P.; Galán, C. Improvement in the accuracy of back trajectories using WRF to identify pollen sources in southern Iberian Peninsula. Int. J. Biometeorol. 2014, 58, 2031–2043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oteros, J.; García-Mozo, H.; Alcázar, P.; Belmonte, J.; Bermejo, D.; Boi, M. A new method for determining the sources of airborne particles. J. Environ. Manag. 2015, 155, 212–218. [Google Scholar] [CrossRef] [PubMed]
- Oteros, J.; Valencia, R.M.; Del Río, S.; Vega, A.M.; García-Mozo, H.; Galán, C.; Gutiérrez, P.; Mandrioli, P.; Fernandez-Gonzalez, D. Concentric Ring Method for generating pollen maps. Quercus as case study. Sci. Total Environ. 2017, 576, 637–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rojo, J.; Orlandi, F.; Pérez-Badia, R.; Aguilera, F.; Dhiab, A.B.; Bouziane, H.; Diaz-de la Guardia, C.; Galan, C.; Gutiérrez-Bustillo, A.M.; Moreno-Grau, S.; et al. Modeling olive pollen intensity in the Mediterranean region through analysis of emission sources. Sci. Total Environ. 2016, 551, 73–82. [Google Scholar] [CrossRef]
- Haworth, J.; Cheng, T. Non-parametric regression for space–time forecasting under missing data. Comput. Environ. Urban Syst. 2012, 36, 538–550. [Google Scholar] [CrossRef]
- Donnelly, A.; Misstear, B.; Broderick, B. Real time air quality forecasting using integrated parametric and non-parametric regression techniques. Atmos. Environ. 2015, 103, 53–65. [Google Scholar] [CrossRef] [Green Version]
- Petit, J.E.; Favez, O.; Albinet, A.; Canonaco, F. A user-friendly tool for comprehensive evaluation of the geographical origins of atmospheric pollution: Wind and trajectory analyses. Environ. Model. Softw. 2017, 88, 183–187. [Google Scholar] [CrossRef]
- Petit, J.E.; Favez, O.; Sciare, J.; Crenn, V.; Sarda-Estève, R.; Bonnaire, N.; Mocnik, G.; Dupont, J.C.; Haeffelin, M.; Leoz-Garziandia, E. Two years of near real-time chemical composition of submicron aerosols in the region of Paris using an Aerosol Chemical Speciation Monitor (ACSM) and a multi-wavelength Aethalometer. Atmos. Chem. Phys. 2015, 15, 2985–3005. [Google Scholar] [CrossRef]
- Mason, R.H.; Si, M.; Chou, C.; Irish, V.E.; Dickie, R.; Elizondo, P.; Wong, R.; Brintnell, M.; Elsasser, M.; Lassar, M.; et al. Size-resolved measurements of ice-nucleating particles at six locations in North America and one in Europe. Atmos. Chem. Phys. 2016, 16, 1637–1651. [Google Scholar] [CrossRef]
- Hirst, J. An automatic volumetric spore trap. Ann. Appl. Boil. 1952, 39, 257–265. [Google Scholar] [CrossRef]
- Jager, S.; Mandroli, P.; Spieksma, F.; Emberlin, J.; Hjelmroos, M.; Rantio-Lehtimaki, A. News. Aerobiologia 1995, 11, 69–70. [Google Scholar]
- Volumetric Pollen and Particle Sampler, VPPS 2000, User’s Manual. 1980, pp. 1–19. Available online: https://www.lanzoni.it/campionatore-pollini (accessed on 30 November 2018).
- Galán, C.; Smith, M.; Thibaudon, M.; Frenguelli, G.; Oteros, J.; Gehrig, R.; Berger, U.; Clot, B.; Brandao, R. Pollen monitoring: Minimum requirements and reproducibility of analysis. Aerobiologia 2014, 30, 385–395. [Google Scholar]
- Caulton, E.; Lacey, M. Airborne Pollens and Spores: A Guide to Trapping and Counting; British Aerobiology Federation: Harpenden, UK, 1995; ISBN 0-9525617-0-0. [Google Scholar]
- Rogers, C.; Muilenberg, M. Comprehensive Guidelines for the Operation of Hirst-Type Suction Bioaerosol Samplers. Pan-American Aerobiology Association, Standardized Protocols. 2001. Available online: http://www.paaa.org/ (accessed on 30 November 2018).
- Henry, R.; Norris, G.A.; Vedantham, R.; Turner, J.R. Source region identification using kernel smoothing. Environ. Sci. Technol. 2009, 43, 4090–4097. [Google Scholar] [CrossRef] [PubMed]
- Olson, D.A.; Vedantham, R.; Norris, G.A.; Brown, S.G.; Roberts, P. Determining source impacts near roadways using wind regression and organic source markers. Atmos. Environ. 2012, 47, 261–268. [Google Scholar] [CrossRef]
- Yamartino, R.J. A comparison of several “single-pass” estimators of the standard deviation of wind direction. J. Clim. Appl. Meteorol. 1984, 23, 1362–1366. [Google Scholar] [CrossRef]
- Weryszko-Chmielewska, E.; Piotrowska, K. Airborne pollen calendar of Lublin, Poland. Ann. Agric. Environ. Med. 2004, 11, 91–97. [Google Scholar] [PubMed]
- Recio, M.; Picornell, A.; Trigo, M.M.; Gharbi, D.; García-Sánchez, J.; Cabezudo, B. Intensity and temporality of airborne Quercus pollen in the southwest Mediterranean area: Correlation with meteorological and phenoclimatic variables, trends and possible adaptation to climate change. Agric. For. Meteorol. 2018, 250, 308–318. [Google Scholar] [CrossRef]
- Spieksma, F.T.M.; Emberlin, J.C.; Hjelmroos, M.; Jäger, S.; Leuschner, R.M. Atmospheric birch (Betula) pollen in Europe: Trends and fluctuations in annual quantities and the starting dates of the seasons. Grana 1995, 34, 51–57. [Google Scholar] [CrossRef]
- García-Mozo, H.; Mestre, A.; Galán, C. Phenological trends in southern Spain: A response to climate change. Agric. For. Meteorol. 2010, 150, 575–580. [Google Scholar] [CrossRef]
- de la Cruz, D.R.; Sánchez-Reyes, E.; Sánchez-Sánchez, J. A contribution to the knowledge of Cupressaceae airborne pollen in the middle west of Spain. Aerobiologia 2015, 31, 435–444. [Google Scholar] [CrossRef]
- Hoebeke, L.; Bruffaerts, N.; Verstraeten, C.; Delcloo, A.; De Smedt, T.; Packeu, A.; Detandt, M.; Hendrickx, M. Thirty-four years of pollen monitoring: An evaluation of the temporal variation of pollen seasons in Belgium. Aerobiologia 2018, 34, 139–155. [Google Scholar] [CrossRef]
- Jato, V.; Rodríguez-Rajo, F.J.; Alcázar, P.; De Nuntiis, P.; Galán, C.; Mandrioli, P. May the definition of pollen season influence aerobiological results? Aerobiologia 2006, 22, 13. [Google Scholar] [CrossRef]
- Alexandre, F. Géographie et écologie végétale. Pour une nouvelle convergence. Ph.D. Thesis, Université Paris 7-Diderot, Paris, France, 2008. [Google Scholar]
- Jones, A.M.; Harrison, R.M. The effects of meteorological factors on atmospheric bioaerosol concentrations—A review. Sci. Total Environ. 2004, 326, 151–180. [Google Scholar] [CrossRef] [PubMed]
- D’Amato, G.; Annesi-Maesano, I.; Cecchi, L.; D’Amato, M. Latest News on relationship between thunderstorms and respiratory allergy, severe asthma, and deaths for asthma. Allergy 2018. [Google Scholar] [CrossRef] [PubMed]
- Davies, J.M.; Thien, F.; Hew, M. Thunderstorm asthma: Controlling (deadly) grass pollen allergy. BMJ 2018, 360, 432. [Google Scholar] [CrossRef]
- İnceoğlu, Ö.; Pinar, N.M.; Şakiyan, N.; Sorkun, K. Airborne pollen concentration in Ankara, Turkey 1990–1993. Grana 1994, 33, 158–161. [Google Scholar] [CrossRef] [Green Version]
- Emberlin, J.; Detandt, M.; Gehrig, R.; Jaeger, S.; Nolard, N.; Rantio-Lehtimäki, A. Responses in the start of Betula (birch) pollen seasons to recent changes in spring temperatures across Europe. Int. J. Biometeorol. 2002, 46, 159–170. [Google Scholar]
- Stach, A.; Emberlin, J.; Smith, M.; Adams-Groom, B.; Myszkowska, D. Factors that determine the severity of Betula spp. pollen seasons in Poland (Poznań and Krakow) and the United Kingdom (Worcester and London). Int. J. Biometeorol. 2008, 52, 311–321. [Google Scholar] [CrossRef]
- Galán, C.; Alcázar, P.; Oteros, J.; García-Mozo, H.; Aira, M.J.; Belmonte, J.; Diaz de la Guardia, C.; Fernández-González, D.; Gutierrez-Bustillo, M.; Moreno-Grau, S.; et al. Airborne pollen trends in the Iberian Peninsula. Sci. Total Environ. 2016, 550, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Ritenberga, O.; Sofiev, M.; Siljamo, P.; Saarto, A.; Dahl, A.; Ekebom, A.; Sauliene, I.; Shalaboda, V.; Severova, E.; Hoebeke, L.; et al. A statistical model for predicting the inter-annual variability of birch pollen abundance in Northern and North-Eastern Europe. Sci. Total Environ. 2018, 615, 228–239. [Google Scholar] [CrossRef] [PubMed]
- Laaidi, K. Predicting days of high allergenic risk during Betula pollination using weather types. Int. J. Biometeorol. 2001, 45, 124–132. [Google Scholar] [CrossRef] [PubMed]
- D’Odorico, P.; Yoo, J.C.; Jaeger, S. Changing seasons: An effect of the North Atlantic Oscillation? J. Clim. 2002, 15, 435–445. [Google Scholar] [CrossRef]
- Dowding, P. Wind pollination mechanisms and aerobiology. Int. Rev. Cytol. 1987, 107, 421–437. [Google Scholar]
- Hjelmroos, M. Evidence of long-distance transport of Betula pollen. Grana 1991, 30, 215–228. [Google Scholar] [CrossRef]
- Cecchi, L.; Torrigiani Malaspina, T.T.; Albertini, R.; Zanca, M.; Ridolo, E.; Usberti, I.; Morabito, M.; Dall’ Aglio, P.; Orlandini, S. The contribution of long-distance transport to the presence of Ambrosia pollen in central northern Italy. Aerobiologia 2007, 23, 145–151. [Google Scholar] [CrossRef]
- Belmonte, J.; Alarcón, M.; Avila, A.; Scialabba, E.; Pino, D. Long-range transport of beech (Fagus sylvatica L.) pollen to Catalonia (north-eastern Spain). Int. J. Biometeorol. 2008, 52, 675–687. [Google Scholar] [CrossRef] [PubMed]
- Bessagnet, B.; Hodzic, A.; Blanchard, O.; Lattuati, M.; Le Bihan, O.; Marfaing, H.; Rouil, L. Origin of particulate matter pollution episodes in wintertime over the Paris Basin. Atmos. Environ. 2005, 39, 6159–6174. [Google Scholar] [CrossRef]
- Baudic, A.; Gros, V.; Sauvage, S.; Locoge, N.; Sanchez, O.; Sarda-Estève, R.; Kalogridis, C.; Petit, J.E.; Bonnaire, N.; Baisnée, D.; et al. Seasonal variability and source apportionment of volatile organic compounds (VOCs) in the Paris megacity (France). Atmos. Chem. Phys. 2016, 16, 11961–11989. [Google Scholar] [CrossRef]
- Agarwal, M.K.; Swanson, M.C.; Reed, C.E.; Yunginger, J.W. Airborne ragweed allergens: Association with various particle sizes and short ragweed plant parts. J. Allergy Clin. Immunol. 1984, 74, 687–693. [Google Scholar] [CrossRef]
- Spieksma, F.T.M.; Kramps, J.A.; Van der Linden, A.C.; Nikkels, B.H.; Plomp, A.; Koerten, H.K.; Dijkman, J.H. Evidence of grass-pollen allergenic activity in the smaller micronic atmospheric aerosol fraction. Clin. Exp. Allergy 1990, 20, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Taylor, P.E.; Flagan, R.C.; Miguel, A.G.; Valenta, R.; Glovsky, M.M. Birch pollen rupture and the release of aerosols of respirable allergens. Clin. Exp. Allergy 2004, 34, 1591–1596. [Google Scholar] [CrossRef] [PubMed]














| Species | Families |
|---|---|
| Alnus | Betulaceae |
| Betula | Betulaceae |
| Carpinus | Betulaceae |
| Corylus | Betulaceae |
| Juniperus | Cupressaceae |
| Cupressus | Cupressaceae |
| Fraxinus | Oleaceae |
| Species | Families |
|---|---|
| All | Poaceae |
| Parietaria | Urticaceae |
| Integrated Period (IP) | Total Rain (mm) | APS (2015 to 2018) (Nb#/m3) |
|---|---|---|
| August 2014 to January 2015 | 291.5 | 54,931 |
| August 2015 to January 2016 | 327.5 | 68,958 |
| August 2016 to January 2017 | 207.2 | 40,861 |
| August 2017 to January 2018 | 465.2 | 86,989 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sarda Estève, R.; Baisnée, D.; Guinot, B.; Petit, J.-E.; Sodeau, J.; O’Connor, D.; Besancenot, J.-P.; Thibaudon, M.; Gros, V. Temporal Variability and Geographical Origins of Airborne Pollen Grains Concentrations from 2015 to 2018 at Saclay, France. Remote Sens. 2018, 10, 1932. https://doi.org/10.3390/rs10121932
Sarda Estève R, Baisnée D, Guinot B, Petit J-E, Sodeau J, O’Connor D, Besancenot J-P, Thibaudon M, Gros V. Temporal Variability and Geographical Origins of Airborne Pollen Grains Concentrations from 2015 to 2018 at Saclay, France. Remote Sensing. 2018; 10(12):1932. https://doi.org/10.3390/rs10121932
Chicago/Turabian StyleSarda Estève, Roland, Dominique Baisnée, Benjamin Guinot, Jean-Eudes Petit, John Sodeau, David O’Connor, Jean-Pierre Besancenot, Michel Thibaudon, and Valérie Gros. 2018. "Temporal Variability and Geographical Origins of Airborne Pollen Grains Concentrations from 2015 to 2018 at Saclay, France" Remote Sensing 10, no. 12: 1932. https://doi.org/10.3390/rs10121932
APA StyleSarda Estève, R., Baisnée, D., Guinot, B., Petit, J. -E., Sodeau, J., O’Connor, D., Besancenot, J. -P., Thibaudon, M., & Gros, V. (2018). Temporal Variability and Geographical Origins of Airborne Pollen Grains Concentrations from 2015 to 2018 at Saclay, France. Remote Sensing, 10(12), 1932. https://doi.org/10.3390/rs10121932