
Distribution Modeling and Gap Analysis of Shorebird Conservation in Northern Brazil

 , ,
, ,
Abstract
:1. Introduction
- To what extent is the habitat of migratory shorebirds currently protected by existing Marine Extractive Reserves?
- Where are important shorebird habitat sites that exist outside current protections?
- How does shifting the conservation priorities between umbrella, flagship, and biodiversity lenses shift the selection of new protected sites?
2. Study Area and Methods
2.1. Study Area: Northern Coast of Brazil
2.2. Survey Methods
2.3. Environmental Parameters
2.4. Species Distribution Modeling
2.5. Validation
2.6. Thresholding Predicted Specie Presence
2.7. Comparing Conservation Strategies
2.8. Identifying New Conservation Zones
3. Results
3.1. Model AUC and Validation Results
3.2. Analysis of Shorebird Habitat within Marine Extractive Reserves
3.3. Comparison of Conservation Strategies
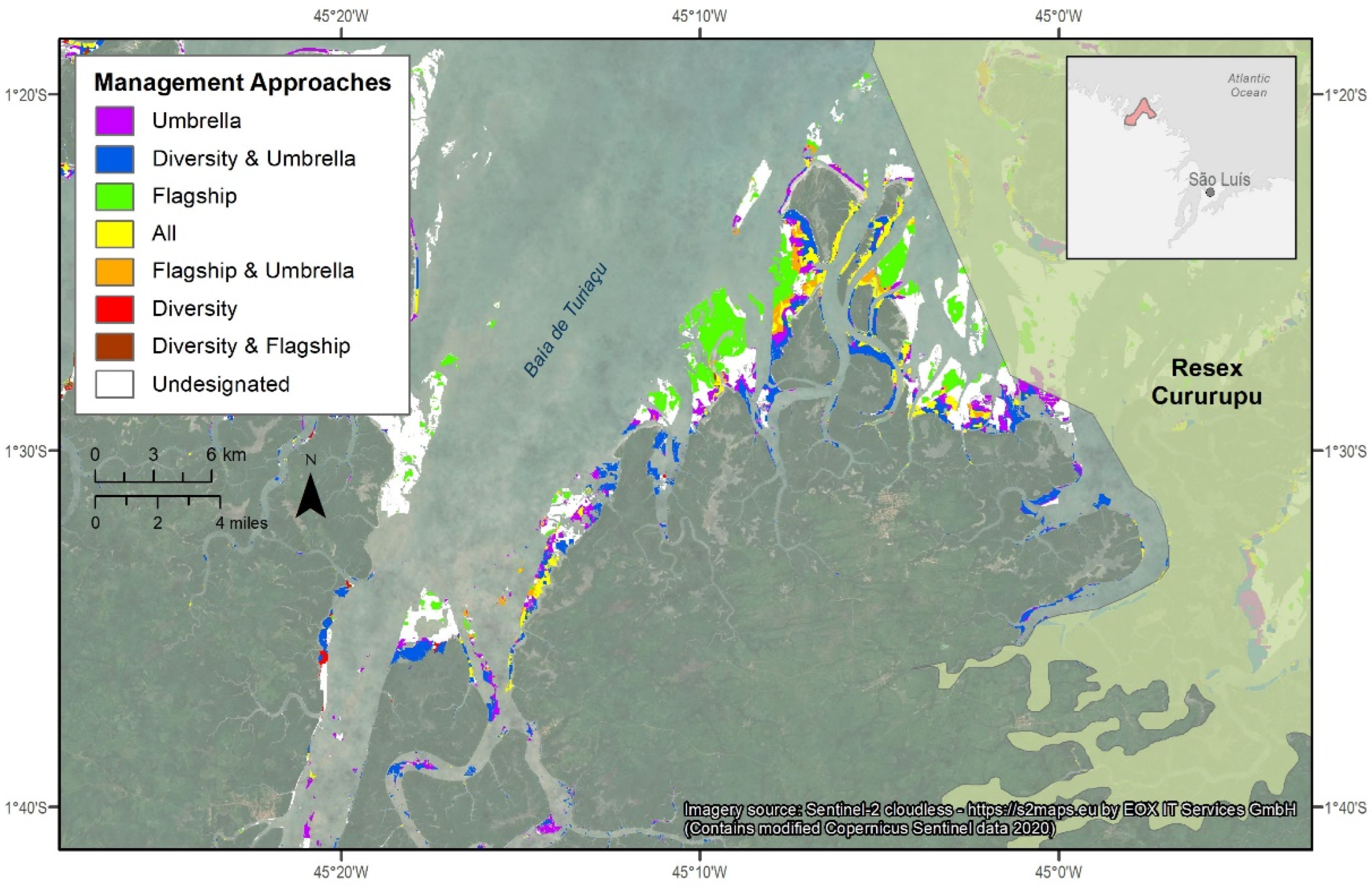
3.4. Identifying New Conservation Zones
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
- Distance to rivers
- Distance to developed areas
- Distance to mangroves
- Tidal exposure
- Permanence
References
- Colwell, M.A. Shorebird Ecology, Conservation, and Management; University of California Press: Berkeley, CA, USA, 2010. [Google Scholar]
- Niles, L.J.; Burger, J.; Porter, R.R.; Dey, A.D.; Minton, C.D.; González, P.M.; Baker, A.J.; Fox, J.W.; Gordon, C. First results using light level geolocators to track Red Knots in the Western Hemisphere show rapid and long intercontinental flights and new details of migration pathways. Wader Study Group Bull. 2010, 117, 123–130. [Google Scholar]
- Gratto-Trevor, C.; Morrison, R.I.G.; Mizrahi, D.; Lank, D.B.; Hicklin, P.; Spaans, A.L. Migratory Connectivity of Semipalmated Sandpipers: Winter Distribution and Migration Routes of Breeding Populations. Waterbirds 2012, 35, 83–95. [Google Scholar] [CrossRef]
- Brown, S.; Gratto-Trevor, C.; Porter, R.; Weiser, E.; Mizrahi, D.; Bentzen, R.; Boldenow, M.; Clay, R.; Freeman, S.; Giroux, M.-A.; et al. Migratory connectivity of Semipalmated Sandpipers and implications for conservation. Condor 2017, 119, 207–224. [Google Scholar] [CrossRef] [Green Version]
- Warnock, N. Stopping vs. staging: The difference between a hop and a jump. J. Avian Biol. 2010, 41, 621–626. [Google Scholar] [CrossRef]
- Mizrahi, D.S.; Peters, K.A.; Hodgetts, P.A. Energetic Condition of Semipalmated and Least Sandpipers during Northbound Migration Staging Periods in Delaware Bay. Waterbirds 2012, 35, 135–145. [Google Scholar] [CrossRef]
- Piersma, T.; Lindström, Å. Migrating shorebirds as integrative sentinels of global environmental change: Shorebirds integrate global environmental information. Ibis 2004, 146, 61–69. [Google Scholar]
- Clemens, R.; Rogers, D.I.; Hansen, B.D.; Gosbell, K.; Minton, C.D.T.; Straw, P.; Bamford, M.; Woehler, E.J.; Milton, D.A.; Weston, M.A.; et al. Continental-scale decreases in shorebird populations in Australia. Emu-Austral Ornithol. 2016, 116, 119–135. [Google Scholar] [CrossRef] [Green Version]
- Studds, C.E.; Kendall, B.E.; Murray, N.J.; Wilson, H.B.; Rogers, D.I.; Clemens, R.S.; Gosbell, K.; Hassell, C.J.; Jessop, R.; Melville, D.S.; et al. Rapid population decline in migratory shorebirds relying on Yellow Sea tidal mudflats as stopover sites. Nat. Commun. 2017, 8, 14895. [Google Scholar] [CrossRef] [Green Version]
- Bart, J.; Brown, S.; Harrington, B.; Morrison, R.I.G. Survey trends of North American shorebirds: Population declines or shifting distributions? J. Avian Biol. 2007, 38, 73–82. [Google Scholar] [CrossRef]
- Andres, B.A.; Gratto-Trevor, C.; Hicklin, P.; Mizrahi, D.; Morrison, R.I.G.; Smith, P.A. Status of the Semipalmated Sandpiper. Waterbirds 2012, 35, 146–148. [Google Scholar] [CrossRef]
- Baker, A.J.; González, P.M.; Piersma, T.; Niles, L.J.; Nascimento, I.D.L.S.D.; Atkinson, P.; Clark, N.A.; Minton, C.D.T.; Peck, M.K.; Aarts, G. Rapid population decline in red knots: Fitness consequences of decreased refuelling rates and late arrival in Delaware Bay. Proc. R. Soc. B Biol. Sci. 2004, 271, 875–882. [Google Scholar] [CrossRef] [Green Version]
- Burger, J.; Niles, L.J.; Porter, R.R.; Dey, A.D.; Koch, S.L.; Gordon, C. Migration and Over-Wintering of Red Knots (Calidris canutus rufa) along the Atlantic Coast of the United States. Condor 2012, 114, 302–313. [Google Scholar] [CrossRef]
- Smith, J.A.M.; Niles, L.J.; Hafner, S.; Modjeski, A.; Dillingham, T. Beach restoration improves habitat quality for American horseshoe crabs and shorebirds in the Delaware Bay, USA. Mar. Ecol. Prog. Ser. 2020, 645, 91–107. [Google Scholar] [CrossRef]
- Burger, J.; Niles, L.; Jeitner, C.; Gochfeld, M. Habitat risk: Use of intertidal flats by foraging red knots (Calidris canutus rufa), ruddy turnstones, (Arenaria interpres), semipalmated sandpipers (Calidris pusilla), and sanderling (Calidris alba) on Delaware Bay beaches. Environ. Res. 2018, 165, 237–246. [Google Scholar]
- Niles, L.; Cooper Ornithological Society. Status of the Red Knot (Calidris canutus rufa) in the Western Hemisphere; Cooper Ornithological Society: Camarillo, CA, USA, 2008. [Google Scholar]
- Shrader-Frechette, K.S.; McCoy, E.D. Method in Ecology: Strategies for Conservation; Cambridge University Press: Cambridge, UK; New York, NY, USA, 1993. [Google Scholar]
- Wilcove, D. Getting Ahead of the Extinction Curve. Ecol. Appl. 1993, 3, 218–220. [Google Scholar] [CrossRef]
- Simberloff, D. Flagships, umbrellas, and keystones: Is single-species management passé in the landscape era? Biol. Conserv. 1998, 83, 247–257. [Google Scholar]
- Caro, T.M.; O’Doherty, G. On the Use of Surrogate Species in Conservation Biology. Conserv. Biol. 1999, 13, 805–814. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- Rodrigues, A.A.F. Seasonal Abundance Of Neartic Shorebirds In The Gulf Of Maranhão, Brazil. J. Field Ornithol. 2000, 71, 665–675. [Google Scholar] [CrossRef]
- Argo, A. Crash: A Tale of Two Species; Public Broadcasting Service: Crystal City, VA, USA, 2008. [Google Scholar]
- US Fish and Wildlife Service. Endangered and Threatened Wildlife and Plants; Proposed Threatened Status for the Rufa Red Knot (Calidris canutus rufa). 2013. Available online: https://www.gpo.gov/fdsys/pkg/FR-2013-09-30/pdf/2013-22700.pdf (accessed on 1 December 2022).
- De Moura, R.L.; Minte-Vera, C.V.; Curado, I.B.; Francini-Filho, R.B.; Rodrigues, H.D.C.L.; Dutra, G.F.; Alves, D.C.; Souto, F.J.B. Challenges and Prospects of Fisheries Co-Management under a Marine Extractive Reserve Framework in Northeastern Brazil. Coast. Manag. 2009, 37, 617–632. [Google Scholar] [CrossRef]
- Pimentel, M.A.S. Comunidades tradicionais em reservas extrativistas marinhas no estado do Pará: Conflitos e resistências. Ambientes 2019, 1, 191–218. [Google Scholar]
- Santos, C.Z.; Schiavetti, A. Assessment of the management in Brazilian Marine Extractive Reserves. Ocean Coast. Manag. 2014, 93, 26–36. [Google Scholar] [CrossRef]
- Kober, K.; Bairlein, F. Habitat Choice and Niche Characteristics Under Poor Food Conditions. A Study on Migratory Nearctic Shorebirds in the Intertidal Flats of Brazil. Ardea 2009, 97, 31–42. [Google Scholar] [CrossRef]
- Mu, T.; Wilcove, D.S. Upper tidal flats are disproportionately important for the conservation of migratory shorebirds. Proc. R. Soc. B Boil. Sci. 2020, 287, 20200278. [Google Scholar] [CrossRef]
- Yates, M.; Jones, A.; McGrorty, S.; Goss-Custard, J. The Use of Satellite Imagery to Determine the Distribution of Intertidal Surface Sediments of The Wash, England. Estuarine Coast. Shelf Sci. 1993, 36, 333–344. [Google Scholar] [CrossRef]
- Rainey, M.; Tyler, A.; Gilvear, D.; Bryant, R.; McDonald, P. Mapping intertidal estuarine sediment grain size distributions through airborne remote sensing. Remote Sens. Environ. 2003, 86, 480–490. [Google Scholar] [CrossRef]
- van der Wal, D.; Herman, P.M.; Dool, A.W.-V.D. Characterisation of surface roughness and sediment texture of intertidal flats using ERS SAR imagery. Remote Sens. Environ. 2005, 98, 96–109. [Google Scholar] [CrossRef]
- van der Wal, D.; Herman, P.M.J. Regression-based synergy of optical, shortwave infrared and microwave remote sensing for monitoring the grain-size of intertidal sediments. Remote Sens. Environ. 2007, 111, 89–106. [Google Scholar] [CrossRef]
- Gade, M.; Melchionna, S.; Stelzer, K.; Kohlus, J. Multi-frequency SAR data help improving the monitoring of intertidal flats on the German North Sea coast. Estuarine Coast. Shelf Sci. 2014, 140, 32–42. [Google Scholar] [CrossRef]
- Zhang, K.; Dong, X.; Liu, Z.; Gao, W.; Hu, Z.; Wu, G. Mapping Tidal Flats with Landsat 8 Images and Google Earth Engine: A Case Study of the China’s Eastern Coastal Zone circa 2015. Remote Sens. 2019, 11, 924. [Google Scholar] [CrossRef] [Green Version]
- Henriques, M.; Catry, T.; Belo, J.R.; Piersma, T.; Pontes, S.; Granadeiro, J.P. Combining Multispectral and Radar Imagery with Machine Learning Techniques to Map Intertidal Habitats for Migratory Shorebirds. Remote Sens. 2022, 14, 3260. [Google Scholar] [CrossRef]
- Lathrop, R.G.; Merchant, D.; Niles, L.; Paludo, D.; Santos, C.D.; Larrain, C.E.; Feigin, S.; Smith, J.; Dey, A. Multi-Sensor Remote Sensing of Intertidal Flat Habitats for Migratory Shorebird Conservation. Remote Sens. 2022, 14, 5016. [Google Scholar] [CrossRef]
- Martinuzzi, S.; Gould, W.A.; Gonzalez, O.M.R. Creating Cloud-Free Landsat ETM+ Data Sets in Tropical Landscapes: Cloud and Cloud-Shadow Removal; U.S. Department of Agriculture, Forest Service, International Institute of Tropical Forestry Gen. Tech. Rep.: Washington, DC, USA, 2007.
- Murray, N.J.; Phinn, S.R.; Clemens, R.S.; Roelfsema, C.M.; Fuller, R.A. Continental Scale Mapping of Tidal Flats across East Asia Using the Landsat Archive. Remote Sens. 2012, 4, 3417–3426. [Google Scholar] [CrossRef] [Green Version]
- Egbert, G.D.; Erofeeva, S.Y. Efficient Inverse Modeling of Barotropic Ocean Tides. J. Atmos. Ocean. Technol. 2002, 19, 183–204. [Google Scholar] [CrossRef]
- Harris, I.; Jones, P.D.; Osborn, T.J.; Lister, D.H. Updated high-resolution grids of monthly climatic observations—The CRU TS3.10 Dataset. Int. J. Climatol. 2014, 34, 623–642. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Guisan, A.; Zimmermann, N.E. Predictive habitat distribution models in ecology. Ecol. Model. 2000, 135, 147–186. [Google Scholar] [CrossRef]
- Elith, J.; Leathwick, J.R. Species Distribution Models: Ecological Explanation and Prediction Across Space and Time. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists: Statistical explanation of MaxEnt. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Golicher, D.; Ford, A.; Cayuela, L.; Newton, A. Pseudo-absences, pseudo-models and pseudo-niches: Pitfalls of model selection based on the area under the curve. Int. J. Geogr. Inf. Sci. 2012, 26, 2049–2063. [Google Scholar] [CrossRef]
- Cao, Y.; DeWalt, R.E.; Robinson, J.L.; Tweddale, T.; Hinz, L.; Pessino, M. Using Maxent to model the historic distributions of stonefly species in Illinois streams: The effects of regularization and threshold selections. Ecol. Model. 2013, 259, 30–39. [Google Scholar] [CrossRef]
- Holmes, I.; McLaren, K.; Wilson, B. Niche modeling for management-ready information in little-studied, threatened frog species assemblages. J. Nat. Conserv. 2015, 28, 26–34. [Google Scholar] [CrossRef]
- Scott, J.M.; Davis, F.; Csuti, B.; Noss, R.; Butterfield, B.; Groves, C.; Anderson, H.; Caicco, S.; D’Erchia, F.; Edwards, T.C.; et al. Gap Analysis: A Geographic Approach to Protection of Biological Diversity. Wildl. Monogr. 1993, 123, 3–41. [Google Scholar]
- Elith, J.H.; Graham, C.P.H.; Anderson, R.P.; Dudík, M.; Ferrier, S.; Guisan, A.; Hijmans, R.J.; Huettmann, F.; Leathwick, J.R.; Lehmann, A.; et al. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef] [Green Version]
- Phillips, S.J.; Dudík, M.; Elith, J.; Graham, C.; Lehmann, A.; Leathwick, J.; Ferrier, S. Sample selection bias and presence-only distribution models: Implications for background and pseudo-absence data. Ecol. Appl. 2009, 19, 181–197. [Google Scholar] [CrossRef]
- Morrison, R.I.G.; Mizrahi, D.S.; Ross, R.K.; Ottema, O.H.; de Pracontal, N.; Narine, A. Dramatic Declines of Semipalmated Sandpipers on their Major Wintering Areas in the Guianas, Northern South America. Waterbirds 2012, 35, 120–134. [Google Scholar] [CrossRef]
- Smith, J.A.M.; Regan, K.; Cooper, N.W.; Johnson, L.; Olson, E.; Green, A.; Tash, J.; Evers, D.C.; Marra, P.P. A green wave of saltmarsh productivity predicts the timing of the annual cycle in a long-distance migratory shorebird. Sci. Rep. 2020, 10, 20658. [Google Scholar] [CrossRef]
- Baker, M.C.; Baker, A.E.M. Niche Relationships Among Six Species of Shorebirds on Their Wintering and Breeding Ranges. Ecol. Monogr. 1973, 43, 193–212. [Google Scholar] [CrossRef]
- Emlen, J.M. The Role of Time and Energy in Food Preference. Am. Nat. 1966, 100, 611–617. [Google Scholar] [CrossRef]
- MacArthur, R.H.; Pianka, E.R. On Optimal Use of a Patchy Environment. Am. Nat. 1966, 100, 603–609. [Google Scholar] [CrossRef] [Green Version]
- Zwarts, L.; Wanink, J.H. How the food supply harvestable by waders in the Wadden Sea depends on the variation in energy density, body weight, biomass, burying depth and behaviour of tidal-flat invertebrates. Neth. J. Sea Res. 1993, 31, 441–476. [Google Scholar] [CrossRef]
- Murray, N.J.; Fuller, R.A. Protecting stopover habitat for migratory shorebirds in East Asia. J. Ornithol. 2015, 156, 217–225. [Google Scholar] [CrossRef]
- Dhanjal-Adams, K.L.; Hanson, J.O.; Murray, N.J.; Phinn, S.R.; Wingate, V.R.; Mustin, K.; Lee, J.R.; Allan, J.R.; Cappadonna, J.L.; Studds, C.E.; et al. The distribution and protection of intertidal habitats in Australia. Emu-Austral Ornithol. 2016, 116, 208–214. [Google Scholar] [CrossRef] [Green Version]
- Burger, J.; Carlucci, S.A.; Jeitner, C.W.; Niles, L. Habitat Choice, Disturbance, and Management of Foraging Shorebirds and Gulls at a Migratory Stopover. J. Coast. Res. 2007, 23, 1159. [Google Scholar] [CrossRef]
- Burger, J.; Niles, L. Shorebirds and stakeholders: Effects of beach closure and human activities on shorebirds at a New Jersey coastal beach. Urban Ecosyst. 2012, 16, 657–673. [Google Scholar] [CrossRef]
- Ottema, O.H.; Spaans, A.L. Challenges and advances in shorebird conservation in the Guianas, with a focus on Suriname. Ornitol. Neotrop. 2008, 19, 339–346. [Google Scholar]
- Andres, B.; Ydenberg, R.C.; Lank, D.B. Shorebird hunting in the Caribbean. In Proceedings of the Western Hemisphere Shorebird Group: Fourth Meeting, Vancouver, BC, Canada, 11–15 August 2011; pp. 11–15. [Google Scholar]
- Gavin, M.C.; McCarter, J.; Berkes, F.; Mead, A.T.P.; Sterling, E.J.; Tang, R.; Turner, N.J. Effective Biodiversity Conservation Requires Dynamic, Pluralistic, Partnership-Based Approaches. Sustainability 2018, 10, 1846. [Google Scholar] [CrossRef] [Green Version]
- de Vasconcellos Pegas, F.; Stronza, A. Ecotourism and sea turtle harvesting in a fishing village of Bahia, Brazil. Conserv. Soc. 2020, 8, 15–25. [Google Scholar] [CrossRef] [Green Version]
- Cusack, C.; Sethi, S.A.; Rice, A.N.; Warren, J.D.; Fujita, R.; Ingles, J.; Flores, J.; Garchitorena, E.; Mesa, S.V. Marine ecotourism for small pelagics as a source of alternative income generating activities to fisheries in a tropical community. Biol. Conserv. 2021, 261, 109242. [Google Scholar] [CrossRef]
- Riera, P.; Stal, L.; Nieuwenhuize, J. Heavy δ15N in Intertidal Benthic Algae and Invertebrates in the Scheldt Estuary (The Netherlands): Effect of River Nitrogen Inputs. Estuarine Coast. Shelf Sci. 2000, 51, 365–372. [Google Scholar] [CrossRef] [Green Version]
- Yamada, F.; Kobayashi, N.; Shirakawa, Y.; Watabe, Y.; Sassa, S.; Tamaki, A. Effects of Tide and River Discharge on Mud Transport on Intertidal Flat. J. Waterw. Port Coastal Ocean Eng. 2012, 138, 172–180. [Google Scholar] [CrossRef]
- Colwell, M.A.; Landrum, S.L. Nonrandom Shorebird Distribution and Fine-Scale Variation in Prey Abundance. Condor 1993, 95, 94–103. [Google Scholar] [CrossRef]
- Thrush, S.; Hewitt, J.; Norkko, A.; Nicholls, P.; Funnell, G.; Ellis, J. Habitat change in estuaries: Predicting broad-scale responses of intertidal macrofauna to sediment mud content. Mar. Ecol. Prog. Ser. 2003, 263, 101–112. [Google Scholar] [CrossRef] [Green Version]
- Kober, K.; Bairlein, F. Shorebirds of the Bragantinian Peninsula I. Prey Availability and Shorebird Consumption at a Tropical Site in Northern Brazil. Ornitol. Neotrop. 2006, 17, 531–548. [Google Scholar]
- Burger, J. The Effect of Human Activity on Shorebirds in Two Coastal Bays in Northeastern United States. Environ. Conserv. 1986, 13, 123–130. [Google Scholar] [CrossRef]
- Lunardi, V.O.; Macedo, R.H.; Granadeiro, J.P.; Palmeirim, J.M. Migratory flows and foraging habitat selection by shorebirds along the northeastern coast of Brazil: The case of Baía de Todos os Santos. Estuar. Coast. Shelf Sci. 2012, 96, 179–187. [Google Scholar] [CrossRef]
- Zwarts, L. Numbers and distribution of coastal waders in Guinea-Bissau. Ardea 1988, 76, 42–55. [Google Scholar]
- Granadeiro, J.P.; Dias, M.; Martins, R.C.; Palmeirim, J. Variation in numbers and behaviour of waders during the tidal cycle: Implications for the use of estuarine sediment flats. Acta Oecologica 2006, 29, 293–300. [Google Scholar] [CrossRef]
- Burger, J.; Howe, M.A.; Hahn, D.C.; Chase, J. Effects of Tide Cycles on Habitat Selection and Habitat Partitioning by Migrating Shorebirds. Ornithology 1977, 94, 743–758. [Google Scholar] [CrossRef]
- Leyrer, J.; Spaans, B.; Camara, M.; Piersma, T. Small home ranges and high site fidelity in red knots (Calidris c. canutus) wintering on the Banc d’Arguin, Mauritania. J. Ornithol. 2006, 147, 376–384. [Google Scholar] [CrossRef]
- Warnock, S.E.; Takekawa, J.Y. Wintering site fidelity and movement patterns of Western Sandpipers Calidris mauri in the San Francisco Bay estuary. Ibis 1996, 138, 160–167. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Common Name | Alpha Code | Latin Name | Number of Individuals Observed | Presence Points | Absence Points | ||
|---|---|---|---|---|---|---|---|
| Calibration | Validation | Total | Validation | ||||
| Black-bellied plover | BBPL | Pluvialis squatarola | 1126 | 126 | 83 | 209 | 159 |
| Red Knot | REKN | Calidris canutus rufa | 1447 | 26 * | 0 | 26 | 232 |
| Ruddy Turnstone | RUTU | Arenaria interpres | 1171 | 76 | 53 | 129 | 189 |
| Sanderling | SAND | Calidris alba | 2572 | 120 | 30 | 150 | 212 |
| Semipalmated Plover | SEPL | Charadrius semipalmatus | 1357 | 109 | 64 | 173 | 178 |
| Semipalmated Sandpiper | SESA | Calidris pusilla | 30,064 | 204 | 75 | 279 | 167 |
| Whimbrel | WHIM | Numenius phaeopus | 2205 | 183 | 105 | 288 | 140 |
| Willet | WILL | Tringa semipalmata | 1844 | 113 | 67 | 180 | 175 |
| AUC | t-Test | |
|---|---|---|
| BBPL | 0.781 | 0.3675 |
| REKN * | 0.743 | 0.0312 |
| RUTU | 0.761 | 0.0011 |
| SAND | 0.744 | 0.0237 |
| SEPL | 0.726 | 0.029 |
| SESA | 0.718 | 0.0034 |
| WHIM | 0.791 | <0.001 |
| WILL | 0.85 | <0.001 |
| # of Species | Total Intertidal | Within MER | ||
|---|---|---|---|---|
| Area (km2) | % of Total | Area (km2) | % of Category | |
| 0 | 261.1 | 19.5 | 83.2 | 31.9 |
| 1 | 358.6 | 26.8 | 85.3 | 23.8 |
| 2 | 217.5 | 16.3 | 59.6 | 27.4 |
| 3 | 131.3 | 9.8 | 33.2 | 25.3 |
| 4 | 108.9 | 8.1 | 27.8 | 25.6 |
| 5 | 103.2 | 7.7 | 20.8 | 20.2 |
| 6 | 103.4 | 7.7 | 19.2 | 18.6 |
| 7 | 47.8 | 3.6 | 14.6 | 30.5 |
| 8 | 5.41 | 0.40 | 3.13 | 57.76 |
| Total | 1337.2 | 346.8 | ||
| Weighted species richness | 1.89 | 2.89 | ||
| Flagship km2 (%) | Umbrella km2 (%) | Diversity km2 (%) | |
|---|---|---|---|
| In MER | 87.9 (44.3) | 82.3 (16.8) | 57.8 (22.2) |
| Out MER | 110.4 (55.7) | 406.0 (83.2) | 202.0 (77.8) |
| Total | 198.3 | 488.2 | 259.8 |
| Overall Weighted Species Richness | 3.23 | 4.35 | 5.83 |
| REKN Habitat | 198.3 (100) | 68.6 (34.6) | 53.5 (27.0) |
| Area km | Flagship km2 (%) | Umbrella km2 (%) | Biodiversity km2 (%) | |
|---|---|---|---|---|
| Baía do Cumã | 35.5 | 2.2 (6.1) | 30.7 (86.7) | 22.7 (64.0) |
| Baía de São José | 90.2 | 0.0 (0.0) | 65.2 (72.3) | 36.1 (40.0) |
| Resex Cururupu | 97.9 | 31.0 (31.6) | 44.1 (45.1) | 30.0 (30.6) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Merchant, D.; Lathrop, R.G.; Santos, C.D.; Paludo, D.; Niles, L.; Smith, J.A.M.; Feigin, S.; Dey, A. Distribution Modeling and Gap Analysis of Shorebird Conservation in Northern Brazil. Remote Sens. 2023, 15, 452. https://doi.org/10.3390/rs15020452
Merchant D, Lathrop RG, Santos CD, Paludo D, Niles L, Smith JAM, Feigin S, Dey A. Distribution Modeling and Gap Analysis of Shorebird Conservation in Northern Brazil. Remote Sensing. 2023; 15(2):452. https://doi.org/10.3390/rs15020452
Chicago/Turabian StyleMerchant, Daniel, Richard G. Lathrop, Carlos David Santos, Danielle Paludo, Larry Niles, Joseph A. M. Smith, Stephanie Feigin, and Amanda Dey. 2023. "Distribution Modeling and Gap Analysis of Shorebird Conservation in Northern Brazil" Remote Sensing 15, no. 2: 452. https://doi.org/10.3390/rs15020452
APA StyleMerchant, D., Lathrop, R. G., Santos, C. D., Paludo, D., Niles, L., Smith, J. A. M., Feigin, S., & Dey, A. (2023). Distribution Modeling and Gap Analysis of Shorebird Conservation in Northern Brazil. Remote Sensing, 15(2), 452. https://doi.org/10.3390/rs15020452