Remote Estimation of Nitrogen Vertical Distribution by Consideration of Maize Geometry Characteristics



Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site
2.2. Experimental Design
2.3. Hyperspectral Reflectance Measurement
2.4. Plant Vertical Layered Sampling and Measurements
2.5. Hyperspectral Vegetation Indices and Data Analysis
3. Results
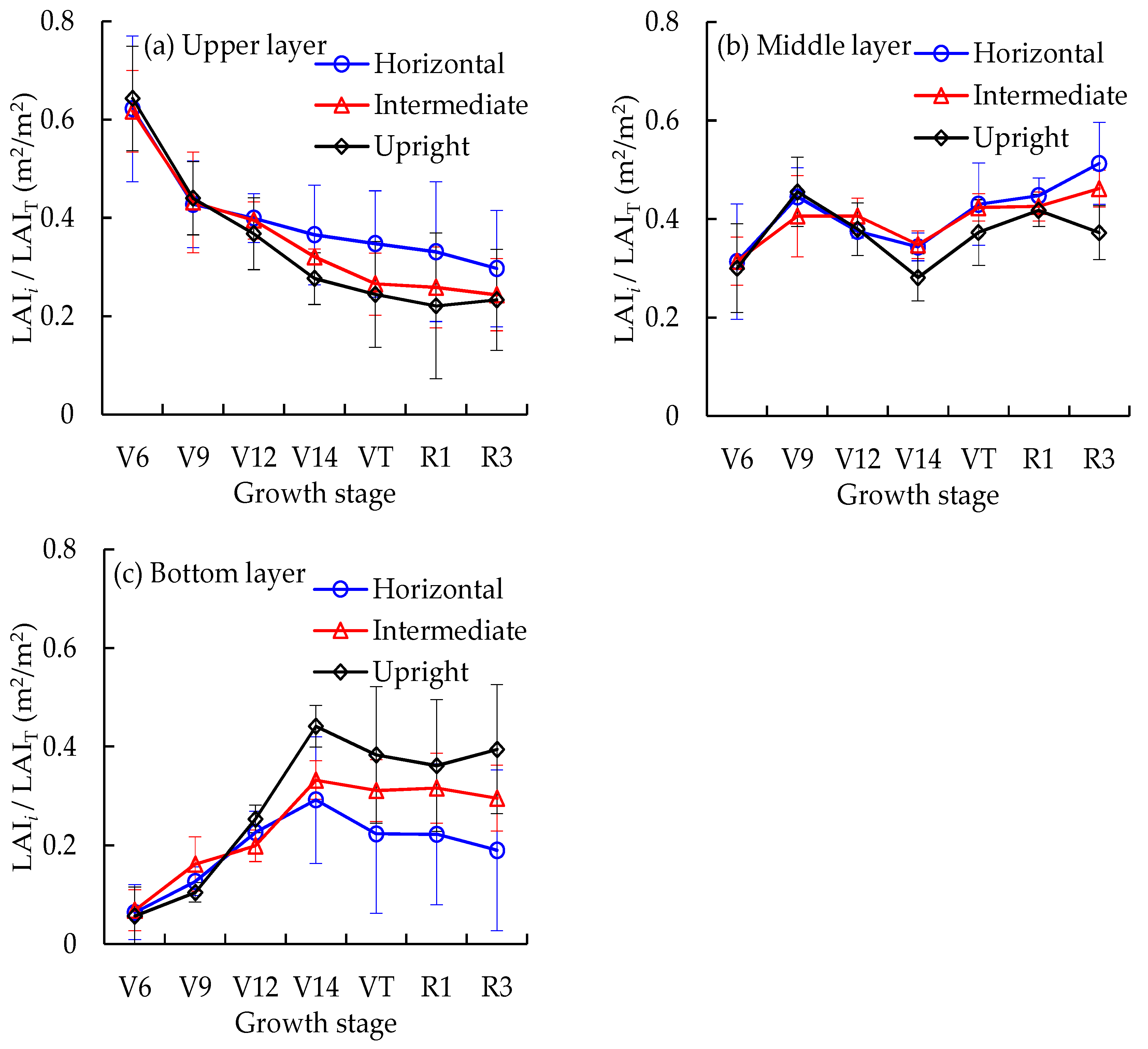
3.1. Canopy Structure Characteristics
3.2. Vertical Distribution Characteristics of Leaf N Density
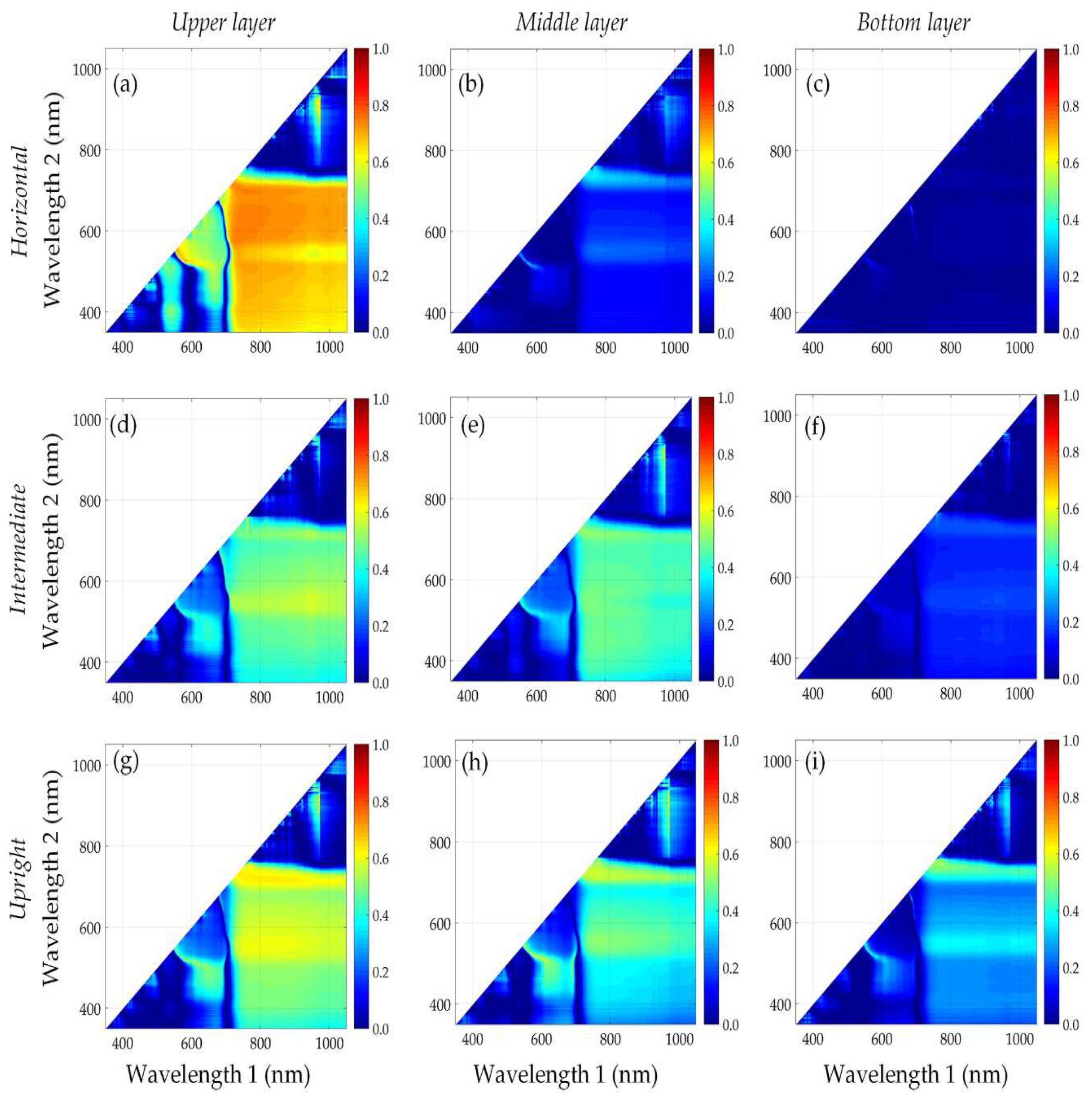
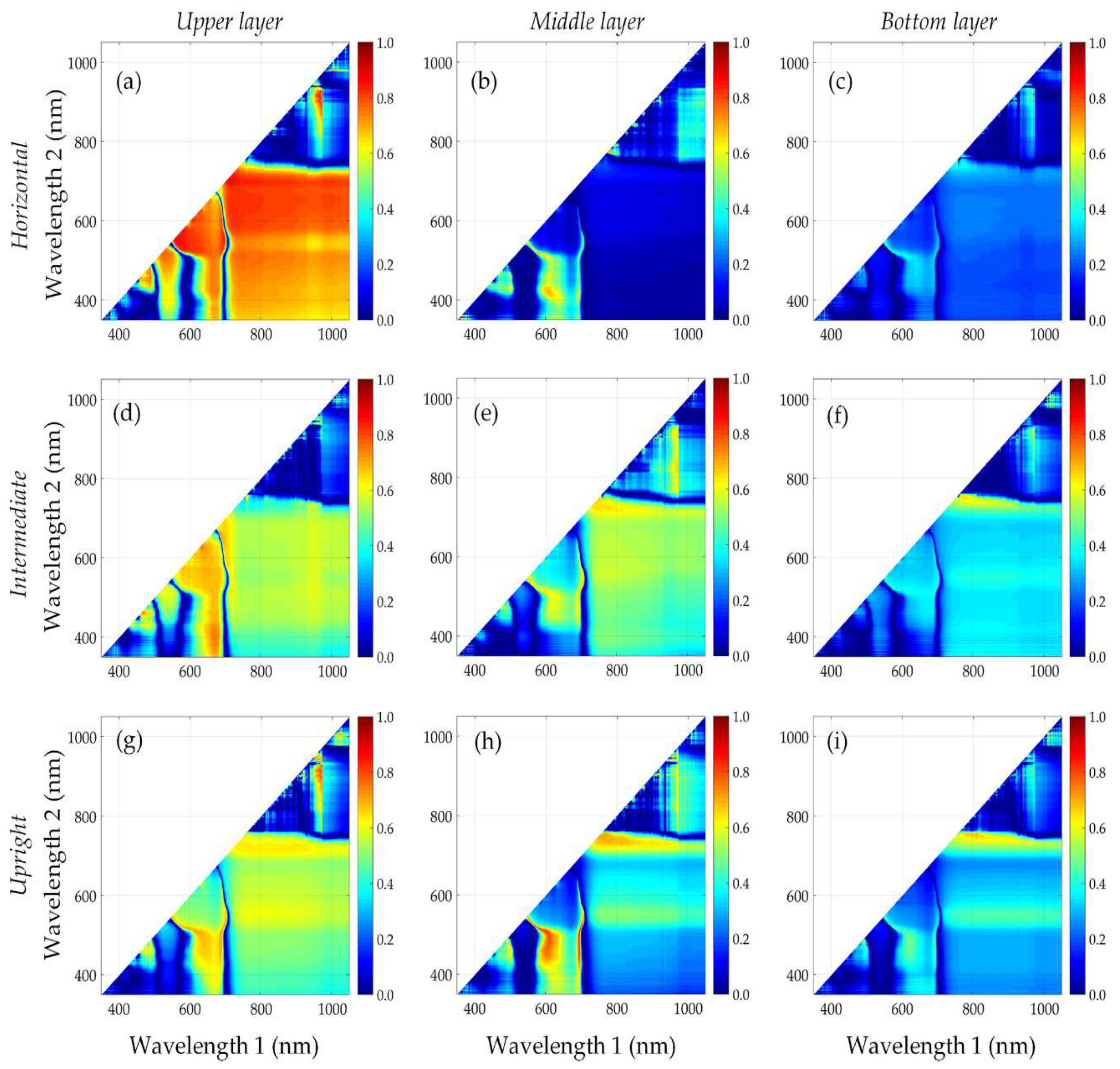
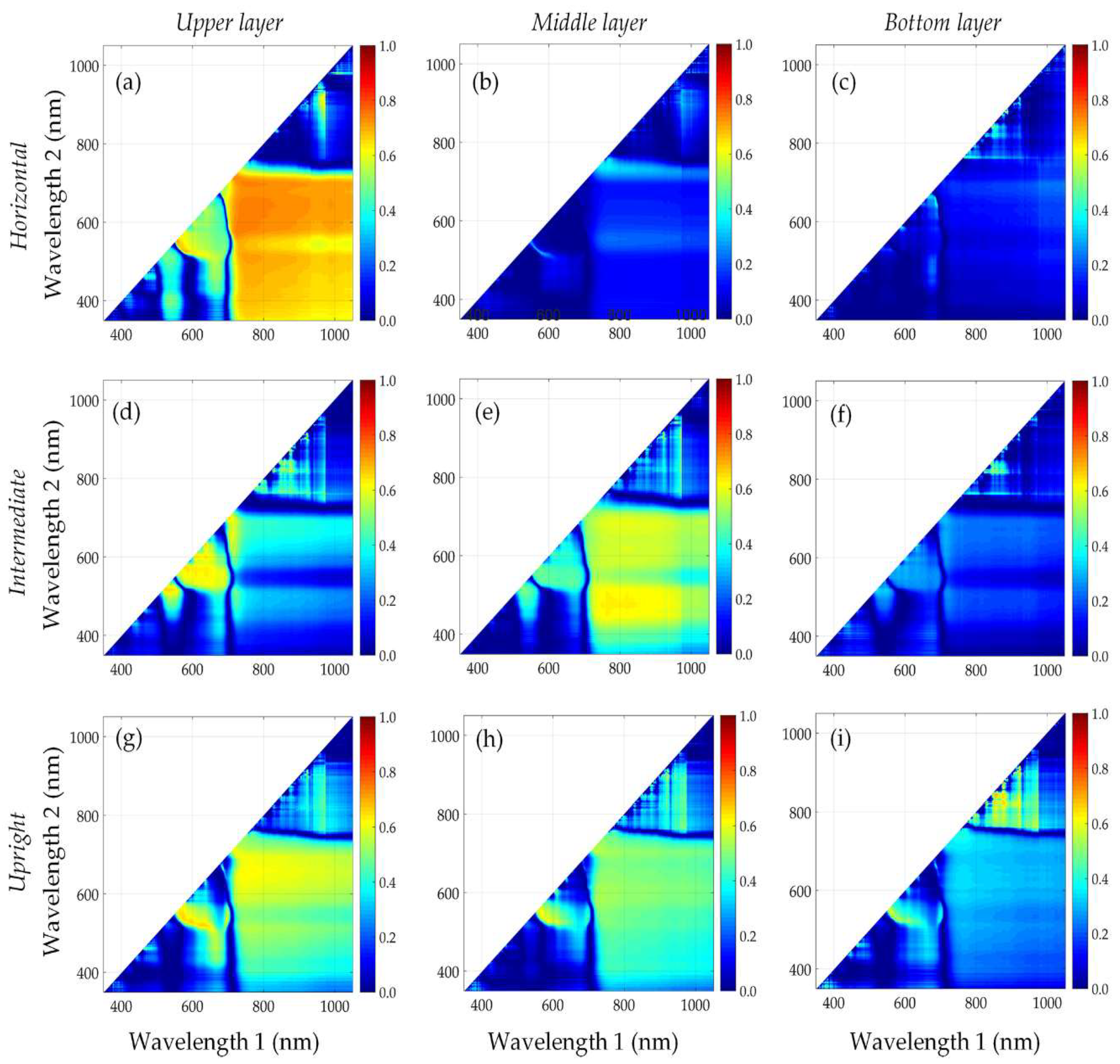
3.3. Performance of Published Vegetation Indices
3.4. Identification of New Vegetation Indices
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sheehy, J.E.; Dionora, M.J.A.; Mitchell, P.L.; Peng, S.; Cassman, K.G.; Lemaire, G.; Williams, R.L. Critical nitrogen concentrations: Implications for high-yielding rice (Oryza sativa L.) cultivars in the tropics. Field Crop Res. 1998, 59, 31–41. [Google Scholar] [CrossRef]
- Ziadi, N.; Bélanger, G.; Claessens, A.; Lefebvre, L.; Cambouris, A.N.; Tremblay, N.; Nolin, M.C.; Parent, L.É. Determination of a critical nitrogen dilution curve for spring wheat. Agron. J. 2010, 102, 241–250. [Google Scholar] [CrossRef]
- Yue, S.C.; Sun, F.L.; Meng, Q.F.; Zhao, R.F.; Li, F.; Chen, X.P.; Zhang, F.S.; Cui, Z.L. Validation of a critical nitrogen curve for summer maize in the North China Plain. Pedosphere 2014, 24, 76–83. [Google Scholar] [CrossRef]
- Scholberg, J.; McNeal, B.L.; Boote, K.J.; Jones, J.W.; Locascio, S.J.; Olson, S.M. Nitrogen stress effects on growth and nitrogen accumulation by field-grown tomato. Agron. J. 2000, 92, 159–167. [Google Scholar] [CrossRef]
- Zhao, D.; Reddy, K.R.; Kakani, V.G.; Reddy, V.R. Nitrogen deficiency effects on plant growth, leaf photosynthesis, and hyperspectral reflectance properties of sorghum. Eur. J. Agron. 2005, 22, 391–403. [Google Scholar] [CrossRef]
- Boussadia, O.; Steppe, K.; Zgallai, H.; El Hadj, S.B.; Braham, M.; Lemeur, R.; Van Labeke, M.C. Effects of nitrogen deficiency on leaf photosynthesis, carbohydrate status and biomass production in two olive cultivars ‘Meski’and ‘Koroneiki’. Sci. Hortic. 2010, 123, 336–342. [Google Scholar] [CrossRef]
- Zhang, Y.; Tan, L.; Zhu, Z.; Yuan, L.; Xie, D.; Sun, C. TOND1 confers tolerance to nitrogen deficiency in rice. Plant J. 2015, 81, 367–376. [Google Scholar] [CrossRef] [Green Version]
- Ju, X.T.; Xing, G.X.; Chen, X.P.; Zhang, S.L.; Zhang, L.J.; Liu, X.J.; Cui, Z.L.; Yin, B.; Christie, P.; Zhu, Z.L.; et al. Reducing environmental risk by improving N management in intensive Chinese agricultural systems. Proc. Natl. Acad. Sci. USA 2009, 106, 3041–3046. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.; Yang, Q.; Pu, R.; Yang, S. Estimation of Nitrogen Vertical Distribution by Bi-Directional Canopy Reflectance in Winter Wheat. Sensors 2014, 14, 20347–20359. [Google Scholar] [CrossRef] [Green Version]
- Qiu, S.J.; He, P.; Zhao, S.C.; Li, W.J.; Xie, J.G.; Hou, Y.P.; Grant, C.A.; Zhou, W.; Jin, J.Y. Impact of Nitrogen Rate on Maize Yield and Nitrogen Use Efficiencies in Northeast China. Agron. J. 2015, 107, 305–313. [Google Scholar] [CrossRef]
- Li, F.; Miao, Y.; Hennig, S.D.; Gnyp, M.L.; Chen, X.; Jia, L.; Bareth, G. Evaluating hyperspectral vegetation indices for estimating nitrogen concentration of winter wheat at different growth stages. Precis. Agric. 2010, 11, 335–357. [Google Scholar] [CrossRef]
- Hansen, P.M.; Schjoerring, J.K. Reflectance measurement of canopy biomass and nitrogen status in wheat crops using normalized difference vegetation indices and partial least square regression. Remote Sens. Environ. 2003, 86, 542–553. [Google Scholar] [CrossRef]
- Xue, L.; Cao, W.; Luo, W.; Dai, T.; Zhu, Y. Monitoring leaf nitrogen status in rice with canopy spectral reflectance. Agron. J. 2004, 96, 135–142. [Google Scholar] [CrossRef]
- Yu, K.; Li, F.; Gnyp, M.L.; Miao, Y.; Bareth, G.; Chen, X. Remotely detecting canopy nitrogen concentration and uptake of paddy rice in the Northeast China Plain. ISPRS-J. Photogramm. Remote Sens. 2013, 78, 102–115. [Google Scholar] [CrossRef]
- Huang, S.; Miao, Y.; Zhao, G.; Yuan, F.; Ma, X.; Tan, C.; Yu, W.; Gnyp, M.L.; Lenz-Wiedemann, V.I.S.; Rascher, U.; et al. Satellite remote sensing-based in-season diagnosis of rice nitrogen status in Northeast China. Remote Sens. 2015, 7, 10646–10667. [Google Scholar] [CrossRef]
- He, L.; Song, X.; Feng, W.; Guo, B.B.; Zhang, Y.S.; Wang, Y.H.; Wang, C.Y.; Guo, T.C. Improved remote sensing of leaf nitrogen concentration in winter wheat using multi-angular hyperspectral data. Remote Sens. Environ. 2016, 174, 122–133. [Google Scholar] [CrossRef]
- Dreccer, M.F.; Slafer, G.A.; Rabbinge, R. Optimization of vertical distribution of canopy nitrogen. J. Crop Prod. 1997, 1, 47–77. [Google Scholar] [CrossRef]
- Dreccer, M.F.; Van Oijen, M.; Schapendonk, A.H.C.M.; Pot, C.S.; Rabbinge, R. Dynamics of vertical leaf nitrogen distribution in a vegetative wheat canopy. Impact on canopy photosynthesis. Ann. Bot. 2000, 86, 821–831. [Google Scholar] [CrossRef]
- Huang, W.; Wang, Z.; Huang, L.; Lamb, D.W.; Ma, Z.; Zhang, J.; Wang, J.; Zhao, C. Estimation of vertical distribution of chlorophyll concentration by bi-directional canopy reflectance spectra in winter wheat. Precis. Agric. 2011, 12, 165–178. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, J.; Zhao, C.; Zhao, M.; Huang, W.; Wang, C. Vertical distribution of nitrogen in different layers of leaf and stem and their relationship with grain quality of winter wheat. J. Plant Nutr. 2012, 28, 73–91. [Google Scholar] [CrossRef]
- Li, H.; Zhao, C.; Huang, W.; Yang, G. Non-uniform vertical nitrogen distribution within plant canopy and its estimation by remote sensing: A review. Field Crop Res. 2013, 142, 75–84. [Google Scholar] [CrossRef]
- Lu, J.L. Plant Nutrition, 1st ed.; the First Volume; Beijing Agricultural University Press (continuing China Agricultural University Press): Beijing, China, 1994. (In Chinese) [Google Scholar]
- Wang, Z.; Wang, J.; Liu, L.; Huang, W.; Zhao, C.; Lu, Y. Estimation of nitrogen status in middle and bottom layers of winter wheat canopy by using ground-measured canopy reflectance. Commun. Soil Sci. Plant 2005, 36, 2289–2302. [Google Scholar] [CrossRef]
- Wang, J.H.; Huang, W.J.; Lao, C.L.; Zhang, L.D.; Luo, C.B.; Wang, T.; Liu, L.Y.; Song, X.Y.; Ma, Z.H. Inversion of winter wheat foliage vertical distribution based on canopy reflected spectrum by partial least squares regression method. Spectrosc. Spect. Anal. 2007, 27, 1319–1322, (In Chinese with English abstract). [Google Scholar]
- Ciganda, V.S.; Gitelson, A.A.; Schepers, J. How deep does a remote sensor sense? expression of chlorophyll content in a maize canopy. Remote Sens. Environ. 2012, 126, 240–247. [Google Scholar] [CrossRef]
- Hirose, T.; Werger, M.J.A.; Pons, T.L.; van Rheenen, J.W.A. Canopy structure and leaf nitrogen distribution in a stand of Lysimachia vulgaris L. as influenced by stand density. Oecologia 1988, 77, 145–150. [Google Scholar] [CrossRef]
- Anten, N.P.R.; Schieving, F.; Werger, M.J.A. Patterns of light and nitrogen distribution in relation to whole canopy carbon gain in C3 and C4 mono- and dicotyledonous species. Oecologia 1995, 101, 504–513. [Google Scholar] [CrossRef] [PubMed]
- Lötscher, M.; Stroh, K.; Schnyder, H. Vertical leaf nitrogen distribution in relation to nitrogen status in grassland plants. Ann. Bot. 2003, 92, 679–688. [Google Scholar] [CrossRef]
- Bertheloot, J.; Martre, P.; Andrieu, B. Dynamics of light and nitrogen distribution during grain filling within wheat canopy. Plant Physiol. 2008, 148, 1707–1720. [Google Scholar] [CrossRef]
- Monsi, M.; Saeki, T. On the factor light in plant communities and its importance for matter production. Ann. Bot. 2005, 95, 549–567. [Google Scholar] [CrossRef]
- Hirose, T.; Werger, M.J.A. Maximizing daily canopy photosynthesis with respect to the leaf nitrogen allocation pattern in the canopy. Oecologia 1987, 72, 520–526. [Google Scholar] [CrossRef]
- Maddonni, G.A.; Otegui, M.E.; Cirilo, A.G. Plant population density, row spacing and hybrid effects on maize canopy architecture and light attenuation. Field Crop. Res. 2001, 71, 183–193. [Google Scholar] [CrossRef]
- Lambert, R.J.; Johnson, R.R. Leaf angle, tassel morphology, and the performance of maize hybrids. Crop Sci. 1978, 18, 499–502. [Google Scholar] [CrossRef]
- Azumi, Y.; Watanabe, A. Evidence for a senescence-associated gene induced by darkness. Plant Physiol. 1991, 95, 577–583. [Google Scholar] [CrossRef] [PubMed]
- Ku, L.X.; Zhao, W.M.; Zhang, J.; Wu, L.C.; Wang, C.L.; Wang, P.A.; Zhang, W.Q.; Chen, Y.H. Quantitative trait loci mapping of leaf angle and leaf orientation value in maize (Zea mays L.). Theor. Appl. Genet. 2010, 121, 951–959. [Google Scholar] [CrossRef] [PubMed]
- Duvick, D.N.; Smith, J.S.C.; Cooper, M. Long-term selection in a commercial hybrid maize breeding program. Plant Breed. Rev. 2004, 24, 109–151. [Google Scholar]
- Ma, D.L.; Xie, R.Z.; Niu, X.K.; Li, S.K.; Long, H.L.; Liu, Y.E. Changes in the morphological traits of maize genotypes in China between the 1950s and 2000s. Eur. J. Agron. 2014, 58, 1–10. [Google Scholar] [CrossRef]
- Tong, P. The course and the achievement of corn variety improvement in the20th century in China. China Hist. Mater. Sci. Technol. 2001, 22, 113–127. [Google Scholar]
- Meghji, M.R.; Dudley, J.W.; Lambert, R.J.; Sprague, G.F. Inbreeding depression, inbred and hybrid grain yields, and other traits of maize genotypes representing three eras. Crop Sci. 1984, 24, 545–549. [Google Scholar] [CrossRef]
- Russell, W.A. Genetic improvement of maize yields. Adv. Agron. 1991, 46, 245–298. [Google Scholar]
- IUSS Working Group WRB. World Reference Base for Soil Resources 2006; World Soil Resources Reports No. 103; FAO: Rome, Italy, 2006. [Google Scholar]
- Soil Survey Division Staff. Soil Survey Manual; United States Department of Agriculture: Washington, DC, USA, 1993.
- Pepper, G.E.; Pearce, R.B.; Mock, J.J. Leaf orientation and yield of maize. Crop Sci. 1977, 17, 883–886. [Google Scholar] [CrossRef]
- Jonckheere, I.; Fleck, S.; Nackaerts, K.; Muys, B.; Coppin, P.; Weiss, M.; Baret, F. Methods for leaf area index determination. Part I: Theories, techniques and instruments. Agric. For. Meteorol. 2004, 121, 19–35. [Google Scholar] [CrossRef]
- Helrich, K. Official Methods of Analysis of the AOAC, 15th ed.; AOAC: Arlington, VA, USA, 1990; pp. 16–17. [Google Scholar]
- Greenwood, D.J.; Neeteson, J.J.; Draycott, A. Quantitative relationships for the dependence of growth rate of arable crops on their nitrogen content, dry weight and aerial environment. In Fundamental, Ecological and Agricultural Aspects of Nitrogen Metabolism in Higher Plants; Springer: Dordrecht, The Netherlands, 1986; Volume 91, pp. 281–301. [Google Scholar]
- Giletto, C.M.; Echeverría, H.E. Critical nitrogen dilution curve for processing potato in Argentinean humid pampas. Am. J. Potato Res. 2012, 89, 102–110. [Google Scholar] [CrossRef]
- Mistele, B.; Schmidhalter, U. Estimating the nitrogen nutrition index using spectral canopy reflectance measurements. Eur. J. Agron. 2008, 29, 184–190. [Google Scholar] [CrossRef]
- Hirose, T.; Werger, M.J.A.; van Rheenen, J.W.A. Canopy development and leaf nitrogen distribution in a stand of Carex acutiformis. Ecology 1989, 70, 1610–1618. [Google Scholar] [CrossRef]
- Archontoulis, S.V.; Vos, J.; Yin, X.; Bastiaans, L.; Danalatos, N.G.; Struik, P.C. Temporal dynamics of light and nitrogen vertical distributions in canopies of sunflower, kenaf and cynara. Field Crop Res. 2011, 122, 186–198. [Google Scholar] [CrossRef]
- Schieving, F.; Pons, T.L.; Werger, M.J.A.; Hirose, T. The vertical distribution of nitrogen and photosynthetic activity at different plant densities in Carex acutiformis. Plant Soil 1992, 14, 9–17. [Google Scholar] [CrossRef]
- Zarco-Tejada, P.J.; Miller, J.R.; Noland, T.L.; Mohammed, G.H.; Sampson, P.H. Scaling-up and model inversion methods with narrowband optical indices for chlorophyll content estimation in closed forest canopies with hyperspectral data. IEEE Trans. Geosci. Remote Sens. 2001, 39, 1491–1507. [Google Scholar] [CrossRef] [Green Version]
- Blackburn, G.A. Quantifying chlorophylls and carotenoids at leaf and canopy scales: An evaluation of some hyperspectral approaches. Remote Sens. Environ. 1998, 66, 273–285. [Google Scholar] [CrossRef]
- Gitelson, A.; Merzlyak, M.N. Quantitative estimation of chlorophyll-a using reflectance spectra: Experiments with autumn chestnut and maple leaves. J. Photochem. Photobiol. B-Biol. 1994, 22, 247–252. [Google Scholar] [CrossRef]
- Fitzgerald, G.J.; Rodriguez, D.; Christensen, L.K.; Belford, K.; Sadras, V.O.; Clarke, T.R. Spectral and thermal sensing for nitrogen and water status in rainfed and irrigated wheat environments. Precis. Agric. 2006, 7, 233–248. [Google Scholar] [CrossRef]
- Rondeaux, G.; Steven, M.; Baret, F. Optimization of soil-adjusted vegetation indices. Remote Sens. Environ. 1996, 55, 95–107. [Google Scholar] [CrossRef]
- Dash, J.; Curran, P.J. The MERIS terrestrial chlorophyll index. Int. J. Remote Sens. 2004, 25, 5403–5413. [Google Scholar] [CrossRef]
- Feng, W.; Guo, B.B.; Wang, Z.J.; He, L.; Song, X.; Wang, Y.H.; Guo, T.C. Measuring leaf nitrogen concentration in winter wheat using double-peak spectral reflection remote sensing data. Field Crop. Res. 2014, 159, 43–52. [Google Scholar] [CrossRef]
- Sims, D.A.; Gamon, J.A. Relationships between leaf pigment content and spectral reflectance across a wide range of species, leaf structures and developmental stages. Remote Sens. Environ. 2002, 81, 331–354. [Google Scholar] [CrossRef]
- Le Maire, G.; François, C.; Soudani, K.; Berveiller, D.; Pontailler, J.Y.; Bréda, N.; Genet, H.; Davi, H.; Dufrêne, E. Calibration and validation of hyperspectral indices for the estimation of broadleaved forest leaf chlorophyll content, leafmass per area, leaf area index and leaf canopy biomass. Remote Sens. Environ. 2008, 112, 3846–3864. [Google Scholar] [CrossRef]
- Wu, C.; Niu, Z.; Tang, Q.; Huang, W. Estimating chlorophyll content from hyperspectral vegetation indices: Modeling and validation. Agric. For. Meteorol. 2008, 148, 1230–1241. [Google Scholar] [CrossRef]
- Daughtry, C.S.T.; Walthall, C.L.; Kim, M.S.; De Colstoun, E.B.; McMurtrey, J.E. Estimating corn leaf chlorophyll concentration from leaf and canopy reflectance. Remote Sens. Environ. 2000, 74, 229–239. [Google Scholar] [CrossRef]
- Drouet, J.L.; Bonhomme, R. Do variations in local leaf irradiance explain changes to leaf nitrogen within row maize canopies? Ann. Bot. 1999, 84, 61–69. [Google Scholar] [CrossRef]
- Shiratsuchi, H.; Yamagishi, T.; Ishii, R. Leaf nitrogen distribution to maximize the canopy photosynthesis in rice. Field Crop Res. 2006, 95, 291–304. [Google Scholar] [CrossRef]
- Feng, W.; Yao, X.; Zhu, Y.; Tian, Y.C.; Cao, W.X. Monitoring leaf nitrogen status with hyperspectral reflectance in wheat. Eur. J. Agron. 2008, 28, 394–404. [Google Scholar] [CrossRef]
- Ottman, M.J.; Welch, L.F. Supplemental radiation effects on senescence, plant nutrients, and yield of field-grown corn. Agron. J. 1988, 80, 619–626. [Google Scholar] [CrossRef]
- Schlemmer, M.; Gitelson, A.; Schepers, J.; Ferguson, R.; Peng, Y.; Shanahan, J.; Rundquistb, D. Remote estimation of nitrogen and chlorophyll contents in maize at leaf and canopy levels. Int. J. Appl. Earth Obs. Geoinf. 2013, 25, 47–54. [Google Scholar] [CrossRef] [Green Version]
- Feng, W.; Qi, S.; Heng, Y.; Zhou, Y.; Wu, Y.; Liu, W.; He, L.; Li, X. Canopy vegetation indices from in situ hyperspectral data to assess plant water status of winter wheat under powdery mildew stress. Front. Plant Sci. 2017, 8, 1219. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index | Formula | Reference |
|---|---|---|
| Simple ratio (SR) 1 | B810/B560 | [13] |
| SR 2 | B750/B710 | [52] |
| Normalized difference vegetation index (NDVI) | (B800 − B680)/(B800 + B680) | [53] |
| Green NDVI | (B750 − B550)/(B750 + B550) | [54] |
| Normalized difference red edge (NDRE) Index | (B790 − B720)/(B790 + B720) | [55] |
| Optimized soil-adjusted vegetation index (OSAVI) | 1.16 × (B800 − B670)/(B800 + B670 + 0.16) | [56] |
| MERIS * terrestrial chlorophyll index (MTCI) | (B754 − B709)/(B709 − B681) | [57] |
| Double-peak nitrogen index (NDDA) | (B680 + B756 − 2 × B718)/(B756 − B680) | [58] |
| Modified red-edge normalized difference vegetation Index (mND705) | (B750 − B705)/(B750 + B705 − 2 × B445) | [59] |
| New double difference (DDn) Index | 2 × B710 − B660 − B760 | [60] |
| Modified chlorophyll absorption ratio index (MCARI) | (B750 − B705 − 0.2 × (B750 − B550)) × (B750/B705) | [61] |
| MCARI/OSAVI | MCARI/OSAVI | [62] |
| Growth Stage | Canopy Layer | n | Min | Max | Mean | SD 1 | CV 2 (%) |
|---|---|---|---|---|---|---|---|
| Horizontal leaf varieties | |||||||
| All stages | Upper layer | 49 | 1.30 | 3.71 | 2.22 | 0.62 | 28.0 |
| Middle layer | 49 | 0.50 | 2.91 | 1.82 | 0.53 | 29.1 | |
| Bottom layer | 49 | 0.13 | 2.66 | 0.93 | 0.74 | 80.0 | |
| V6 to V12 stages | Upper layer | 21 | 1.41 | 3.71 | 2.37 | 0.74 | 31.2 |
| Middle layer | 21 | 0.50 | 2.44 | 1.59 | 0.56 | 35.4 | |
| Bottom layer | 21 | 0.13 | 1.33 | 0.57 | 0.38 | 66.4 | |
| V14 to R3 stages | Upper layer | 28 | 1.30 | 2.80 | 2.06 | 0.43 | 21.1 |
| Middle layer | 28 | 1.39 | 2.91 | 2.02 | 0.42 | 20.9 | |
| Bottom layer | 28 | 0.17 | 2.66 | 1.18 | 0.84 | 70.6 | |
| Intermediate leaf varieties | |||||||
| All stage | Upper layer | 49 | 0.88 | 3.74 | 2.17 | 0.72 | 33.2 |
| Middle layer | 49 | 0.71 | 3.24 | 2.18 | 0.61 | 27.8 | |
| Bottom layer | 49 | 0.25 | 3.01 | 1.08 | 0.74 | 68.3 | |
| V6 to V12 stages | Upper layer | 21 | 1.33 | 3.74 | 2.59 | 0.74 | 28.7 |
| Middle layer | 21 | 0.71 | 3.24 | 2.02 | 0.61 | 30.1 | |
| Bottom layer | 21 | 0.25 | 1.29 | 0.65 | 0.35 | 53.4 | |
| V14 to R3 stages | Upper layer | 28 | 0.88 | 2.72 | 1.86 | 0.53 | 28.7 |
| Middle layer | 28 | 1.25 | 3.15 | 2.30 | 0.59 | 25.8 | |
| Bottom layer | 28 | 0.37 | 3.01 | 1.40 | 0.79 | 56.8 | |
| Upright leaf varieties | |||||||
| All stage | Upper layer | 49 | 0.81 | 3.68 | 2.03 | 0.71 | 34.8 |
| Middle layer | 49 | 0.93 | 2.96 | 2.09 | 0.53 | 25.3 | |
| Bottom layer | 49 | 0.12 | 3.38 | 1.18 | 0.96 | 81.4 | |
| V6 to V12 stages | Upper layer | 21 | 1.32 | 3.68 | 2.36 | 0.70 | 29.8 |
| Middle layer | 21 | 0.93 | 2.94 | 1.98 | 0.66 | 33.2 | |
| Bottom layer | 21 | 0.12 | 1.91 | 0.64 | 0.48 | 75.0 | |
| V14 to R3 stages | Upper layer | 28 | 0.81 | 3.35 | 1.79 | 0.62 | 34.5 |
| Middle layer | 28 | 1.28 | 2.96 | 2.18 | 0.41 | 18.7 | |
| Bottom layer | 28 | 0.18 | 3.38 | 1.56 | 1.04 | 66.5 | |
| Index | All Stages | V6 to V12 Stages | V14 to R3 Stages | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Upper Layer | Middle Layer | Bottom Layer | Upper Layer | Middle Layer | Bottom Layer | Upper Layer | Middle Layer | Bottom Layer | |
| Horizontal leaf varieties | |||||||||
| SR 1 | 0.63 ** | 0.23 ** | 0.00 | 0.68 ** | 0.06 | 0.20 | 0.41 ** | 0.38 ** | 0.09 |
| SR 2 | 0.67 ** | 0.23 ** | 0.00 | 0.78 ** | 0.14 | 0.27 * | 0.34 ** | 0.27 ** | 0.11 |
| NDVI | 0.75 ** | 0.14 * | 0.01 | 0.83 ** | 0.09 | 0.24 | 0.57 ** | 0.19 * | 0.17 |
| Green NDVI | 0.69 ** | 0.22 ** | 0.02 | 0.75 ** | 0.08 | 0.22 | 0.42 ** | 0.39 ** | 0.08 |
| NDRE | 0.60 ** | 0.30 ** | 0.02 | 0.73 ** | 0.15 | 0.31 | 0.21 * | 0.31 ** | 0.10 |
| OSAVI | 0.75 ** | 0.14 * | 0.01 | 0.83 ** | 0.09 | 0.24 | 0.58 ** | 0.20 * | 0.16 |
| MTCI | 0.23 ** | 0.30 ** | 0.02 | 0.38 * | 0.33 * | 0.30 * | 0.10 | 0.19 * | 0.03 |
| NDDA | 0.25 ** | 0.34 ** | 0.02 | 0.40 * | 0.29 * | 0.31 * | 0.06 | 0.24 * | 0.05 |
| mND705 | 0.69 ** | 0.23 ** | 0.01 | 0.83 ** | 0.19 | 0.30 * | 0.22 * | 0.16 | 0.15 |
| DDn | 0.39 ** | 0.07 | 0.00 | 0.54 ** | 0.02 | 0.31 * | 0.33 ** | 0.36 ** | 0.13 |
| MCARI | 0.46 ** | 0.06 | 0.00 | 0.64 ** | 0.03 | 0.29 * | 0.37 ** | 0.30 ** | 0.15 |
| MCARI/OSAVI | 0.40 ** | 0.05 | 0.01 | 0.58 ** | 0.03 | 0.29 * | 0.33 ** | 0.30 ** | 0.14 |
| Intermediate leaf varieties | |||||||||
| SR 1 | 0.59 ** | 0.50 ** | 0.20 ** | 0.55 ** | 0.50 ** | 0.50 ** | 0.38 ** | 0.51 ** | 0.07 |
| SR 2 | 0.52 ** | 0.54 ** | 0.20 ** | 0.51 ** | 0.60 ** | 0.50 ** | 0.32 ** | 0.50 ** | 0.15 * |
| NDVI | 0.45 ** | 0.46 ** | 0.15 ** | 0.57 ** | 0.53 ** | 0.35 * | 0.19 | 0.60 ** | 0.22 * |
| Green NDVI | 0.55 ** | 0.50 ** | 0.18 ** | 0.52 ** | 0.56 ** | 0.40 * | 0.37 ** | 0.53 ** | 0.11 |
| NDRE | 0.49 ** | 0.49 ** | 0.20 ** | 0.43 * | 0.64 ** | 0.49 ** | 0.20 * | 0.41 ** | 0.05 |
| OSAVI | 0.46 ** | 0.46 ** | 0.15 * | 0.56 ** | 0.53 ** | 0.35 * | 0.21 * | 0.59 ** | 0.22 * |
| MTCI | 0.41 ** | 0.43 ** | 0.18 ** | 0.22 | 0.63 ** | 0.49 ** | 0.30 ** | 0.25 * | 0.03 |
| NDDA | 0.35 ** | 0.36 ** | 0.16 ** | 0.18 | 0.61 ** | 0.43 * | 0.12 | 0.16 | 0.00 |
| mND705 | 0.45 ** | 0.48 ** | 0.17 ** | 0.46 ** | 0.63 ** | 0.37 * | 0.27 ** | 0.41 ** | 0.16 * |
| DDn | 0.18 * | 0.42 ** | 0.13 * | 0.13 | 0.50 ** | 0.47 ** | 0.06 | 0.43 ** | 0.07 |
| MCARI | 0.26 ** | 0.47 ** | 0.13 * | 0.28 | 0.57 ** | 0.54 ** | 0.17 | 0.50 ** | 0.10 |
| MCARI/OSAVI | 0.20 ** | 0.44 ** | 0.12 * | 0.19 | 0.54 ** | 0.52 ** | 0.15 | 0.47 ** | 0.09 |
| Upright leaf varieties | |||||||||
| SR 1 | 0.58 ** | 0.51 ** | 0.44 ** | 0.60 ** | 0.50 ** | 0.54 ** | 0.47 ** | 0.53 ** | 0.34 ** |
| SR 2 | 0.61 ** | 0.56 ** | 0.42 ** | 0.64 ** | 0.57 ** | 0.53 ** | 0.55 ** | 0.54 ** | 0.39 ** |
| NDVI | 0.53 ** | 0.39 ** | 0.24 ** | 0.54 ** | 0.33 * | 0.28 * | 0.59 ** | 0.50 ** | 0.32 ** |
| Green NDVI | 0.59 ** | 0.50 ** | 0.37 ** | 0.59 ** | 0.48 ** | 0.44 ** | 0.50 ** | 0.51 ** | 0.30 ** |
| NDRE | 0.63 ** | 0.56 ** | 0.45 ** | 0.63 ** | 0.64 ** | 0.52 ** | 0.52 ** | 0.50 ** | 0.36 ** |
| OSAVI | 0.53 ** | 0.39 ** | 0.24 ** | 0.54 ** | 0.33 * | 0.27 * | 0.59 ** | 0.49 ** | 0.32 ** |
| MTCI | 0.53 ** | 0.56 ** | 0.48 ** | 0.49 ** | 0.76 ** | 0.61 ** | 0.50 ** | 0.48 ** | 0.39 ** |
| NDDA | 0.52 ** | 0.56 ** | 0.45 ** | 0.48 ** | 0.77 ** | 0.56 ** | 0.44 ** | 0.45 ** | 0.33 ** |
| mND705 | 0.61 ** | 0.58 ** | 0.36 ** | 0.65 ** | 0.59 ** | 0.43 * | 0.58 ** | 0.52 ** | 0.36 ** |
| DDn | 0.43 ** | 0.43 ** | 0.27 ** | 0.56 ** | 0.53 ** | 0.55 ** | 0.35 ** | 0.34 ** | 0.20 * |
| MCARI | 0.47 ** | 0.46 ** | 0.29 ** | 0.61 ** | 0.47 ** | 0.54 ** | 0.42 ** | 0.46 ** | 0.28 ** |
| MCARI/OSAVI | 0.43 ** | 0.45 ** | 0.28 ** | 0.59 ** | 0.50 ** | 0.59 ** | 0.40 ** | 0.42 ** | 0.26 ** |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ye, H.; Huang, W.; Huang, S.; Wu, B.; Dong, Y.; Cui, B. Remote Estimation of Nitrogen Vertical Distribution by Consideration of Maize Geometry Characteristics. Remote Sens. 2018, 10, 1995. https://doi.org/10.3390/rs10121995
Ye H, Huang W, Huang S, Wu B, Dong Y, Cui B. Remote Estimation of Nitrogen Vertical Distribution by Consideration of Maize Geometry Characteristics. Remote Sensing. 2018; 10(12):1995. https://doi.org/10.3390/rs10121995
Chicago/Turabian StyleYe, Huichun, Wenjiang Huang, Shanyu Huang, Bin Wu, Yingying Dong, and Bei Cui. 2018. "Remote Estimation of Nitrogen Vertical Distribution by Consideration of Maize Geometry Characteristics" Remote Sensing 10, no. 12: 1995. https://doi.org/10.3390/rs10121995
APA StyleYe, H., Huang, W., Huang, S., Wu, B., Dong, Y., & Cui, B. (2018). Remote Estimation of Nitrogen Vertical Distribution by Consideration of Maize Geometry Characteristics. Remote Sensing, 10(12), 1995. https://doi.org/10.3390/rs10121995