


Quantifying Temperate Forest Diversity by Integrating GEDI LiDAR and Multi-Temporal Sentinel-2 Imagery



Abstract
:1. Introduction
2. Materials
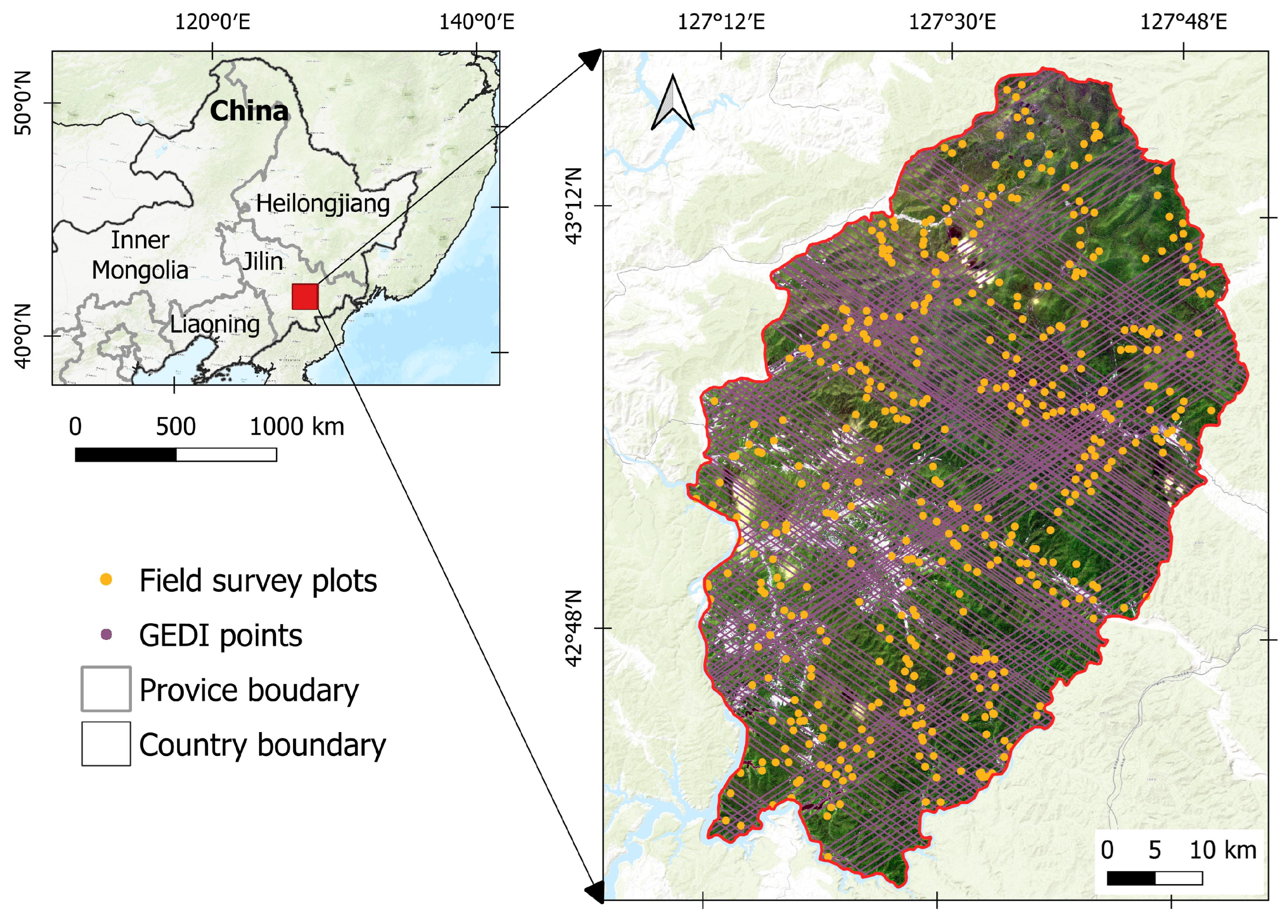
2.1. Study Area
2.2. Field Botanical Surveys
2.3. Data Source and Processing
2.3.1. Diversity Index Data
2.3.2. Sentinel-2 Images
2.3.3. GEDI LiDAR Data
3. Methods
3.1. Variable Importance Assessment
3.2. Algorithms for Forest Diversity Mapping
3.3. Accuracy Assessment
4. Results
4.1. Optimal Features from SENTINEL-2 Images and GEDI LiDAR Data
4.2. Diversity Indices Modelling Using Machine Learning Algorithms
4.3. Spatial Variability of the Predicted Diversity Indices
5. Discussion
5.1. Prospects of GEDI LiDAR and Sentinel-2 Data on Forest Diversity
5.2. Machine Learning Algorithms for Forest Diversity Mapping
5.3. Prediction Performance and Uncertainty for Forest Diversity
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Simpson | Shannon | Pielou | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Rank | Variables | BRT (%) | MDG (%) | Variables | BRT (%) | MDG (%) | Variables | BRT (%) | MDG (%) |
| 1 | FHD_GS | 16.49 | 14.07 | FHD_GS | 15.59 | 12.14 | NDVI_Jun | 11.43 | 11.34 |
| 2 | NDVI_Jun | 12.16 | 12.37 | NDVI_Jun | 12.78 | 8.52 | FHD_GS | 9.33 | 7.48 |
| 3 | NDWI_May | 7.74 | 8.91 | NDWI_May | 10.48 | 9.38 | NDWI_Jun | 8.51 | 3.97 |
| 4 | PAI_GS | 7.55 | 6.33 | B12_May | 7.59 | 7.09 | NDWI_May | 5.38 | 5.07 |
| 5 | NDWI_Jun | 5.25 | 5.66 | NDVI_Oct | 6.40 | 6.84 | PAI_GS | 4.72 | 3.77 |
| 6 | B12_May | 4.58 | 5.55 | PAI_GS | 2.10 | 4.73 | B12_May | 3.97 | 2.94 |
| 7 | EVI_May | 5.18 | 3.84 | B11_Oct | 4.11 | 3.40 | EVI_May | 2.95 | 2.38 |
| 8 | NDVI_May | 3.75 | 3.18 | EVI_May | 3.52 | 2.70 | B11_Oct | 2.83 | 2.42 |
| 9 | B12_Oct | 3.21 | 2.76 | B7_Jun | 2.77 | 2.37 | B7_Jun | 2.77 | 2.40 |
| 10 | B7_Jun | 2.91 | 2.47 | B11_Jun | 2.36 | 2.25 | B11_Jun | 2.70 | 2.06 |
| 11 | B11_Oct | 2.58 | 2.25 | B12_Oct | 2.07 | 2.23 | B12_Oct | 1.66 | 2.02 |
| 12 | B8A_Jun | 1.97 | 2.18 | NDVI_May | 1.83 | 1.94 | NDVI_May | 1.59 | 1.68 |
| 13 | B6_Jun | 1.71 | 1.80 | B8A_Jun | 1.43 | 1.85 | B5_May | 1.59 | 1.45 |
| 14 | B2_Jun | 1.46 | 1.56 | B8A_May | 1.32 | 1.46 | B8A_May | 1.50 | 1.41 |
| 15 | FHD_NGS | 1.27 | 1.10 | FHD_NGS | 1.20 | 1.46 | B5_Jun | 1.49 | 1.40 |
| 16 | B11_May | 1.15 | 1.02 | EVI_Oct | 1.26 | 1.34 | B8A_Jun | 1.44 | 1.29 |
| 17 | B1_Jun | 0.92 | 0.99 | B1_May | 1.14 | 1.29 | B5_Sep | 1.29 | 1.18 |
| 18 | NDWI_Sep | 0.85 | 0.87 | B8_Jun | 1.08 | 1.19 | NDVI_Oct | 1.16 | 1.17 |
| 19 | B4_May | 0.83 | 0.85 | DVI_Sep | 1.02 | 1.05 | B3_Sep | 1.15 | 1.17 |
| 20 | B3_May | 0.78 | 0.84 | B5_Oct | 1.02 | 1.03 | B3_Oct | 1.11 | 1.16 |
| 21 | B1_May | 0.76 | 0.79 | EVI_Jun | 0.98 | 1.00 | B4_Oct | 1.09 | 1.13 |
| 22 | B7_Sep | 0.75 | 0.78 | DVI_Oct | 0.96 | 0.82 | B7_Sep | 1.07 | 1.12 |
| 23 | B1_Oct | 0.74 | 0.77 | B3_May | 0.89 | 0.80 | B6_Sep | 1.05 | 1.11 |
| 24 | EVI_Oct | 0.74 | 0.71 | B4_May | 0.89 | 0.78 | B1_May | 1.05 | 1.11 |
| 25 | B6_Sep | 0.71 | 0.69 | B5_Jun | 0.84 | 0.75 | B1_Oct | 1.04 | 1.09 |
| 26 | B5_Jun | 0.70 | 0.67 | B1_Oct | 0.78 | 0.75 | B4_Sep | 0.99 | 1.06 |
| 27 | DVI_Sep | 0.69 | 0.60 | B12_Jun | 0.75 | 0.72 | B2_Oct | 0.97 | 1.05 |
| 28 | B12_Jun | 0.68 | 0.60 | B4_Sep | 0.66 | 0.72 | B1_Jun | 0.97 | 1.05 |
| 29 | B5_Sep | 0.65 | 0.59 | B2_Jun | 0.65 | 0.69 | DVI_Sep | 0.97 | 1.04 |
| 30 | B11_Jun | 0.65 | 0.58 | B3_Sep | 0.62 | 0.68 | B2_Sep | 0.96 | 1.03 |
| 31 | DVI_Oct | 0.60 | 0.58 | B5_May | 0.61 | 0.68 | PAI_NGS | 0.95 | 1.03 |
| 32 | B8_Jun | 0.56 | 0.55 | B1_Jun | 0.61 | 0.65 | DVI_Jun | 0.93 | 1.00 |
| 33 | NDVI_Oct | 0.56 | 0.55 | B2_Sep | 0.57 | 0.64 | DVI_Oct | 0.93 | 0.99 |
| 34 | PAI_NGS | 0.55 | 0.49 | B2_Oct | 0.55 | 0.63 | B6_May | 0.85 | 0.98 |
| 35 | DVI_Jun | 0.54 | 0.49 | B7_Sep | 0.50 | 0.61 | B6_Jun | 0.84 | 0.98 |
| 36 | B2_Oct | 0.51 | 0.49 | NDWI_Sep | 0.48 | 0.61 | B4_May | 0.82 | 0.96 |
| 37 | B8_Oct | 0.48 | 0.48 | B8_May | 0.45 | 0.61 | EVI_Oct | 0.82 | 0.94 |
| 38 | B9_Oct | 0.48 | 0.47 | NDWI_Oct | 0.44 | 0.56 | B9_Oct | 0.80 | 0.92 |
| 39 | B3_Jun | 0.47 | 0.44 | B6_May | 0.44 | 0.54 | B3_Jun | 0.77 | 0.91 |
| 40 | B3_Sep | 0.47 | 0.43 | B8A_Sep | 0.43 | 0.52 | B1_Sep | 0.73 | 0.88 |
| 41 | B3_Oct | 0.46 | 0.42 | NDWI_Jun | 0.41 | 0.51 | B5_Oct | 0.73 | 0.88 |
| 42 | B5_Oct | 0.46 | 0.41 | B11_May | 0.40 | 0.51 | B8_Jun | 0.72 | 0.88 |
| 43 | B2_Sep | 0.31 | 0.40 | B1_Sep | 0.39 | 0.50 | B2_May | 0.71 | 0.86 |
| 44 | B1_Sep | 0.31 | 0.40 | B6_Oct | 0.37 | 0.49 | EVI_Jun | 0.71 | 0.85 |
| 45 | B8A_Sep | 0.30 | 0.39 | B12_Sep | 0.37 | 0.48 | B12_Jun | 0.70 | 0.85 |
| 46 | B4_Oct | 0.30 | 0.39 | B6_Jun | 0.36 | 0.48 | NDVI_Sep | 0.70 | 0.85 |
| 47 | B2_May | 0.28 | 0.39 | DVI_Jun | 0.36 | 0.47 | B8A_Sep | 0.70 | 0.84 |
| 48 | B8_May | 0.28 | 0.39 | B8_Oct | 0.31 | 0.47 | B11_May | 0.68 | 0.83 |
| 49 | EVI_Jun | 0.27 | 0.38 | B5_Sep | 0.30 | 0.46 | B3_May | 0.65 | 0.83 |
| 50 | B4_Sep | 0.26 | 0.38 | NDVI_Sep | 0.30 | 0.45 | SAVI_May | 0.56 | 0.83 |
| 51 | NDWI_Oct | 0.24 | 0.37 | B6_Sep | 0.30 | 0.45 | SAVI_Sep | 0.55 | 0.81 |
| 52 | B5_May | 0.24 | 0.37 | B2_May | 0.29 | 0.45 | B8_Oct | 0.53 | 0.81 |
| 53 | B7_Oct | 0.21 | 0.37 | B11_Sep | 0.28 | 0.43 | FHD_NGS | 0.52 | 0.81 |
| 54 | B8A_May | 0.20 | 0.37 | PAI_NGS | 0.26 | 0.42 | B6_Oct | 0.52 | 0.81 |
| 55 | B6_Oct | 0.19 | 0.37 | B7_May | 0.23 | 0.42 | B11_Sep | 0.50 | 0.79 |
| 56 | B11_Sep | 0.19 | 0.36 | B9_Oct | 0.22 | 0.42 | B8_Sep | 0.48 | 0.79 |
| 57 | B8_Sep | 0.18 | 0.36 | B3_Oct | 0.21 | 0.41 | B4_Jun | 0.48 | 0.77 |
| 58 | B12_Sep | 0.18 | 0.35 | B4_Jun | 0.19 | 0.41 | B7_May | 0.48 | 0.77 |
| 59 | NDVI_Sep | 0.16 | 0.33 | SAVI_Sep | 0.15 | 0.41 | B2_Jun | 0.47 | 0.77 |
| 60 | B4_Jun | 0.15 | 0.33 | B4_Oct | 0.07 | 0.40 | B8_May | 0.45 | 0.75 |
| 61 | B6_May | 0.13 | 0.32 | DVI_May | 0.02 | 0.40 | DVI_May | 0.33 | 0.73 |
| 62 | DVI_May | 0.03 | 0.31 | B7_Oct | 0.02 | 0.39 | B12_Sep | 0.30 | 0.72 |
| 63 | SAVI_May | 0.01 | 0.29 | B8_Sep | 0.02 | 0.39 | B7_Oct | 0.22 | 0.72 |
| 64 | B7_May | 0.00 | 0.27 | SAVI_May | 0.01 | 0.38 | NDWI_Oct | 0.17 | 0.67 |
| 65 | SAVI_Sep | 0.00 | 0.27 | B3_Jun | 0.00 | 0.37 | SAVI_Jun | 0.00 | 0.66 |
| 66 | SAVI_Jun | 0.00 | 0.25 | SAVI_Jun | 0.00 | 0.34 | EVI_Sep | 0.00 | 0.65 |
| 67 | EVI_Sep | 0.00 | 0.24 | EVI_Sep | 0.00 | 0.33 | SAVI_Oct | 0.00 | 0.65 |
| 68 | SAVI_Oct | 0.00 | 0.20 | SAVI_Oct | 0.00 | 0.32 | NDWI_Sep | 0.00 | 0.65 |



References
- Qi, W.; Saarela, S.; Armston, J.; Ståhl, G.; Dubayah, R. Forest biomass estimation over three distinct forest types using TanDEM-X InSAR data and simulated GEDI lidar data. Remote Sens. Environ. 2019, 232, 111283. [Google Scholar] [CrossRef]
- Madonsela, S.; Cho, M.A.; Ramoelo, A.; Mutanga, O. Remote sensing of species diversity using Landsat 8 spectral variables. ISPRS J. Photogramm. Remote Sens. 2017, 133, 116–127. [Google Scholar] [CrossRef] [Green Version]
- Oldeland, J.; Wesuls, D.; Rocchini, D.; Schmidt, M.; Jürgens, N. Does using species abundance data improve estimates of species diversity from remotely sensed spectral heterogeneity? Ecol. Indic. 2010, 10, 390–396. [Google Scholar] [CrossRef]
- Brown, K.A.; Gurevitch, J. Long-term impacts of logging on forest diversity in Madagascar. Proc. Natl. Acad. Sci. USA 2004, 101, 6045–6049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asner, G.P.; Martin, R.E.; Knapp, D.E.; Tupayachi, R.; Anderson, C.B.; Sinca, F.; Vaughn, N.R.; Llactayo, W. Airborne laser-guided imaging spectroscopy to map forest trait diversity and guide conservation. Science 2017, 355, 385–389. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhang, Z.; Lutz, J.A.; Chu, C.; Hu, J.; Shen, G.; Li, B.; Yang, Q.; Lian, J.; Zhang, M.; et al. Drone-acquired data reveal the importance of forest canopy structure in predicting tree diversity. For. Ecol. Manag. 2022, 505, 119945. [Google Scholar] [CrossRef]
- Nagendra, H.; Rocchini, D.; Ghate, R.; Sharma, B.; Pareeth, S. Assessing Plant Diversity in a Dry Tropical Forest: Comparing the Utility of Landsat and Ikonos Satellite Images. Remote Sens. 2010, 2, 478–496. [Google Scholar] [CrossRef] [Green Version]
- Stenzel, S.; Fassnacht, F.E.; Mack, B.; Schmidtlein, S. Identification of high nature value grassland with remote sensing and minimal field data. Ecol. Indic. 2017, 74, 28–38. [Google Scholar] [CrossRef]
- Almeida, D.R.A.d.; Broadbent, E.N.; Ferreira, M.P.; Meli, P.; Zambrano, A.M.A.; Gorgens, E.B.; Resende, A.F.; de Almeida, C.T.; do Amaral, C.H.; Corte, A.P.D.; et al. Monitoring restored tropical forest diversity and structure through UAV-borne hyperspectral and lidar fusion. Remote Sens. Environ. 2021, 264, 112582. [Google Scholar] [CrossRef]
- Fauvel, C.; Weizman, O.; Trimaille, A.; Mika, D.; Pommier, T.; Pace, N.; Douair, A.; Barbin, E.; Fraix, A.; Bouchot, O.; et al. Pulmonary embolism in COVID-19 patients: A French multicentre cohort study. Eur. Heart J. 2020, 41, 3058–3068. [Google Scholar] [CrossRef]
- Leutner, B.F.; Reineking, B.; Müller, J.; Bachmann, M.; Beierkuhnlein, C.; Dech, S.; Wegmann, M. Modelling forest α-diversity and floristic composition—On the added value of LiDAR plus hyperspectral remote sensing. Remote Sens. 2012, 4, 2818–2845. [Google Scholar] [CrossRef] [Green Version]
- Hakkenberg, C.R.; Zhu, K.; Peet, R.K.; Song, C. Mapping multi-scale vascular plant richness in a forest landscape with integrated Li DAR and hyperspectral remote-sensing. Ecology 2018, 99, 474–487. [Google Scholar] [CrossRef]
- Chrysafis, I.; Korakis, G.; Kyriazopoulos, A.P.; Mallinis, G. Predicting Tree Species Diversity Using Geodiversity and Sentinel-2 Multi-Seasonal Spectral Information. Sustainability 2020, 12, 9250. [Google Scholar] [CrossRef]
- Hauser, L.T.; Féret, J.-B.; An Binh, N.; van der Windt, N.; Sil, Â.F.; Timmermans, J.; Soudzilovskaia, N.A.; van Bodegom, P.M. Towards scalable estimation of plant functional diversity from Sentinel-2: In-situ validation in a heterogeneous (semi-)natural landscape. Remote Sens. Environ. 2021, 262, 112505. [Google Scholar] [CrossRef]
- Xi, Y.; Ren, C.; Tian, Q.; Ren, Y.; Dong, X.; Zhang, Z. Exploitation of Time Series Sentinel-2 Data and Different Machine Learning Algorithms for Detailed Tree Species Classification. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2021, 14, 7589–7603. [Google Scholar] [CrossRef]
- Chen, C.; Ma, Y.; Ren, G.; Wang, J. Aboveground biomass of salt-marsh vegetation in coastal wetlands: Sample expansion of in situ hyperspectral and Sentinel-2 data using a generative adversarial network. Remote Sens. Environ. 2022, 270, 112885. [Google Scholar] [CrossRef]
- Shamshiri, R.; Eide, E.; Høyland, K.V. Spatio-temporal distribution of sea-ice thickness using a machine learning approach with Google Earth Engine and Sentinel-1 GRD data. Remote Sens. Environ. 2022, 270, 112851. [Google Scholar] [CrossRef]
- Kowalski, K.; Senf, C.; Hostert, P.; Pflugmacher, D. Characterizing spring phenology of temperate broadleaf forests using Landsat and Sentinel-2 time series. Int. J. Appl. Earth Obs. Geoinf. 2020, 92, 102172. [Google Scholar] [CrossRef]
- Li, H.; Kato, T.; Hayashi, M.; Wu, L. Estimation of Forest Aboveground Biomass of Two Major Conifers in Ibaraki Prefecture, Japan, from PALSAR-2 and Sentinel-2 Data. Remote Sens. 2022, 14, 468. [Google Scholar] [CrossRef]
- Coyle, D.B.; Stysley, P.R.; Poulios, D.; Clarke, G.B.; Kay, R.B. Laser transmitter development for NASA’s Global Ecosystem Dynamics Investigation (GEDI) lidar. In Lidar Remote Sensing for Environmental Monitoring XV; SPIE: Bellingham, WA, USA, 2015; pp. 19–25. [Google Scholar] [CrossRef]
- Qi, W.; Dubayah, R.O. Combining Tandem-X InSAR and simulated GEDI lidar observations for forest structure mapping. Remote Sens. Environ. 2016, 187, 253–266. [Google Scholar] [CrossRef]
- Lang, N.; Kalischek, N.; Armston, J.; Schindler, K.; Dubayah, R.; Wegner, J. Global canopy height regression and uncertainty estimation from GEDI LIDAR waveforms with deep ensembles. Remote Sens. Environ. 2022, 268, 112760. [Google Scholar] [CrossRef]
- Potapov, P.; Li, X.; Hernandez-Serna, A.; Tyukavina, A.; Hansen, M.C.; Kommareddy, A.; Pickens, A.; Turubanova, S.; Tang, H.; Silva, C.E.; et al. Mapping Global Forest Canopy Height Through Integration of GEDI and Landsat Data. Remote Sens. Environ. 2020, 253, 112165. [Google Scholar] [CrossRef]
- Liang, M.; Duncanson, L.; Silva, J.A.; Sedano, F. Quantifying aboveground biomass dynamics from charcoal degradation in Mozambique using GEDI Lidar and Landsat. Remote Sens. Environ. 2023, 284, 113367. [Google Scholar] [CrossRef]
- Zeng, W.; Tomppo, E.; Healey, S.P.; Gadow, K.V. The national forest inventory in China: History—results—international context. For. Ecosyst. 2015, 2, 23. [Google Scholar] [CrossRef] [Green Version]
- Mi, X.; Guo, J.; Hao, Z.; Xie, Z.; Guo, K.; Ma, K. Chinese forest biodiversity monitoring: Scientific foundations and strategic planning. Biodivers. Sci. 2016, 24, 1203. [Google Scholar] [CrossRef] [Green Version]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- Simpson, E.H. Measurement of diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Pielou, E.C. The measurement of diversity in different types of biological collections. J. Theor. Biol. 1966, 13, 131–144. [Google Scholar] [CrossRef]
- Bereta, K.; Caumont, H.; Daniels, U.; Goor, E.; Koubarakis, M.; Pantazi, D.-A.; Stamoulis, G.; Ubels, S.; Venus, V.; Wahyudi, F. The Copernicus App Lab Project: Easy Access to Copernicus Data; EDBT: Lisbon, Portugal, 2019; pp. 501–511. [Google Scholar]
- Main-Knorn, M.; Pflug, B.; Louis, J.; Debaecker, V.; Müller-Wilm, U.; Gascon, F. Sen2Cor for sentinel-2. In Image and Signal Processing for Remote Sensing XXIII; SPIE Remote Sensing: Warsaw, Poland, 2017; pp. 37–48. [Google Scholar]
- Zuhlke, M.; Fomferra, N.; Brockmann, C.; Peters, M.; Veci, L.; Malik, J.; Regner, P. SNAP (sentinel application platform) and the ESA sentinel 3 toolbox. In Sentinel-3 for Science Workshop; ESA: Venice, Italy, 2015; p. 21. [Google Scholar]
- Rouse, J.W., Jr.; Haas, R.H.; Deering, D.W.; Schell, J.A.; Harlan, J.C. Monitoring the Vernal Advancement and Retrogradation (Green Wave Effect) of Natural Vegetation; Texas A&M university: College Station, TX, USA, 1974; No. E75-10354. [Google Scholar]
- Gao, G.; Ting-Toomey, S.; Gudykunst, W.B. Chinese Communication Processes; Oxford University Press: Oxford, UK, 1996. [Google Scholar]
- Tucker, C.J.; Elgin Jr, J.H.; McMurtrey Iii, J.E.; Fan, C.J. Monitoring corn and soybean crop development with hand-held radiometer spectral data. Remote Sens. Environ. 1979, 8, 237–248. [Google Scholar] [CrossRef]
- Huete, A.; Didan, K.; Miura, T.; Rodriguez, E.P.; Gao, X.; Ferreira, L.G. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar] [CrossRef]
- Huete, A.R. A soil-adjusted vegetation index (SAVI). Remote Sens. Environ. 1988, 25, 295–309. [Google Scholar] [CrossRef]
- Hancock, S.; Armston, J.; Hofton, M.; Sun, X.; Tang, H.; Duncanson, L.I.; Kellner, J.R.; Dubayah, R. The GEDI Simulator: A Large-Footprint Waveform Lidar Simulator for Calibration and Validation of Spaceborne Missions. Earth Space Sci. 2019, 6, 294–310. [Google Scholar] [CrossRef] [PubMed]
- MacArthur, R.H.; Horn, H.S. Foliage profile by vertical measurements. Ecology 1969, 50, 802–804. [Google Scholar] [CrossRef] [Green Version]
- Lu, G.Y.; Wong, D.W. An adaptive inverse-distance weighting spatial interpolation technique. Comput. Geosci. 2008, 34, 1044–1055. [Google Scholar] [CrossRef]
- Bartier, P.M.; Keller, C.P. Multivariate interpolation to incorporate thematic surface data using inverse distance weighting (IDW). Comput. Geosci. 1996, 22, 795–799. [Google Scholar] [CrossRef]
- Hallman, T.A.; Robinson, W.D. Comparing multi-and single-scale species distribution and abundance models built with the boosted regression tree algorithm. Landsc. Ecol. 2020, 35, 1161–1174. [Google Scholar] [CrossRef]
- Valbuena, R.; Eerikäinen, K.; Packalen, P.; Maltamo, M. Gini coefficient predictions from airborne lidar remote sensing display the effect of management intensity on forest structure. Ecol. Indic. 2016, 60, 574–585. [Google Scholar] [CrossRef]
- Maxwell, A.E.; Warner, T.A.; Fang, F. Implementation of machine-learning classification in remote sensing: An applied review. Int. J. Remote Sens. 2018, 39, 2784–2817. [Google Scholar] [CrossRef] [Green Version]
- Vafaei, S.; Soosani, J.; Adeli, K.; Fadaei, H.; Naghavi, H.; Pham, T.; Tien Bui, D. Improving Accuracy Estimation of Forest Aboveground Biomass Based on Incorporation of ALOS-2 PALSAR-2 and Sentinel-2A Imagery and Machine Learning: A Case Study of the Hyrcanian Forest Area (Iran). Remote Sens. 2018, 10, 172. [Google Scholar] [CrossRef]
- Mitchell, M.W. Bias of the Random Forest Out-of-Bag (OOB) Error for Certain Input Parameters. Open J. Stat. 2011, 1, 205–211. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Ren, C.; Zhang, B.; Wang, Z.; Liu, M.; Man, W.; Liu, J. Improved estimation of forest stand volume by the integration of GEDI LiDAR data and multi-sensor imagery in the Changbai Mountains Mixed forests Ecoregion (CMMFE), northeast China. Int. J. Appl. Earth Obs. Geoinf. 2021, 100, 102326. [Google Scholar] [CrossRef]
- Hastie, T.; Tibshirani, R.; Friedman, J.H.; Friedman, J.H. The Elements of Statistical Learning: Data Mining, Inference, and Prediction; Springer: New York, NY, USA, 2009; Volume 2, pp. 1–758. [Google Scholar]
- Fauvel, M.; Lopes, M.; Dubo, T.; Rivers-Moore, J.; Frison, P.-L.; Gross, N.; Ouin, A. Prediction of plant diversity in grasslands using Sentinel-1 and -2 satellite image time series. Remote Sens. Environ. 2020, 237, 111536. [Google Scholar] [CrossRef]
- Yue, J.; Tian, Q.; Dong, X.; Xu, N. Using broadband crop residue angle index to estimate the fractional cover of vegetation, crop residue, and bare soil in cropland systems. Remote Sens. Environ. 2020, 237, 111538. [Google Scholar] [CrossRef]
- Sothe, C.; Almeida, C.; Liesenberg, V.; Schimalski, M. Evaluating Sentinel-2 and Landsat-8 Data to Map Sucessional Forest Stages in a Subtropical Forest in Southern Brazil. Remote Sens. 2017, 9, 838. [Google Scholar] [CrossRef] [Green Version]
- Grabska, E.; Frantz, D.; Ostapowicz, K. Evaluation of machine learning algorithms for forest stand species mapping using Sentinel-2 imagery and environmental data in the Polish Carpathians. Remote Sens. Environ. 2020, 251, 112103. [Google Scholar] [CrossRef]
- Fassnacht, F.E.; Latifi, H.; Stereńczak, K.; Modzelewska, A.; Lefsky, M.; Waser, L.T.; Straub, C.; Ghosh, A. Review of studies on tree species classification from remotely sensed data. Remote Sens. Environ. 2016, 186, 64–87. [Google Scholar] [CrossRef]
- Belgiu, M.; Drăguţ, L. Random forest in remote sensing: A review of applications and future directions. ISPRS J. Photogramm. Remote Sens. 2016, 114, 24–31. [Google Scholar] [CrossRef]
- Wessel, M.; Brandmeier, M.; Tiede, D. Evaluation of Different Machine Learning Algorithms for Scalable Classification of Tree Types and Tree Species Based on Sentinel-2 Data. Remote Sens. 2018, 10, 1419. [Google Scholar] [CrossRef] [Green Version]
- Kuter, S. Completing the machine learning saga in fractional snow cover estimation from MODIS Terra reflectance data: Random forests versus support vector regression. Remote Sens. Environ. 2021, 255, 112294. [Google Scholar] [CrossRef]
- Poorazimy, M.; Shataee, S.; McRoberts, R.E.; Mohammadi, J. Integrating airborne laser scanning data, space-borne radar data and digital aerial imagery to estimate aboveground carbon stock in Hyrcanian forests, Iran. Remote Sens. Environ. 2020, 240, 111669. [Google Scholar] [CrossRef]
- Latifi, H.; Nothdurft, A.; Koch, B. Non-parametric prediction and mapping of standing timber volume and biomass in a temperate forest: Application of multiple optical/LiDAR-derived predictors. Forestry 2010, 83, 395–407. [Google Scholar] [CrossRef] [Green Version]
- Cesarz, S.; Ruess, L.; Jacob, M.; Jacob, A.; Schaefer, M.; Scheu, S. Tree species diversity versus tree species identity: Driving forces in structuring forest food webs as indicated by soil nematodes. Soil Biol. Biochem. 2013, 62, 36–45. [Google Scholar] [CrossRef]
- Peng, Y.; Fan, M.; Song, J.; Cui, T.; Li, R. Assessment of plant species diversity based on hyperspectral indices at a fine scale. Sci. Rep. 2018, 8, 4776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabrero-González, C.; Garrido-Almonacid, A.; Esquivel, F.J.; Cámara-Serrano, J.A. A model of spatial location: New data for the Gor River megalithic landscape (Spain) from LiDAR technology and field survey. Archaeol. Prospect. 2022, 1–15. [Google Scholar] [CrossRef]
- Lechner, A.M.; Foody, G.M.; Boyd, D.S. Applications in remote sensing to forest ecology and management. One Earth 2020, 2, 405–412. [Google Scholar] [CrossRef]







| Diversity Index | Equation | Reference | Description |
|---|---|---|---|
| Shannon index (H′, based e) | [27] | Species richness and equitability in distribution in a plot | |
| Simpson index (λ form) | [28] | The dominance of a species in a plot | |
| Pielou evenness index (J′) | [29] | How close in numbers each species in a plot |
| Vegetation Indices | Expression | References |
|---|---|---|
| Normalized Difference Vegetation Index (NDVI) | [33] | |
| Normalized Difference Water Index (NDWI) | [34] | |
| Difference Vegetation Index (DVI) | [35] | |
| Enhanced Vegetation Index (EVI) | [36] | |
| Soil Adjusted Vegetation Index (SAVI) | [37] |
| Data Type | Variables | Time | Description |
|---|---|---|---|
| Sentinel-2 | B1 | May. Jun. Sep. and Oct. | Coastal aerosol, 443 nm |
| B2 | May. Jun. Sep. and Oct. | Blue, 490 nm | |
| B3 | May. Jun. Sep. and Oct. | Green, 560 nm | |
| B4 | May. Jun. Sep. and Oct. | Red, 665 nm | |
| B5 | May. Jun. Sep. and Oct. | Red edge, 705 nm | |
| B6 | May. Jun. Sep. and Oct. | Red edge, 740 nm | |
| B7 | May. Jun. Sep. and Oct. | Red edge, 783 nm | |
| B8 | May. Jun. Sep. and Oct. | Near infrared, 842 nm | |
| B8A | May. Jun. Sep. and Oct. | Near infrared, 865 nm | |
| B11 | May. Jun. Sep. and Oct. | Short-wave infrared, 1610 nm | |
| B12 | May. Jun. Sep. and Oct. | Short-wave infrared, 2190 nm | |
| Vegetation indices | NDVI | May. Jun. Sep. and Oct. | Normalized Difference Vegetation Index |
| NDWI | May. Jun. Sep. and Oct. | Normalized Difference Water Index | |
| EVI | May. Jun. Sep. and Oct. | Enhanced Vegetation Index | |
| DVI | May. Jun. Sep. and Oct. | Difference Vegetation Index | |
| SAVI | May. Jun. Sep. and Oct. | Soil Adjusted Vegetation Index | |
| GEDI LiDAR | FHD_NGS | Non-growing season | Foliage height diversity in non-growing season |
| FHD_GS | Growing season | Foliage height diversity in growing season | |
| PAI_NGS | Non-growing season | Plant area index in non-growing season | |
| PAI_GS | Growing season | Plant area index in growing season |
| Model | Abbr. | Parameters | Feature Rank Criteria |
|---|---|---|---|
| Lasso regression | LR | — | Absolute value of coefficients |
| K-Nearest Neighbors | KNN | K values = 3, 5, 7, 9, 11 | Minimum error rate |
| Support Vector Machine | SVM | cost = 0.1, 0.5, 1, 2, 4, 10 | Squared weights |
| kernel = linear, radial, sigmoid, rbf. | |||
| Random Forest | RF | ntree = 200, 500, 800, 1000 | Increase in mean squared error by permuting a variable |
| mtry = 2, 5, 10, 20, or k/3 |
| Combined Variables | H′ Index | λ Index | J′ Index | |||
|---|---|---|---|---|---|---|
| R2 | RMSE | R2 | RMSE | R2 | RMSE | |
| GEDI | 0.51 | 0.78 | 0.54 | 0.26 | 0.48 | 0.35 |
| Sentinel-2 &VIs | 0.66 | 0.56 | 0.57 | 0.15 | 0.63 | 0.18 |
| GEDI & Sentinel-2 &VIs | 0.72 | 0.46 | 0.78 | 0.14 | 0.86 | 0.11 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ren, C.; Jiang, H.; Xi, Y.; Liu, P.; Li, H. Quantifying Temperate Forest Diversity by Integrating GEDI LiDAR and Multi-Temporal Sentinel-2 Imagery. Remote Sens. 2023, 15, 375. https://doi.org/10.3390/rs15020375
Ren C, Jiang H, Xi Y, Liu P, Li H. Quantifying Temperate Forest Diversity by Integrating GEDI LiDAR and Multi-Temporal Sentinel-2 Imagery. Remote Sensing. 2023; 15(2):375. https://doi.org/10.3390/rs15020375
Chicago/Turabian StyleRen, Chunying, Hailing Jiang, Yanbiao Xi, Pan Liu, and Huiying Li. 2023. "Quantifying Temperate Forest Diversity by Integrating GEDI LiDAR and Multi-Temporal Sentinel-2 Imagery" Remote Sensing 15, no. 2: 375. https://doi.org/10.3390/rs15020375
APA StyleRen, C., Jiang, H., Xi, Y., Liu, P., & Li, H. (2023). Quantifying Temperate Forest Diversity by Integrating GEDI LiDAR and Multi-Temporal Sentinel-2 Imagery. Remote Sensing, 15(2), 375. https://doi.org/10.3390/rs15020375