
Global Vegetation Photosynthetic Phenology Products Based on MODIS Vegetation Greenness and Temperature: Modeling and Evaluation


Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Site-Level VPP Observations from FLUXNET Measurements
2.3. Global VPP Estimates from MODIS EVI and Land Surface Temperature
2.4. Evaluating Differences between VPP Estimates and MODIS-LSP Products
2.4.1. Comparison with MODIS-Derived LSP
2.4.2. Comparing the Relationship of VPP and LSP with GPP Products
2.5. Analytical Methods
3. Results
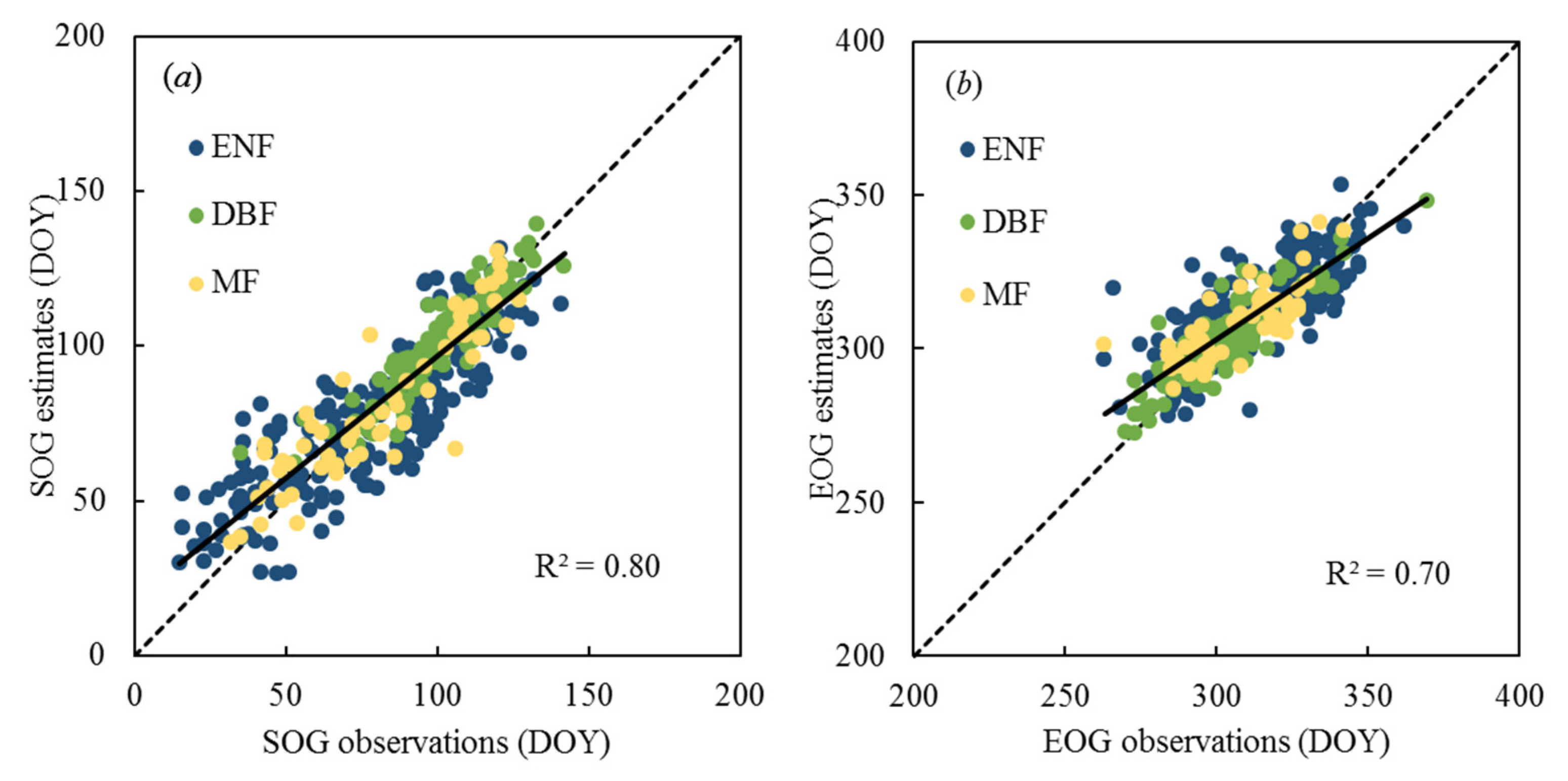
3.1. Comparison of Site-Level VPP Estimates with VPP Observations from FLUXNET GPP
3.2. Spatial Distribution of Global VPP Estimates
3.3. Comparisons of VPP Estimates with MODIS-LSP
3.4. Relationship between VPP, MODIS-LSP, and GPP
4. Discussion
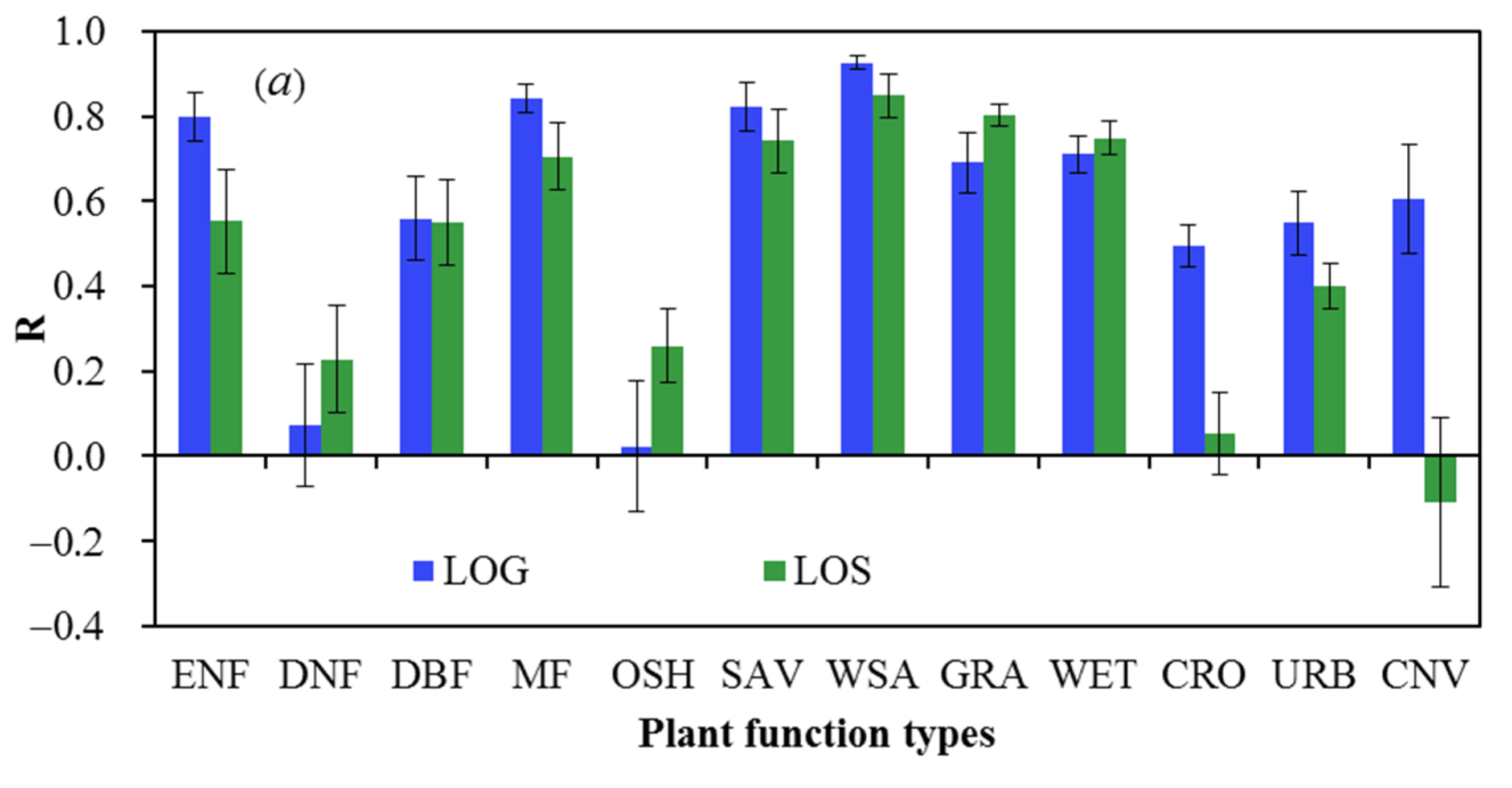
4.1. Accuracy of Site-Level VPP Estimates across PFTs
4.2. Discrepancies between VPP Estimates from the Regression Models and MODIS-LSP
4.3. Limitations of the Method
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| LSP | Land surface phenology |
| MODIS-LSP | Land surface phenology downloaded from the MODIS dataset |
| SOS | Start of growing season, refers to MODIS-SOS |
| EOS | End of growing season, refers to MODIS-EOS |
| LOS | Length of growing season, equals EOS minus SOS, and refers to MODIS-LOS |
| VPP | Vegetation photosynthetic phenology, implying the key transitions of vegetation carbon fluxes |
| VPP obs | VPP derived from the FLUXNET daily gross primary productivity (GPP) data |
| VPP est | VPP estimated based on the relationship between VPP observations and MODIS products using the regression models |
| SOG | Starting days of GPP, from VPPobs or VPPest |
| EOG | Ending days of GPP, from VPPobs or VPPest |
| LOG | Length of the photosynthetic phenology season, equal to EOG minus SOG. |
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Function Types | Model Formula | |
|---|---|---|
| Forest | ENF | SOG = 684.40 − 115.13 × mEVIsp − 165.15 × sEVIsp + 70.16 × EVI243 − 2.09 × LST60 |
| DBF | SOG = 237.01 − 116.82 × mEVIsp + 86.70 × mEVIsu − 0.43 × LST243 | |
| DNF | ||
| MF | SOG = −73.94 − 94.49 × EVI334 − 2.20 × LST60 + 2.77 × mLSTsu − 2362.37 × cLSTsu | |
| Non-forest | CSH | SOG = −51.57 − 243.87 × mEVIsp + 117.05 × EVI243 + 2.88 × LST152 − 2.30 × LST243 + 221.19 × sEVIsu |
| OSH | ||
| SAV | ||
| WSA | ||
| GRA | SOG = 180.18 − 180.55 × mEVIsp + 207.39 × sEVIsu − 3.25 × sLSTau | |
| CRO | SOG = 88.28 − 114.86 × mEVIsp + 141.83 × maxEVIsu −51.35 × EVI152 | |
| WET | SOG = 521.47 − 1.40 × mLSTsp + 352.64 × sEVIsu − 62.34 × mEVIsp | |
Appendix B
| Plant Function Types | Model Formula | |
|---|---|---|
| Forest | ENF | EOG = −1.80 + 40.66 × mEVIsp + 175.16 × sEVIsp − 1.47 × maxLSTsu + 2.51 × mLSTau |
| DBF DNF | EOG = −82.16− 15.40 × EVI152 + 98.14 × mEVIau + 1.17 × mLSTau | |
| MF | EOG = −287.59 + 52.09 × mEVIsp + 1.93 × mLSTau | |
| Non-forest | CSH OSH SAV WSA | EOG =459.51 + 149.45 × mEVIau + 2.40 × LST152 − 1.71 × LST243 + 2.84 × mLSTsp −6.31 × mLSTau + 2.05 × LST334 − 58.60 × mEVIsp |
| GRA | EOG = 290.49 − 10.46 × sEVIsu + 48.94 × mEVIau + 501.01 × cLSTsp − 6.02 × sLSTau | |
| CRO | EOG = 208.56 + 189.10 × EVI243 − 185.12 × EVI152 + 390.12 × sEVIsp-362.92 × sEVIau + 59.43 × mEVIsu | |
| WET | EOG = −400.64 + 1.15 × LST334 − 175.05 × sEVIsp + 1.28 × LST243 | |
References
- Friedl, M.; Henebry, G.M.; Reed, B.; Huete, A.; White, M.; Morisette, J.; Nemani, R.; Zhang, X.; Myneni, R. Land Surface Phenology. A Community White Paper Requested by NASA. 2006. Available online: https://lcluc.umd.edu/documents/friedl-m-g-henebry-b-reed-huete-m-white-j-morisette-r-nemani-x-zhang-and-r-myneni-2006 (accessed on 15 May 2021).
- Zhang, X.; Liu, L.; Liu, Y.; Jayavelu, S.; Wang, J.; Moon, M.; Henebry, G.M.; Friedl, M.A.; Schaaf, C.B. Generation and evaluation of the VIIRS land surface phenology product. Remote Sens. Environ. 2018, 216, 212–229. [Google Scholar] [CrossRef]
- Keenan, T.F.; Gray, J.; Friedl, M.A.; Toomey, M.; Bohrer, G.; Hollinger, D.Y.; Munger, J.M.; O’Keefe, J.; Schmid, H.P.; Wing, I.S.; et al. Net carbon uptake has increased through warming-induced changes in temperate forest phenology. Nat. Clim. Chang. 2014, 4, 598–604. [Google Scholar] [CrossRef]
- Xia, J.; Niu, S.; Ciais, P.; Janssens, I.A.; Chen, J.; Ammann, C.; Arain, A.; Blanken, P.D.; Cescatti, A.; Bonal, D.; et al. Joint control of terrestrial gross primary productivity by plant phenology and physiology. Proc. Natl. Acad. Sci. USA 2015, 112, 2788–2793. Available online: http://ir.itpcas.ac.cn/handle/131C11/7356 (accessed on 20 May 2021). [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Friedl, M.A.; Schaaf, C.B.; Strahler, A.H.; Hodges, J.C.F.; Gao, F.; Reed, B.C.; Huete, A. Monitoring vegetation phenology using MODIS. Remote Sens. Environ. 2003, 84, 471–475. [Google Scholar] [CrossRef]
- Zhang, X.; Friedl, M.A.; Schaaf, C.B. Global vegetation phenology from moderate resolution imaging spectroradiometer (MODIS): Evaluation of global patterns and comparison with in situ measurements. J. Geophys. Res. Biogeosci. 2006, 111, G04017. [Google Scholar] [CrossRef]
- Gonsamo, A.; Chen, J.M.; Wu, C.; Dragoni, D. Predicting deciduous forest carbon uptake phenology by upscaling FLUXNET measurements using remote sensing data. Agric. For. Meteorol. 2012, 165, 127–135. [Google Scholar] [CrossRef]
- Richardson, A.D.; Hufkens, K.; Milliman, T.; Aubrecht, D.M.; Frolking, S. Tracking vegetation phenology across diverse North American biomes using phenocam imagery. Sci. Data 2018, 5, 180028. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, J.; Gao, F.; Liu, Y.; Schaaf, C.; Friedl, M.; Yu, Y.; Jayavelu, S.; Gray, J.; Liu, L.; et al. Exploration of scaling effects on coarse resolution land surface phenology. Remote Sens. Environ. 2017, 190, 318–330. [Google Scholar] [CrossRef] [Green Version]
- Richardson, A.D.; Black, T.A.; Ciais, P.; Delbart, N.; Friedl, M.A.; Gobron, N.; Hollinger, D.Y.; Kutsch, W.L.; Longdoz, B.; Luyssaert, S.; et al. Influence of spring and autumn phenological transitions on forest ecosystem productivity. Philos. Trans. R. Soc. Lond. 2010, 365, 3227–3246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Odorico, P.; Gonsamo, A.; Gough, C.M.; Bohrer, G.; Morison, J.; Wilkinson, J.; Hanson, P.J.; Gianelle, D.; Fuentes, J.D.; Buchmann, N. The match and mismatch between photosynthesis and land surface phenology of deciduous forests. Agric. For. Meteorol. 2015, 214–215, 25–38. [Google Scholar] [CrossRef]
- Wang, X.; Xiao, J.; Li, X.; Cheng, G.; Ma, M.; Zhu, G.; Mehnaz, A.; Black, T.; Rachhpal, J. No trends in spring and autumn phenology during the global warming hiatus. Nat. Commun. 2019, 10, 2389. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Noormets, A. Standardized flux seasonality metrics: A companion dataset for FLUXNET annual product. Earth Syst. Sci. Data Discuss. 2020, 13, 1461–1475. [Google Scholar] [CrossRef]
- Gonsamo, A.; Chen, J.M.; D’Odorico, P. Deriving land surface phenology indicators from CO2 eddy covariance measurements. Ecol. Indic. 2013, 29, 203–207. [Google Scholar] [CrossRef]
- Wu, C.; Chen, J.M.; Gonsamo, A.; Price, D.T.; Black, T.A.; Kurz, W.A. Interannual variability of net carbon exchange is related to the lag between the end-dates of net carbon uptake and photosynthesis: Evidence from long records at two contrasting forest stands. Agric. For. Meteorol. 2012, 164, 29–38. [Google Scholar] [CrossRef]
- Balzarolo, M.; Vicca, S.; Nguy-Robertson, A.L.; Bonal, D.; Elbers, J.A.; Fu, Y.H.; Grünwald, T.; Horemans, J.A.; Papale, D.; Peñuelas, J.; et al. Matching the phenology of Net Ecosystem Exchange and vegetation indices estimated with MODIS and FLUXNET in-situ observations. Remote Sens. Environ. 2016, 174, 290–300. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.; Peng, D.; Soudani, K.; Siebicke, L.; Gough, C.M.; Arain, M.A.; Bohrer, G.; Lafleur, P.M.; Gonsamo, A.; Xu, S.; et al. Land surface phenology derived from normalized difference vegetation index (NDVI) at global FLUXNET sites. Agric. For. Meteorol. 2017, 233, 171–182. [Google Scholar] [CrossRef]
- Richardson, A.D.; Keenan, T.F.; Migliavacca, M.; Ryu, Y.; Sonnentag, O.; Toomey, M. Climate change, phenology, and phenological control of vegetation feedbacks to the climate system. Agric. For. Meteorol. 2013, 169, 156–173. [Google Scholar] [CrossRef]
- Gonsamo, A.; Croft, H.; Chen, J.M.; Wu, C.; Froelich, N.; Staebler, R.M. Radiation contributed more than temperature to increased decadal autumn and annual carbon uptake of two eastern North America mature forests. Agric. For. Meteorol. 2015, 201, 740–749. [Google Scholar] [CrossRef]
- Fu, Z.; Stoy, P.C.; Luo, Y.; Chen, J.; Sun, J.; Montagnani, L.; Wohlfahrt, G.; Rahman, F.; Rambal, S.; Bernhoferet, C.; et al. Climate controls over the net carbon uptake period and amplitude of net ecosystem production in temperate and boreal ecosystems. Agric. For. Meteorol. 2017, 243, 9–18. [Google Scholar] [CrossRef] [Green Version]
- Garrity, S.R.; Bohrer, G.; Maurer, K.D.; Mueller, K.L.; Vogel, C.S.; Curtis, P.S. A comparison of multiple phenology data sources for estimating seasonal transitions in deciduous forest carbon exchange. Agric. For. Meteorol. 2011, 151, 1741–1752. [Google Scholar] [CrossRef]
- Chen, B.; Che, M. Improving vegetation phenological parameterization of, a land surface model. Biogeosc. Discuss. 2016. preprint. [Google Scholar] [CrossRef]
- Peng, B.; Guan, K.; Chen, M.; Lawrence, D.M.; Pokhrel, Y.; Suyker, A.; Arkebauer, T.; Lu, Q. Improving maize growth processes in the community land model: Implementation and evaluation. Agric. For. Meteorol. 2018, 250–251, 64–89. [Google Scholar] [CrossRef]
- Liu, Y.; Wu, C.; Peng, D.; Xu, S.; Gonsamo, A.; Jassal, R.S.; Arain, M.A.; Lu, L.; Fang, B.; Chen, M. Improved modeling of land surface phenology using MODIS land surface reflectance and temperature at evergreen needleleaf forests of central North America. Remote Sens. Environ. 2016, 176, 152–162. [Google Scholar] [CrossRef]
- Reichstein, M.; Falge, E.; Baldocchi, D.; Papale, D.; Valentini, R. On the Separation of Net Ecosystem Exchange into Assimilation and Ecosystem Respiration: Review and Improved Algorithm. Glob. Chang. Biol. 2005, 11, 1424–1439. [Google Scholar] [CrossRef]
- Papale, D.; Reichstein, M.; Aubinet, M.; Canfora, E.; Bernhofer, C.; Kutsch, W.; Longdoz, B.; Rambal, S.; Valentini, R.; Vesala, T.; et al. Towards a standardized processing of Net Ecosystem Exchange measured with eddy covariance technique: Algorithms and uncertainty estimation. Biogeosciences 2006, 3, 571–583. [Google Scholar] [CrossRef] [Green Version]
- Lasslop, G.; Reichstein, M.; Papale, D.; Richardson, A.D.; Arneth, A.; Barr, A.; Paul, S.; Gerog, W. Separation of net ecosystem exchange into assimilation and respiration using a light response curve approach: Critical issues and global evaluation. Glob. Chang. Biol. 2010, 16, 187–208. [Google Scholar] [CrossRef] [Green Version]
- Belward, A.S.; Estes, J.E.; Kline, K.D. The IGBP-DIS 1-Km Land-Cover Data Set DISCover: A Project Overview. Photogramm. Eng. Remote Sens. 1999, 65, 1013–1020. [Google Scholar]
- Chen, J.; Jönsson, P.; Tamura, M.; Gu, Z.; Matsushita, B.; Eklundh, L. A simple method for reconstructing a high-quality NDVI time-series data set based on the Savitzky-Golay filter. Remote Sens. Environ. 2004, 91, 332–344. [Google Scholar] [CrossRef]
- Pan, Z.K.; Huang, J.F.; Zhou, Q.B.; Wang, L.M.; Cheng, Y.X.; Zhang, H.K.; Blackburn, G.A.; Yan, J.; Liu, J.H. Mapping Crop Phenology Using NDVI Time-Series Derived from HJ-1 A/B Data. Int. J. Appl. Earth Obs. Geoinf. 2015, 34, 188–197. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Zhou, G.; Du, H.; Mao, F.; Xu, L.; Li, X.; Liu, L. Combined MODIS land surface temperature and greenness data for modeling vegetation phenology, physiology, and gross primary production in terrestrial ecosystems. Sci. Total Environ. 2020, 726, 137948. [Google Scholar] [CrossRef]
- Zhou, S.; Zhang, Y.; Caylor, K.K.; Luo, Y.; Xiao, X.; Ciais, P.; Huang, Y.; Wang, G. Explaining inter-annual variability of gross primary productivity from plant phenology and physiology. Agric. For. Meteorol. 2016, 226–227, 246–256. [Google Scholar] [CrossRef] [Green Version]
- Cong, N.; Wang, T.; Nan, H.; Ma, Y.; Wang, X.; Myneni, R.B.; Piao, S. Changes in satellite-derived spring vegetation green-up date and its linkage to climate in china from 1982 to 2010: A multimethod analysis. Glob. Chang. Biol. 2013, 19, 881–891. [Google Scholar] [CrossRef]
- Gray, J.; Sulla-Menashe, D.; Friedl, M. MCD12Q2 MODIS/Terra+Aqua Land Cover Dynamics Yearly L3 Global 500m SIN Grid V006; NASA EOSDIS Land Processes DAAC: Sioux Falls, SD, USA, 2019. [CrossRef]
- Sims, D.A.; Rahman, A.F.; Cordova, V.D.; El-Masri, B.Z.; Baldocchi, D.D.; Flanagan, L.B.; Goldstein, A.H.; Hollinger, D.Y.; Misson, L.; Monson, R.K.; et al. On the use of MODIS EVI to assess gross primary productivity of North American ecosystems. J. Geophys. Res. 2006, 111, G04015. [Google Scholar] [CrossRef] [Green Version]
- Sims, D.A.; Rahman, A.F.; Cordova, V.D.; El-Masri, B.Z.; Baldocchi, D.D.; Bolstad, P.V.; Flanagan, L.B.; Goldstein, A.H.; Hollinger, D.Y.; Misson, L.; et al. A new model of gross primary productivity for North American ecosystems based solely on the enhanced vegetation index and land surface temperature from MODIS. Remote Sens. Environ. 2008, 112, 1633–1646. [Google Scholar] [CrossRef]
- Didan, K. MOD13Q1 MODIS/Terra Vegetation Indices 16-Day L3 Global 250m SIN Grid; V006; NASA EOSDIS LP DAAC: Sioux Falls, SD, USA, 2015.
- Wan, Z.; Hook, S.; Hulley, G. MOD11A2 MODIS/Terra Land Surface Temperature/Emissivity 8-Day L3 Global 1 km SIN Grid V006; NASA EOSDIS LP DAAC: Sioux Falls, SD, USA, 2015. [CrossRef]
- Zhang, Y.; Xiao, X.; Wu, X.; Zhou, S.; Zhang, G.; Qin, Y.; Dong, J. A global moderate resolution dataset of gross primary production of vegetation for 2000–2016. Sci. Data 2017, 4, 170165. [Google Scholar] [CrossRef] [Green Version]
- Shabanov, N.V.; Zhou, L.; Knyazikhin, Y.; Myneni, R.B.; Tucker, C.J. Analysis of interannual changes in northern vegetation activity observed in AVHRR data from 1981 to 1994. IEEE Trans. Geosci. Remote Sens. 2002, 40, 115–130. [Google Scholar] [CrossRef] [Green Version]
- Dye, D.G.; Tucker, C.J. Seasonality and trends of snow-cover, vegetation index, and temperature in northern Eurasia. Geophys. Res. Lett. 2003, 30, 1405. [Google Scholar] [CrossRef]
- Böttcher, K.; Kervinen, M.; Aurela, M.; Mattila, O.-P.; Markkanen, T.; Pullianinen, J. Monitoring spring phenology of boreal coniferous forest in Finland using MODIS time-series. In Proceedings of the 31st EARSeL Symposium of Remote Sensing and Geoinformation Not Only for Scientific Cooperation, Prague, Czech Republic, 30 May–2 June 2011; pp. 125–134, ISBN 978-80-01-04868-9. [Google Scholar]
- Zhu, W.Q.; Tian, H.Q.; Xu, X.F.; Pan, Y.Z.; Chen, G.S.; Lin, W.P. Extension of the growing season due to delayed autumn over mid and high latitudes in North America during 1982–2006. Glob. Ecol. Biogeogr. 2012, 21, 260–271. [Google Scholar] [CrossRef]
- D’Odorico, P.; Gonsamo, A.; Damm, A.; Schaepman, M.E. Experimental evaluation of sentinel-2 spectral response functions for NDVI time-series continuity. IEEE Trans. Geosci. Remote Sens. 2013, 51, 1336–1348. [Google Scholar] [CrossRef]
- Kobayashi, H.; Yunus, A.P.; Nagai, S.; Sugiura, K.; Kim, Y.; Dam, B.V.; Nagano, H.; Zona, D.; Harazono, Y.; Bret-Harte, M.S.; et al. Latitudinal gradient of spruce forest understory and tundra phenology in Alaska as observed from satellite and ground-based data. Remote Sens. Environ. 2016, 177, 160–170. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.L.; Huang, Y.F.; Wang, G.Q. Response of vegetation carbon uptake to snow-induced phenological and physiological changes across temperate China. Sci. Total Environ. 2019, 692, 188–200. [Google Scholar] [CrossRef]
- Ganguly, S.; Friedl, M.A.; Tan, B.; Zhang, X.; Verma, M. Land surface phenology from MODIS: Characterization of the Collection 5 global land cover dynamics product. Remote Sens. Environ. 2010, 114, 1805–1816. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Shen, R.; Wang, Y.; Li, X.; Liu, S.; Liang, S.; Chen, J.M.; Ju, W.; Zhang, L.; Yuan, W. Improved estimate of global gross primary production for reproducing its long-term variation, 1982–2017. Earth Syst. Sci. Data Discuss. 2019, 12, 2725–2746. [Google Scholar] [CrossRef]
- Melaas, E.K.; Sulla-Menashe, D.; Gray, J.M.; Black, T.A.; Friedl, M.A. Multisite analysis of land surface phenology in North American temperate and boreal deciduous forests from Landsat. Remote Sens. Environ. 2016, 186, 452–464. [Google Scholar] [CrossRef]
- Zhang, Y.; Xiao, X.; Zhou, S.; Ciais, P.; McCarthy, H.; Luo, Y. Canopy and physiological limitation of GPP during drought and heat wave. Geophys. Res. Lett. 2016, 43, 3325–3333. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Hill, M.J.; Zhang, X.; Wang, Z.; Richardson, A.D.; Hufkens, K.; Filippa, G.; Baldocchi, D.D.; Ma, S.; Verfaillie, J.; et al. Using data from Landsat, MODIS, VIIRS and PhenoCams to monitor the phenology of California oak/grass savanna and open grassland across spatial scales. Agric. For. Meteorol. 2017, 237–238, 311–325. [Google Scholar] [CrossRef]
- Ma, S.; Baldocchi, D.D.; Xu, L.; Hehn, T. Inter-annual variability in carbon dioxide exchange of an oak/grass savanna and open grassland in California. Agric. For. Meteorol. 2007, 147, 157–171. [Google Scholar] [CrossRef]
- Huete, A.; Didan, K.; Miura, T.; Rodriguez, E.P.; Gao, X.; Ferreira, L.G. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar] [CrossRef]
- Wu, W.; Shibasaki, R.; Yang, P.; Zhou, Q.; Tang, H. Characterizing spatial patterns of phenology in China’s cropland based on remotely sensed data. In Proceedings of the 2008 International Workshop on Earth Observation and Remote Sensing Applications, Beijing, China, 30 June–2 July 2008; pp. 1–6. [Google Scholar] [CrossRef]
- Dash, J.; Jeganathan, C.; Atkinson, P.M. The use of MERIS Terrestrial Chlorophyll index to study spatio-temporal variation in vegetation phenology over India. Remote Sens. Environ. 2010, 114, 1388–1402. [Google Scholar] [CrossRef]
- Adole, T.; Dash, J.; Atkinson, P.M. Characterising the land surface phenology of Africa using 500 m MODIS EVI. Appl. Geogr. 2018, 90, 187–199. [Google Scholar] [CrossRef] [Green Version]
- Niinemets, ü. Responses of forest trees to single and multiple environmental stresses from seedlings to mature plants: Past stress history, stress interactions, tolerance and acclimation. For. Ecol. Manag. 2010, 260, 1623–1639. [Google Scholar] [CrossRef]
- Teuling, A.J.; Seneviratne, S.I.; Stöckli, R.; Reichstein, M.; Moors, E.; Ciais, P.; Luyssaert, S.; Hurk, B.D.; Ammann, C.; Bernhofer, C.; et al. Contrasting response of European forest and grassland energy exchange to heatwaves. Nat. Geosci. 2010, 3, 722–727. [Google Scholar] [CrossRef]
- Shen, M.; Tang, Y.; Desai, A.R.; Gough, C.; Chen, J. Can derived land-surface phenology be used as a surrogate for phenology of canopy photosynthesis? Int. J. Remote Sens. 2014, 35, 1162–1174. [Google Scholar] [CrossRef]
- Ahrends, H.E.; Etzold, S.; Kutsch, W.L.; Stoeckli, R.; Eugster, W. Tree phenology and carbon dioxide fluxes: Use of digital photography for process-based interpretation at the ecosystem scale. Clim. Res. 2009, 39, 261–274. [Google Scholar] [CrossRef] [Green Version]
- Lang, W.; Chen, X.; Qian, S.; Liu, G.; Piao, S. A new process-based model for predicting autumn phenology: How is leaf senescence controlled by photoperiod and temperature coupling? Agric. For. Meteorol. 2019, 268, 124–135. [Google Scholar] [CrossRef]
- Wilson, K.B.; Baldocchi, D.D.; Hanson, P.J. Leaf age affects the seasonal pattern of photosynthetic capacity and net ecosystem exchange of carbon in a deciduous forest. Plant Cell Environ. 2001, 24, 571–583. [Google Scholar] [CrossRef]
- Knohl, A.; Schulze, E.D.; Kolle, O.; Buchmann, N. Large carbon uptake by an unmanaged 250-year-old deciduous forest in Central Germany. Agric. For. Meteorol. 2003, 118, 151–167. [Google Scholar] [CrossRef]
- Keel, S.G.; Schädel, C. Expanding leaves of mature deciduous forest trees rapidly become autotrophic. Tree Physiol. 2010, 30, 1253–1259. [Google Scholar] [CrossRef] [Green Version]
- Koike, T.; Kitao, M.; Maruyama, Y.; Mori, S.; Lei, T.T. Leaf morphology and photosynthetic adjustments among deciduous broad-leaved trees within the vertical canopy profile. Tree Physiol. 2001, 21, 951–958. [Google Scholar] [CrossRef] [PubMed]
- Melaas, E.K.; Friedl, M.A.; Zhu, Z. Detecting interannual variation in deciduous broadleaf forest phenology using Landsat TM/ETM+ data. Remote Sens. Environ. 2013, 132, 176–185. [Google Scholar] [CrossRef]
- Chang, Q.; Xiao, X.; Jiao, W.; Wu, X.; Doughty, R.; Wang, J.; Du, L.; Zou, Z.; Qin, Y. Assessing consistency of spring phenology of snow-covered forests as estimated by vegetation indices, gross primary production, and solar-induced chlorophyll fluorescence. Agric. For. Meteorol. 2019, 275, 305–316. [Google Scholar] [CrossRef]
- Park, H.; Su, J.J.; Chang, H.H.; Park, C.E.; Kim, J. Slowdown of spring green-up advancements in boreal forests. Remote Sens. Environ. 2018, 217, 191–202. [Google Scholar] [CrossRef]
- Filippa, G.; Cremonese, E.; Galvagno, M.; Isabellon, M.; Bayle, A.; Choler, P.; Carlson, B.Z.; Gabellani, S.; Cella, U.M.; Migliavacca, M. Climatic Drivers of Greening Trends in the Alps. Remote Sens. 2019, 11, 2527. [Google Scholar] [CrossRef] [Green Version]
- Monson, R.K.; Sparks, J.P.; Rosenstiel, T.N.; Scott-Denton, L.E.; Huxman, T.E.; Harley, P.C.; Turnipseed, A.A.; Burns, S.P.; Backlund, B.; Hu, J. Climatic influences on net ecosystem CO2 exchange during the transition from wintertime carbon source to springtime carbon sink in a high-elevation, subalpine forest. Oecologia 2005, 146, 130–147. [Google Scholar] [CrossRef]
- Hmimina, G.; Dufrêne, E.; Pontailler, J.Y.; Delpierre, N.; Soudani, K. Evaluation of the potential of MODIS satellite data to predict vegetation phenology in different biomes: An investigation using ground-based NDVI measurements. Remote Sens. Environ. 2013, 132, 145–158. [Google Scholar] [CrossRef]
- Du, Q.; Liu, H.Z.; Li, Y.H.; Xu, L.J.; Diloksumpun, S. The effect of phenology on the carbon exchange process in grassland and maize cropland ecosystems across a semiarid area of China. Sci. Total Environ. 2019, 695, 133868. [Google Scholar] [CrossRef] [PubMed]
- Yan, D.; Zhang, X.; Nagai, S.; Yu, Y.; Akitsu, T.; Nasahara, K.N.; Ide, R.; Maeda, T. Evaluating land surface phenology from the advanced himawari imager using observations from modis and the phenological eyes network. Int. J. Appl. Earth Obs. 2019, 79, 71–83. [Google Scholar] [CrossRef]
- Dong, J.; Xiao, X.; Wagle, P.; Zhang, G.; Zhou, Y.; Jin, C.; Torn, M.S.; Meyers, T.P.; Suyker, A.E.; Wang, J.; et al. Comparison of four evi-based models for estimating gross primary production of maize and soybean croplands and tallgrass prairie under severe drought. Remote Sens. Environ. 2015, 162, 154–168. [Google Scholar] [CrossRef] [Green Version]
- Xin, F.; Xiao, X.; Zhao, B.; Miyata, A.; Baldocchi, D.; Knox, S.; Kang, M.; Shim, K.; Min, S.; Chen, B.; et al. Modeling gross primary production of paddy rice cropland through analyses of data from CO2 eddy flux tower sites and MODIS images. Remote Sens. Environ. 2017, 190, 42–55. [Google Scholar] [CrossRef] [Green Version]
- Running, S.W.; Nemani, R.R.; Heinsch, F.A.; Zhao, M.S.; Reeves, M.; Hashimoto, H. A continuous satellite-derived measure of global terrestrial primary production. Bioscience 2004, 54, 547–560. [Google Scholar] [CrossRef]









| PFT | SOG | EOG | ||||
|---|---|---|---|---|---|---|
| R2 | Bias (Days) | RMSE (Days) | R2 | Bias (Days) | RMSE (Days) | |
| ENF | 0.72 | 0.7 | 15.4 | 0.65 | 0.2 | 12.0 |
| DBF | 0.85 | 0.0 | 6.6 | 0.77 | 0.1 | 7.5 |
| MF | 0.82 | 0.0 | 11.5 | 0.59 | 0.0 | 10.3 |
| All | 0.80 | 0.4 | 12.7 | 0.70 | 0.2 | 10.5 |
| PFT | SOG | EOG | ||||
|---|---|---|---|---|---|---|
| R2 | Bias (Days) | RMSE (Days) | R2 | Bias (Days) | RMSE (Days) | |
| OSH + CSH | 0.79 | 2.1 | 23.0 | 0.24 | 4.5 | 39.0 |
| SAV + WSA | 0.73 | −0.9 | 25.2 | 0.68 | 4.7 | 27.5 |
| GRA | 0.71 | 0.7 | 19.3 | 0.59 | −0.1 | 13.4 |
| CRO | 0.90 | −3.4 | 15.6 | 0.53 | 1.4 | 25.4 |
| WET | 0.60 | 1.0 | 18.3 | 0.58 | −1.2 | 16.5 |
| ALL | 0.78 | −0.5 | 19.4 | 0.64 | 1.1 | 22.9 |
| PFT | R | Bias (Days) | RMSD (Days) | RMSDb (Days) | N | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | |
| ENF | 0.74 | 0.12 | −10.60 | 4.82 | 16.04 | 3.27 | 11.51 | 0.91 | 8936 | 2389 |
| DNF | 0.40 | 0.12 | −5.94 | 3.15 | 9.73 | 2.03 | 7.21 | 1.44 | 4255 | 1256 |
| DBF | 0.84 | 0.04 | 4.95 | 1.77 | 9.79 | 0.71 | 8.28 | 0.54 | 16,160 | 4168 |
| MF | 0.78 | 0.12 | −1.02 | 5.45 | 19.52 | 2.12 | 18.74 | 2.42 | 20,662 | 5399 |
| OSH | 0.10 | 0.12 | 5.75 | 6.17 | 17.86 | 2.50 | 15.74 | 3.03 | 29,941 | 7881 |
| SAV | 0.84 | 0.03 | −4.34 | 3.84 | 13.94 | 1.67 | 12.76 | 1.20 | 61,135 | 15,842 |
| WSA | 0.87 | 0.04 | 0.39 | 3.29 | 16.51 | 2.00 | 16.19 | 2.05 | 63,596 | 16,477 |
| GRA | 0.74 | 0.04 | −12.30 | 2.01 | 25.47 | 2.66 | 22.23 | 2.54 | 25,361 | 6633 |
| WET | 0.76 | 0.09 | 9.83 | 8.43 | 15.64 | 5.37 | 10.19 | 2.22 | 5391 | 1438 |
| CRO | 0.68 | 0.07 | −0.32 | 3.65 | 23.87 | 2.55 | 23.60 | 2.61 | 34,867 | 9216 |
| URB | 0.66 | 0.07 | −5.55 | 2.91 | 18.21 | 1.79 | 17.13 | 1.58 | 2550 | 661 |
| CNV | 0.63 | 0.10 | −1.26 | 3.11 | 16.12 | 2.61 | 15.77 | 2.72 | 3156 | 870 |
| PFT | R | Bias (Days) | RMSD (Days) | RMSDb (Days) | N | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | |
| ENF | 0.26 | 0.10 | 9.07 | 7.04 | 17.84 | 3.65 | 14.08 | 2.12 | 8936 | 2389 |
| DNF | 0.50 | 0.16 | −17.10 | 1.62 | 17.88 | 1.57 | 5.16 | 0.60 | 4255 | 1256 |
| DBF | 0.69 | 0.10 | −5.14 | 2.85 | 11.41 | 1.67 | 9.83 | 1.48 | 16,160 | 4168 |
| MF | 0.69 | 0.14 | −7.45 | 3.69 | 15.29 | 3.34 | 13.04 | 2.49 | 20,662 | 5399 |
| OSH | 0.12 | 0.11 | 0.33 | 4.46 | 19.63 | 4.02 | 19.02 | 4.62 | 29,941 | 7881 |
| SAV | 0.28 | 0.16 | −2.35 | 6.64 | 23.98 | 2.85 | 23.02 | 2.53 | 61,135 | 15,842 |
| WSA | 0.67 | 0.10 | 0.94 | 4.55 | 25.63 | 3.46 | 25.21 | 3.61 | 63,596 | 16,477 |
| GRA | 0.73 | 0.04 | −1.23 | 2.38 | 17.47 | 1.31 | 17.28 | 1.36 | 25,361 | 6633 |
| WET | 0.50 | 0.13 | −32.86 | 7.12 | 36.86 | 7.17 | 16.56 | 2.34 | 5391 | 1438 |
| CRO | −0.33 | 0.12 | 2.73 | 7.84 | 61.62 | 6.24 | 61.07 | 6.45 | 34,867 | 9216 |
| URB | 0.31 | 0.17 | −4.09 | 4.01 | 31.36 | 3.81 | 30.80 | 4.22 | 2550 | 661 |
| CNV | 0.17 | 0.13 | −45.94 | 6.71 | 55.04 | 3.82 | 29.19 | 6.37 | 3156 | 870 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, X.; Tang, Y.; Qu, Y.; Zhou, Z.; Hu, J. Global Vegetation Photosynthetic Phenology Products Based on MODIS Vegetation Greenness and Temperature: Modeling and Evaluation. Remote Sens. 2021, 13, 5080. https://doi.org/10.3390/rs13245080
Xu X, Tang Y, Qu Y, Zhou Z, Hu J. Global Vegetation Photosynthetic Phenology Products Based on MODIS Vegetation Greenness and Temperature: Modeling and Evaluation. Remote Sensing. 2021; 13(24):5080. https://doi.org/10.3390/rs13245080
Chicago/Turabian StyleXu, Xiaojun, Yan Tang, Yiling Qu, Zhongsheng Zhou, and Junguo Hu. 2021. "Global Vegetation Photosynthetic Phenology Products Based on MODIS Vegetation Greenness and Temperature: Modeling and Evaluation" Remote Sensing 13, no. 24: 5080. https://doi.org/10.3390/rs13245080
APA StyleXu, X., Tang, Y., Qu, Y., Zhou, Z., & Hu, J. (2021). Global Vegetation Photosynthetic Phenology Products Based on MODIS Vegetation Greenness and Temperature: Modeling and Evaluation. Remote Sensing, 13(24), 5080. https://doi.org/10.3390/rs13245080