
A Validated and Accurate Method for Quantifying and Extrapolating Mangrove Above-Ground Biomass Using LiDAR Data


Abstract
:
1. Introduction
- Collect field data of mangrove structure at Darwin Harbour as a training set to model relationships with remotely sensed LiDAR data;
- Establish optimal relationships between field-derived mangrove structural measurements and remotely sensed data describing mangrove structure;
- Validate relationships against field-based validation data;
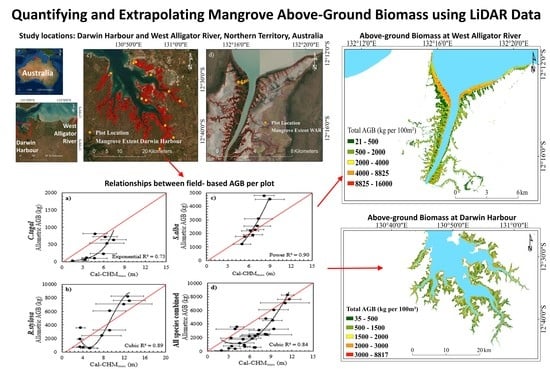
- Apply optimal relationships to LiDAR data, estimate mangrove above-ground biomass at Darwin Harbour, and compare with published estimates of mangrove above-ground biomass [26]; and
- Extrapolate relationships to the West Alligator River and compare with published estimates of mangrove above-ground biomass.
2. Materials and Methods
2.1. Study Sites
2.1.1. Darwin Harbour, Northern Territory, Australia
2.1.2. West Alligator River, Northern Territory, Australia
2.2. Study Approach, LiDAR Data, and Processing
2.3. Classifying Species Distribution
2.4. Field-Based Measurements of Mangrove Forest Characteristics
- DBH = diameter at breast height.
- β0 and β1 = regression coefficients.
- β0 and β1 for Rhizophora stylora = −0.696 and 2.465.
- β0 and β1 for Ceriops tagal = −0.494 and 2.056.
- β0 and β1 for Sonneratia alba = 0.258 and 2.287.
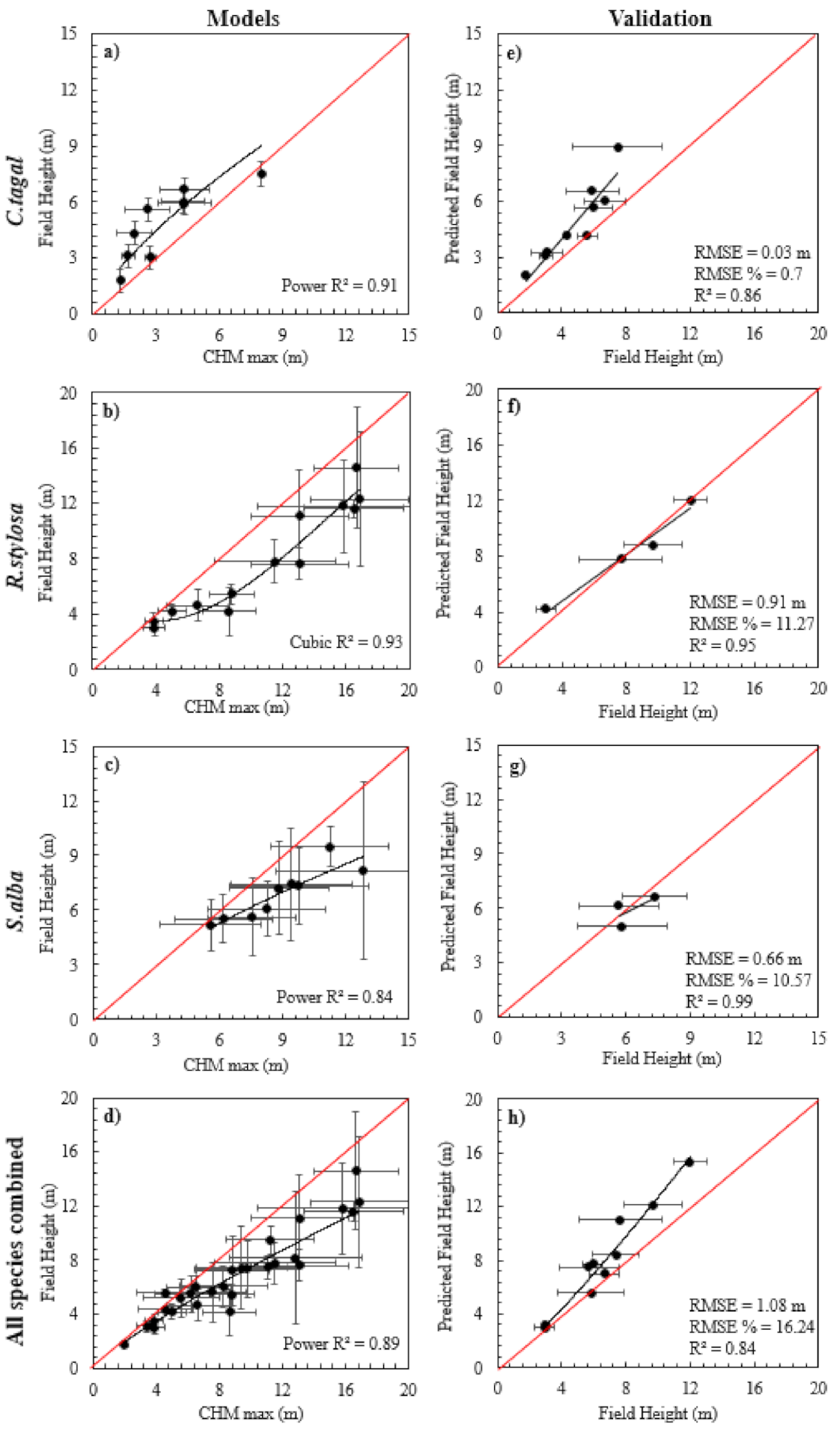
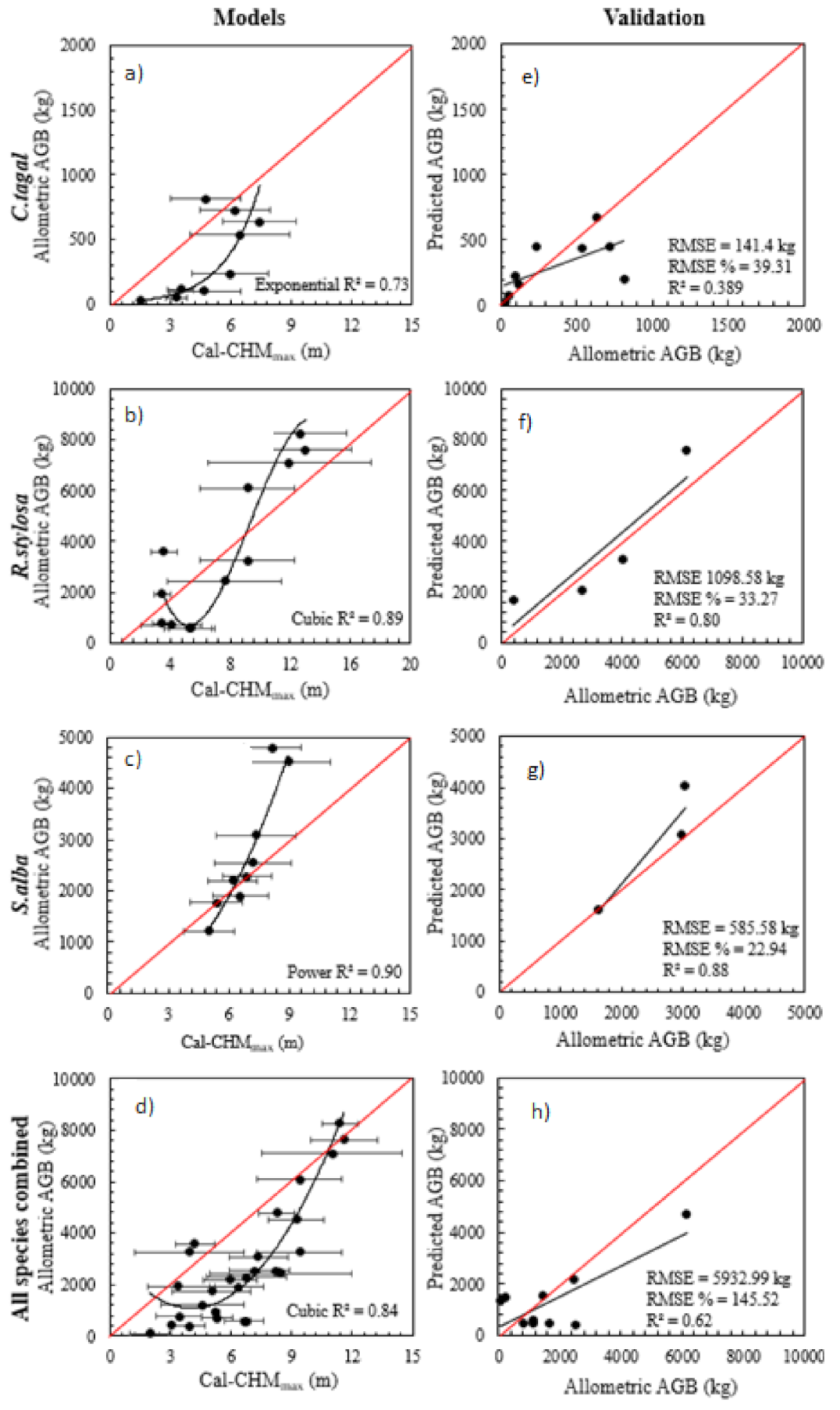
2.5. Modelling and Validating Relationships between Mangrove Forest Characteristics and LiDAR Data
2.6. Model Extrapolation and Comparison at West Alligator River
3. Results
3.1. Mangrove Forest Characteristics
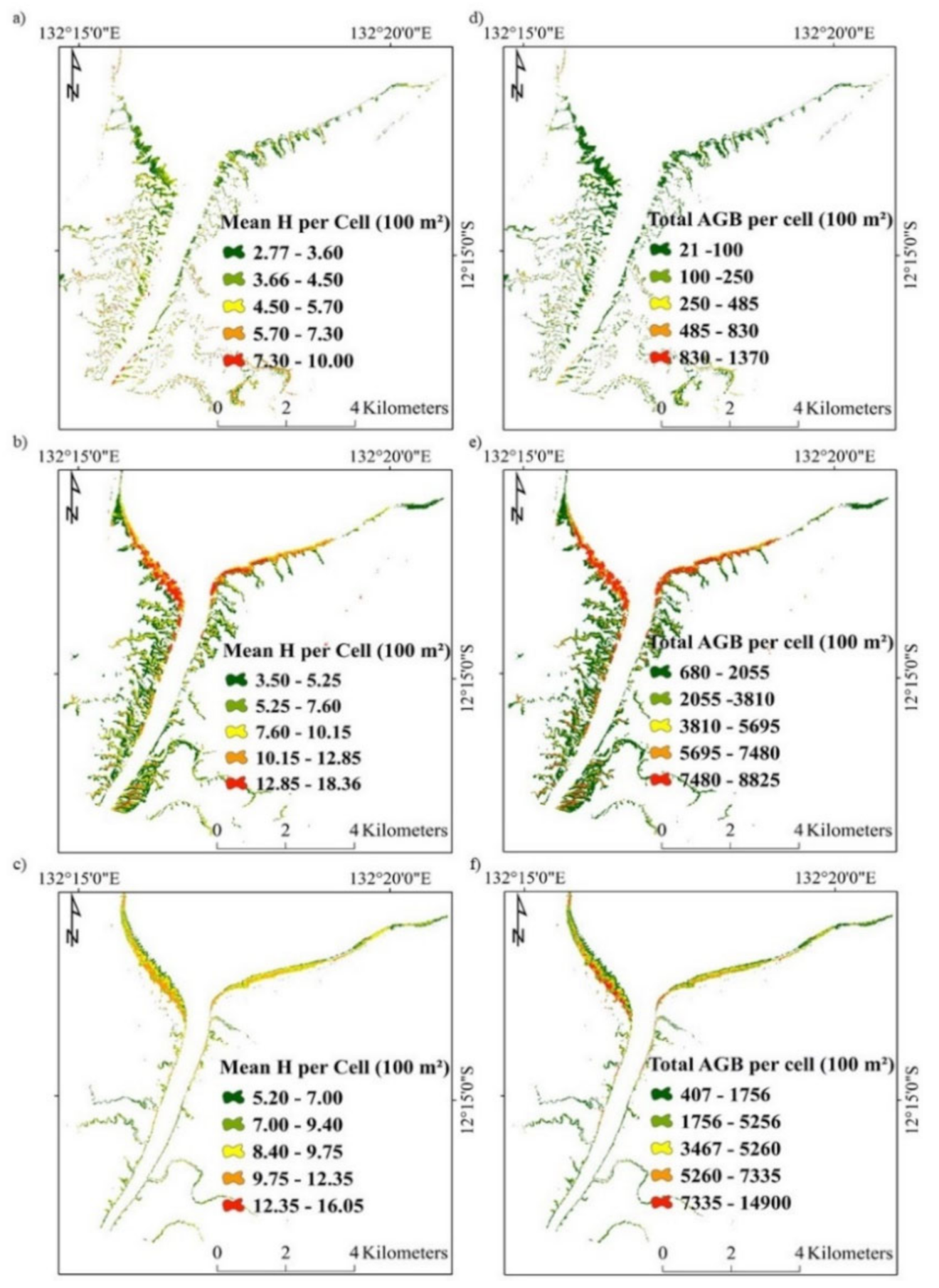
3.2. Model Extrapolation and Comparison at the West Alligator River
4. Discussion
4.1. Quantifying and Extrapolating Mangrove Biomass Using LiDAR Data
4.2. Implications for Blue Carbon Assessments
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
Appendix B
Appendix C
Appendix D
References
- Donato, D.C.; Kauffman, J.B.; Murdiyarso, D.; Kurnianto, S.; Stidham, M.; Kanninen, M. Mangroves among the most carbon-rich forests in the tropics. Nat. Geosci. 2011, 4, 293–297. [Google Scholar] [CrossRef]
- Komiyama, A. Common allometric equations for estimating the tree weight of mangroves. J. Trop. Ecol. 2005, 21, 471–477. [Google Scholar] [CrossRef]
- Chave, J.; Andalo, C.; Brown, S.; Cairns, M.A.; Chambers, J.Q.; Eamus, D.; Folster, H.; Fromard, F.; Higuchi, N.; Kira, T.; et al. Tree allometry and improved estimation of carbon stocks and balance in tropical forests. Oecologia 2005, 145, 87–99. [Google Scholar] [CrossRef] [PubMed]
- Shaffer-Novelli, Y.; Molero-Cintron, G. Guia PARA ESTUDO DE AREAS DE MANGUEZAL: Estrutura, Funcao e Flora; Caribbean Ecological Research: Sao Paulo, Brazil, 1985. [Google Scholar]
- Brocklehurst, P.S.; Edmeades, B.F. The Mangrove Communities of Darwin Harbor; Department of Lands, Planning and Environment: Darwin, Australia, 1996; p. 171. [Google Scholar]
- Salum, R.B.; Souza-Filho, P.W.M.; Simard, M.; Silva, C.A.; Fernandes, M.E.B.; Cougo, M.F.; do Nascimento, W.; Rogers, K. Improving mangrove above-ground biomass estimates using LiDAR. Estuarine Coast. Shelf Sci. 2020, 236. [Google Scholar] [CrossRef]
- Hu, T.; Zhang, Y.; Su, Y.; Zheng, Y.; Lin, G.; Guo, Q. Mapping the Global Mangrove Forest Aboveground Biomass Using Multisource Remote Sensing Data. Remote Sens. 2020, 12, 1690. [Google Scholar] [CrossRef]
- Wang, D.; Wan, B.; Liu, J.; Su, Y.; Guo, Q.; Qiu, P.; Wu, X. Estimating aboveground biomass of the mangrove forests on northeast Hainan Island in China using an upscaling method from field plots, UAV-LiDAR data and Sentinel-2 imagery. Int. J. Appl. Earth Observ. Geoinf. 2020, 85. [Google Scholar] [CrossRef]
- Qiu, P.; Wang, D.; Zou, X.; Yang, X.; Xie, G.; Xu, S.; Zhong, Z. Finer Resolution Estimation and Mapping of Mangrove Biomass Using UAV LiDAR and WorldView-2 Data. Forests 2019, 10, 871. [Google Scholar] [CrossRef] [Green Version]
- Simard, M.; Fatoyinbo, L.; Smetanka, C.; Rivera-Monroy, V.H.; Castañeda-Moya, E.; Thomas, N.; Van der Stocken, T. Mangrove canopy height globally related to precipitation, temperature and cyclone frequency. Nat. Geosci. 2018, 12, 40–45. [Google Scholar] [CrossRef]
- Tang, W.; Zheng, M.; Zhao, X.; Shi, J.; Yang, J.; Trettin, C. Big Geospatial Data Analytics for Global Mangrove Biomass and Carbon Estimation. Sustainability 2018, 10, 472. [Google Scholar] [CrossRef] [Green Version]
- Hickey, S.M.; Callow, N.J.; Phinn, S.; Lovelock, C.E.; Duarte, C.M. Spatial complexities in aboveground carbon stocks of a semi-arid mangrove community: A remote sensing height-biomass-carbon approach. Estuarine Coast. Shelf Sci. 2018, 200, 194–201. [Google Scholar] [CrossRef] [Green Version]
- Fatoyinbo, T.; Feliciano, E.A.; Lagomasino, D.; Lee, S.K.; Trettin, C. Estimating mangrove aboveground biomass from airborne LiDAR data: A case study from the Zambezi River delta. Environ. Res. Lett. 2018, 13. [Google Scholar] [CrossRef] [Green Version]
- Staben, G.; Lucieer, A.; Scarth, P. Modelling LiDAR derived tree canopy height from Landsat TM, ETM+ and OLI satellite imagery—A machine learning approach. Int. J. Appl. Earth Observ. Geoinf. 2018, 73, 666–681. [Google Scholar] [CrossRef]
- Rocha de Souza Pereira, F.; Kampel, M.; Gomes Soares, M.; Estrada, G.; Bentz, C.; Vincent, G. Reducing Uncertainty in Mapping of Mangrove Aboveground Biomass Using Airborne Discrete Return Lidar Data. Remote Sens. 2018, 10, 637. [Google Scholar] [CrossRef] [Green Version]
- Fatoyinbo, T.E.; Simard, M. Height and biomass of mangroves in Africa from ICESat/GLAS and SRTM. Int. J. Remote Sens. 2013, 34, 668–681. [Google Scholar] [CrossRef]
- Heenkenda, M.; Maier, S.; Joyce, K. Estimating Mangrove Biophysical Variables Using WorldView-2 Satellite Data: Rapid Creek, Northern Territory, Australia. J. Imaging 2016, 2, 24. [Google Scholar] [CrossRef] [Green Version]
- Cougo, M.; Souza-Filho, P.; Silva, A.; Fernandes, M.; Santos, J.; Abreu, M.; Nascimento, W.; Simard, M. Radarsat-2 Backscattering for the Modeling of Biophysical Parameters of Regenerating Mangrove Forests. Remote Sens. 2015, 7, 5873. [Google Scholar] [CrossRef] [Green Version]
- Proisy, C.; Couteron, P.; Fromard, F. Predicting and mapping mangrove biomass from canopy grain analysis using Fourier-based textural ordination of IKONOS images. Remote Sens. Environ. 2007, 109, 379–392. [Google Scholar] [CrossRef]
- Lucas, R.; Van De Kerchove, R.; Otero, V.; Lagomasino, D.; Fatoyinbo, L.; Omar, H.; Satyanarayana, B.; Dahdouh-Guebas, F. Structural characterisation of mangrove forests achieved through combining multiple sources of remote sensing data. Remote Sens. Environ. 2020, 237. [Google Scholar] [CrossRef]
- Owers, C.J.; Rogers, K.; Woodroffe, C.D. Terrestrial laser scanning to quantify above-ground biomass of structurally complex coastal wetland vegetation. Estuarine Coast. Shelf Sci. 2018, 204, 164–176. [Google Scholar] [CrossRef] [Green Version]
- Olagoke, A.; Proisy, C.; Féret, J.-B.; Blanchard, E.; Fromard, F.; Mehlig, U.; de Menezes, M.M.; Dos Santos, V.F.; Berger, U. Extended biomass allometric equations for large mangrove trees from terrestrial LiDAR data. Trees 2016, 30, 935–947. [Google Scholar] [CrossRef] [Green Version]
- Calders, K.; Newnham, G.; Burt, A.; Murphy, S.; Raumonen, P.; Herold, M.; Culvenor, D.; Avitabile, V.; Disney, M.; Armston, J. Nondestructive estimates of above-ground biomass using terrestrial laser scanning. Methods Ecol. Evol. 2015, 6, 198–208. [Google Scholar] [CrossRef]
- Kargar, A.R.; MacKenzie, R.A.; Apwong, M.; Hughes, E.; van Aardt, J. Stem and root assessment in mangrove forests using a low-cost, rapid-scan terrestrial laser scanner. Wetlands Ecol. Manag. 2020, 28, 883–900. [Google Scholar] [CrossRef]
- Rogers, K.; Lymburner, L.; Salum, R.; Brooke, B.P.; Woodroffe, C.D. Mapping mangrove extent and zonation using high and low tide composites of Landsat data. Hydrobiologia 2017, 803, 49–68. [Google Scholar] [CrossRef]
- Brocklehurst, P.S.; Edmeades, B.F. The Mangrove Communities of Darwin Harbor Biomass Update; NT Government: Darwin, Australia, 2013. [Google Scholar]
- Leong, R.C.; Friess, D.A.; Crase, B.; Lee, W.K.; Webb, E.L. High-resolution pattern of mangrove species distribution is controlled by surface elevation. Estuarine Coast. Shelf Sci. 2018, 202, 185–192. [Google Scholar] [CrossRef]
- Lucas, R.; Finlayson, C.M.; Bartolo, R.; Rogers, K.; Mitchell, A.; Woodroffe, C.D. Historical perspectives on the mangroves of Kakadu National Park. Marine Freshwater Res. 2017, 69, 1047–1063. [Google Scholar] [CrossRef]
- Crase, B.; Liedloff, A.; Vesk, P.A.; Burgman, M.A.; Wintle, B.A. Hydroperiod is the main driver of the spatial pattern of dominance in mangrove communities. Global Ecol. Biogeogr. 2013, 22, 806–817. [Google Scholar] [CrossRef]
- Woodroffe, C.D.; Bardsley, K.N.; Ward, P.J.; Hanley, J.R. Production of Mangrove Litter in a Macrotidal Embayment, Darwin Harbor, N.T., Australia. Estuarine Coast. Shelf Sci. 1988, 26, 581–598. [Google Scholar] [CrossRef]
- Semeniuk, V. Mangrove enviroments of Port Darwin Northern Territory: The physical framework and habitats. J. R. Soc. West. Aust. 1985, 67, 81–97. [Google Scholar]
- Williams, D.; Wolanski, E.; Spagnol, S. Hydrodynamics of Darwin harbour. In The Environment in Asia Pacific Harbours; Springer: Berlin, Germany, 2006; pp. 461–476. [Google Scholar]
- Walpole, B.P.; Crohn, P.W.; Dunn, P.R.; Randal, M.A. Geology of the Katherine-Darwin Region Northen Territory; Bureau of Mineral Resources, Geology and Geophysics: Canberra, Australia, 1968; p. 304. [Google Scholar]
- Woodroffe, C.; Bardsley, K. The distribution and productivity of mangroves in Creek H, Darwin Harbor. In Proceedings of the Workshop on Research and Management, Darwin, Australia, 2–3 September 1987; pp. 2–3. [Google Scholar]
- Woodroffe, C.D.; Chappell, J.; Thom, B.G.; Wallensky, E. Depositional model of a macrotidal estuary and floodplain, South Alligator River, Northern Australia. Sedimentology 1989, 36, 737–756. [Google Scholar] [CrossRef]
- Woodroffe, C.D.; Thom, B.G.; Chappell, J. Development of widespread mangrove swamps in mid-Holocene times in Northern Australia. Nature 1985, 317, 711–713. [Google Scholar] [CrossRef]
- Woodroffe, C.D. Mangrove response to sea level rise: Palaeoecological insights from macrotidal systems in Northern Australia. Mar. Freshwater Res. 2018, 69. [Google Scholar] [CrossRef] [Green Version]
- Asbridge, E.; Bartolo, R.; Finlayson, C.; Lucas, R.; Rogers, K.; Woodroffe, C. Assessing the distribution and drivers of mangrove dieback in Kakadu National Park, northern Australia. Estuarine Coast. Shelf Sci. 2019, 228, 106353. [Google Scholar] [CrossRef]
- Asbridge, E. Mangrove Response to Environmental Changes Predicted Under Varying Climates: Case Studies from Australia. Curr. For. Rep. 2015, 1, 178–194. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, A.L. Remote Sensing Techniques for Assessment of Mangrove Forest Structure, Species Composition and Biomass, and Response to Environmental Change. Ph.D. Thesis, The University of New South Wales High Street, Kensington, NSW, Australia, 2003. [Google Scholar]
- Temby, O. Project Report for Aerial LiDAR Survey of Darwin, and Surrounds; Fugro Spatial Solutions Pty Ltd.: Perth, Australia, 2009. [Google Scholar]
- Schroeder, T.; Anstee, J.; Botha, H.; Byrne, G.; Dyce, P. Kakadu LIDAR Project 2011 0.5m Contours maps. v3; CSIRO and Geoscience Australia: Canberra, Australia, 2013. [Google Scholar] [CrossRef]
- Geoscience Australia. Digital Elevation Model (DEM) of Australia Derived from LiDAR 5 Metre Grid; Geoscience Australia: Canberra, Australia, 2015. [Google Scholar]
- Khosravipour, A.; Skidmore, A.K.; Isenburg, M.; Wang, T.; Hussin, Y.A. Generating Pit-free Canopy Height Models from Airborne Lidar. Photogramm. Eng. Remote Sens. 2014, 80, 863–872. [Google Scholar] [CrossRef]
- Comley, B.W.T.; McGuinness, K.A. Above- and below-ground biomass, and allometry, of four common northern Australian mangroves. Aust. J. Bot. 2005, 53, 431. [Google Scholar] [CrossRef]
- Owers, C.J.; Rogers, K.; Woodroffe, C.D. Spatial variation of above-ground carbon storage in temperate coastal wetlands. Estuar. Coast. Shelf Sci. 2018, 2010, 55–67. [Google Scholar] [CrossRef]
- Kusmana, C.; Hidayat, T.; Tiryana, T.; Rusdiana, O. Allometric models for above- and below-ground biomass of Sonneratia spp. Glob. Ecol. Conserv. 2018, 15. [Google Scholar] [CrossRef]
- Clough, B.; Scott, K. Allometrics relationships for estiamting above ground biomass in six mangrove speceis. For. Ecol. Manag. 1989, 27, 117–127. [Google Scholar] [CrossRef]
- Rogers, K.; Krauss, K.W. Moving from Generalisations to Specificity about Mangrove–Saltmarsh Dynamics. Wetlands 2018. [Google Scholar] [CrossRef] [Green Version]
- Chadwick, J. Integrated LiDAR and IKONOS multispectral imagery for mapping mangrove distribution and physical properties. Int. J. Remote Sens. 2011, 32, 6765–6781. [Google Scholar] [CrossRef]
- Barbier, E.B.; Hacker, S.D.; Kennedy, C.; Koch, E.W.; Stier, A.C.; Silliman, B.R. The value of estuarine and coastal ecosystem services. Ecol. Monogr. 2011, 81, 169–193. [Google Scholar] [CrossRef]
- Rogers, K.; Boon, P.I.; Branigan, S.; Duke, N.C.; Field, C.D.; Fitzsimons, J.A.; Kirkman, H.; Mackenzie, J.R.; Saintilan, N. The state of legislation and policy protecting Australia’s mangrove and salt marsh and their ecosystem services. Mar. Policy 2016, 72, 139–155. [Google Scholar] [CrossRef]
- Kelleway, J.; Serrano, O.; Baldock, J.; Cannard, T.; Lavery, P.; Lovelock, C.; Macreadie, P.; Masqué, P.; Saintilan, N.; Steven, A.J.C. Technical Review of Opportunities for Including Blue Carbon in the Australian Government’s Emissions Reduction Fund; CSIRO: Canberra, Australia, 2017. [Google Scholar]
- Kelleway, J.J.; Serrano, O.; Baldock, J.A.; Burgess, R.; Cannard, T.; Lavery, P.S.; Lovelock, C.E.; Macreadie, P.I.; Masqué, P.; Newnham, M.J.G.E.C. A national approach to greenhouse gas abatement through blue carbon management. Glob. Environ. Chang. 2020, 63, 102083. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LiDAR Details (Fugro Spatial Solutions) | Darwin Harbour Study Site | West Alligator River Study Site |
|---|---|---|
| Laser model | ALS50 | ALS60 |
| Nominal flying height | 2012 m AGL | 1425 m AGL |
| Full swath width | 1425 m | 997 m |
| Distance between runs | 1276 m | 468 m |
| Airspeed | 145 knots | 145 knots |
| Average point density | 1.1 pts/m2 | 2 pts/m2 |
| Footprint diameter | 0.45 m | 0.32 m |
| Navigation mode | GPS-based | GPS-based |
| Position mode | DGPS | DGPS |
| Vertical accuracy | ±0.15 m | ±0.3 m |
| Metadata source | [43] | [44] |
| Parameter | Species | Field Measurements (Per 10 m × 10 m) | |||
|---|---|---|---|---|---|
| Minimum | x | Sx | Maximum | ||
| Height (m) | C. tagal | 1.78 | 4.88 | 1.92 | 7.48 |
| R. stylosa | 2.95 | 8.02 | 3.71 | 14.57 | |
| S. alba | 5.18 | 6.74 | 1.29 | 9.47 | |
| DBH (cm) | C. tagal | 5.03 | 6.13 | 0.97 | 7.59 |
| R. stylosa | 6.34 | 11.66 | 5.92 | 26.95 | |
| S. alba | 13.91 | 19.52 | 4.16 | 27.72 | |
| Density (trees with DBH >5 cm per plot) | C. tagal | 16 | 76 | 48 | 140 |
| R. stylosa | 8 | 28 | 15 | 55 | |
| S. alba | 5 | 10 | 4 | 19 | |
| AGB (kg) | C. tagal | 110 | 1439 | 1263 | 3257 |
| R. stylosa | 374 | 3747 | 3222 | 10,930 | |
| S. alba | 592 | 2576 | 1225 | 4793 | |
| Species | Area (ha) | AGB (Mt) | AGB (t ha−1) |
|---|---|---|---|
| C. tagal | 6161 | 0.05 | 7.4 |
| R. stylosa | 9656 | 1.72 | 179 |
| S. alba | 766 | 0.19 | 252 |
| All species | 16,584 | 1.94 | 118 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salum, R.B.; Robinson, S.A.; Rogers, K. A Validated and Accurate Method for Quantifying and Extrapolating Mangrove Above-Ground Biomass Using LiDAR Data. Remote Sens. 2021, 13, 2763. https://doi.org/10.3390/rs13142763
Salum RB, Robinson SA, Rogers K. A Validated and Accurate Method for Quantifying and Extrapolating Mangrove Above-Ground Biomass Using LiDAR Data. Remote Sensing. 2021; 13(14):2763. https://doi.org/10.3390/rs13142763
Chicago/Turabian StyleSalum, Rafaela B., Sharon A. Robinson, and Kerrylee Rogers. 2021. "A Validated and Accurate Method for Quantifying and Extrapolating Mangrove Above-Ground Biomass Using LiDAR Data" Remote Sensing 13, no. 14: 2763. https://doi.org/10.3390/rs13142763
APA StyleSalum, R. B., Robinson, S. A., & Rogers, K. (2021). A Validated and Accurate Method for Quantifying and Extrapolating Mangrove Above-Ground Biomass Using LiDAR Data. Remote Sensing, 13(14), 2763. https://doi.org/10.3390/rs13142763