
Integrated Fire Management as a Renewing Agent of Native Vegetation and Inhibitor of Invasive Plants in Vereda Habitats: Diagnosis by Remotely Piloted Aircraft Systems


Abstract
:1. Introduction
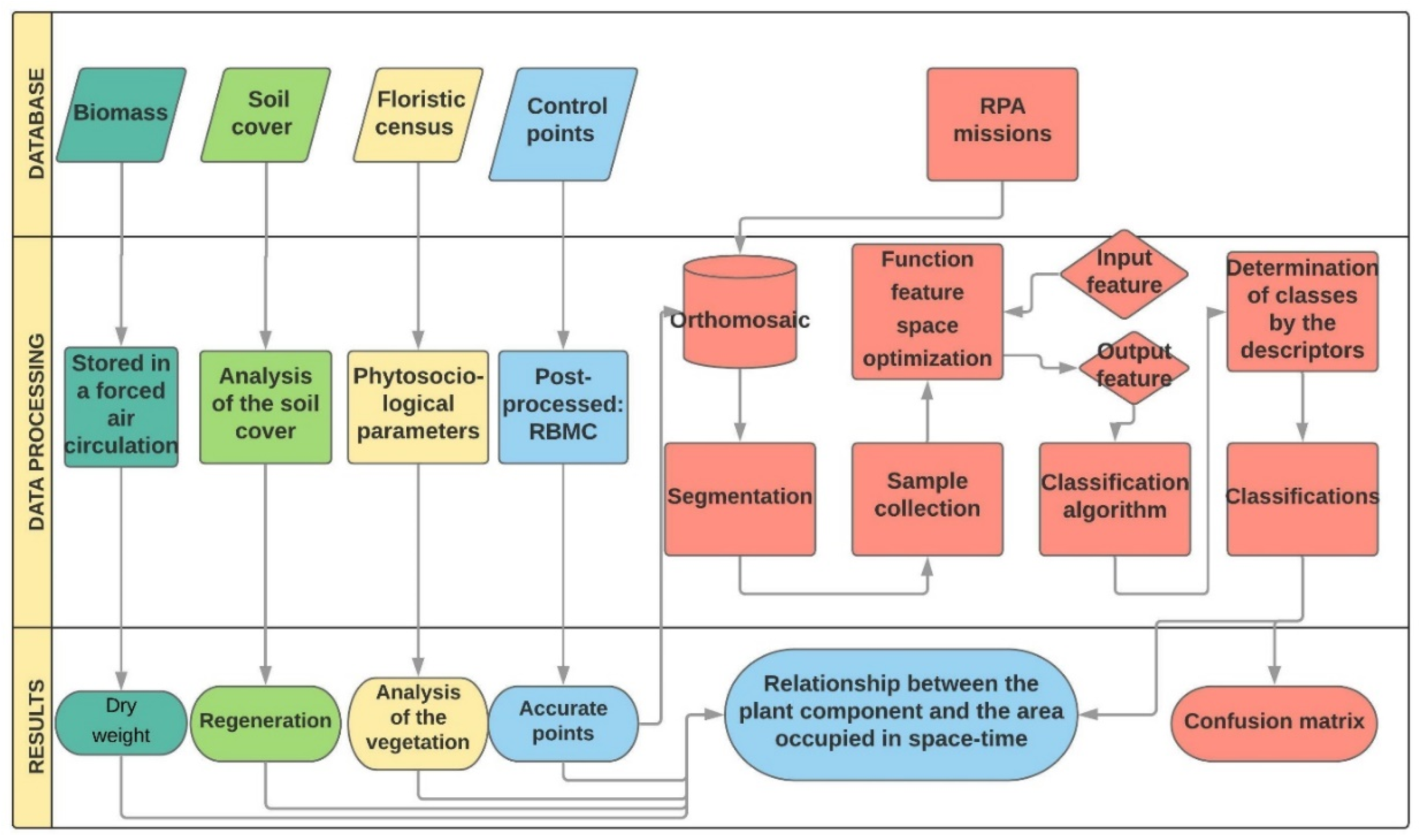
2. Materials and Methods
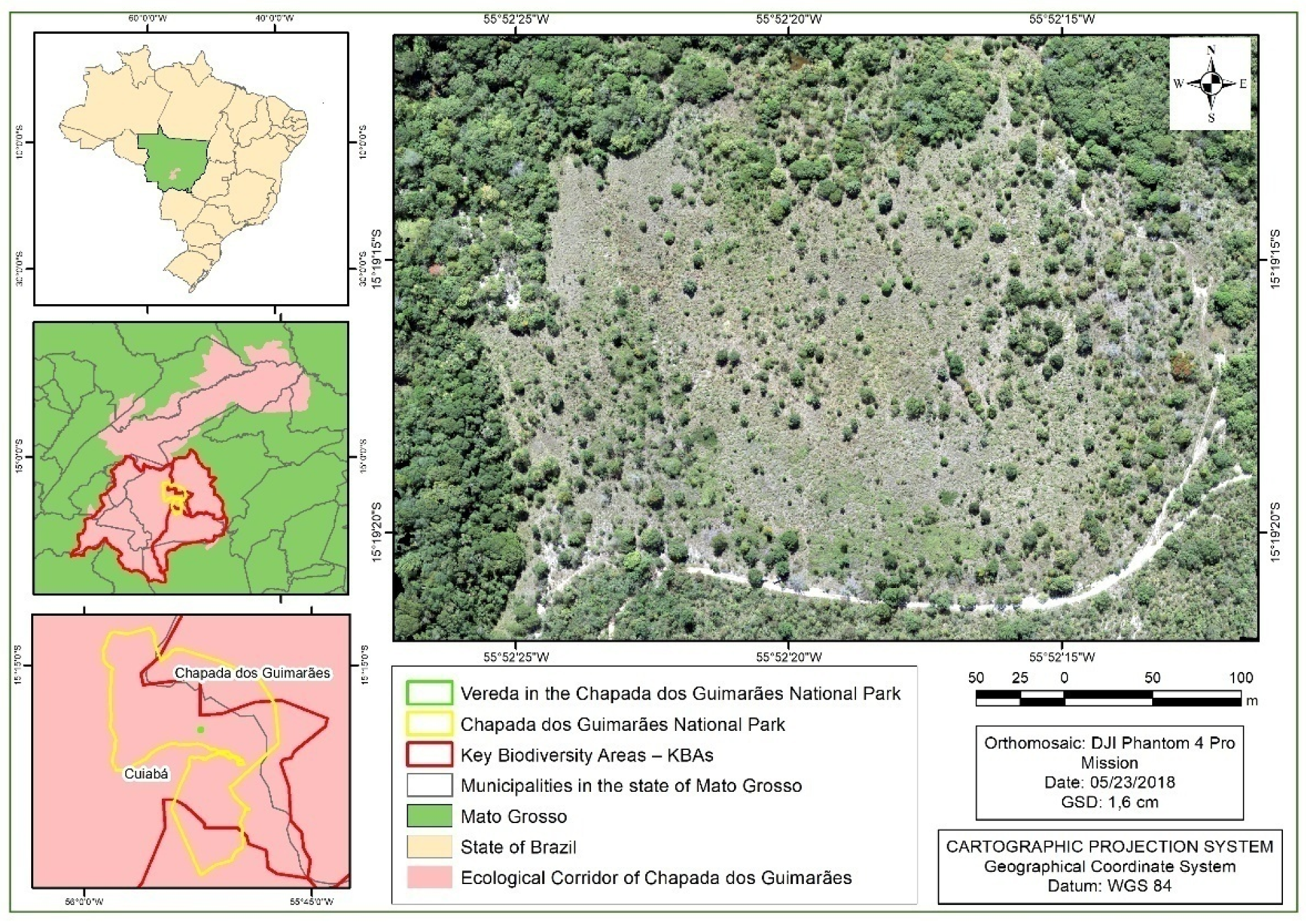
2.1. Study Area
2.2. Database
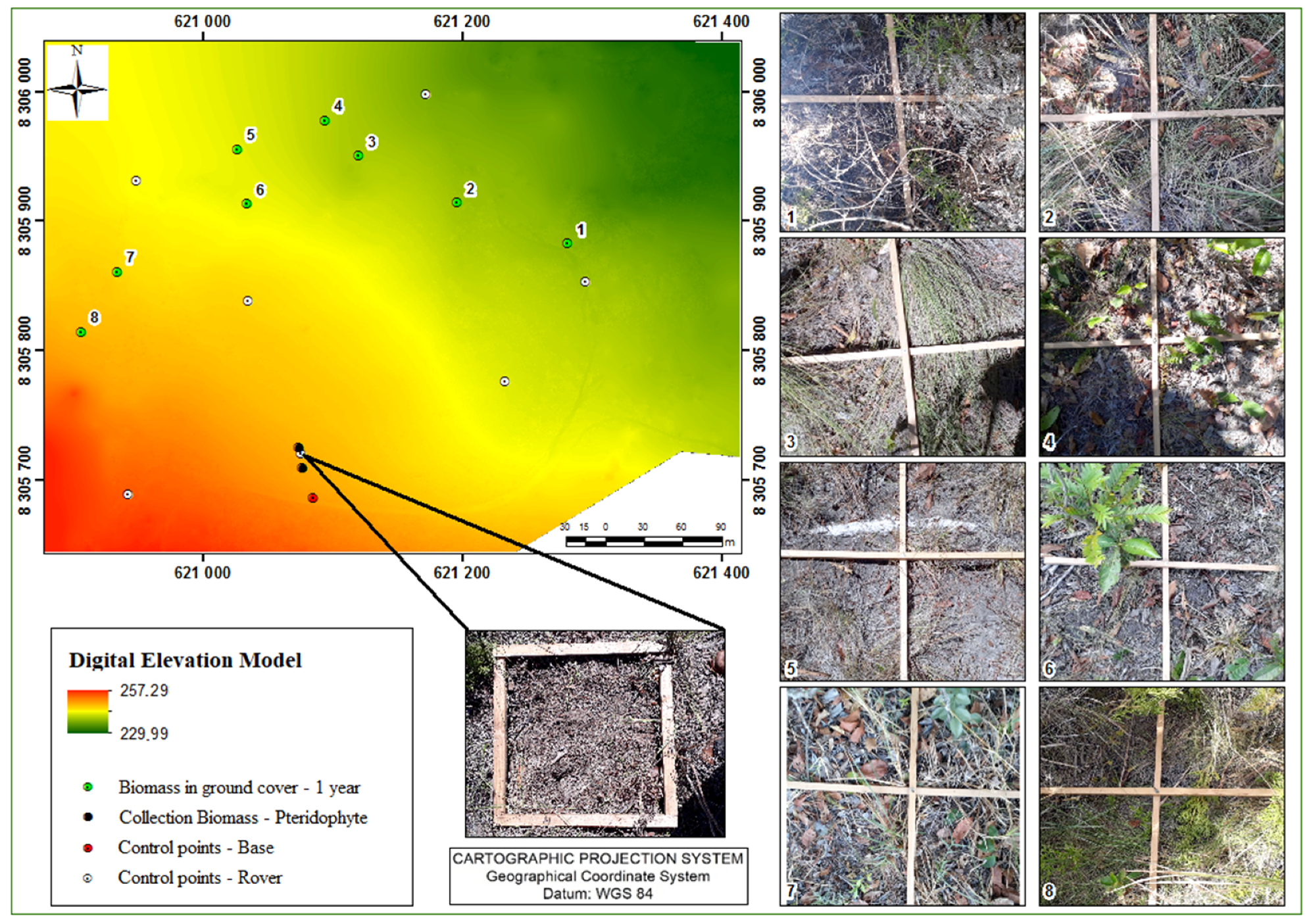
2.2.1. Vegetable Aspects
2.2.2. Aerophotogrammetry
2.3. Data Prossessing
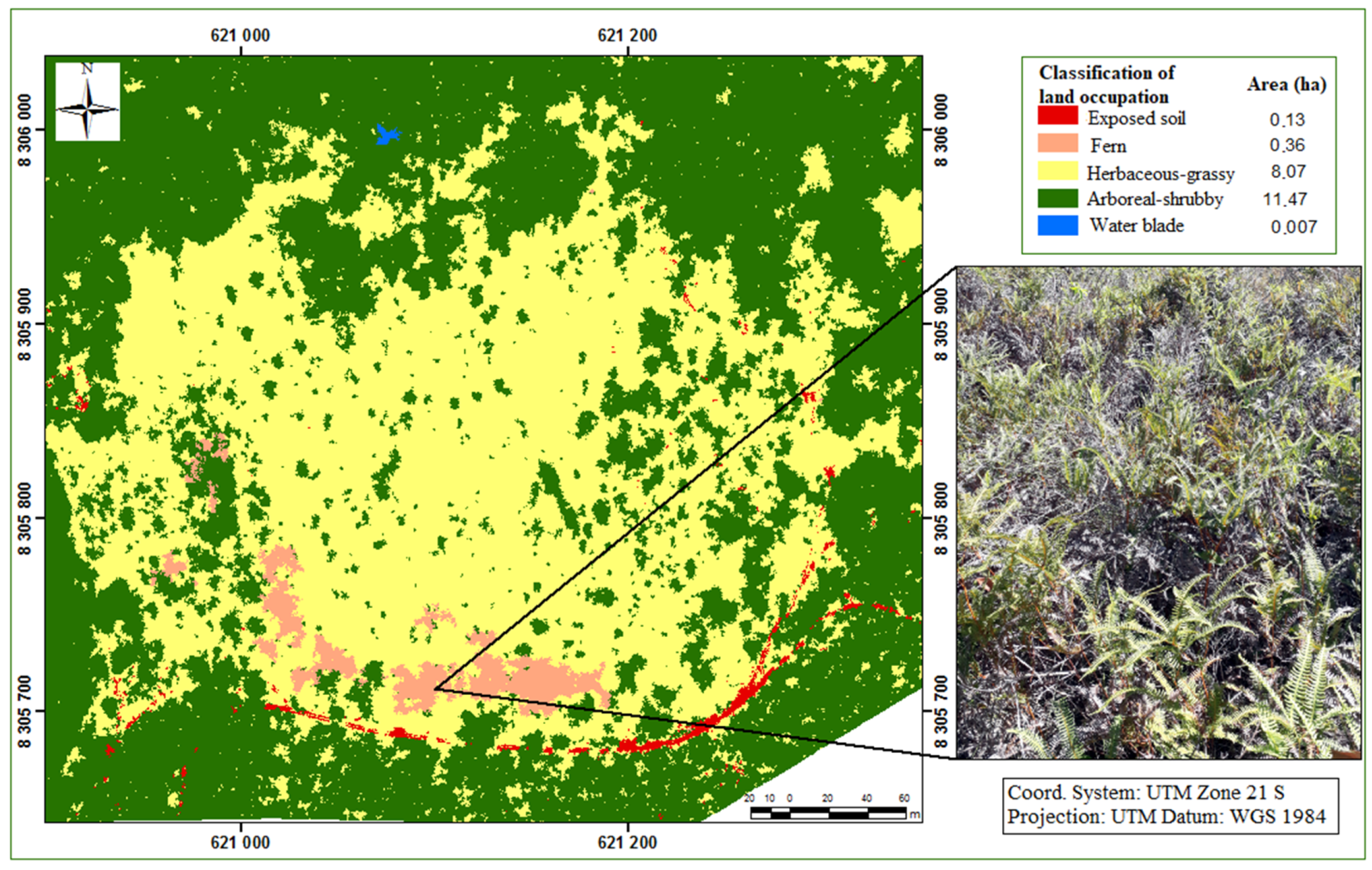
2.3.1. Vegetation Aspects
2.3.2. Aerophotogrammetry
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Leão, T.C.C.; Almeida, W.R.; Dechoum, M.; Ziller, S.R. Espécies Exóticas Invasoras no Nordeste do Brasil: Contextualização, Manejo e Políticas Públicas; Centro de Pesquisas Ambientais do Nordeste e Instituto Hórus de Desenvolvimento e Conservação Ambiental: Recife, Brazil, 2011; Volume 1, 99p. [Google Scholar]
- Richardson, D.; Pysek, P.; Rejmanek, M.; Barbour, M.G.; Panetta, F.D.; West, C.J. Naturalization and invasion of alien plants: Concepts and definitions. Divers. Distrib. 2000, 6, 93–107. [Google Scholar] [CrossRef]
- Archer, S.R.; Andersen, E.M.; Predick, K.I.; Schwinning, S.; Steidl, R.J.; Woods, S.R. Woody Plant Encroachment: Causes and Consequences. In Rangeland Systems: Processes, Management and Challenges; Briske, D.D., Ed.; Springer International Publishing: Cham, Switzerland, 2017; pp. 25–84. [Google Scholar] [CrossRef] [Green Version]
- Lehmann, D.R.M. Estudos sobre a propagação de Gleichenella pectinata (Willd.) ching (pteridófita—Gleicheniaceae). In Dissertação (Mestrado em Biologia Vegetal); Universidade Federal de Santa Catarina: Florianópolis, Brazil, 2008; 54p. [Google Scholar]
- Arantes, A.A.; Prado, J.; Ranal, M.A. Monilófitas da Estação Ecológica do Panga, Uberlândia, Estado de Minas Gerais, Brasil: Dennstaedtiaceae, Dryopteridaceae, Gleicheniaceae, Hymenophyllaceae e Lindsaeaceae. Hoehnea 2008, 35, 367–378. [Google Scholar] [CrossRef] [Green Version]
- Melo, L.C.; Salino, A. Pteridófitas de duas áreas de floresta da bacia do Rio Doce no estado de Minas Gerais, Brasil. Lundiana Int. J. Biodivers. 2002, 3, 129–139. [Google Scholar] [CrossRef]
- Lima, L.V.; Salino, A. Gleicheniaceae in Flora do Brasil 2019 em Construção. Jardim Botânico do Rio de Janeiro. Available online: http://floradobrasil.jbrj.gov.br/reflora/floradobrasil/FB91164 (accessed on 18 February 2020).
- Windisch, P.G. Pteridófitas do Estado de Mato Grosso: Gleicheniaceae. BRADEA—Bol. Do Herb. Bradeanum 1994, 6, 304–311. [Google Scholar]
- Carvalho, A.C.A. Análise ambiental de um subsistema de vereda em unidade de conservação: Reserva Ecológica do IBGE- RECOR e Estação Ecológica de Águas emendadas ESECAE/DF Brasília/DF. Master’s Thesis, Universidade de Brasília, Distrito Federal, Brasilia, Brazil, 2015; 127p. [Google Scholar] [CrossRef] [Green Version]
- BRASIL. Lei nº 12.651, De 25 De Maio De 2012. Available online: http://www.planalto.gov.br/ccivil_03/_ato2011-2014/2012/lei/l12651.htm (accessed on 15 February 2022).
- Borges, S.L.; Eloy, L.; Schmidt, I.B.; Barradas, A.C.S.; Dos Santos, I.A. Fire Management in Veredas (Palm Swamps): New Perspectives on Traditional Farming Systems in Jalapão, Brazil. Ambient. Soc. 2016, 19, 269–294. [Google Scholar] [CrossRef] [Green Version]
- Miguel, E.P.; Rezende, A.V.; Leal, F.A.; Matricardi, E.A.T.; Vale, A.T.D.; Pereira, R.S. Redes neurais artificiais para a modelagem do volume de madeira e biomassa do cerradão com dados de satélite. Pesqui. Agropecuária Bras. 2015, 50, 829–839. [Google Scholar] [CrossRef] [Green Version]
- Alba, E.; Mello, E.P.; Marchesan, J.; Silva, E.A.; Tramontina, J.; Pereira, R.S. Spectral characterization of forest plantations with Landsat 8/OLI images for forest planning and management. Pesqui. Agropecuária Bras. 2017, 52, 1072–1079. [Google Scholar] [CrossRef] [Green Version]
- De Almeida, A.Q.; De Mello, A.A.; Neto, A.L.D.; Ferraz, R.C. Relações empíricas entre características dendrométricas da Caatinga brasileira e dados TM Landsat 5. Pesqui. Agropecuária Bras. 2014, 49, 306–315. [Google Scholar] [CrossRef] [Green Version]
- Furtado, L.F.D.A.; Silva, T.S.F.; Fernandes, P.J.F.; Novo, E.M.L.D.M. Land cover classification of Lago Grande de Curuai floodplain (Amazon, Brazil) using multi-sensor and image fusion techniques. Acta Amaz. 2015, 45, 195–202. [Google Scholar] [CrossRef] [Green Version]
- Kurtz, B.C.; Magalhães, A.M.; Seabra, V.D.S.; Richter, M.; Caris, E.A.P. Integrating remote sensing and phytosociology of the Atlantic Forest to map a small continental island in southeastern Brazil: Subsidies to protect the habitat of critically endangered species. Rodriguésia 2018, 69, 2081–2092. [Google Scholar] [CrossRef] [Green Version]
- Alvarado, S.T.; Fornazari, T.; Costola, A.; Morellato, P.; Silva, T. Drivers of fire occurrence in a mountainous Brazilian cerrado savanna: Tracking long-term fire regimes using remote sensing. Ecol. Indic. 2017, 78, 270–281. [Google Scholar] [CrossRef] [Green Version]
- de Almeida Furtado, L.F.; Francisco, C.N.; de Almeida, C.M. Análise de imagem baseada em objeto para classificação das fisionomias da vegetação em imagens de alta resolução espacial. Geociências 2013, 32, 3, 441–451. [Google Scholar]
- Souza, J.M.F.; dos Reis, E.F.; Martins, A.S.; Santos, A.L.F. Avaliação dos conflitos no uso da terra na bacia hidrográfica do ribeirão Lamarão, Distrito Federal. Ciência Florest. 2019, 29, 950–964. [Google Scholar] [CrossRef] [Green Version]
- Maillard, P.; Pereira, D.B.; Souza, C.G. Incêndios florestais em veredas: Conceitos e estudo de caso no Peruaçu. Rev. Bras. Cartogr. 2009, 61, 321–330. [Google Scholar]
- Fontes, J.C.; Pozzetti, V.C. O uso dos veículos não tripulados no monitoramento ambiental na Amazônia. Rev. Direito Sustentabilidade 2016, 2, 149–164. [Google Scholar] [CrossRef]
- Longhitano, G.A. VANTs Para Sensoriamento Remoto: Aplicabilidade na Avaliação e Monitoramento de Impactos Ambientais Causados por Acidentes de Carga. Master’s Thesis, Escola Politécnica da Universidade de São Paulo, São Paulo, Brazil, 2010; 163p. [Google Scholar]
- MMA; ICMBIO; PNCG. Plano de Manejo Parque Nacional da Chapada dos Guimarães; Chapada dos Guimarães, Mato Grosso, Brazil, 2009. Available online: https://www.icmbio.gov.br/portal/images/stories/imgs-unidades-coservacao/parna_chapada-dos-guimaraes.pdf (accessed on 1 February 2022).
- ICMBio. Plano de Prevenção e Combate a Incêndios Florestais e Manejo Integrado e Adaptativo do Fogo do Parque Nacional da Chapada dos Guimarães. 2016. Available online: http://reflorestamentoecarbono.com.br/wp-content/uploads/2016/04/plano-de-protecao-pncg.pdf (accessed on 18 February 2020).
- Silva, R.; Paixão, E.; Cunha, C.; Finger, Z. Fitossociologia da comunidade arbórea de cerrado sensu stricto do Parque Nacional da Chapada dos Guimarães. Nativa 2016, 4, 82–86. [Google Scholar] [CrossRef] [Green Version]
- Spera, S.T.; Reatto, A.; Martins, E.D.S.; Correia, J.R.; Cunha, T.J.F. Solos areno-quartzosos no cerrado: Características, problemas e limitações ao solo. Planaltina Embrapa Cerrados 1999, 7, 48. [Google Scholar]
- Neves, W.V.; Menezes, E.P.; de Oliveira, F.S.; Augustin, C.H.R.R.; Aranha, P.R.A. Estudos das coberturas superficiais na interface cerrado-vereda no norte de Minas Gerais. Rev. Bras. Geomorf. 2015, 16, 351–367. [Google Scholar] [CrossRef] [Green Version]
- Alvares, C.A.; Stape, J.L.; Sentelhas, P.C.; de Moraes Gonçalves, J.L.; Sparovek, G. Köppen’s climate classification map for Brazil. Meteorol. Z. 2013, 22, 711–728. [Google Scholar] [CrossRef]
- ANA—Agência Nacional de Águas; GEF; PNUMA; OEA. Implementação de Práticas de Gerenciamento Integrado de Bacia Hidrográfica para o Pantanal e Bacia do Alto Paraguai ANA/GEF/PNUMA/OEA: Programa de Ações Estratégicas para o Gerenciamento Integrado do Pantanal e Bacia do Alto Paraguai: Relatório Final 2004; Campo Grande, Mato Grosso do Sul, Brazil, 2005; 137p. Available online: https://www.ecoa.org.br/wp-content/uploads/2015/11/RE_51MS.pdf (accessed on 1 February 2022).
- ICMBio—Instituto Chico Mendes de Conservação da Biodiversidade. Plano de Manejo: Parque Nacional da Chapada dos Guimarães. 2009. Available online: http://www.icmbio.gov.br/parnaguimaraes/images/stories/downloads/capa_apresentacao_e_indice.pdf (accessed on 7 October 2019).
- Ribeiro, J.F.; Walter, B.M.T. As principais fitofisionomias do Bioma Cerrado. In Cerrado: Ecologia e Flora; Sano, S.M., Almeida, S.P., Ribeiro, J.F., Eds.; Embrapa Cerrados: Brasília, Brazil, 2008; pp. 151–212. [Google Scholar]
- Felfili, J.M.; Carvalho, F.A.; Haidar, R.F. Manual Para o Monitoramento de Parcelas Permanentes nos Biomas Cerrado e Pantanal; Universidade de Brasília, Departamento de Engenharia Florestal: Brasília, Brazil, 2005; 55p. [Google Scholar]
- dos Santos, L.F.B. Avaliação de Modelo Digital de Terreno Gerado Através de VANT em Planícies Pantaneiras; Monografia (Graduação em Engenharia Florestal). Master’s Thesis, Universidade Federal de Mato Grosso—UFMT, Cuiabá, Brazil, 2016; 53p. [Google Scholar]
- Prina, B.Z.; Trenti, R. GPS x GNSS: Constelação GLONASS maximizando a precisão e acurácia dos dados. Rev. Eletrônica Gestão Educ. E Tecnol. Ambient. Santa Maria 2015, 19, 344–352. [Google Scholar]
- Arevalo, L.A.; Alegre, J.C.; Vilcahuaman, L.J.M. Metodologia para Estimar o Estoque de Carbono em Diferentes Sistemas de uso da Terra; Colombo Embrapa Florestas: Colombo, Brazil, 2002; Volume 73, 41p. [Google Scholar]
- Meschede, D. Avaliação de diferentes coberturas na supressão de plantas daninhas no cerrado. Planta Daninha 2007, 25, 465–471. [Google Scholar] [CrossRef] [Green Version]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; University of Illinois Press: Urbana, IL, USA, 1949. [Google Scholar]
- Brower, J.E.; Zar, J.H. Field and Laboratory Methods for General ecology; Wm. C. Brown Company: Dubuque, Iowa, 1984; p. 226. [Google Scholar]
- Mueller-Dombois, D.; Ellenberg, H. Aims and Methods of Vegetation Ecology; John Wiley & Sons: New York, NY, USA, 1974; p. 547. [Google Scholar]
- Mcguinnes, W.G. A relação entre índice de frequência e abundância aplicada a populações de plantas em uma região semi-árida. Ecology 1934, 16, 263–282. [Google Scholar]
- Collischonn, C.; de Lima, E.M.; Ruiz, L.F.C.; Araujo, T.D.; Matsuoka, M.T. Posicionamento GNSS: Comparação entre coordenadas oficiais de estações da RBMC e as obtidas por PPP e posicionamento relativo e ajustamento. In Simpósio Brasileiro de Sensoriamento Remoto—SBSR; INPE: João Pessoa, Brazil, 2015. [Google Scholar]
- Fernandes, R.R.; Nunes, G.M.; Silva, T.S.F. Classificação orientada a objetos aplicada na caracterização da cobertura da terra no Araguaia. Pesqui. Agropecuária Bras. 2012, 47, 1251–1260. [Google Scholar] [CrossRef] [Green Version]
- Definiens, T.G. Trimble eCognition Developer for Windows Operating System; 2018. Available online: https://www.geospatial.trimble.com/ecognition-download (accessed on 15 February 2022).
- Baatz, M.; Schäpe, A. Multiresolution Segmentation—an optimization approach for high quality multi-scale image segmentation. In Angewandte Geographische Informationsverarbeitung XII; Strobl, J., Blaschke, T., Griesbner, G., Eds.; Wichmann, Verlag: Karlsruhe, Germany, 2000; pp. 12–23. [Google Scholar]
- Yang, Z.; Willis, P.E.; Mueller, R. Impact of band-ratio enhanced awifs image to crop classification accuracy. Proc. Pecora 2008, 11, 1–11. [Google Scholar]
- Klock, U.; Muñiz, G.I.B.; de Hernandez, J.A.; Andrade, A.S. Química da Madeira. In Manual Didático: Curitiba, 3rd ed.; Revisada; Universidade Federal do Pará: Curitiba, Brazil, 2005. [Google Scholar]
- Corpo de Bombeiros Militar. Manual Operacional de Bombeiros: Perícia de Incêndio; Corpo de Bombeiros Militar: Goiânia, Brazil, 2017; 276p.
- Landis, J.R.; Koch, G.G. The measurements of observer agreement for categorical data. Biometrics 1977, 33, 159–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, J.; Wan, S.; Li, Z.; Shao, Y.; Xu, G.; Liu, Z.; Zhou, L.; Fu, S. Dicranopteris-dominated understory as major driver of intensive forest ecosystem in humid subtropical and tropical region. Soil Biol. Biochem. 2012, 49, 78–87. [Google Scholar] [CrossRef]
- Ramos, M.V.V.; Haridasan, M.; De Araújo, G.M. Caracterização dos Solos e da Estrutura Fitossociológica da Vegetação de Veredas da Chapada no Triângulo Mineiro. Front. J. Social, Technol. Environ. Sci. 2014, 3, 180. [Google Scholar] [CrossRef]
- Moreira, S.N.; Pott, V.J.; Pott, A.; Da Silva, R.H.; Júnior, G.A.D. Flora and vegetation structure of vereda in southwestern cerrado. Oecologia Aust. 2019, 23, 776–798. [Google Scholar] [CrossRef]
- Guimarães, A.J.M.; De Araújo, G.M.; Corrêa, G.F. Estrutura fitossociológica em área natural e antropizada de uma vereda em Uberlândia, MG. Acta Bot. Bras. 2002, 16, 317–329. [Google Scholar] [CrossRef]
- Resende, I.L.D.M.; Dos Santos, F.P.; Chaves, L.J.; Nascimento, J.L.D. Estrutura etária de populações de Mauritia flexuosa L. F. (Arecaceae) de veredas da região central de Goiás, Brasil. Rev. Árvore 2012, 36, 103–112. [Google Scholar] [CrossRef] [Green Version]
- Marimon, B.S.; Lima, E.D.S.; Duarte, T.G.; Chieregatto, L.C.; Ratter, J.A. Observations on the vegetation of northeastern mato grosso, brazil. IV. An analysis of the cerrado–amazonian forest ecotone. Edinb. J. Bot. 2006, 63, 323–341. [Google Scholar] [CrossRef]
- Miranda, I.S.; Almeida, S.S.; Dantas, P.J. Florística e estrutura de comunidades arbóreas em cerrados de Rondônia, Brasil. Acta Amaz. 2006, 36, 419–430. [Google Scholar] [CrossRef] [Green Version]
- Valdés-Sáenz, M.A.; García-Quintana, Y.; Escarré-Esteve, A.; Flores, J.; Geada-López, G.; Arteaga-Crespo, Y.; Valdés-Sáenz, C.R. Estructura de un bosque natural perturbado de Pinus tropicalis Morelet en Galalón, Cuba. Bot. Sci. 2014, 92, 417. [Google Scholar] [CrossRef] [Green Version]
- De Souza, C.R.; Rossi, L.M.B.; de Souza, C.R.; de Azevedo, C.P.; Yared, J.A.G. Taxi-Branco (Sclerolobium Paniculatum Vogel). In Manaus: Embrapa Amazônia Ocidental; Documentos 34; Manaus, Amazônia, Brazil, 2004; 23p, Available online: https://www.infoteca.cnptia.embrapa.br/bitstream/doc/675654/1/Doc34.pdf (accessed on 16 February 2020).
- Pires, I.P.; Marcati, C.R. Anatomia e uso da madeira de duas variedades de Sclerolobium paniculatum Vog. do sul do Maranhão, Brasil. Acta Bot. Bras. 2005, 19, 669–678. [Google Scholar] [CrossRef] [Green Version]
- Ávila, M.A.; Souza, S.R.; Veloso, M.D.D.M.; Santos, R.M.; Fernandes, L.A.; Nunes, Y.R.F. Structure of natural regeneration in relation to soil properties and disturbance in two swamp forests. Cerne 2016, 22, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Albor Pinto, C.; Tun Garrido, J.; Ortiz Díaz, J.J. Diversidad, estructura y afinidades florísticas de un bosque temporalmente inundable de la Península de Yucatán. Rev. Biol. Trop. 2017, 65, 68–880. [Google Scholar] [CrossRef] [Green Version]
- Lorenzi, H. Árvores Brasileiras: Manual de Identificação e Cultivo de Plantas Arbóreas do Brasil, 5th ed.; Instituto Plantarum: Nova Odessa, Brazil, 2008; Volume 1. [Google Scholar]
- Carvalho, P.E.R. Espécies arbóreas brasileiras. In Coleção Espécies Arbóreas Brasileiras, v. 1; Embrapa Informação Tecnológica: Brasília, Brazil; Embrapa Florestas: Colombo, Brazil, 2003; 1039p. [Google Scholar]
- Araújo, G.M.; Barbosa, A.A.A.; Arantes, A.A.; Amaral, A.F. Composição florística de veredas no Município de Uberlândia, MG. Braz. J. Bot. 2002, 25, 475–493. [Google Scholar] [CrossRef]
- Munhoz, C.B.R.; Felfili, J.M. Fitossociologia do estrato herbáceo-subarbustivo em campo limpo úmido no Brasil Central. Acta Bot. Bras. 2008, 22, 905–913. [Google Scholar] [CrossRef]
- Townsend, C.; Begon, M.; Harper, J.L. Fundamentos em Ecologia, 3rd ed.; Artmed: Porto Alegre, Brazil, 2010; p. 576. [Google Scholar]
- Miranda, S.C.; Carvalho, P.S.; Bustamante, M.M.C.; Silva Júnior, M.C. Variação temporal na estrutura da vegetação lenhosa de cerrado sentido restrito sobre Neossolos Quartzarênicos. Rev. Espac. 2017, 38, 3. [Google Scholar]
- da Silva, M.P.; Santana, N.C.; Guedes, S.R.A.; de Paula Laranja, R.E. Levantamento fitossociológico em ambiente de Vereda na apa ribeirões do gama e cabeça de veado, Brasília-DF. Rev. Geoaraguaia 2016, 6, 84–98. [Google Scholar]
- dos Santos, H.G.; Jacomine, P.K.T.; dos Anjos, L.H.C.; De Oliveira, V.A.; Lumbreras, J.F.; Coelho, M.R.; De Almeida, J.A.; de Araujo Filho, J.C.; de Oliveira, J.D.; Cunha, T.J.F. Sistema Brasileiro de Classificação de Solos; Embrapa: Brasília, Brazil, 2018. [Google Scholar]
- Tarantino, E.; Figorito, B. Mapping Rural Areas with Widespread Plastic Covered Vineyards Using True Color Aerial Data. Remote Sens. 2012, 4, 1913–1928. [Google Scholar] [CrossRef] [Green Version]
- Meneses, P.R.; de Almeida, T. Introdução ao Processamento de Imagens de Sensoriamento Remoto; UnB, CNPq: Brasília, Brazil, 2012; Available online: http://www.cnpq.br/documents/10157/56b578c4-0fd5-4b9f-b82a-e9693e4f69d8 (accessed on 7 October 2020).
- Queiroz, M.L. Nascentes, Veredas e Áreas Úmidas—Revisão Conceitual e Metodologia de Caracterização e Determinação: Estudo de caso na Estação Ecológica de Águas Emendadas—Distrito Federal. Master’s Thesis, Universidade de Brasília, Distrito Federal, Brazil, 2015; 161p. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, A.C.A.; de Paula Laranja, R.E.; de Oliveira Nascimento, R.; Rodrigues, F.P. Delimitação e caracterização de um transecto na vertente do subsistema de vereda na reserva ecológica do IBGE/ RECOR-DF. Rev. Caminhos Geogr. 2018, 19, 233–249. [Google Scholar] [CrossRef]
- Cunha, C.N.; da Piedade, M.T.F.; Junk, W.J. Classificação e Delineamento das Áreas Úmidas Brasileiras e de Seus Macrohabitats; EdUFMT: Cuiabá, Brazil, 2015; 165p. [Google Scholar]
- De Pires, P.T.L.; Loper, A.A.; Mendes, C.J.; Peters, E.L.; Maia, G.N.; de Abreu, L.M. Dicionário de Termos Florestais; FUPEF: Curitiba, Brazil, 2018. [Google Scholar]
- Carvalho, P.E.R. Espécies arbóreas brasileiras. In Coleção Espécies Arbóreas Brasileiras, v. 2; Embrapa Informação Tecnológica: Brasília, Brazil; Embrapa Florestas: Colombo, Brazil, 2006; 627p, ISBN 85-7383-373-4. [Google Scholar]
- Sartorelli, P.A.R.; Campos Filho, E.M. Guia de Plantas da Regeneração Natural do Cerrado e da Mata Atlântica; Agroicone: São Paulo, Brazil, 2017; Available online: https://www.inputbrasil.org/wp-content/uploads/2017/05/INPUT_Agroicone_Guia-de-Plantas-da-Regeneracao-Natural-do-Cerrado-e-da-Mata-Atlantica.pdf (accessed on 18 February 2020).
- Hoffmann, W.A. Post-Establishment Seedling Success in the Brazilian Cerrado: A Comparison of Savanna and Forest Species1. Biotropica 2000, 32, 62–69. [Google Scholar] [CrossRef]
- Hoffmann, W.A. Fire and population dynamics of woody plants in a neotropical savanna: Matrix model projections. Ecology 1999, 80, 1354–1369. [Google Scholar] [CrossRef]
- Ribeiro, G.A. Estudos do Comportamento do Fogo e de Alguns Efeitos da Queima Controlada em Povoamentos de Eucalyptus Viminalis Labil em Três Barras. Ph.D. Thesis, Universidade Federal do Paraná, Curitiba, Brazil, 1997. [Google Scholar]
- Myers, R. Convivendo Com o Fogo: Manutenção dos Ecossistemas & Subsistência com o Manejo Integrado do Fogo; The Nature Conservancy: Tallahassee, FL, USA, 2006. [Google Scholar]
- Couto, E.G.; Chig, L.A.; Cunha, C.D.; Loureiro, M.D.F. Conhecendo o Pantanal 2: Estudo Sobre o Impacto do Fogo na Disponibilidade de Nutrientes, no Banco de Sementes e na Biota de Solos da RPPN SESC Pantanal; Serviço Social do Comércio: Rio de Janeiro, Brazil, 2006; 56p. [Google Scholar]
- Matos, D.M.S.; Pivello, V.R. O impacto das plantas invasoras nos recursos naturais de ambientes terrestres—Alguns casos brasileiros. Cienc. Cult. 2009, 61, 21–30. [Google Scholar]
- Henriques, R.P.B.; Hay, J.D. Patterns and Dynamics of Plant Populations. In The Cerrados of Brazil: Ecology and Natural History of a Neotropical Savana; Oliveira, P.S., Marquis, R.J., Eds.; Columbia University Press: New York, NY, USA, 2002. [Google Scholar]
- Fank-de-Carvalho, S.M.; Báo, S.N.; Marchioretto, M.S. Amaranthaceae as a Bioindicator of Neotropical Savannah Diversity. In Biodiversity Enrichment in a Diverse World; IntechOpen: London, UK, 2012; pp. 235–262. [Google Scholar] [CrossRef] [Green Version]
- Silva, D.M.; de Paula Loiola, P.; Rosatti, N.B.; Silva, I.A.; Cianciaruso, M.V.; Batalha, M.A. Os efeitos dos regimes de fogo sobre a vegetação de cerrado no parque nacional das emas, GO: Considerações para a conservação da diversidade. Biodivers. Bras. 2011, 2, 26–39. [Google Scholar] [CrossRef]
- Montoya, E.; Rull, V.; Nogué, S.; Diaz, W.A. Paleoecología del Holoceno en la Gran Sabana, SE Venezuela: Análisis preliminar de polen y microcarbones en la Laguna Encantada. Collect. Bot. 2009, 28, 65–79. [Google Scholar] [CrossRef] [Green Version]
- Montoya, E.; Rull, V.; Stansell, N.; Abbott, M.B.; Nogué, S.; Bird, B.W.; Díaz, W.A. Forest–savanna–morichal dynamics in relation to fire and human occupation in the southern Gran Sabana (SE Venezuela) during the last millennia. Quat. Res. 2011, 76, 335–344. [Google Scholar] [CrossRef] [Green Version]
- Campos, J.M.S.; Davide, L.C.; Soares, G.L.G.; Viccini, L.F. Mutagenic effects due to allelopathic action of fern (Gleicheniaceae) extracts. Allelopath. J. 2008, 22, 143–151. [Google Scholar]
- de Werneck, M.S. Conservação da Flora e Planejamento Ambiental no Contexto da Expansão Urbana da Região Metropolitana de Belo Horizonte, Minas Gerais. Ph.D. Thesis, Universidade Federal de Minas Gerais, Belo Horizonte, Brazil, 2006. [Google Scholar]
- de Paula Athayde Filho, F.; Agostinho, A.A. Pteridoflora de duas Veredas no Município de Campinápolis, Mato Grosso, Brasil; Pesquisas, Botânica. N° 56: 145-160; Instituto Anchietano de Pesquisas: São Leopoldo, Brazil, 2005. [Google Scholar]
- Gonçalves, J.R.; Ferreira, J.F.R. Pteridófitas: Uma investigação de táxons na vereda do córrego Mogi no município de Uberlândia-MG. E-RAC 2012, 2, 1–17. [Google Scholar]
- BRASIL. Resolução nº 7, De 29 De Maio De 2018. Brasília, DF, Mai 2018. Available online: http://www.institutohorus.org.br/download/marcos_legais/Resolucao_CONABIO%2007%20-%202018.pdf (accessed on 15 February 2022).
- BRASIL. Decreto nº 2.519, De 16 De Março De 1998. Brasília, DF, Mar. 1998. Available online: http://www.planalto.gov.br/ccivil_03/decreto/d2519.htm (accessed on 15 February 2022).
- ICMBIO; MMA. Guia de Orientação para o Manejo de Espécies Exóticas Invasoras em Unidades de Conservação Federais. 2019. Available online: https://www.icmbio.gov.br/cbc/images/stories/Publica%C3%A7%C3%B5es/EEI/Guia_de_Manejo_de_EEI_em_UC_v3.pdf (accessed on 18 February 2020).
- Carvajal-Ramírez, F.; Marques da Silva, J.R.; Agüera-Vega, F.; Martínez-Carricondo, P.; Serrano, J.; Moral, F.J. Evaluation of Fire Severity Indices Based on Pre- and Post-Fire Multispectral Imagery Sensed from UAV. Remote Sens. 2019, 11, 993. [Google Scholar] [CrossRef] [Green Version]
- McKenna, P.; Erskine, P.D.; Lechner, A.M.; Phinn, S. Measuring fire severity using UAV imagery in semi-arid central Queensland, Australia. Int. J. Remote Sens. 2017, 38, 4244–4264. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
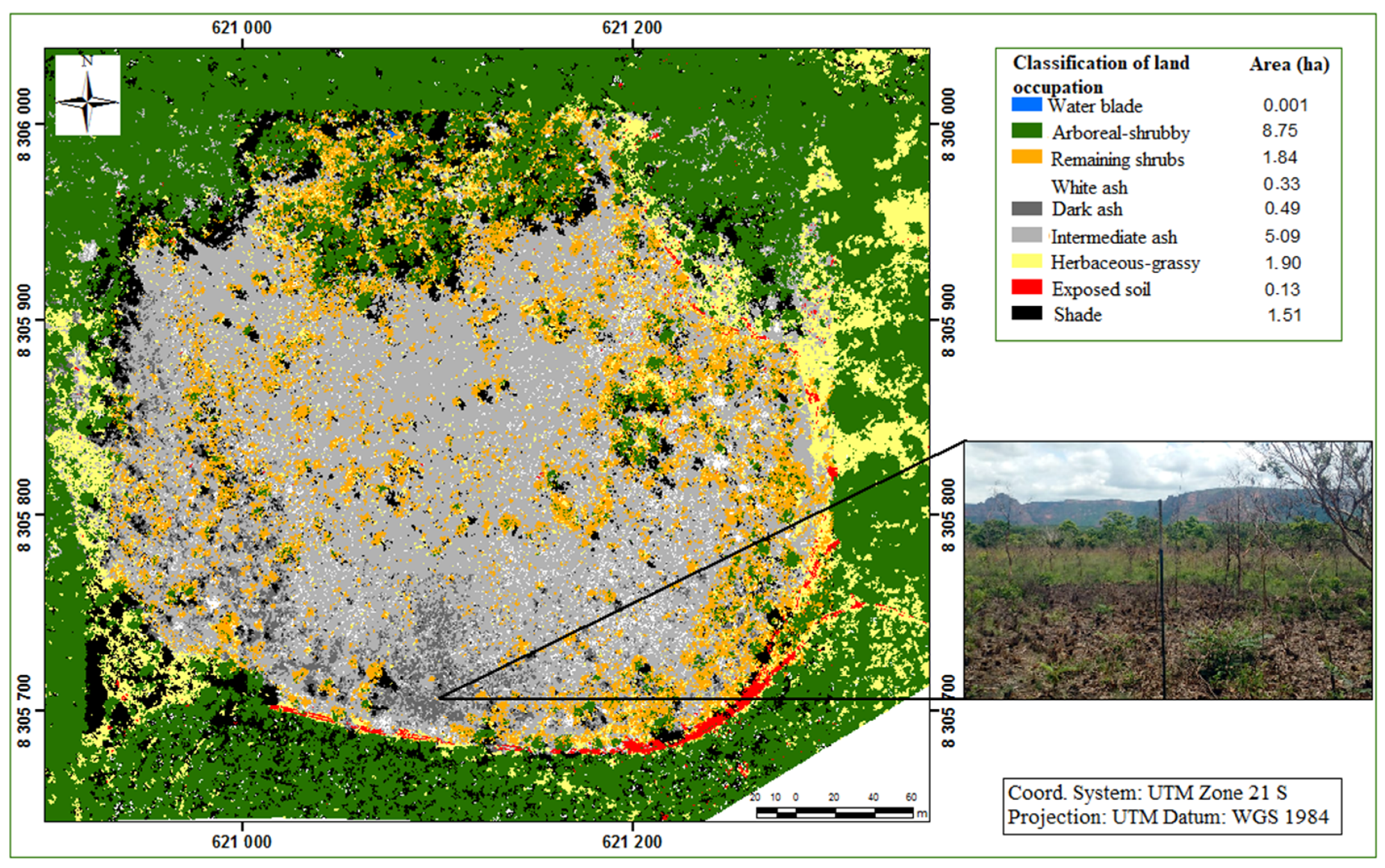{kind=link}
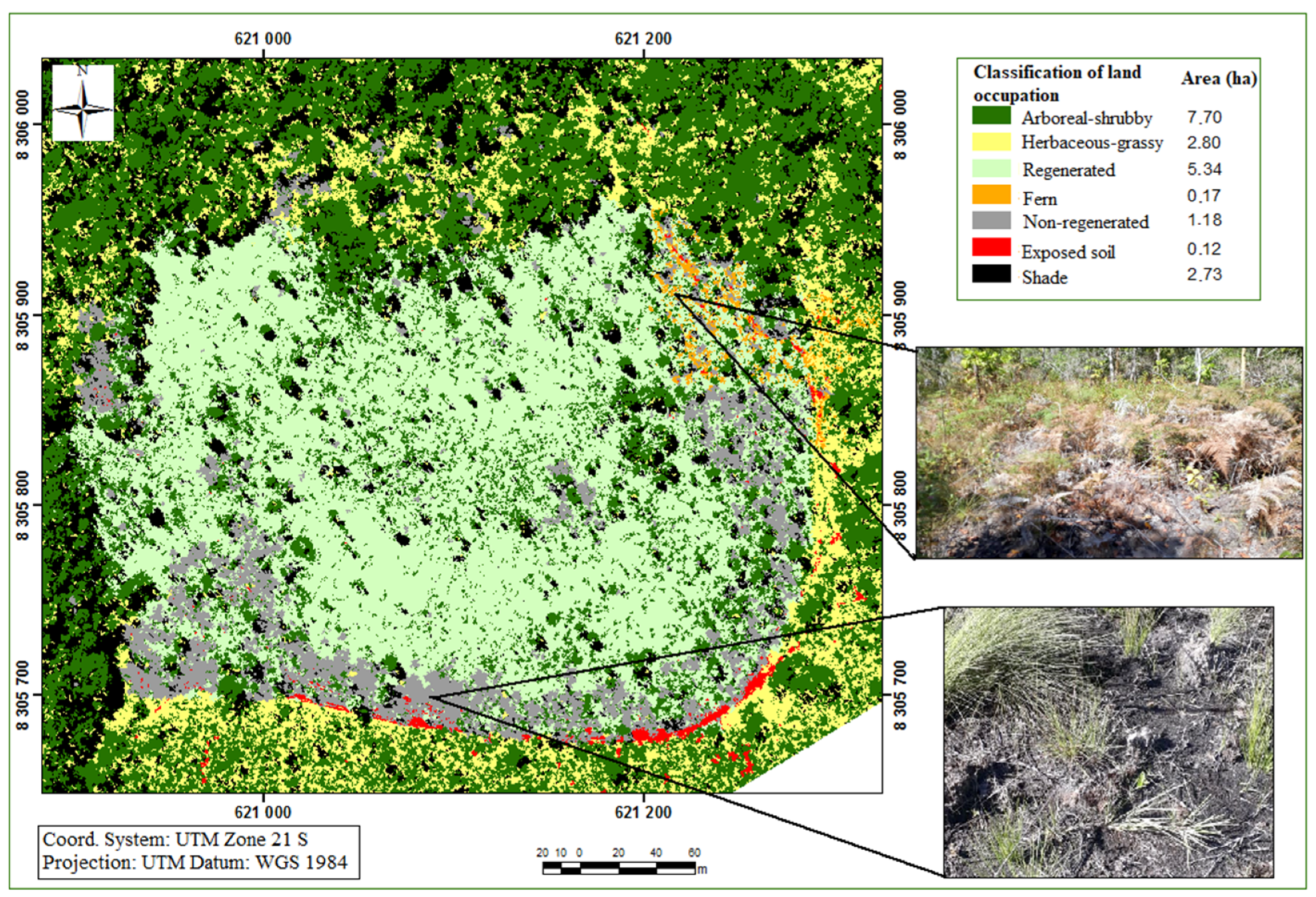{kind=link}
| Pre-Burn | Post-Burn | Monitoring | |
|---|---|---|---|
| Descriptors | Standard deviation of band 2 | Average of band 1 | Customized MPRI |
| Maximum difference | Maximum difference | Average of band 1 average | |
| Average of band 2 | Density | Rectangular shape | |
| Standard deviation of band 1 | Standard deviation of band 2 | Standard deviation of band 1 | |
| Customized MPRI | Average of layer 2 | Maximum difference |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nascente, J.C.; Ferreira, M.E.; Nunes, G.M. Integrated Fire Management as a Renewing Agent of Native Vegetation and Inhibitor of Invasive Plants in Vereda Habitats: Diagnosis by Remotely Piloted Aircraft Systems. Remote Sens. 2022, 14, 1040. https://doi.org/10.3390/rs14041040
Nascente JC, Ferreira ME, Nunes GM. Integrated Fire Management as a Renewing Agent of Native Vegetation and Inhibitor of Invasive Plants in Vereda Habitats: Diagnosis by Remotely Piloted Aircraft Systems. Remote Sensing. 2022; 14(4):1040. https://doi.org/10.3390/rs14041040
Chicago/Turabian StyleNascente, Jéssika Cristina, Manuel Eduardo Ferreira, and Gustavo Manzon Nunes. 2022. "Integrated Fire Management as a Renewing Agent of Native Vegetation and Inhibitor of Invasive Plants in Vereda Habitats: Diagnosis by Remotely Piloted Aircraft Systems" Remote Sensing 14, no. 4: 1040. https://doi.org/10.3390/rs14041040
APA StyleNascente, J. C., Ferreira, M. E., & Nunes, G. M. (2022). Integrated Fire Management as a Renewing Agent of Native Vegetation and Inhibitor of Invasive Plants in Vereda Habitats: Diagnosis by Remotely Piloted Aircraft Systems. Remote Sensing, 14(4), 1040. https://doi.org/10.3390/rs14041040