1. Introduction
Forests cover around 30% of the Earth’s land area [
1], including semi-arid areas. Iran is climatically diverse and encompasses a considerable portion of the world’s climate zones [
2]. However, the country is known to be dominated by semi-arid, arid, and fragile vegetation ecosystems [
3]. The Zagros mountains are home to a significant portion of Iran’s semi-arid forests and woodlands, which encompass > 40% of Iran’s forests, accounting for ca. six million ha [
4,
5]. These forests are particularly important from the socioeconomic, soil conservation, and water-quality perspectives, as well as for providing considerable non-market ecosystem services [
6]. However, their quantity and quality have been almost constantly deteriorating over the last few decades due to a variety of natural and human-induced factors [
7,
8,
9,
10,
11], including, in particular, the high level of dependency of local livelihood on these forests [
12,
13].
This close connection between people and nature in northern Zagros (located in the West Azerbaijan and Kurdistan provinces, Iran) [
14] has led to significant alterations in the vegetative composition [
15] and the primary structure of both trees and forest stands [
16]. Historically, the high dependence of local forest dwellers on natural resources has made inventive utilization of the forest to supply both human and livestock inevitable [
17]. Most forest dwellers practice some sort of silvopastoralism and highly depend on oak forests to produce forage for their livestock throughout the year [
18]. In particular, traditional animal husbandry in northern Zagros faces a lack of pastures and fodder in the cold seasons. To adapt their livelihoods to such limitations, the locals have developed a traditional silvopastoral method called pollarding (Kurdish: Galazani) [
6,
19]. Pollarding is a generic name for pruning tree branches to provide both fodder (for domestic animals) and fuelwood [
20]. Pollarding has been reported to be practiced in parts of Europe, Asia, Africa, and America, especially for the benefit of using leaves and branches as animal fodder. Pollarding is hence an important factor in livestock management and relates to this important source of income in the farming economy [
21,
22,
23,
24,
25]. Oak pollards are a part of traditional forest management in many ecosystems and have a very long history of about 4000 years [
26]. In Iran, and as a part of the traditional management system in the northern Zagros, each rural family traditionally owns, i.e., tenures, a part of the forest (Kurdish: Galajar) [
27]. Each Galajar is divided into three or four parcels with equal fodder productivity, termed Shanegala (in Kurdish) [
28]. This is due to the fact that a given Shanegala is used to perform pollarding for winter fodder each year. Each Shanegala is pollarded in a three or four year pollarding cycle [
4,
28], which annually occurs from mid-September to early October (pre-fall). However, pollarding is not carried out in this way everywhere and is carried out irregularly [
29]. During pollarding, forest dwellers cut leafy branches (Kurdish: Bakhe) of oak trees, mostly
Quercus libani Olivier and
Q. infectoria Olivier., and store them on large trees (Kurdish: Dargala), on the ground, or on rock fragments in a cone-shaped formation known as a Gala or a Loya Gala (in Kurdish). In winter, dried leafy Loya Gala branches are used to feed livestock (especially Markhoz goats and sheep) [
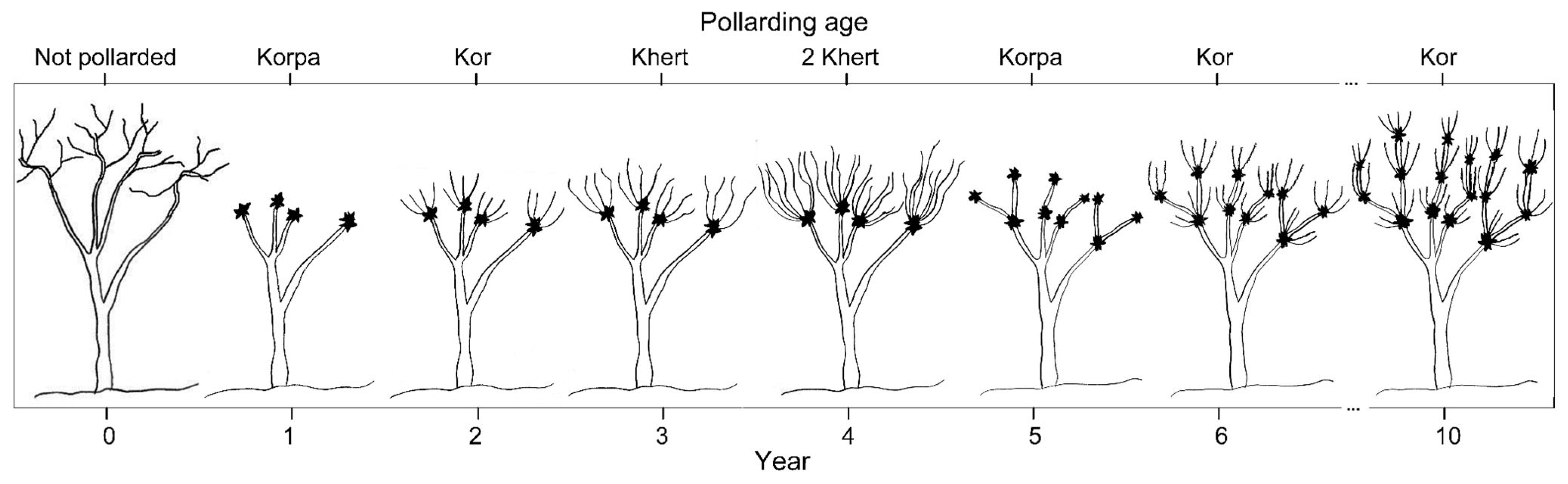
16]. The locals estimate pollarding ages to conduct the above-mentioned pollarding cycle. Whereas a freshly pollarded tree is called Korpa, the tree is termed Kor, Khert, and 2 Khert following one, two, and three years from the time of its pollarding, respectively [
30] (
Figure 1).
One of the initial consequences of pollarding is reducing the tree canopy, which causes stress on the tree and changes the typical pattern of tree growth [
31,
32]. However, some studies [
6,
16,
32,
33,
34] have shown that pollarding may result in a decrease in height, trunk, crown area, canopy vigor, health, seed and coppice regeneration, and density.
However, scientific knowledge on this traditional silvicultural approach is very sparse, and the exact effects of pollarding on tree structure and vitality are not yet fully understood (an overview of earlier studies focusing on this topic is summarized in
Table 1). The first step to thoroughly examining the pollarding approach and its temporal trajectories is to adequately capture the tree structure of pollarded trees [
35,
36]. Field inventories typically involve measuring the diameter at breast height (DBH), tree height, and the tree crown diameter [
37,
38] but without considering the intrinsic features of pollarded trees, like their distorted DBH~height and DBH~crown area relationships. With respect to potential monitoring approaches, timely field measurements are challenging, as pollarding typically takes place on mountain slopes [
16,
30,
39], a problem that is exacerbated not only by the subjectivity and required logistics for conducting field measurements (particularly for attributes like tree height) but also by the fact that measurements might be prevented or opposed by the local dwellers that assert a customary right to forest tenure over Galajars.
As an alternative or complement, remote sensing- and photogrammetry-based approaches may partly replace or amend field measurements [
40]. Remote sensing has been historically used in a variety of fields related to vegetation monitoring [
41,
42]. Well-known pattern recognition algorithms in combination with space-borne active [
43,
44,
45] and passive [
46,
47,
48] data have been largely employed yet face difficulties in small-scale applications. Estimating the structural variables of single trees can be accomplished using very-high-resolution (VHR) space-borne data [
49], which are, however, often prohibitive due to their high price and the lack of accessibility for politically sensitive areas like Iran. As an alternative, three-dimensional ground sensors (e.g., terrestrial laser scanners) [
50] or aerial sensors (e.g., aerial laser scanners) [
51] can be used. However, these tools are often either unavailable, costly, or practically infeasible for large-area assessments or subsequent spatial upscaling to larger domains [
52]. Consumer-grade unmanned aerial vehicles (UAVs) may offer an intermediate solution [
53] at an affordable cost [
54]. Compared with very-high-resolution spaceborne data, their application is associated with fewer limitations regarding atmospheric correction and cloud cover because of their relatively low flight altitudes [
55]. Additionally, 3D point clouds can be created from raw UAV data by using stereo image-matching techniques [
56]. This enhances the level of structural analysis from a planar space to a vertical space [
57] and allows tree heights to be obtain with reasonable accuracy, particularly in open forest environments. UAV photogrammetry is not limited to forests when it comes to tree attribute estimation. Several studies have successfully employed UAV photogrammetry to estimate the height and diameter of orchard trees as well [
58,
59,
60].
The overall workflow of UAV photogrammetry in structural tree attribute estimation involves mission planning, acquisition of aerial images using a UAV, preprocessing of the images to correct distortions, image matching to determine position and orientation, generation of a point cloud and digital surface model, subtraction of the digital surface model to create a canopy height model, and estimation of tree heights using the canopy height model [
53,
61]. The accuracy of the estimated tree heights depends on the quality of the images, the image-matching accuracy, and the effectiveness of segmentation and measurement algorithms.
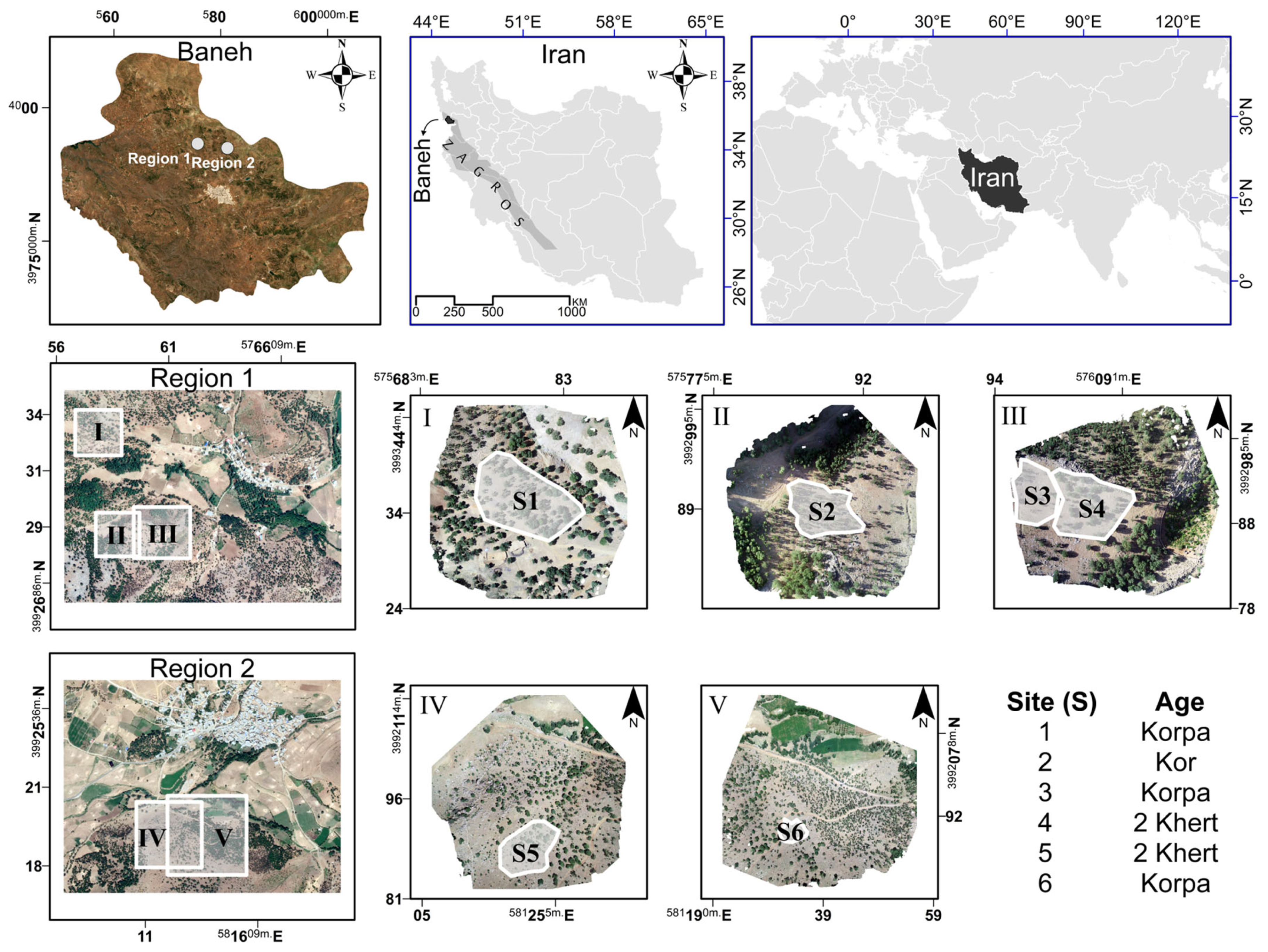
Despite all of these benefits, it is important to also mention the limitations of UAV data. The very small coverage of most UAVs is the primary obstacle, which is a consequence of several factors, including (1) limited battery power/flight time, (2) limited flight area that can be used for stereo analysis, and (3) the significant data volumes needed for creating digital orthomosaics and digital terrain/surface models [
62,
63]. Despite these limitations, a few earlier studies applied UAVs in the Zagros region to assess various forest attributes (
Table 1).
Table 1.
The results of a systematic search on Web of Science® on relevant UAV-based studies within Zagros. AB in the “Search Query” column stands for abstract.
Table 1.
The results of a systematic search on Web of Science® on relevant UAV-based studies within Zagros. AB in the “Search Query” column stands for abstract.
| Search Query | Result |
|---|
| Reference | Year | Description | Region |
|---|
| AB = (UAV OR UAS) AND AB = (Zagros) | [61] | 2022 | Edge detection-based method for delineating overlapped crowns of coppice trees using UAV photogrammetry to monitor tree decline | Middle and South Zagros |
| [53] | 2021 | Structural variable extraction of Zagros single trees | Middle and South Zagros |
| [64] | 2021 | Intra- and interspecific interaction investigation and their contribution to growth inhibition in the patches of Pistacia trees and Amygdalus shrubs | South Zagros |
| [65] | 2017 | Mapping part of a wild pistachio nature reserve in Zagros open woodlands | South Zagros |
| [66] | 2017 | Challenges and quality of landslide assessment based on
remotely sensed data | Zagros Mountains |
| AB = (pollarded) AND AB = (Zagros) | [25] | 2018 | Quantitative description of the pollarding process in the northern Zagros, with a focus on foliage stacks resulting from pollarding | North Zagros |
| [67] | 2018 | Structural analysis of pollarded trees to evaluate the effect of aspect on biometric indices of oak trees | North Zagros |
| [16] | 2017 | Investigation of the effect of pollarding on the increment in diameter of Lebanon oak trees by comparing them to less disturbed stands | North Zagros |
| [6] | 2014 | Investigating the tree-pollarding process in Galajars | North Zagros |
| [18] | 2012 | Allometric relationship introduction for estimating the crown and leaf biomass of Q. libani using the DBH, tree height, crown length, and crown width | North Zagros |
| [27] | 2010 | Evaluation of the capability of IRS-P6 data to separate the pollarding areas | North Zagros |
However, the utility of UAVs as practical and affordable tools for structural assessments of pollarded trees has not yet been examined and was set as the main objective of this research. We were particularly interested in exploring the relationship between UAV-derived tree heights and DBHs of the trees. This is particularly interesting for potential future use cases to conduct simplified forest inventories exclusively based on UAV flights to estimate wood volume and biomass.
Therefore, the specific objectives of this research are:
To test the feasibility of consumer-grade UAV data to estimate the height of pollarded trees and to use these heights to estimate the DBH of the trees;
To understand how the essential DBH~H relationship is affected by the oak species and the stage of pollarding and whether there is a difference between multi- and mono-stemmed trees.
4. Discussion
The general objective of our study was to examine whether height derived from UAV photogrammetry can predict the DBH of pollarded oak trees, which is challenging due to the strongly altered canopy structures [
27,
67]. In the following, we first discuss the technical aspects of UAV-based tree height determination and then reflect on the identified DBH~H relationships.
Earlier studies discussed the challenges of producing realistic 3D models in forests using UAV photogrammetry [
101,
102]. One challenge may be that tree crowns are observed only with a limited number of pixels in UAV images. This complicates solving the collinearity equations of automatic image-matching techniques [
102]. Increasing the number of sampled pixels has been suggested in such conditions [
102]. Tu et al., (2021) showed that the combined use of oblique and nadir flights (as compared to a nadir flight alone) can improve 3D photogrammetry models by 35% [
103]. However, this approach might also be associated with image-matching issues such as changes in wind speed/direction and the sun exposure that may occur due to the time lag between flights, hence complicating the image-matching procedure. Here, we coped with these two issues by taking images using continuous imaging under constant and calm wind conditions. One may also note that pollarded trees are less sensitive to wind because of the generally sparser canopy foliage compared with unpollarded trees. Additionally, we conducted the flights with the shortest possible time lag of approximately one minute between the two flights, due to which changes in sun exposure could be regarded as negligible. In order to overcome many common image-matching problems in forests, we processed images of two oblique (60°) and vertical flights simultaneously to increase the number of candidate pixels in the image-matching process [
56].
All in all, our analysis returned accurate estimates of the tree height using UAV photogrammetric models, with an average difference of 0.3 m to 1.2 m compared to the values extracted from the CHM and the field measurement. This was in the range reported by other studies, which obtained differences of between 0.3 m to 2.9 m [
104] and 0.1 m to 0.5 m [
57] when estimating tree height from UAV data. The average heights of pollarded trees in our study were in the range of 3.5–6.7 m, which can be considered at the lower end of the height values reported in the literature for these silvopastoral systems [
6,
18,
32]. The tree height of pollarded trees is strongly influenced by the local inhabitants who practice pollarding. The traditional owners of Galajars do not allow the crown of the tree to be out of their reach due to the increase in height [
30]. Furthermore, another reason for the low height of these trees may be the low canopy cover of the stands. As a result, there is less competition between trees to receive light (a factor influencing height growth) in pollarded stands, and thus the trees tend to expand their crown horizontally rather than vertically (height growth) [
105].
In terms of height–DBH relationships, previous studies on other forest types generally showed that DBH is correlated with height [
106,
107]. Ref. [
29] showed that the DBHs and heights of pollarded trees of the species
Quercus cerris L. were linearly correlated (correlation coefficient = 0.464). Furthermore, Niemczyk and Bruchwald (2017) presented relationships for estimating the DBH of
Populus spp. trees and hybrids in coppice stands using height and the form factor [
108]. Finally, Ref. [
109] investigated and compared the relationship between the DBH and the height of
Populus spp. in short rotations in northern Poland in two cycles of 5 and 6 years.
In field inventories, height measurement is generally more difficult than DBH measurement [
110], often with lower accuracy of measured height than that of DBH [
38,
111]. Thus, allometries are often used to estimate height using the DBH in classical inventories of standard forests (see
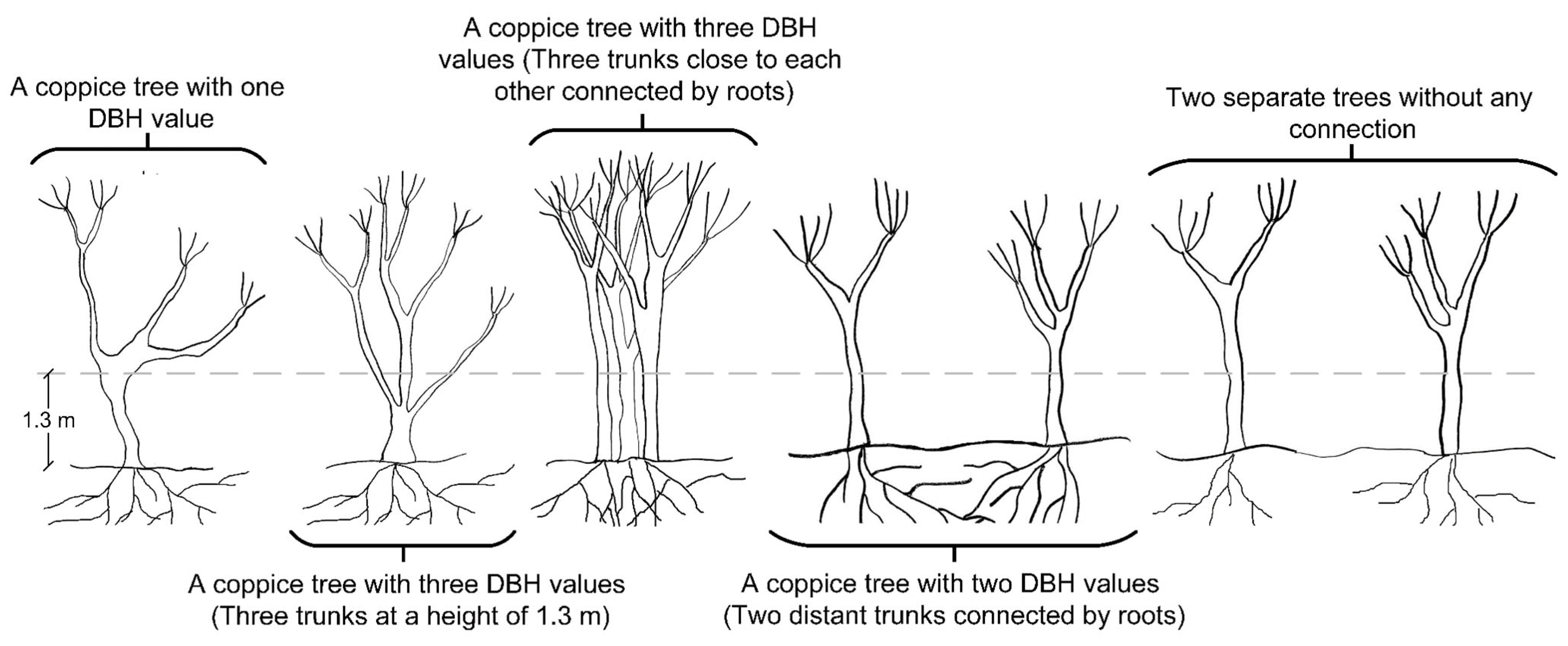
Table 6). We argue that the contrary is true in our case and other comparable cases in stands comprising both single- and multi-stem coppice structures. The use of photogrammetric UAV allows for the estimation of tree heights with reasonable accuracies, whereas DBH measurements may be more time-consuming or prohibited. Therefore, we estimated DBH using the UAV-derived tree heights for single- and multi-stem individuals in this study. In our special case, we used the maximum DBH of pollarded coppice groups (where trees are generally associated with more than one stem) as previously indicated in
Section 2.2. We first checked whether distinguishing between single- and multi-stem trees (categorical variable of Tr in
Table 7) improved the model quality. The high
p-value for the Tr variable throughout the variable selection suggested that this categorization was not helping the model. As a result, we adopted the idea of using the maximum DBH.
The maximum DBH may also be suitable from an ecological perspective. In multi-stemmed coppice oak clumps, dominant stems benefit more from the well-developed parent-tree root system. Therefore, the dominant stem with maximum DBH typically grows faster both in diameter and height. Although multiple stems often persist within a clump for decades [
112], as coppice stems grow larger, competition within the clump increases and reduces the diameter and height growth of stems. Finally, only one or two stems within each clump usually maintain their dominance in the crown position, and other stems are oppressed or excluded due to natural thinning or traditional silvicultural treatments [
113]. Thus, we focused on establishing an allometry between the maximum DBH of the coppice oak trees and our UAV-derived H measurements.
In one of the few other studies focusing on DBH~H relationships in coppice stands, Niemczyk et al., (2016) used the allometric relationship suggested by [
114] to estimate height from DBH. This relationship was nonlinear and included two fitted coefficients that were obtained by the samples measured in short rotations in northern Poland [
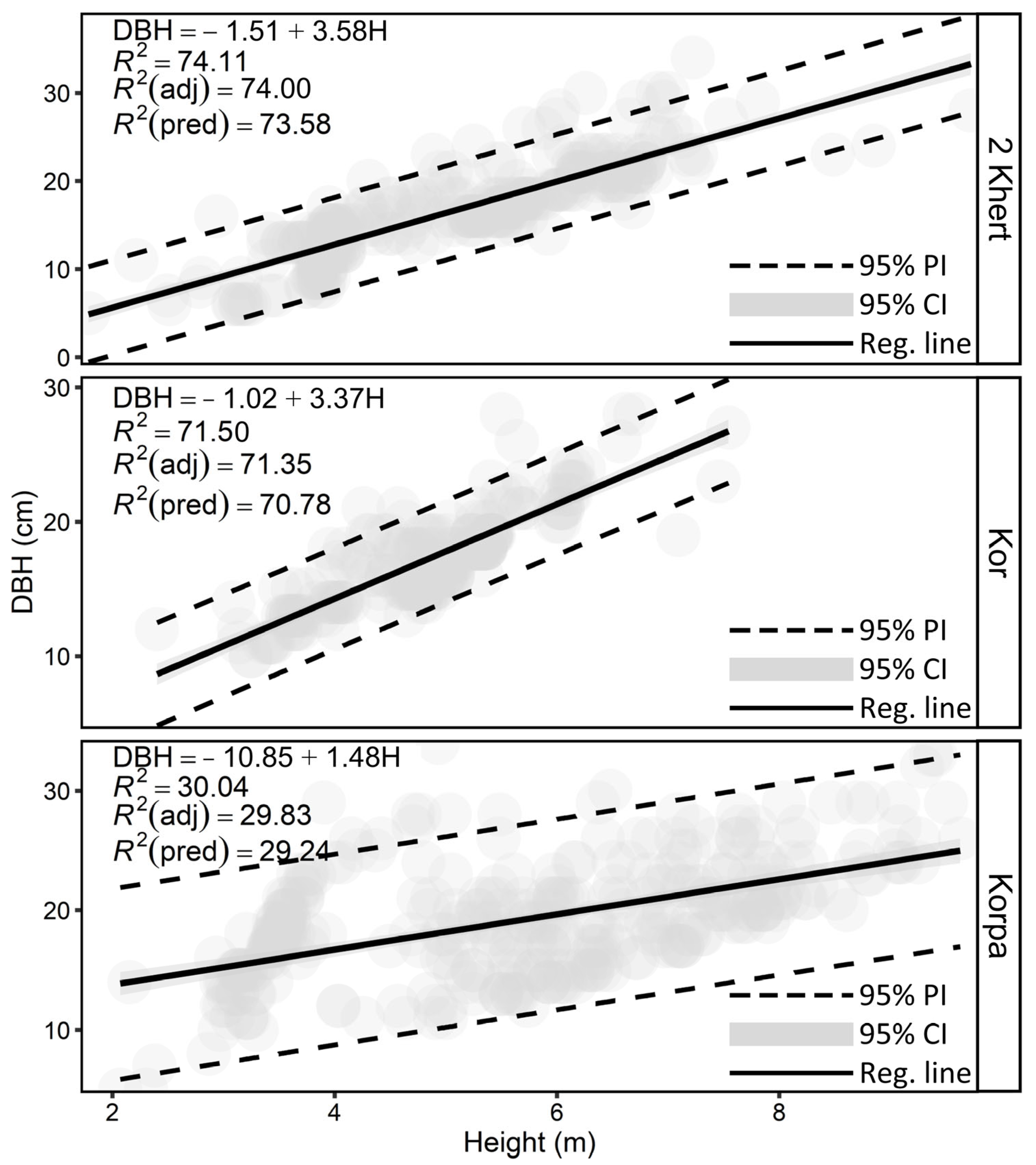
109]. In northern Zagros, such allometric equations do not exist, especially not for different stages of pollarding. Therefore, we applied a linear model to explore the link between DBH and height (
Figure 5), in which we used the SE coefficient, T-value, and VIF to evaluate the model coefficients (
Table 9). The SE coefficient explains how using different sampling techniques might lead to changes in the estimated coefficients [
95]. Here, we observed a higher SE coefficient for the pollarding age (Age) class 2 Khert, i.e., Galajar’s DBH at the 2 Khert age was estimated more accurately. These linear models are not ecologically realistic because the increase in tree height decreases with time, especially when the tree reaches the forest canopy [
115]. But this is only true if the forests are minimally disturbed, whereas our studied forests are regularly changed by humans (see
Figure 1). However, we additionally tested nonlinear allometries (
Table 6). Although the findings demonstrated that nonlinear models exhibited slightly better accuracy than linear models, we chose to utilize the linear model. This decision was based on the simplicity, comprehensibility, and superior interpretability of linear models compared to nonlinear models.
In our study sites, pollarding results in the complete debranching of oak trees. Therefore, one year later, pollarding trees have almost no crown and their height decreases by 1 to 1.5 m. With time, the tree crown recovers by producing new twigs and branches both apically and laterally [
33]. Just 4 years after pollarding (2 Khert class), the tree crown and height reach their typical shape and condition. Thus, the relationship between DBH and height is more reliable in such stands.
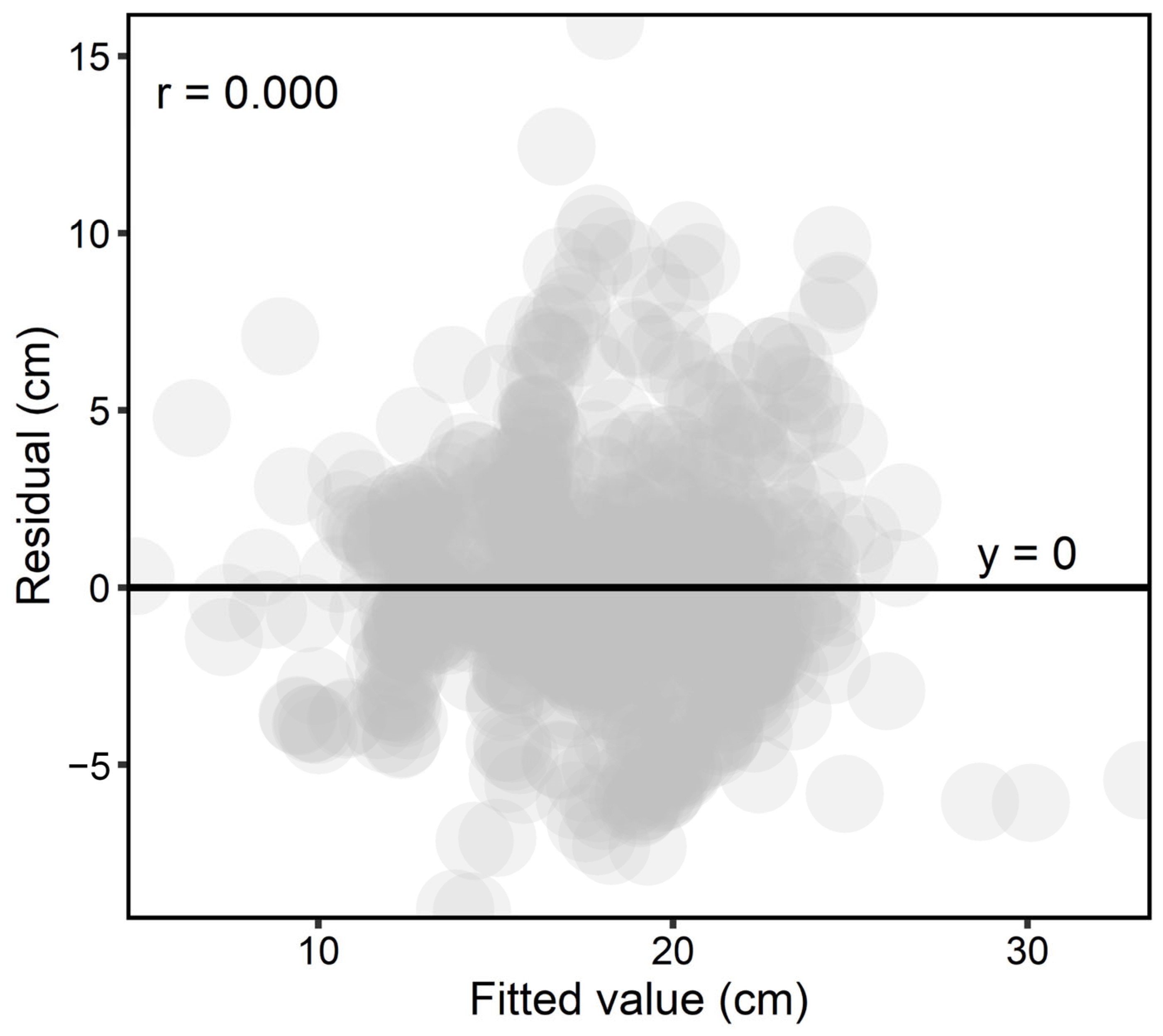
The linear model (
Table 11) showed a reliable R
2 in the overall assessment due to the small difference between R
2 and R
2adj. The R
2pred is essentially a validation that denotes the degree of model reliability [
116]. Furthermore, comparing k-fold S and k-fold R
2 with S and R
2, respectively, can warn of overfitting. We set k to 10, for which the small difference between the values rejected the hypothesis of model overfitting. In addition, the ANOVA (
Table 12) showed relatively high SSError, i.e., changes in DBHMax that cannot be explained by the model. Since we chose DBHMax as the DBH value of the tree (except that the tree was multi-stem), it added an error to the model, which can be seen in SSError. We instead used DBHMax due to the fact that, in oak clumps, the dominant trees are significantly taller than the oppressed trees and stay dominant until all others are thinned or die.
The linear model suggested in this study allows for the prediction of DBH with reasonable accuracies in coppice stands at the age of Kor and 2 Khert, but the accuracy for stands in the Korpa age class is notably worse. Given that the models are also nearly unbiased, there seems to be a certain potential to inventory the DBH and hence also the biomass of oak coppice stands in Northern Zagros. Given the large areas covered by these coppice stands and given the frequently underestimated forest cover of semi-arid forests in global forest maps [
117], this may be a relevant contribution to improve our understanding of global biomass distribution patterns and the corresponding carbon stocks. In future studies, it may be worthwhile to think about developing automated approaches to identify the pollarding age of coppice stands from remote sensing data. This may be possible with very-high-resolution UAV data but could also examined for other VHR data (if available).



 ,
, 


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





