The Continental Impact of European Forest Conservation Policy and Management on Productivity Stability




Abstract
:
1. Introduction
2. Materials and Methods
2.1. Forest Protected Areas: Treatment Applied
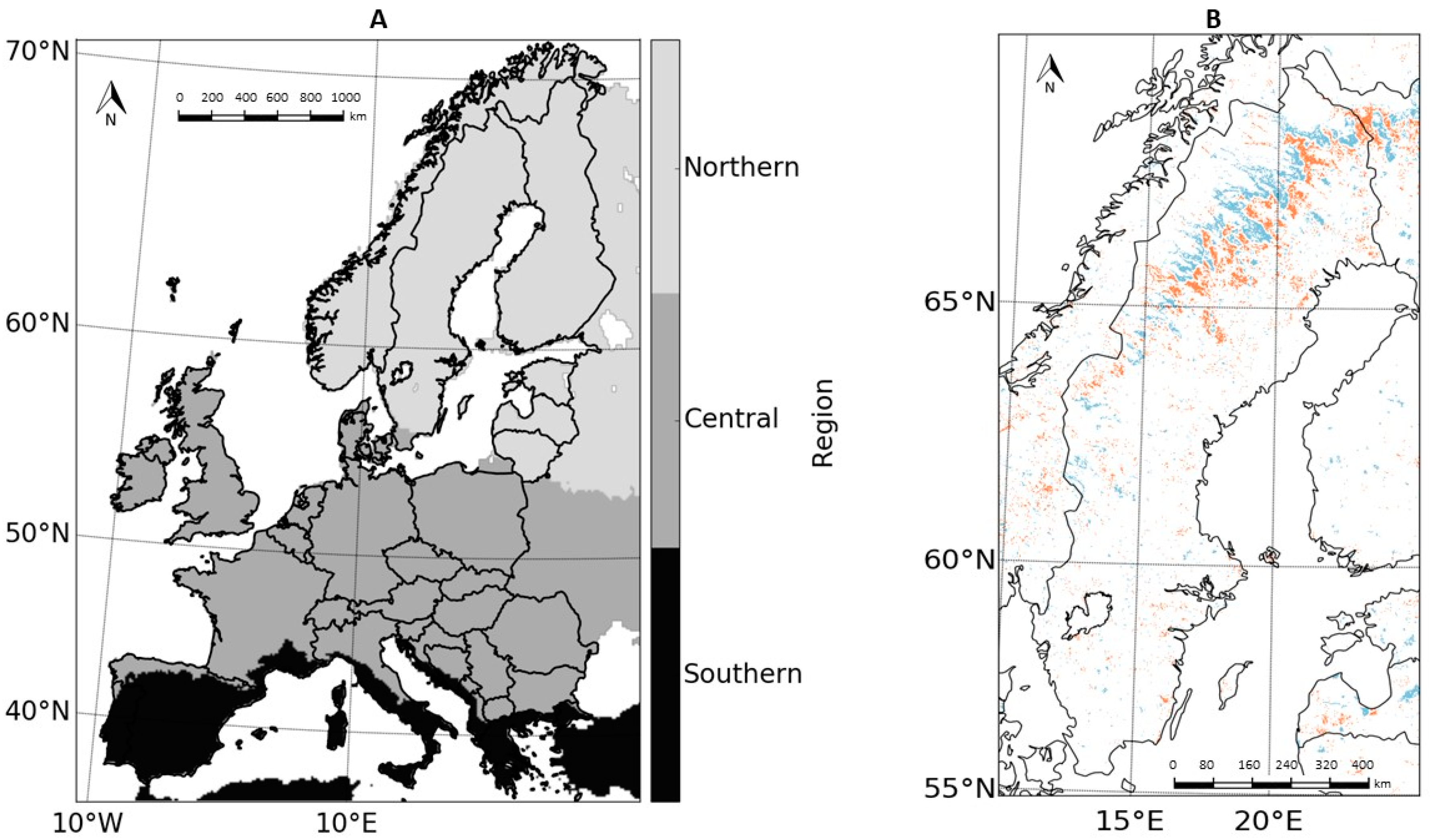
2.2. Land Characteristics: Independent Variables
2.3. Average NPP Response to Climate Anomalies: Response Variable
3. Results
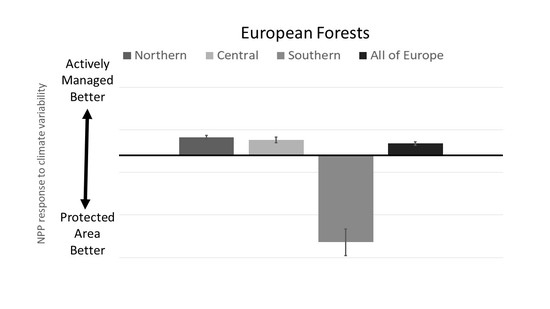
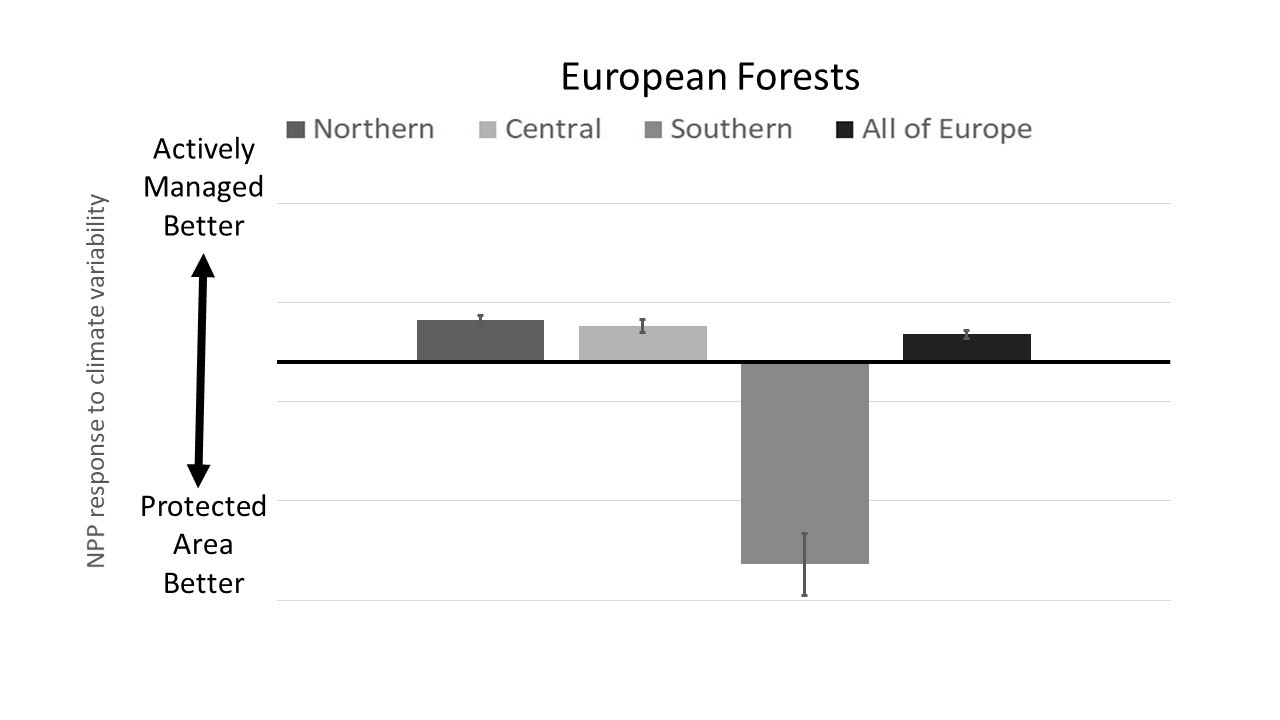
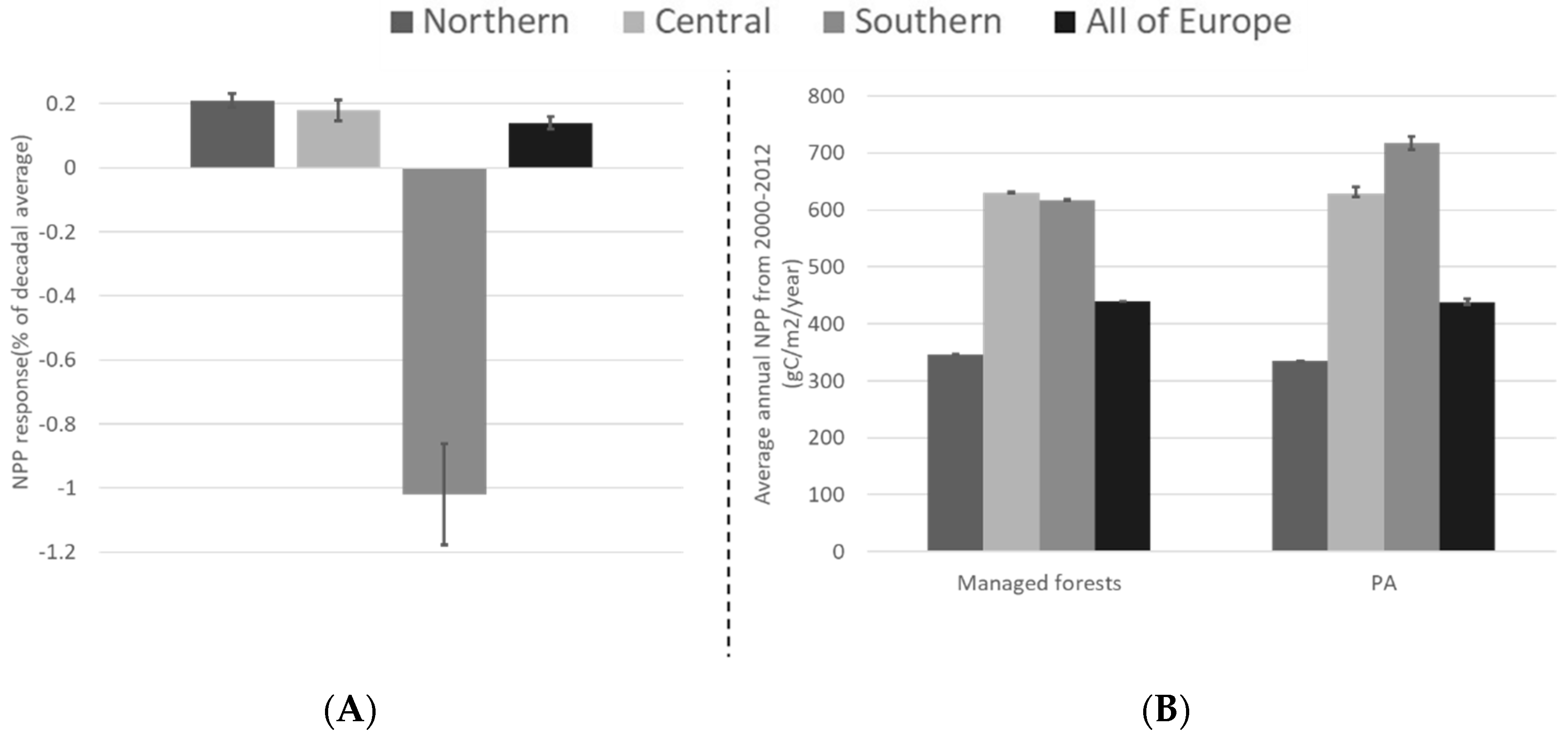
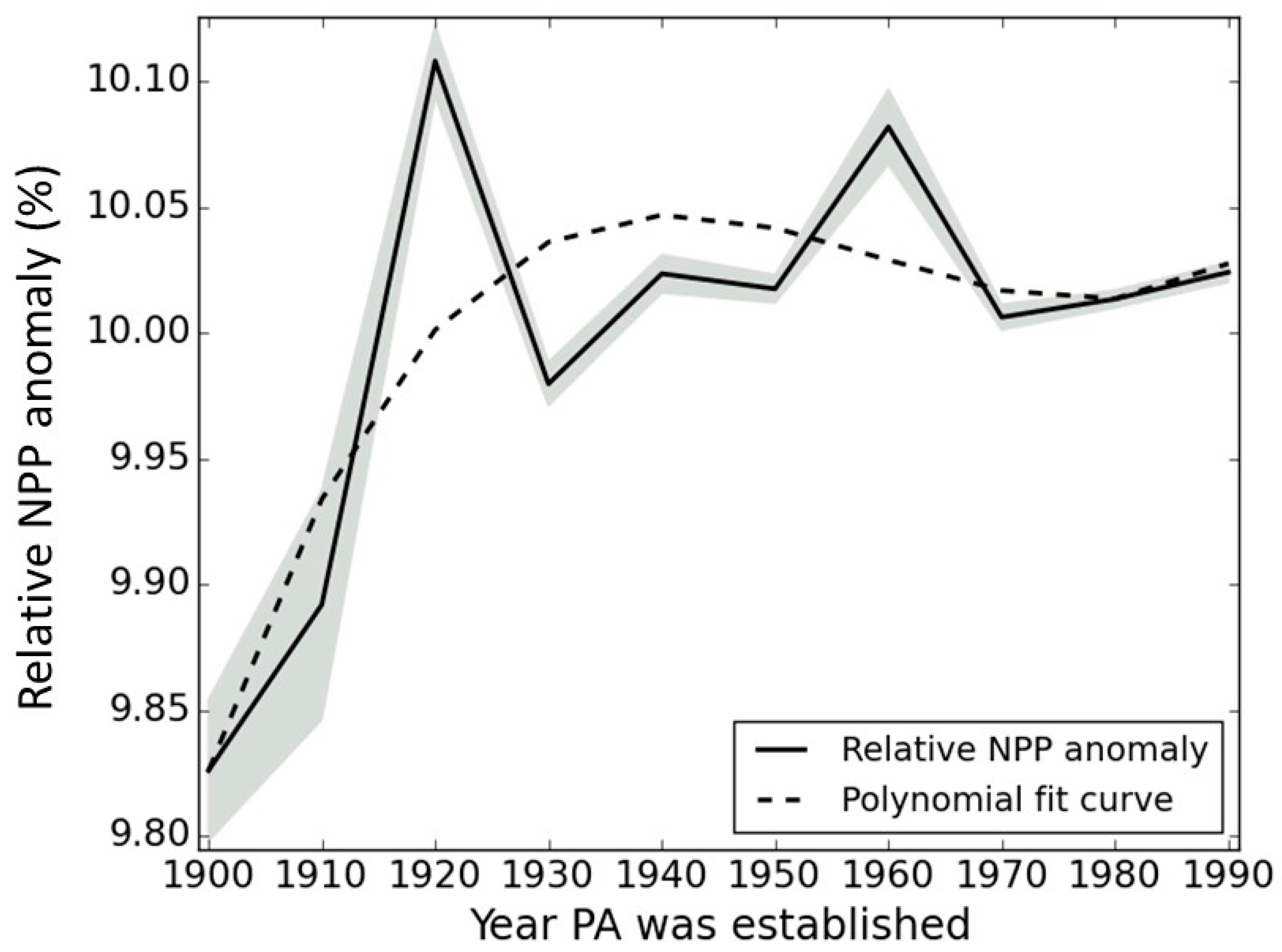
3.1. PA Impact across Europe
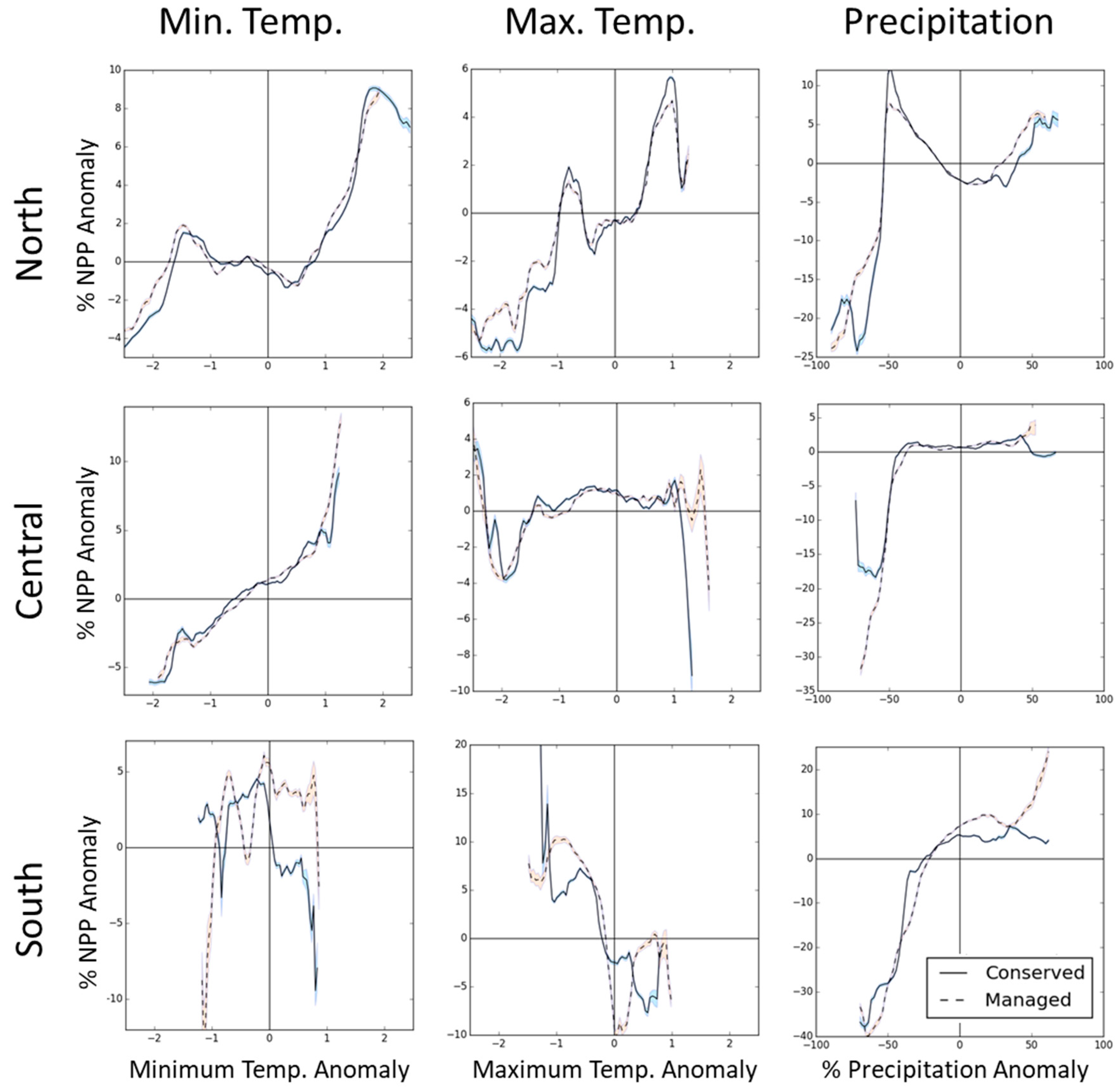
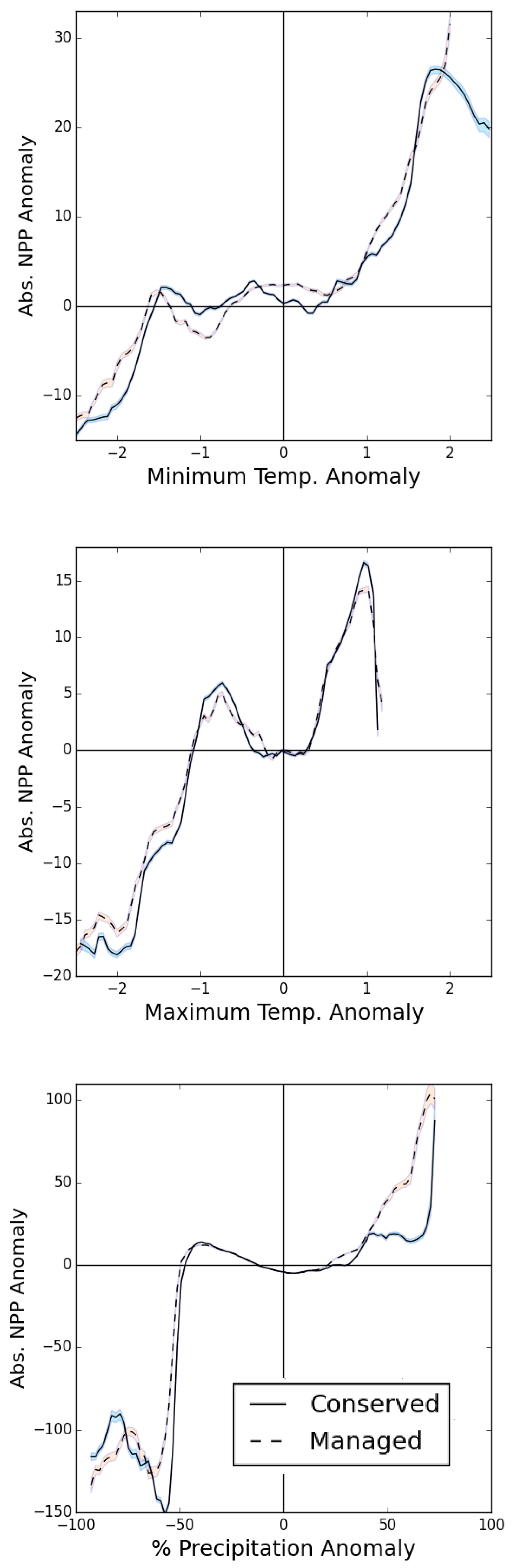
3.2. PA Regional Impacts
4. Discussion
4.1. Conservation Monitoring and Evaluation
4.2. Within the Context of European Conservation
4.3. Regional Differences in PA Impacts
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Managed | Conserved | ||||||
|---|---|---|---|---|---|---|---|
| N | % Ann. | STD | NPP | % Ann. | STD | NPP | |
| Northern | 816,023 | 7.45 | 10.05 | 346.00 | 7.66 | 10.39 | 334.51 |
| Central | 326,209 | 6.38 | 8.94 | 630.65 | 6.45 | 9.55 | 629.00 |
| Southern | 80,652 | 15.72 | 21.67 | 617.35 | 14.70 | 22.93 | 716.72 |
| All | 1,222,897 | 7.69 | 10.94 | 439.94 | 7.83 | 11.45 | 438.27 |
| N | Mean | STD | Variance | Skewness | Kurtosis | |
|---|---|---|---|---|---|---|
| Solar Radiation (W/km2) | ||||||
| All managed forests | 9,352,192 | 2118.58 | 662.78 | 439,274.00 | −0.13 | −0.65 |
| PA-forests | 94,069 | 1712.79 | 684.97 | 469,188.00 | 0.69 | −1.00 |
| Managed forests post-match | 94,069 | 1741.14 | 716.86 | 513,893.00 | 0.75 | −0.90 |
| Elevation (m) | ||||||
| All managed forests | 9,352,192 | 323.71 | 366.23 | 134,124.82 | 2.47 | 8.41 |
| PA-forests | 94,069 | 578.61 | 498.38 | 248,383.10 | 1.53 | 2.30 |
| Managed forests post-match | 94,069 | 562.25 | 484.98 | 235,207.74 | 1.54 | 2.24 |
| Precipitation (cm/yr) | ||||||
| All managed forests | 9,352,192 | 1.98 | 0.82 | 67.99 | 2.71 | 10.80 |
| PA-forests | 94,069 | 1.92 | 0.79 | 63.00 | 2.62 | 8.39 |
| Managed forests post-match | 94,069 | 1.91 | 0.74 | 54.12 | 2.34 | 6.66 |
| Minimum Temperature (°C) | ||||||
| All managed forests | 9,352,192 | 3.95 | 4.43 | 1959.66 | −0.34 | −0.43 |
| PA-forests | 94,069 | −0.48 | 4.65 | 2157.98 | 0.96 | 0.25 |
| Managed forests post-match | 94,069 | −0.48 | 4.31 | 1860.91 | 0.82 | −0.16 |
| Maximum Temperature (°C) | ||||||
| All managed forests | 9,352,192 | 12.79 | 5.28 | 2785.68 | −0.01 | −0.46 |
| PA-forests | 94,069 | 7.67 | 4.64 | 2154.80 | 1.00 | 0.28 |
| Managed forests post-match | 94,069 | 7.98 | 4.75 | 2257.38 | 1.01 | 0.25 |
| Vapor Pressure Deficit (Pa) | ||||||
| All managed forests | 9,352,192 | 5549.63 | 2309.87 | 5,335,510.00 | 0.69 | −0.17 |
| PA-forests | 94,069 | 3923.85 | 1494.05 | 2,232,180.00 | 1.90 | 4.83 |
| Managed forests post-match | 94,069 | 4113.03 | 1746.33 | 3,049,660.00 | 1.89 | 3.66 |
References
- European Commission Natura 2000 Newsletter. Available online: http://ec.europa.eu/environment/nature/info/pubs/natura2000nl_en.htm (accessed on 14 October 2018).
- Millennium Ecosystem Assessment. Ecosystems and Human Well-being: Synthesis; Island Press: Washington, DC, USA, 2005; ISBN 1597260401. [Google Scholar]
- Beresford, A.E.; Buchanan, G.M.; Sanderson, F.J.; Jefferson, R.; Donald, P.F. The Contributions of the EU Nature Directives to the CBD and Other Multilateral Environmental Agreements. Conserv. Lett. 2016, 9, 479–488. [Google Scholar] [CrossRef] [Green Version]
- Daily, G.C.; Alexander, S.; Ehrlich, P.R.; Goulder, L.; Lubchenco, J.; Matson, P.A.; Mooney, H.A.; Postel, S.; Schneider, S.H.; Tilman, D.; et al. Ecosystem Services: Benefits Supplied to Human Societies by Natural Ecosystems. Issues Ecol. 1997, 2, 1–18. [Google Scholar]
- Day, J.; Dudley, N.; Hockings, M.; Holmes, G.; Laffoley, D.; Stolton, S. Guidelines for Applying the IUCN Protected Area Management Categories to Marine Protected Areas; IUCN: Gland, Switzerland, 2012. [Google Scholar]
- European Commission Natura 2000 and Forests. Part I-II. Tech. Rep. Environ. 2015, 88, 114.
- Joppa, L.N.; Pfaff, A. Global protected area impacts. Proc. R. Soc. B Biol. Sci. 2011, 278, 1633–1638. [Google Scholar] [CrossRef] [PubMed]
- Geldmann, J.; Barnes, M.; Coad, L.; Craigie, I.D.; Hockings, M.; Burgess, N.D. Effectiveness of terrestrial protected areas in reducing habitat loss and population declines. Biol. Conserv. 2013, 161, 230–238. [Google Scholar] [CrossRef]
- Gray, C.L.; Hill, S.L.L.; Newbold, T.; Hudson, L.N.; Börger, L.; Contu, S.; Hoskins, A.J.; Ferrier, S.; Purvis, A.; Scharlemann, J.P.W. Local biodiversity is higher inside than outside terrestrial protected areas worldwide. Nat. Commun. 2016, 7, 12306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balmford, A.; Bennun, L.; Ten Brink, B.; Cooper, D.; Côté, I.M.; Crane, P.; Dobson, A.; Dudley, N.; Dutton, I.; Green, R.E.; et al. The Convention on Biological Diversity’s 2010 Target. Science 2005, 307, 212–213. [Google Scholar] [CrossRef]
- Trouwborst, A.; Chapron, G.; Fleurke, F.; Epstein, Y.; López-Bao, J.V. Europe’s biodiversity avoids fatal setback. Science 2017, 355, 140. [Google Scholar] [CrossRef]
- Orlikowska, E.H.; Roberge, J.-M.; Blicharska, M.; Mikusiński, G. Gaps in ecological research on the world’s largest internationally coordinated network of protected areas: A review of Natura 2000. Biol. Conserv. 2016, 200, 216–227. [Google Scholar] [CrossRef]
- PAILLET, Y.; BERGÈS, L.; HJÄLTÉN, J.; ÓDOR, P.; AVON, C.; BERNHARDT-RÖMERMANN, M.; BIJLSMA, R.-J.; DE BRUYN, L.; FUHR, M.; GRANDIN, U.; et al. Biodiversity Differences between Managed and Unmanaged Forests: Meta-Analysis of Species Richness in Europe. Conserv. Biol. 2010, 24, 101–112. [Google Scholar] [CrossRef] [Green Version]
- De Koning, J.; Winkel, G.; Sotirov, M.; Blondet, M.; Borras, L.; Ferranti, F.; Geitzenauer, M. Natura 2000 and climate change—Polarisation, uncertainty, and pragmatism in discourses on forest conservation and management in Europe. Environ. Sci. Policy 2014, 39, 129–138. [Google Scholar] [CrossRef]
- Salinger, M.J. Climate Variability and Change: Past, Present and Future—An Overview. Clim. Chang. 2005, 70, 9–29. [Google Scholar] [CrossRef]
- Anderegg, W.R.L.; Berry, J.A.; Smith, D.D.; Sperry, J.S.; Anderegg, L.D.L.; Field, C.B. The roles of hydraulic and carbon stress in a widespread climate-induced forest die-off. Proc. Natl. Acad. Sci. USA 2012, 109, 233–237. [Google Scholar] [CrossRef] [PubMed]
- Neumann, M.; Mues, V.; Moreno, A.; Hasenauer, H.; Seidl, R. Climate variability drives recent tree mortality in Europe. Glob. Chang. Biol. 2017, 23, 4788–4797. [Google Scholar] [CrossRef] [PubMed]
- Miteva, D.A.; Pattanayak, S.K.; Ferraro, P.J. Evaluation of biodiversity policy instruments: What works and what doesn’t? Oxf. Rev. Econ. Policy 2012, 28, 69–92. [Google Scholar] [CrossRef]
- Faith, D.P.; Walker, P.A.; Ive, J.R.; Belbin, L. Integrating conservation and forestry production: Exploring trade-offs between biodiversity and production in regional land-use assessment. For. Ecol. Manag. 1996, 85, 251–260. [Google Scholar] [CrossRef]
- Seidl, R.; Schelhaas, M.-J.; Rammer, W.; Verkerk, P.J. Increasing forest disturbances in Europe and their impact on carbon storage. Nat. Clim. Chang. 2014, 4, 806–810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tilman, D.; Reich, P.B.; Isbell, F. Biodiversity impacts ecosystem productivity as much as resources, disturbance, or herbivory. Proc. Natl. Acad. Sci. USA 2012, 109, 10394–10397. [Google Scholar] [CrossRef] [Green Version]
- Hooper, D.U.; Chapin, F.S., III; Ewel, J.J. Effects of biodiversity on ecosystem functioning: a consensus of current knowledge. Ecol. Monogr. 2005, 75, 3–35. [Google Scholar] [CrossRef]
- Robinson, N.P.; Allred, B.W.; Smith, W.K.; Jones, M.O.; Moreno, A.; Erickson, T.A.; Naugle, D.E.; Running, S.W. Terrestrial primary production for the conterminous United States derived from Landsat 30 m and MODIS 250 m. Remote Sens. Ecol. Conserv. 2018, 1–17. [Google Scholar] [CrossRef]
- Joppa, L.N.; Pfaff, A. High and Far: Biases in the Location of Protected Areas. PLoS ONE 2009, 4, e8273. [Google Scholar] [CrossRef] [PubMed]
- Cleveland, C.C.; Townsend, A.R.; Taylor, P.; Alvarez-Clare, S.; Bustamante, M.M.C.; Chuyong, G.; Dobrowski, S.Z.; Grierson, P.; Harms, K.E.; Houlton, B.Z.; et al. Relationships among net primary productivity, nutrients and climate in tropical rain forest: A pan-tropical analysis. Ecol. Lett. 2011, 14, 939–947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chapin, F.S., III; Zavaleta, E.S.; Eviner, V.T.; Naylor, R.L.; Vitousek, P.M.; Reynolds, H.L.; Hooper, D.U.; Lavorel, S.; Sala, O.E.; Hobbie, S.E.; et al. Consequences of changing biodiversity. Nature 2000, 405, 234–242. [Google Scholar] [CrossRef] [PubMed]
- Thuiller, W. Climate change and the ecologist. Nature 2007, 448, 550–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Environment Agency Nationally Designated Areas (CDDA). Available online: https://www.eea.europa.eu (accessed on 14 October 2018).
- Forest Europe. State of Europe’s Forests 2015. In Proceedings of the Ministerial Conference on the Protection of Forests in Europe FOREST EUROPE Liaison Unit, Madrid, Spain, 20–21 October 2015. [Google Scholar]
- European Environment Agency European Biogeographical Regions. Available online: https://www.eea.europa.eu/data-and-maps/data/biogeographical-regions-europe-3 (accessed on 14 October 2018).
- Friedl, M.; McIver, D.K.; Hodges, J.C.F.; Zhang, X.; Muchoney, D.; Strahler, A.H.; Woodcock, C.E.; Gopal, S.; Schneider, A.; Cooper, A.; et al. Global land cover mapping from MODIS: algorithms and early results. Remote Sens. Environ. 2002, 83, 287–302. [Google Scholar] [CrossRef]
- Moreno, A.; Hasenauer, H. Spatial downscaling of European climate data. Int. J. Climatol. 2016, 36, 1444–1458. [Google Scholar] [CrossRef]
- USGS GTOPO30 Global 30 Arc-Second Elevation (GTOPO30). Available online: https://lta.cr.usgs.gov/GTOPO30 (accessed on 14 October 2018).
- Newbold, T.; Hudson, L.N.; Hill, S.L.; Contu, S.; Lysenko, I.; Senior, R.; Börger, L.; Bennett, D.; Choimes, A.; Collen, B.; et al. Global effects of land use on local terrestrial biodiversity. Nature 2015, 520, 45. [Google Scholar] [CrossRef]
- Neumann, M.; Moreno, A.; Thurnher, C.; Mues, V.; Härkönen, S.; Mura, M.; Bouriaud, O.; Lang, M.; Cardellini, G.; Thivolle-Cazat, A.; et al. Creating a Regional MODIS Satellite-Driven Net Primary Production Dataset for European Forests. Remote Sens. 2016, 8, 554. [Google Scholar] [CrossRef]
- Neumann, M.; Smith, P. Carbon uptake by European agricultural land is variable, and in many regions could be increased: Evidence from remote sensing, yield statistics and models of potential productivity. Sci. Total Environ. 2018, 643, 902–911. [Google Scholar] [CrossRef]
- Moreno, A.; Neumann, M.; Hasenauer, H. Forest structures across Europe. Geosci. Data J. 2017, 4, 17–28. [Google Scholar] [CrossRef] [Green Version]
- Biodiversity Information System for Europe. Available online: https://biodiversity.europa.eu/ (accessed on 14 October 2018).
- Kallimanis, A.S.; Touloumis, K.; Tzanopoulos, J.; Mazaris, A.D.; Apostolopoulou, E.; Stefanidou, S.; Scott, A.V.; Potts, S.G.; Pantis, J.D. Vegetation coverage change in the EU: patterns inside and outside Natura 2000 protected areas. Biodivers. Conserv. 2015, 24, 579–591. [Google Scholar] [CrossRef]
- Gerard, F.; Petit, S.; Smith, G.; Thomson, A.; Brown, N.; Manchester, S.; Wadsworth, R.; Bugar, G.; Halada, L.; Bezák, P.; et al. Land cover change in Europe between 1950 and 2000 determined employing aerial photography. Prog. Phys. Geogr. 2010, 34, 183–205. [Google Scholar] [CrossRef] [Green Version]
- Nagendra, H. Do Parks Work? Impact of Protected Areas on Land Cover Clearing. AMBIO J. Hum. Environ. 2008, 37, 330–337. [Google Scholar] [CrossRef]
- Vincent, J.R. Impact Evaluation of Forest Conservation Programs: Benefit-Cost Analysis, Without the Economics. Environ. Resour. Econ. 2016, 63, 395–408. [Google Scholar] [CrossRef]
- Ho, D.E.; Imai, K.; King, G.; Stuart, E.A. Matching as nonparametric preprocessing for reducing model dependence in parametric causal inference. Political Anal. 2007, 15, 199–236. [Google Scholar] [CrossRef]
- Mohebalian, P.M.; Aguilar, F.X. Beneath the Canopy: Tropical Forests Enrolled in Conservation Payments Reveal Evidence of Less Degradation. Ecol. Econ. 2018, 143, 64–73. [Google Scholar] [CrossRef]
- Pearse, P.H. The Optimum Forest Rotation. For. Chron. 1967, 43, 178–195. [Google Scholar] [CrossRef]
- Davis, M.; Naumann, S.; McFarland, K.; Graf, A.; Evans, D. Literature Review The ecological effectiveness of the Natura 2000 Network. Tech. Pap. Eur. Environ. Agency 2014, 5, 30. [Google Scholar]
- Tittensor, D.P.; Walpole, M.; Hill, S.L.L.; Boyce, D.G.; Britten, G.L.; Burgess, N.D.; Butchart, S.H.M.; Leadley, P.W.; Regan, E.C.; Alkemade, R.; et al. A mid-term analysis of progress toward international biodiversity targets. Science 2014, 346, 241–244. [Google Scholar] [CrossRef]
- Butchart, S.H.M.; Clarke, M.; Smith, R.J.; Sykes, R.E.; Scharlemann, J.P.W.; Harfoot, M.; Buchanan, G.M.; Angulo, A.; Balmford, A.; Bertzky, B.; et al. Shortfalls and Solutions for Meeting National and Global Conservation Area Targets. Conserv. Lett. 2015, 8, 329–337. [Google Scholar] [CrossRef] [Green Version]
- Fady-Welterlen, B. Is There Really More Biodiversity in Mediterranean Forest Ecosystems? Taxon 2005, 54, 905. [Google Scholar] [CrossRef]
- Mauri, A.; Strona, G.; San-Miguel-Ayanz, J. EU-Forest, a high-resolution tree occurrence dataset for Europe. Sci. Data 2017, 4, 160123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paquette, A.; Messier, C. The effect of biodiversity on tree productivity: from temperate to boreal forests. Glob. Ecol. Biogeogr. 2011, 20, 170–180. [Google Scholar] [CrossRef]
- Dallemand, E.J.F.; Petersen, J.E.; Karp, A. Short Rotation Forestry, Short Rotation Coppice and perennial grasses in the European Union: Agro-environmental aspects, present use and perspectives. JRC Sci. Tech. Rep. 2008, 166. Available online: https://iet.jrc.ec.europa.eu/remea/events/short-rotation-forestry-short-rotation-coppice-and-perennial-grasses-european-union-agro (accessed on 26 December 2018).
- Laughlin, J.; Messina, M. Swedisk Forestry and Forest Policy. J. For. 1988, 86, 17–22. [Google Scholar] [CrossRef]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moreno, A.; Neumann, M.; Mohebalian, P.M.; Thurnher, C.; Hasenauer, H. The Continental Impact of European Forest Conservation Policy and Management on Productivity Stability. Remote Sens. 2019, 11, 87. https://doi.org/10.3390/rs11010087
Moreno A, Neumann M, Mohebalian PM, Thurnher C, Hasenauer H. The Continental Impact of European Forest Conservation Policy and Management on Productivity Stability. Remote Sensing. 2019; 11(1):87. https://doi.org/10.3390/rs11010087
Chicago/Turabian StyleMoreno, Adam, Mathias Neumann, Phillip M. Mohebalian, Christopher Thurnher, and Hubert Hasenauer. 2019. "The Continental Impact of European Forest Conservation Policy and Management on Productivity Stability" Remote Sensing 11, no. 1: 87. https://doi.org/10.3390/rs11010087
APA StyleMoreno, A., Neumann, M., Mohebalian, P. M., Thurnher, C., & Hasenauer, H. (2019). The Continental Impact of European Forest Conservation Policy and Management on Productivity Stability. Remote Sensing, 11(1), 87. https://doi.org/10.3390/rs11010087