
Early Detection of Myrtle Rust on Pōhutukawa Using Indices Derived from Hyperspectral and Thermal Imagery
 , , , , , , ,
, , , , , , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Setup
2.2. Visual Assessment of Symptoms
2.3. Thermal Measurements
2.4. Hyperspectral Measurements
2.4.1. Data Acquisition
2.4.2. Processing of Data and Extraction of Narrowband Hyperspectral Indices
2.5. Physiological Measurements
2.6. Data Analysis
2.6.1. Treatment Differences in Measured Variables
2.6.2. Classification Model
2.6.3. Relationships between Physiological Variables and Indices
2.7. Software Used
3. Results
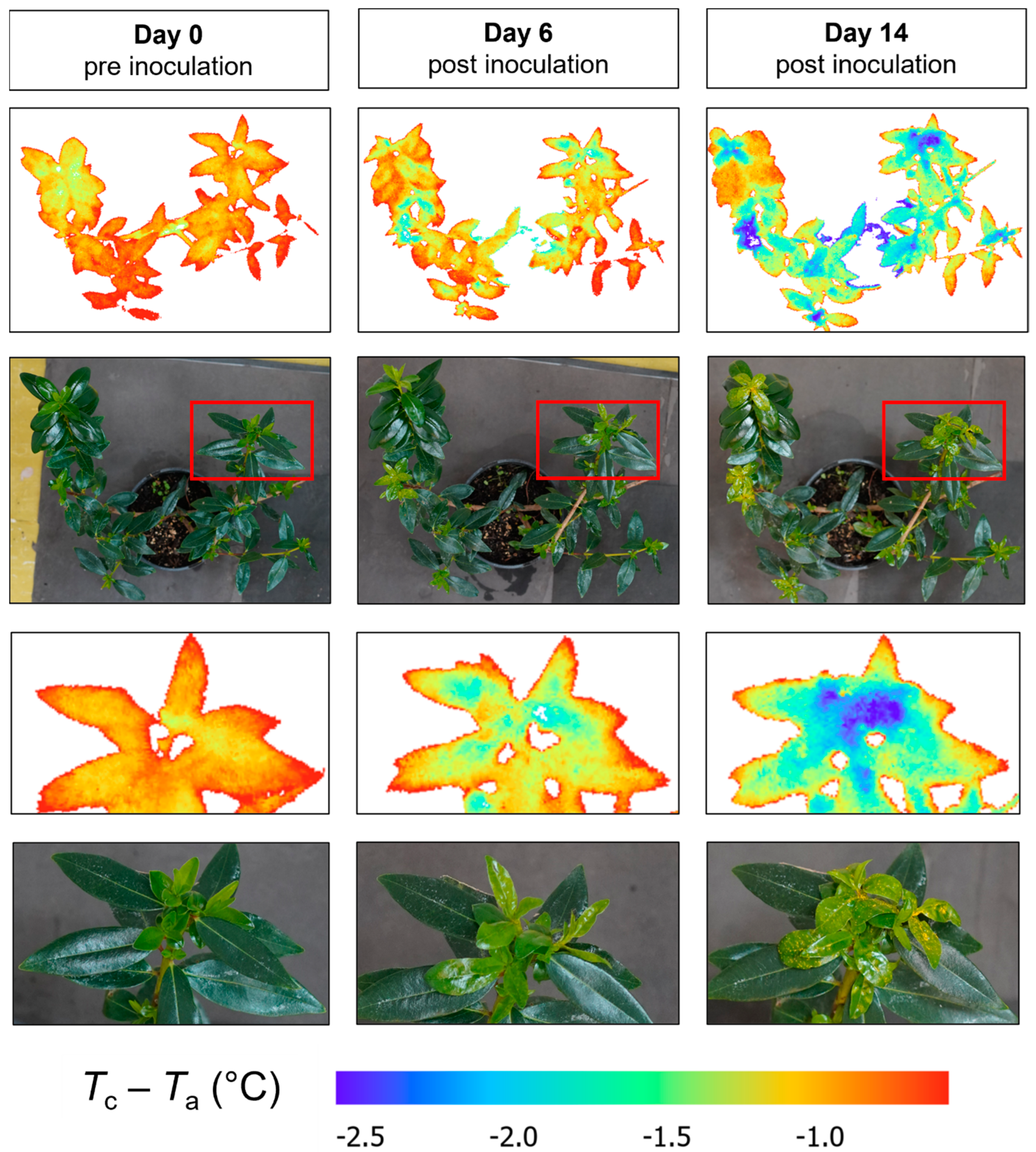
3.1. Disease Symptoms
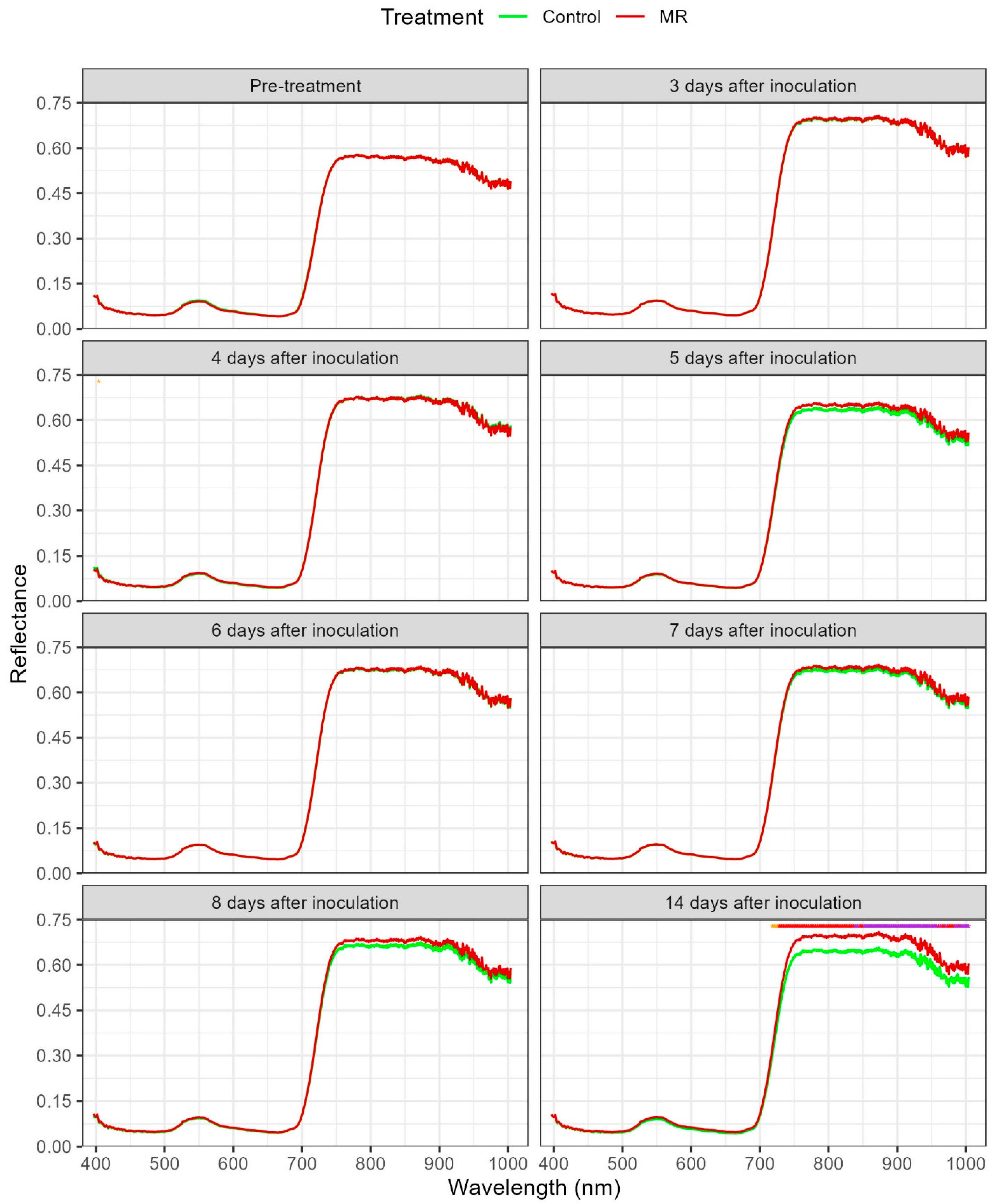
3.2. Hyperspectral Spectra
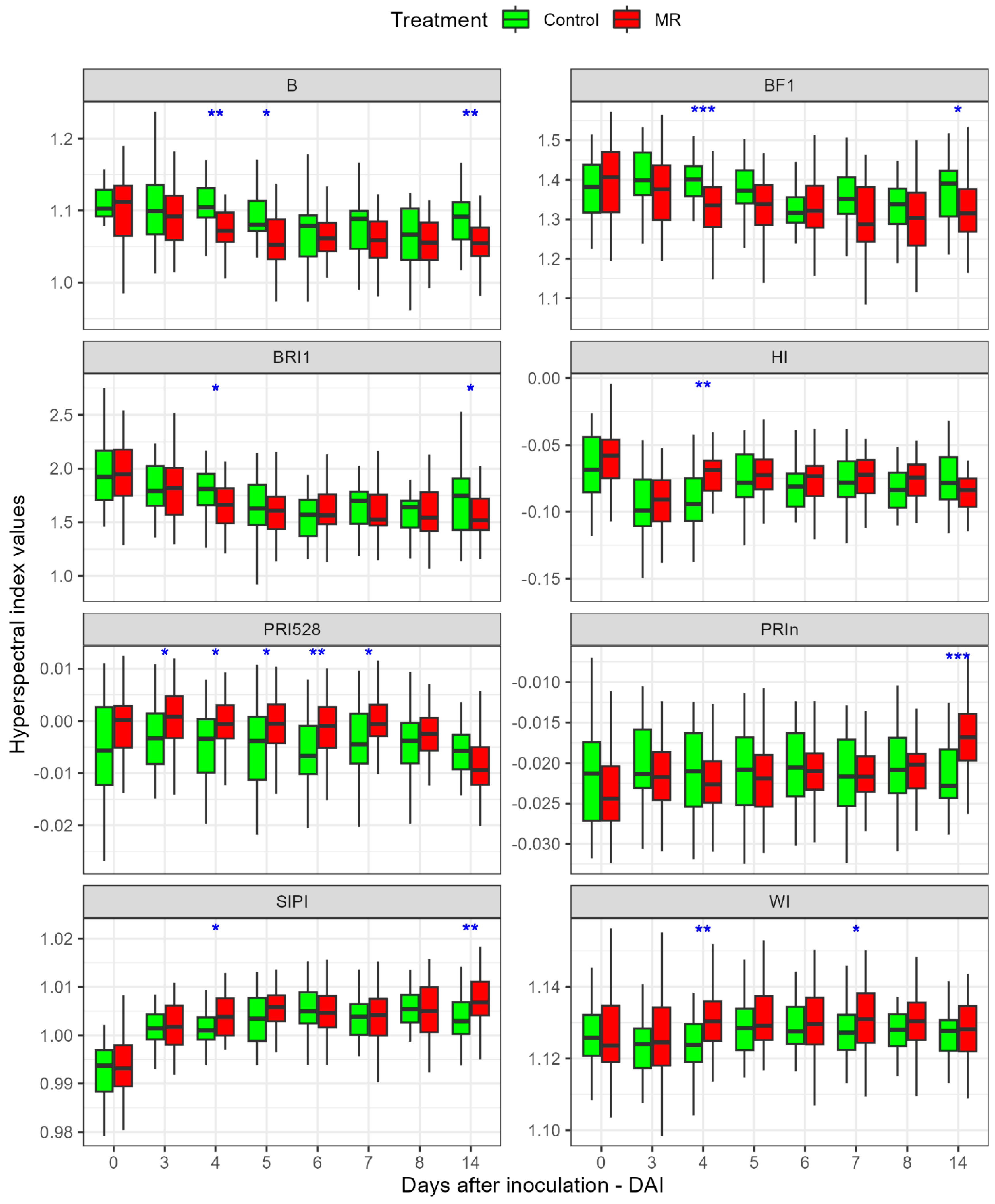
3.3. Variation in Indices between Treatments
3.3.1. Thermal Indices
3.3.2. Hyperspectral Indices
3.4. Model Predictions
3.4.1. Models Using Thermal Indices
3.4.2. Models using Hyperspectral Indices
3.4.3. Models Using Both Thermal and Hyperspectral Indices
3.5. Tree Physiology and Relationships with Thermal Indices and NBHIs
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Indices | Equation | Ref. |
|---|---|---|
| Thermal indices | ||
| Normalised canopy temperature | Tc − Ta | [66] |
| Standard deviation normalised temp. | TSD = std dev (Tc − Ta) | [66] |
| Xanthophyll indices | ||
| Photochemical Refl. Index (570) | [67] | |
| Photochemical Refl. Index (515) | [68] | |
| Photochemical Refl. Index (528) | [67] | |
| Photochemical Refl. Index (550) | [67] | |
| Photochemical Refl. Index m1 | [68] | |
| Photochemical Refl. Index m2 | [67] | |
| Photochemical Refl. Index m3 | [67] | |
| Photochemical Refl. Index m4 | [68] | |
| Normalized Photoch. Refl. Index | [69] | |
| Ratio of PRI to Simple Ratio | [70] | |
| Carotenoid/Chlorophyll Ratio Index | [71] | |
| R/G/B indices | ||
| Redness Index | [72] | |
| Greenness Index | [47] | |
| Greenness Index 2 | [73] | |
| Blue Index | [47] | |
| Blue/green indices | [73] | |
| [73] | ||
| Blue/red indices | [74] | |
| [74] | ||
| BF1 | [25] | |
| BF2 | [25] | |
| BF3 | [25] | |
| BF4 | [25] | |
| BF5 | [25] | |
| Red/green index | [73] | |
| Ratio Analysis of Reflectance Spectra | [75] | |
| Lichtenthaler indices | [76] | |
| [76] | ||
| [76] | ||
| Plant disease indices | ||
| Cercospora leaf spot index | [22] | |
| Healthy-index | [22] | |
| Powdery mildew index | [22] | |
| Sugar beet rust–index | [22] | |
| Water Indices | ||
| Floating position Water Band index | [77] | |
| Water Band Index | [78] | |
| Water Index | [79] | |
| Curvature index | ||
| Curvature index | [80] | |
| Structural indices | ||
| Normalized Difference Veg. Index | [81] | |
| Renormalized Difference Veg. Index | [82] | |
| Optimized Soil-Adjusted Veg. Index | [83] | |
| Modified Soil-Adjusted Vegetation Index | [84] | |
| Triangular Vegetation Index | [85] | |
| Modified Triangular Veg. Index 1 | [86] | |
| Modified Triangular Veg. Index 2 | [86] | |
| Chlorophyll Abs. Reflectance Index | [87] | |
| Modified Chlorophyll Abs. Index | [86] | |
| Modified Chlorophyll Abs. Index 1 | [86] | |
| Modified Chlorophyll Abs. Index 2 | [86] | |
| Modified Chlorophyll Abs. Index 3 | [88] | |
| Simple Ratio | [89] | |
| Modified Simple Ratio | [90] | |
| Enhanced Vegetation Index | [91] | |
| Pigment indices | ||
| Vogelmann indices | [92] | |
| [92] | ||
| [92] | ||
| Gitelson & Merzlyak indices | [53] | |
| [53] | ||
| [93] | ||
| Transformed Chlorophyll Absorption in Reflectance Index | [94] | |
| TCARI/OSAVI | [94] | |
| Chlorophyll Index Red Edge | [94] | |
| Simple Ratio Pigment Index | [51,95] | |
| Normalized Phaeophytinization Index | [51,95] | |
| Normalized Pigments Index | [95] | |
| Carter indices | [96] | |
| [97] | ||
| Reflectance band ratio indices | [98] | |
| [98] | ||
| Structure-Insensitive Pigment Index | [95] | |
| Carotenoid Reflectance Indices | [99,100] | |
| [99] | ||
| [99] | ||
| [99] | ||
| [99,100] | ||
| [99,100] | ||
| Plant Senescing Reflectance Index | [101] | |
| Pigment Specific Simple Ratio Chlorophyll a | [102] | |
| Pigment Spec. Simple Ratio Chl. b | [102] | |
| Pigment Specific Simple Ratio Carotenoid | [102] | |
| Pigment Specific Normalized Difference | [102] | |
| Reciprocal reflectance | [103] |
| Index Type | Variable | Days after Inoculation | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Pre-Treat | 3 | 4 | 5 | 6 | 7 | 8 | 14 | ||
| Thermal indices | Tc − Ta | 0.9339 | 0.9973 | 0.0151 | 0.0202 | 0.0001 | 8.0 × 10−10 | 1.5 × 10−10 | 4.9 × 10−12 |
| TSD | 0.7341 | 0.7462 | 0.7547 | 0.1267 | 0.5007 | 0.0438 | 0.7740 | 0.4121 | |
| Xanthophyll | PRI570 | 0.1696 | 0.3332 | 0.8786 | 0.4486 | 0.6389 | 0.9258 | 0.6662 | 0.0001 |
| indices | PRI515 | 0.6713 | 0.9850 | 0.7761 | 0.9359 | 0.8190 | 0.7972 | 0.8824 | 0.9736 |
| PRI528 | 0.1164 | 0.0105 | 0.0249 | 0.0161 | 0.0045 | 0.0159 | 0.2869 | 0.0632 | |
| PRI550 | 0.7438 | 0.8201 | 0.8678 | 0.7875 | 0.5777 | 0.7670 | 0.4621 | 0.0119 | |
| PRIm1 | 0.5321 | 0.9617 | 0.8583 | 0.8523 | 0.8947 | 0.6965 | 0.6662 | 0.7227 | |
| PRIm2 | 0.8027 | 0.5339 | 0.5010 | 0.9919 | 0.9516 | 0.7548 | 0.7028 | 0.0016 | |
| PRIm3 | 0.4172 | 0.9760 | 0.4142 | 0.7622 | 0.7877 | 0.9519 | 0.8988 | 0.1006 | |
| PRIm4 | 0.2592 | 0.6061 | 0.4235 | 0.5254 | 0.9872 | 0.9792 | 0.8863 | 0.4564 | |
| PRIn | 0.1495 | 0.3196 | 0.2400 | 0.5044 | 0.6155 | 0.9495 | 0.6615 | 4.2 × 10−5 | |
| DRI PRI | 0.1120 | 0.1363 | 0.0659 | 0.2706 | 0.3367 | 0.9111 | 0.3130 | 3.3 × 10−5 | |
| PRI CI | 0.2810 | 0.4728 | 0.8524 | 0.3062 | 0.7850 | 0.9060 | 0.6340 | 0.0252 | |
| R/G/B Indices | R | 0.2691 | 0.6516 | 0.0221 | 0.5573 | 0.7884 | 0.6937 | 0.4057 | 0.3709 |
| G | 0.3597 | 0.9302 | 0.4649 | 0.6874 | 0.8223 | 0.9647 | 0.9066 | 0.4573 | |
| GI | 0.4279 | 0.9213 | 0.8493 | 0.7929 | 0.5656 | 0.7646 | 0.7260 | 0.3613 | |
| B | 0.3526 | 0.4589 | 0.0027 | 0.0156 | 0.5287 | 0.2028 | 0.6033 | 0.0084 | |
| BGI1 | 0.8036 | 0.5767 | 0.0114 | 0.4078 | 0.8960 | 0.4431 | 0.7473 | 0.0561 | |
| BGI2 | 0.6555 | 0.7916 | 0.2865 | 0.4779 | 0.9907 | 0.8035 | 0.8828 | 0.0849 | |
| BRI1 | 0.8712 | 0.7311 | 0.0330 | 0.4418 | 0.5525 | 0.6060 | 0.8989 | 0.0245 | |
| BRI2 | 0.6972 | 0.9057 | 0.9400 | 0.4729 | 0.3743 | 0.8033 | 0.2886 | 0.0078 | |
| BF1 | 0.9467 | 0.1314 | 0.0001 | 0.3741 | 0.4419 | 0.1247 | 0.2108 | 0.0128 | |
| BF2 | 0.9729 | 0.4207 | 0.0010 | 0.3413 | 0.8676 | 0.1249 | 0.6432 | 0.0309 | |
| BF3 | 0.9967 | 0.3856 | 0.0018 | 0.3317 | 0.5423 | 0.2856 | 0.7265 | 0.0588 | |
| BF4 | 0.8910 | 0.4000 | 0.0020 | 0.2861 | 0.6901 | 0.2605 | 0.5967 | 0.0458 | |
| BF5 | 0.9584 | 0.5672 | 0.0019 | 0.5733 | 0.6533 | 0.3933 | 0.6790 | 0.1121 | |
| RGI | 0.7144 | 0.6174 | 0.1288 | 0.7800 | 0.2476 | 0.5355 | 0.3252 | 0.4593 | |
| RARS | 0.6215 | 0.8175 | 0.1998 | 0.7759 | 0.5264 | 0.6026 | 0.5225 | 0.8905 | |
| LIC1 | 0.9935 | 0.8217 | 0.3202 | 0.9349 | 0.6021 | 0.6564 | 0.9279 | 0.8995 | |
| LIC2 | 0.6516 | 0.6119 | 0.8630 | 0.9425 | 0.3314 | 0.5478 | 0.2442 | 0.0339 | |
| LIC3 | 0.8452 | 0.9957 | 0.9865 | 0.8679 | 0.7182 | 0.8787 | 0.5647 | 0.2464 | |
| Plant disease | CLS | 0.9084 | 0.5916 | 0.1020 | 0.2650 | 0.5584 | 0.3057 | 0.1284 | 0.0038 |
| indices | HI | 0.2138 | 0.3411 | 0.0033 | 0.5509 | 0.1879 | 0.3993 | 0.1174 | 0.1151 |
| PMI | 0.9385 | 0.5872 | 0.6885 | 0.1753 | 0.6217 | 0.3406 | 0.2644 | 0.0961 | |
| SBRI | 0.3296 | 0.6373 | 0.6180 | 0.7500 | 0.5532 | 0.6695 | 0.7972 | 0.1207 | |
| Water indices | fWBI | 0.6527 | 0.2858 | 0.0027 | 0.6518 | 0.3417 | 0.9034 | 0.1109 | 0.1139 |
| WBI | 0.9572 | 0.3024 | 0.0041 | 0.2341 | 0.6796 | 0.0348 | 0.1291 | 0.4479 | |
| WI | 0.9567 | 0.2952 | 0.0039 | 0.2300 | 0.6722 | 0.0340 | 0.1273 | 0.4415 | |
| Curvature index | CUR | 0.1417 | 0.8552 | 0.4686 | 0.8227 | 0.0479 | 0.1769 | 0.4135 | 0.2775 |
| Index type | Variable | Days after inoculation | |||||||
| Pre-treat | 3 | 4 | 5 | 6 | 7 | 8 | 14 | ||
| Structural | NDVI | 0.9698 | 0.8664 | 0.1813 | 0.9312 | 0.9171 | 0.9239 | 0.8942 | 0.7090 |
| Indices | RDVI | 0.9671 | 0.7938 | 0.6754 | 0.2942 | 0.8220 | 0.4784 | 0.2238 | 0.0020 |
| OSAVI | 0.9256 | 0.7527 | 0.2883 | 0.4071 | 0.8518 | 0.6110 | 0.4268 | 0.0307 | |
| MSAVI | 0.8501 | 0.7599 | 0.3139 | 0.4204 | 0.9937 | 0.7376 | 0.5517 | 0.0291 | |
| TVI | 0.8454 | 0.8632 | 0.9636 | 0.3120 | 0.9335 | 0.5332 | 0.2683 | 0.0031 | |
| MTVI1 | 0.7978 | 0.8614 | 0.9223 | 0.3258 | 0.8473 | 0.4990 | 0.2281 | 0.0020 | |
| MTVI2 | 0.5411 | 0.8222 | 0.2969 | 0.4758 | 0.9762 | 0.7379 | 0.4683 | 0.0342 | |
| CARI | 0.2392 | 0.6730 | 0.4107 | 0.9809 | 0.7583 | 0.8778 | 0.9167 | 0.4113 | |
| MCARI | 0.2241 | 0.6262 | 0.1932 | 0.8480 | 0.7474 | 0.8398 | 0.7488 | 0.7026 | |
| MCARI1 | 0.7978 | 0.8614 | 0.9223 | 0.3258 | 0.8473 | 0.4990 | 0.2281 | 0.0020 | |
| MCARI2 | 0.5411 | 0.8222 | 0.2969 | 0.4758 | 0.9762 | 0.7379 | 0.4683 | 0.0342 | |
| MCARI3 | 0.4349 | 0.8673 | 0.8378 | 0.5370 | 0.9843 | 0.7256 | 0.4463 | 0.1168 | |
| SR | 0.9745 | 0.9173 | 0.1201 | 0.7887 | 0.8755 | 0.8529 | 0.7028 | 0.5369 | |
| MSR | 0.9737 | 0.9753 | 0.1325 | 0.8257 | 0.9306 | 0.9124 | 0.7517 | 0.5790 | |
| EVI | 0.8383 | 0.7887 | 0.8961 | 0.3333 | 0.9987 | 0.6844 | 0.3790 | 0.0030 | |
| Pigment indices | VOG1 | 0.5178 | 0.6816 | 0.5840 | 0.7091 | 0.4973 | 0.5356 | 0.7814 | 0.9248 |
| VOG2 | 0.5644 | 0.4978 | 0.3041 | 0.5141 | 0.3558 | 0.3667 | 0.7560 | 0.8244 | |
| VOG3 | 0.5705 | 0.5047 | 0.3254 | 0.5310 | 0.3678 | 0.3828 | 0.7456 | 0.8214 | |
| GM1 | 0.5232 | 0.8457 | 0.3251 | 0.8766 | 0.7383 | 0.8431 | 0.6565 | 0.9276 | |
| GM2 | 0.3531 | 0.8788 | 0.9008 | 0.9224 | 0.9619 | 0.9567 | 0.9495 | 0.8588 | |
| GM4 | 0.5139 | 0.8536 | 0.3366 | 0.8726 | 0.7511 | 0.8240 | 0.6782 | 0.9259 | |
| TCARI | 0.2916 | 0.7881 | 0.6687 | 0.9170 | 0.8386 | 0.9381 | 0.9517 | 0.3950 | |
| TCARI/OSAVI | 0.2856 | 0.7514 | 0.7335 | 0.9951 | 0.8127 | 0.8762 | 0.9781 | 0.5465 | |
| CI1 | 0.4346 | 0.8919 | 0.8206 | 0.9602 | 0.7672 | 0.8840 | 0.9508 | 0.9723 | |
| SRPI | 0.5624 | 0.9834 | 0.0301 | 0.5735 | 0.4467 | 0.8662 | 0.6757 | 0.0166 | |
| NPQI | 0.5400 | 0.7747 | 0.0682 | 0.5994 | 0.2253 | 0.1706 | 0.9208 | 0.6934 | |
| NPCI | 0.5349 | 0.9904 | 0.0308 | 0.6467 | 0.3982 | 0.8710 | 0.6789 | 0.0193 | |
| CTR1 | 0.3898 | 0.8225 | 0.6454 | 0.9615 | 0.3569 | 0.7955 | 0.5274 | 0.0895 | |
| CAR | 0.2471 | 0.6085 | 0.7877 | 0.7402 | 0.7532 | 0.7448 | 0.7507 | 0.9543 | |
| DCabCxc | 0.4016 | 0.8245 | 0.9726 | 0.6721 | 0.7013 | 0.7063 | 0.5185 | 0.2347 | |
| DNIRCabCxc | 0.6054 | 0.6545 | 0.3816 | 0.5901 | 0.6013 | 0.6187 | 0.4315 | 0.2525 | |
| SIPI | 0.4444 | 0.8275 | 0.0383 | 0.4006 | 0.3902 | 0.9822 | 0.6538 | 0.0054 | |
| CRI550 | 0.9078 | 0.9725 | 0.5531 | 0.6046 | 0.9243 | 0.7201 | 0.2542 | 0.2296 | |
| CRI700 | 0.9602 | 0.8092 | 0.1635 | 0.4966 | 0.7319 | 0.5870 | 0.1478 | 0.2726 | |
| CRI550 515 | 0.7129 | 0.8853 | 0.3813 | 0.5831 | 0.6905 | 0.5869 | 0.3172 | 0.1489 | |
| CRI700 515 | 0.9113 | 0.6749 | 0.0726 | 0.4598 | 0.5230 | 0.4612 | 0.1597 | 0.2026 | |
| RNIR CRI550 | 0.9417 | 0.8506 | 0.3818 | 0.9991 | 0.9523 | 0.9022 | 0.4103 | 0.4427 | |
| RNIR CRI700 | 0.8965 | 0.8554 | 0.0674 | 0.7858 | 0.7020 | 0.7004 | 0.2222 | 0.4876 | |
| PSRI | 0.3947 | 0.8987 | 0.9464 | 0.9815 | 0.0838 | 0.2245 | 0.0703 | 0.0101 | |
| PSSRa | 0.9999 | 0.9173 | 0.1518 | 0.7881 | 0.9974 | 0.9851 | 0.7758 | 0.5944 | |
| PSSRb | 0.7417 | 0.9129 | 0.1274 | 0.8405 | 0.7811 | 0.7951 | 0.5879 | 0.5271 | |
| PSSRc | 0.7563 | 0.9371 | 0.2229 | 0.9127 | 0.8068 | 0.8923 | 0.5529 | 0.6924 | |
| PSNDc | 0.9207 | 0.8700 | 0.4106 | 0.9046 | 0.9841 | 0.8072 | 0.7608 | 0.4588 | |
| RR | 0.4156 | 0.9581 | 0.9406 | 0.6472 | 0.8838 | 0.7913 | 0.5692 | 0.0741 | |

References
- Wingfield, M.J.; Slippers, B.; Wingfield, B.D.; Barnes, I. The unified framework for biological invasions: A forest fungal pathogen perspective. Biol. Invasions 2017, 19, 3201–3214. [Google Scholar] [CrossRef]
- Beenken, L. Austropuccinia: A new genus name for the myrtle rust Puccinia psidii placed within the redefined family Sphaerophragmiaceae (Pucciniales). Phytotaxa 2017, 297, 53–61. [Google Scholar] [CrossRef]
- Stewart, J.E.; Ross-Davis, A.L.; Graça, R.N.; Alfenas, A.C.; Peever, T.L.; Hanna, J.W.; Uchida, J.Y.; Hauff, R.D.; Kadooka, C.Y.; Kim, M.S.; et al. Genetic diversity of the myrtle rust pathogen (Austropuccinia psidii) in the Americas and Hawaii: Global implications for invasive threat assessments. For. Pathol. 2017, 48, e12378. [Google Scholar] [CrossRef]
- Glen, M.; Alfenas, A.C.; Zauza, E.A.V.; Wingfield, M.J.; Mohammed, C. Puccinia psidii: A threat to the Australian environment and economy—A review. Australas. Plant Pathol. 2007, 36, 1–16. [Google Scholar] [CrossRef]
- Carnegie, A.J.; Pegg, G.S. Lessons from the Incursion of Myrtle Rust in Australia. Annu. Rev. Phytopathol. 2018, 56, 457–478. [Google Scholar] [CrossRef]
- Berthon, K.A.; Fernandez Winzer, L.; Sandhu, K.; Cuddy, W.; Manea, A.; Carnegie, A.J.; Leishman, M.R. Endangered species face an extra threat: Susceptibility to the invasive pathogen Austropuccinia psidii (myrtle rust) in Australia. Australas. Plant Pathol. 2019, 48, 385–393. [Google Scholar] [CrossRef]
- Soewarto, J.; Giblin, F.; Carnegie, A.J. Austropuccinia psidii (Myrtle Rust) Global Host List, Version 4; Australian Network for Plant Conservation: Canberra, ACT, Australia, 2019. [Google Scholar]
- Almeida, R.F.; Machado, P.S.; Damacena, M.B.; Santos, S.A.; Guimarães, L.M.S.; Klopfenstein, N.B.; Alfenas, A.C. A new, highly aggressive race of Austropuccinia psidii infects a widely planted, myrtle rust-resistant, eucalypt genotype in Brazil. For. Pathol. 2021, 51, e12679. [Google Scholar] [CrossRef]
- Fensham, R.J.; Radford-Smith, J. Unprecedented extinction of tree species by fungal disease. Biol. Conserv. 2021, 261, 109276. [Google Scholar] [CrossRef]
- Heim, R.H.J.; Wright, I.J.; Allen, A.P.; Geedicke, I.; Oldeland, J. Developing a spectral disease index for myrtle rust (Austropuccinia psidii). Plant Pathol. 2019, 68, 738–745. [Google Scholar] [CrossRef]
- Soewarto, J.; Carriconde, F.; Hugot, N.; Bocs, S.; Hamelin, C.; Maggia, L.; Klopfenstein, N.B. Impact of Austropuccinia psidii in New Caledonia, a biodiversity hotspot. For. Pathol. 2018, 48, e12402. [Google Scholar] [CrossRef]
- Sutherland, R.; Soewarto, J.; Beresford, R.; Ganley, B. Monitoring Austropuccinia psidii (myrtle rust) on New Zealand Myrtaceae in native forest. N. Z. J. Ecol. 2020, 44, 1–5. [Google Scholar] [CrossRef]
- Soewarto, J.; Somchit, C.; du Plessis, E.; Barnes, I.; Granados, G.M.; Wingfield, M.J.; Shuey, L.; Bartlett, M.; Fraser, S.; Scott, P.; et al. Susceptibility of native New Zealand Myrtaceae to the South African strain of Austropuccinia psidii: A biosecurity threat. Plant Pathol. 2021, 70, 667–675. [Google Scholar] [CrossRef]
- Beresford, R.M.; Shuey, L.S.; Pegg, G.S. Symptom development and latent period of Austropuccinia psidii (myrtle rust) in relation to host species, temperature, and ontogenic resistance. Plant Pathol. 2020, 69, 484–494. [Google Scholar] [CrossRef]
- Baskarathevan, J.; Taylor, R.K.; Ho, W.; McDougal, R.L.; Shivas, R.G.; Alexander, B.J.R. Real-Time PCR Assays for the Detection of Puccinia psidii. Plant Dis. 2016, 100, 617–624. [Google Scholar] [CrossRef]
- Bini, A.P.; Quecine, M.C.; da Silva, T.M.; Silva, L.D.; Labate, C.A. Development of a quantitative real-time PCR assay using SYBR Green for early detection and quantification of Austropuccinia psidii in Eucalyptus grandis. Eur. J. Plant Pathol. 2017, 150, 735–746. [Google Scholar] [CrossRef]
- Carnegie, A.J.; Cooper, K. Emergency response to the incursion of an exotic myrtaceous rust in Australia. Australas. Plant Pathol. 2011, 40, 346–359. [Google Scholar] [CrossRef]
- Langrell, S.R.H.; Glen, M.; Alfenas, A.C. Molecular diagnosis of Puccinia psidii (guava rust)—A quarantine threat to Australian eucalypt and Myrtaceae biodiversity. Plant Pathol. 2008, 57, 687–701. [Google Scholar] [CrossRef]
- Roux, J.; Greyling, I.; Coutinho, T.A.; Verleur, M.; Wingfield, M.J. The Myrtle rust pathogen, Puccinia psidii, discovered in Africa. IMA Fungus 2013, 4, 155–159. [Google Scholar] [CrossRef]
- Hernández-Clemente, R.; Hornero, A.; Mottus, M.; Penuelas, J.; González-Dugo, V.; Jiménez, J.C.; Suárez, L.; Alonso, L.; Zarco-Tejada, P.J. Early Diagnosis of Vegetation Health From High-Resolution Hyperspectral and Thermal Imagery: Lessons Learned From Empirical Relationships and Radiative Transfer Modelling. Curr. For. Rep. 2019, 5, 169–183. [Google Scholar] [CrossRef]
- Hornero, A.; Zarco-Tejada, P.J.; Quero, J.L.; North, P.R.J.; Ruiz-Gómez, F.J.; Sánchez-Cuesta, R.; Hernandez-Clemente, R. Modelling hyperspectral-and thermal-based plant traits for the early detection of Phytophthora-induced symptoms in oak decline. Remote Sens. Environ. 2021, 263, 112570. [Google Scholar] [CrossRef]
- Mahlein, A.K.; Rumpf, T.; Welke, P.; Dehne, H.W.; Plümer, L.; Steiner, U.; Oerke, E.C. Development of spectral indices for detecting and identifying plant diseases. Remote Sens. Environ. 2013, 128, 21–30. [Google Scholar] [CrossRef]
- Poblete, T.; Camino, C.; Beck, P.S.A.; Hornero, A.; Kattenborn, T.; Saponari, M.; Boscia, D.; Navas-Cortes, J.A.; Zarco-Tejada, P.J. Detection of Xylella fastidiosa infection symptoms with airborne multispectral and thermal imagery: Assessing bandset reduction performance from hyperspectral analysis. ISPRS J. Photogramm. Remote Sens. 2020, 162, 27–40. [Google Scholar] [CrossRef]
- Tian, L.; Xue, B.; Wang, Z.; Li, D.; Yao, X.; Cao, Q.; Zhu, Y.; Cao, W.; Cheng, T. Spectroscopic detection of rice leaf blast infection from asymptomatic to mild stages with integrated machine learning and feature selection. Remote Sens. Environ. 2021, 257, 112350. [Google Scholar] [CrossRef]
- Zarco-Tejada, P.J.; Camino, C.; Beck, P.S.A.; Calderon, R.; Hornero, A.; Hernández-Clemente, R.; Kattenborn, T.; Montes-Borrego, M.; Susca, L.; Morelli, M. Previsual symptoms of Xylella fastidiosa infection revealed in spectral plant-trait alterations. Nat. Plants 2018, 4, 432–439. [Google Scholar] [CrossRef]
- Calderón, R.; Navas-Cortés, J.A.; Zarco-Tejada, P.J. Early detection and quantification of Verticillium wilt in olive using hyperspectral and thermal imagery over large areas. Remote Sens. 2015, 7, 5584–5610. [Google Scholar] [CrossRef]
- Watt, M.S.; Bartlett, M.; Soewarto, J.; de Silva, D.; Estarija, H.J.C.; Massam, P.; Cajes, D.; Yorston, W.; Graevskaya, E.; Dobbie, K. Previsual and early detection of myrtle rust on rose apple using indices derived from thermal imagery and visible-to-short-infrared spectroscopy. Phytopathology 2023, 113, 1405–1416. [Google Scholar] [CrossRef]
- Bylsma, R.J.; Clarkson, B.D.; Efford, J.T. Biological flora of New Zealand 14: Metrosideros excelsa, pōhutukawa, New Zealand Christmas tree. N. Z. J. Bot. 2014, 52, 365–385. [Google Scholar] [CrossRef]
- Black, A.; Garner, G.; Mark-Shadbolt, M.; Balanovic, J.; MacDonald, E.; Mercier, O.; Wright, J.; Calver, M. Indigenous peoples’ attitudes and social acceptability of invasive species control in New Zealand. Pac. Conserv. Biol. 2021, 28, 481–490. [Google Scholar] [CrossRef]
- Teulon, D.A.J.; Alipia, T.T.; Ropata, H.T.; Green, J.M.; Viljanen, S.L.H.; Cromey, M.G.; Arthur, K.; MacDairmid, R.M.; Waipara, N.W.; Marsh, A.T. The threat of myrtle rust to Māori taonga plant species in New Zealand. N. Z. Plant Prot. 2015, 68, 66–75. [Google Scholar]
- Dawson, M.; Hobbs, J.; Platt, G.; Rumbal, J. Metrosideros in cultivation: Pōhutukawa. N. Z. Gard. J. 2010, 13, 10–22. [Google Scholar]
- Beresford, R.M.; Turner, R.; Tait, A.; Paul, V.; Macara, G.; Yu, Z.D.; Lima, L.; Martin, R. Predicting the climatic risk of myrtle rust during its first year in New Zealand. N. Z. Plant Prot. 2018, 71, 332–347. [Google Scholar] [CrossRef]
- Toome-Heller, M.; Ho, W.W.H.; Ganley, R.J.; Elliott, C.E.A.; Quinn, B.; Pearson, H.G.; Alexander, B.J.R. Chasing myrtle rust in New Zealand: Host range and distribution over the first year after invasion. Australas. Plant Pathol. 2020, 49, 221–230. [Google Scholar] [CrossRef]
- Ho, W.H.; Baskarathevan, J.; Griffin, R.L.; Quinn, B.D.; Alexander, B.J.R.; Havell, D.; Ward, N.A.; Pathan, A.K. First report of myrtle rust caused by Austropuccinia psidii on Metrosideros kermadecensis on Raoul Island and on M. excelsa in Kerikeri, New Zealand. Plant Dis. 2019, 103, 2128. [Google Scholar] [CrossRef]
- Ministry for Primary Industries. Species Infected with Myrtle Rust in New Zealand. 2024. Available online: https://myrtlerust-uat.biosites.mpi.govt.nz/about-myrtle-rust/species-infected-with-myrtle-rust-in-new-zealand/ (accessed on 12 February 2024).
- Rasband, W.S. ImageJ; U.S. National Institutes of Health: Bethesda, MD, USA, 2012. Available online: https://imagej.nih.gov/ij/ (accessed on 11 December 2023).
- Buddenbaum, H.; Watt, M.S.; Scholten, R.C.; Hill, J. Preprocessing ground-based visible/near infrared imaging spectroscopy data affected by smile effects. Sensors 2019, 19, 1543. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2023; Available online: https://www.R-project.org/ (accessed on 2 February 2024).
- Pedregosa, F.; Varoquaux, G.; Gramfort, A.; Michel, V.; Thirion, B.; Grisel, O.; Blondel, M.; Prettenhofer, P.; Weiss, R.; Dubourg, V. Scikit-learn: Machine Learning in Python. J. Mach. Learn. Res. 2011, 12, 2825–2830. [Google Scholar]
- Yuan, X.; Chen, S.; Sun, C.; Yuwen, L. A novel early diagnostic framework for chronic diseases with class imbalance. Sci. Rep. 2022, 12, 8614. [Google Scholar] [CrossRef] [PubMed]
- Still, C.; Powell, R.; Aubrecht, D.; Kim, Y.; Helliker, B.; Roberts, D.; Richardson, A.D.; Goulden, M. Thermal imaging in plant and ecosystem ecology: Applications and challenges. Ecosphere 2019, 10, e02768. [Google Scholar] [CrossRef]
- Chock, M.K. The global threat of Myrtle rust (Austropuccinia psidii): Future prospects for control and breeding resistance in susceptible hosts. Crop Protect. 2020, 136, 105176. [Google Scholar] [CrossRef]
- Smith, R.C.G.; Heritage, A.D.; Stapper, M.; Barrs, H.D. Effect of stripe rust (Puccinia striiformis West.) and irrigation on the yield and foliage temperature of wheat. Field Crops Res. 1986, 14, 39–51. [Google Scholar] [CrossRef]
- Nichol, C.J.; Rascher, U.; Matsubara, S.; Osmond, B. Assessing photosynthetic efficiency in an experimental mangrove canopy using remote sensing and chlorophyll fluorescence. Trees 2006, 20, 9–15. [Google Scholar] [CrossRef]
- Peguero-Pina, J.J.; Morales, F.; Flexas, J.; Gil-Pelegrín, E.; Moya, I. Photochemistry, remotely sensed physiological reflectance index and de-epoxidation state of the xanthophyll cycle in Quercus coccifera under intense drought. Oecologia 2008, 156, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Zarco-Tejada, P.J.; Berni, J.A.J.; Suárez, L.; Sepulcre-Cantó, G.; Morales, F.; Miller, J.R. Imaging chlorophyll fluorescence with an airborne narrow-band multispectral camera for vegetation stress detection. Remote Sens. Environ. 2009, 113, 1262–1275. [Google Scholar] [CrossRef]
- Calderón, R.; Navas-Cortés, J.A.; Lucena, C.; Zarco-Tejada, P.J. High-resolution airborne hyperspectral and thermal imagery for early detection of Verticillium wilt of olive using fluorescence, temperature and narrow-band spectral indices. Remote Sens. Environ. 2013, 139, 231–245. [Google Scholar] [CrossRef]
- López-López, M.; Calderón, R.; González-Dugo, V.; Zarco-Tejada, P.J.; Fereres, E. Early detection and quantification of almond red leaf blotch using high-resolution hyperspectral and thermal imagery. Remote Sens. 2016, 8, 276. [Google Scholar] [CrossRef]
- Watt, M.S.; Poblete, T.; de Silva, D.; Estarija, H.J.C.; Hartley, R.J.L.; Leonardo, E.M.C.; Massam, P.; Buddenbaum, H.; Zarco-Tejada, P.J. Prediction of the severity of Dothistroma needle blight in radiata pine using plant based traits and narrow band indices derived from UAV hyperspectral imagery. Agric. For. Meteorol. 2023, 330, 109294. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Rinderle, U. The role of chlorophyll fluorescence in the detection of stress conditions in plants. CRC Crit. Rev. Anal. Chem. 1988, 19, S29–S85. [Google Scholar] [CrossRef]
- Barnes, J.D.; Balaguer, L.; Manrique, E.; Elvira, S.; Davison, A.W. A reappraisal of the use of DMSO for the extraction and determination of chlorophylls a and b in lichens and higher plants. Environ. Exp. Bot. 1992, 32, 85–100. [Google Scholar] [CrossRef]
- Penuelas, J.; Filella, I.; Lloret, P.; Mun Oz, F.; Vilajeliu, M. Reflectance assessment of mite effects on apple trees. Int. J. Remote Sens. 1995, 16, 2727–2733. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Merzlyak, M.N. Signature analysis of leaf reflectance spectra: Algorithm development for remote sensing of chlorophyll. J. Plant Physiol. 1996, 148, 494–500. [Google Scholar] [CrossRef]
- Oliva, J.; Stenlid, J.; Martinez-Vilalta, J. The effect of fungal pathogens on the water and carbon economy of trees: Implications for drought-induced mortality. New Phytol. 2014, 203, 1028–1035. [Google Scholar] [CrossRef]
- Skoneczny, H.; Kubiak, K.; Spiralski, M.; Kotlarz, J.; Mikiciński, A.; Puławska, J. Fire blight disease detection for apple trees: Hyperspectral analysis of healthy, infected and dry leaves. Remote Sens. 2020, 12, 2101. [Google Scholar] [CrossRef]
- Kim, S.-R.; Lee, W.-K.; Lim, C.-H.; Kim, M.; Kafatos, M.C.; Lee, S.-H.; Lee, S.-S. Hyperspectral analysis of pine wilt disease to determine an optimal detection index. Forests 2018, 9, 115. [Google Scholar] [CrossRef]
- Buddenbaum, H.; Rock, G.; Hill, J.; Werner, W. Measuring stress reactions of beech seedlings with PRI, fluorescence, temperatures and emissivity from VNIR and thermal field imaging spectroscopy. Eur. J. Remote Sens. 2015, 48, 263–282. [Google Scholar] [CrossRef]
- Pearse, G.D.; Watt, M.S.; Soewarto, J.; Tan, A.Y.S. Deep learning and phenology enhance large-scale tree species classification in aerial imagery during a biosecurity response. Remote Sens. 2021, 13, 1789. [Google Scholar] [CrossRef]
- Duarte, A.; Borralho, N.; Cabral, P.; Caetano, M. Recent advances in forest insect pests and diseases monitoring using UAV-based data: A systematic review. Forests 2022, 13, 911. [Google Scholar] [CrossRef]
- Watt, M.S.; Pearse, G.D.; Dash, J.P.; Melia, N.; Leonardo, E.M.C. Application of remote sensing technologies to identify impacts of nutritional deficiencies on forests. ISPRS J. Photogramm. Remote Sens. 2019, 149, 226–241. [Google Scholar] [CrossRef]
- Neinavaz, E.; Schlerf, M.; Darvishzadeh, R.; Gerhards, M.; Skidmore, A.K. Thermal infrared remote sensing of vegetation: Current status and perspectives. Int. J. Appl. Earth Obs. Geoinf. 2021, 102, 102415. [Google Scholar] [CrossRef]
- Smigaj, M.; Agarwal, A.; Bartholomeus, H.; Decuyper, M.; Elsherif, A.; de Jonge, A.; Kooistra, L. Thermal Infrared Remote Sensing of Stress Responses in Forest Environments: A Review of Developments, Challenges, and Opportunities. Curr. For. Rep. 2024, 10, 56–76. [Google Scholar] [CrossRef]
- Asner, G.P. Biophysical and biochemical sources of variability in canopy reflectance. Remote Sens. Environ. 1998, 64, 234–253. [Google Scholar] [CrossRef]
- Ollinger, S.V. Sources of variability in canopy reflectance and the convergent properties of plants. New Phytol. 2011, 189, 375–394. [Google Scholar] [CrossRef]
- Cao, B.; Liu, Q.; Du, Y.; Roujean, J.-L.; Gastellu-Etchegorry, J.-P.; Trigo, I.F.; Zhan, W.; Yu, Y.; Cheng, J.; Jacob, F. A review of earth surface thermal radiation directionality observing and modeling: Historical development, current status and perspectives. Remote Sens. Environ. 2019, 232, 111304. [Google Scholar] [CrossRef]
- Idso, S.B.; Jackson, R.D.; Pinter Jr, P.J.; Reginato, R.J.; Hatfield, J.L. Normalizing the stress-degree-day parameter for environmental variability. Agric. Meteorol. 1981, 24, 45–55. [Google Scholar] [CrossRef]
- Gamon, J.A.; Penuelas, J.; Field, C.B. A narrow-waveband spectral index that tracks diurnal changes in photosynthetic efficiency. Remote Sens. Environ. 1992, 41, 35–44. [Google Scholar] [CrossRef]
- Hernández-Clemente, R.; Navarro-Cerrillo, R.M.; Suárez, L.; Morales, F.; Zarco-Tejada, P.J. Assessing structural effects on PRI for stress detection in conifer forests. Remote Sens. Environ. 2011, 115, 2360–2375. [Google Scholar] [CrossRef]
- Zarco-Tejada, P.J.; Morales, A.; Testi, L.; Villalobos, F.J. Spatio-temporal patterns of chlorophyll fluorescence and physiological and structural indices acquired from hyperspectral imagery as compared with carbon fluxes measured with eddy covariance. Remote Sens. Environ. 2013, 133, 102–115. [Google Scholar] [CrossRef]
- Dotzler, S.; Hill, J.; Buddenbaum, H.; Stoffels, J. The potential of EnMAP and Sentinel-2 data for detecting drought stress phenomena in deciduous forest communities. Remote Sens. 2015, 7, 14227–14258. [Google Scholar] [CrossRef]
- Garrity, S.R.; Eitel, J.U.H.; Vierling, L.A. Disentangling the relationships between plant pigments and the photochemical reflectance index reveals a new approach for remote estimation of carotenoid content. Remote Sens. Environ. 2011, 115, 628–635. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Yacobi, Y.Z.; Schalles, J.F.; Rundquist, D.C.; Han, L.; Stark, R.; Etzion, D. Remote estimation of phytoplankton density in productive waters. Adv. Limnol. Stuttg. 2000, 55, 121–136. [Google Scholar]
- Zarco-Tejada, P.J.; Berjón, A.; Lopez-Lozano, R.; Miller, J.R.; Martín, P.; Cachorro, V.; González, M.R.; De Frutos, A. Assessing vineyard condition with hyperspectral indices: Leaf and canopy reflectance simulation in a row-structured discontinuous canopy. Remote Sens. Environ. 2005, 99, 271–287. [Google Scholar] [CrossRef]
- Zarco-Tejada, P.J.; González-Dugo, V.; Berni, J.A.J. Fluorescence, temperature and narrow-band indices acquired from a UAV platform for water stress detection using a micro-hyperspectral imager and a thermal camera. Remote Sens. Environ. 2012, 117, 322–337. [Google Scholar] [CrossRef]
- Chappelle, E.W.; Kim, M.S.; Mcmurtrey, J.E. Ratio analysis of reflectance spectra (RARS): An algorithm for the remote estimation of the concentrations of Chlorophyll A, Chlorophyll B, and Carotenoids in Soybean leaves. Remote Sens. Environ. 1992, 39, 239–247. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Vegetation stress: An introduction to the stress concept in plants. J. Plant Physiol. 1996, 148, 4–14. [Google Scholar] [CrossRef]
- Strachan, I.B.; Pattey, E.; Boisvert, J.B. Impact of nitrogen and environmental conditions on corn as detected by hyperspectral reflectance. Remote Sens. Environ. 2002, 80, 213–224. [Google Scholar] [CrossRef]
- Peñuelas, J.; Filella, I.; Biel, C.; Serrano, L.; Save, R. The reflectance at the 950–970 nm region as an indicator of plant water status. Int. J. Remote Sens. 1993, 14, 1887–1905. [Google Scholar] [CrossRef]
- Peñuelas, J.; Pinol, J.; Ogaya, R.; Filella, I. Estimation of plant water concentration by the reflectance water index WI (R900/R970). Int. J. Remote Sens. 1997, 18, 2869–2875. [Google Scholar] [CrossRef]
- Zarco-Tejada, P.J.; Miller, J.R.; Mohammed, G.H.; Noland, T.L. Chlorophyll fluorescence effects on vegetation apparent reflectance: I. Leaf-level measurements and model simulation. Remote Sens. Environ. 2000, 74, 582–595. [Google Scholar] [CrossRef]
- Rouse, J.W.; Haas, R.H.; Schell, J.A.; Deering, D.W. Monitoring vegetation systems in the Great Plains with ERTS. NASA Spec. Publ. 1974, 351, 309. [Google Scholar]
- Roujean, J.-L.; Breon, F.-M. Estimating PAR absorbed by vegetation from bidirectional reflectance measurements. Remote Sens. Environ. 1995, 51, 375–384. [Google Scholar] [CrossRef]
- Rondeaux, G.; Steven, M.; Baret, F. Optimization of soil-adjusted vegetation indices. Remote Sens. Environ. 1996, 55, 95–107. [Google Scholar] [CrossRef]
- Qi, J.; Chehbouni, A.; Huete, A.R.; Kerr, Y.H.; Sorooshian, S. A modified soil adjusted vegetation index. Remote Sens. Environ. 1994, 48, 119–126. [Google Scholar] [CrossRef]
- Broge, N.H.; Leblanc, E. Comparing prediction power and stability of broadband and hyperspectral vegetation indices for estimation of green leaf area index and canopy chlorophyll density. Remote Sens. Environ. 2001, 76, 156–172. [Google Scholar] [CrossRef]
- Haboudane, D.; Miller, J.R.; Pattey, E.; Zarco-Tejada, P.J.; Strachan, I.B. Hyperspectral vegetation indices and novel algorithms for predicting green LAI of crop canopies: Modeling and validation in the context of precision agriculture. Remote Sens. Environ. 2004, 90, 337–352. [Google Scholar] [CrossRef]
- Kim, M.S. The Use of Narrow Spectral Bands for Improving Remote Sensing Estimations of Fractionally Absorbed Photosynthetically Active Radiation. Ph.D. Dissertation, University of Maryland, College Park, MD, USA, 1994. [Google Scholar]
- Wu, C.; Niu, Z.; Tang, Q.; Huang, W. Estimating chlorophyll content from hyperspectral vegetation indices: Modeling and validation. Agric. For. Meteorol. 2008, 148, 1230–1241. [Google Scholar] [CrossRef]
- Jordan, C.F. Derivation of leaf-area index from quality of light on the forest floor. Ecology 1969, 50, 663–666. [Google Scholar] [CrossRef]
- Chen, J.M. Evaluation of vegetation indices and a modified simple ratio for boreal applications. Can. J. Remote Sens. 1996, 22, 229–242. [Google Scholar] [CrossRef]
- Liu, H.Q.; Huete, A. A feedback based modification of the NDVI to minimize canopy background and atmospheric noise. IEEE Trans. Geosci. Remote Sens. 1995, 33, 457–465. [Google Scholar] [CrossRef]
- Vogelmann, T.C. Plant tissue optics. Annu. Rev. Plant Biol. 1993, 44, 231–251. [Google Scholar] [CrossRef]
- Gitelson, A.; Merzlyak, M.N. Spectral reflectance changes associated with autumn senescence of Aesculus hippocastanum L. and Acer platanoides L. leaves. Spectral features and relation to chlorophyll estimation. J. Plant Physiol. 1994, 143, 286–292. [Google Scholar] [CrossRef]
- Haboudane, D.; Miller, J.R.; Tremblay, N.; Zarco-Tejada, P.J.; Dextraze, L. Integrated narrow-band vegetation indices for prediction of crop chlorophyll content for application to precision agriculture. Remote Sens. Environ. 2002, 81, 416–426. [Google Scholar] [CrossRef]
- Penuelas, J.; Baret, F.; Filella, I. Semi-empirical indices to assess carotenoids/chlorophyll a ratio from leaf spectral reflectance. Photosynthetica 1995, 31, 221–230. [Google Scholar]
- Carter, G.A. Ratios of leaf reflectances in narrow wavebands as indicators of plant stress. Remote Sens. 1994, 15, 697–703. [Google Scholar] [CrossRef]
- Carter, G.A.; Cibula, W.G.; Dell, T.R. Spectral reflectance characteristics and digital imagery of a pine needle blight in the southeastern United States. Can. J. For. Res. 1996, 26, 402–407. [Google Scholar] [CrossRef]
- Datt, B. Remote sensing of chlorophyll a, chlorophyll b, chlorophyll a+ b, and total carotenoid content in eucalyptus leaves. Remote Sens. Environ. 1998, 66, 111–121. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Keydan, G.P.; Merzlyak, M.N. Three-band model for noninvasive estimation of chlorophyll, carotenoids, and anthocyanin contents in higher plant leaves. Geophys. Res. Lett. 2006, 33, 1–5. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Gritz, Y.; Merzlyak, M.N. Relationships between leaf chlorophyll content and spectral reflectance and algorithms for non-destructive chlorophyll assessment in higher plant leaves. J. Plant Physiol. 2003, 160, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Merzlyak, M.N.; Gitelson, A.A.; Chivkunova, O.B.; Rakitin, V.Y.U. Non-destructive optical detection of pigment changes during leaf senescence and fruit ripening. Physiol. Plant. 1999, 106, 135–141. [Google Scholar] [CrossRef]
- Blackburn, G.A. Spectral indices for estimating photosynthetic pigment concentrations: A test using senescent tree leaves. Int. J. Remote Sens. 1998, 19, 657–675. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Buschmann, C.; Lichtenthaler, H.K. The chlorophyll fluorescence ratio F735/F700 as an accurate measure of the chlorophyll content in plants. Remote Sens. Environ. 1999, 69, 296–302. [Google Scholar] [CrossRef]







| Data | DAI | Variables in the Model |
|---|---|---|
| Thermal | 3, 8 | Tc − Ta |
| indices | All others | Tc − Ta, TSD |
| NBHIs | 3 | PRI528, CUR, PRI CI, RARS, PRIm1, VOG3 |
| 4 | HI, BF1, BGI1, B, fWBI | |
| 5 | B, PRI528, NPQI, R, DRI PRI | |
| 6 | CUR, PRI528, RGI, RR | |
| 7 | PRI528, NPQI, CUR, WBI, B, CTR1, PRI CI, BF4 | |
| 8 | PRI528, HI, RDVI, CRI700 515, RR, WI, R, SIPI | |
| 14 | DRI PRI, PRIn, EVI, BF2, RR, BF1 | |
| NBHIs + | 3 | PRI CI, PRI528, CUR |
| Thermal | 4 | HI, BF1, BGI1, fWBI, PRI528, RGI, R |
| indices | 5 | NPQI, PRI528, STD, B, DRI PRI, RR, PRI CI, Tc − Ta |
| 6 | PRI528, CUR, Tc − Ta | |
| 7 | Tc − Ta, PRI528, B, CUR, BF4 | |
| 8 | Tc − Ta | |
| 14 | Tc − Ta, DRI PRI, PRIn, EVI, BF5 |
| Software | Modules | Methods Sections |
|---|---|---|
| Matlab version 2022a | None | 2.4.2. |
| R version 4.2.3 | ggplot2, dplyr, tidyverse, broom, gridExtra | 2.6.1., 2.6.3. |
| Python 3.8.5. | pandas, numpy, sklearn | 2.6.2. |
| Index | Days after | Confusion Matrix (%) | Classification Statistics | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Inoculation | TN | FP | FN | TP | Prec. | Recall | Acc. (%) | F1 Score | |
| Thermal indices | 3 | 4.8 | 28.5 | 9.3 | 57.3 | 0.67 | 0.86 | 62 | 0.75 |
| 4 | 2.1 | 31.2 | 8.3 | 58.4 | 0.65 | 0.88 | 61 | 0.75 | |
| 5 | 6.4 | 26.9 | 6.5 | 60.1 | 0.69 | 0.90 | 67 | 0.78 | |
| 6 | 18.8 | 14.5 | 13.1 | 53.6 | 0.79 | 0.80 | 72 | 0.80 | |
| 7 | 23.2 | 10.1 | 5.9 | 60.8 | 0.86 | 0.91 | 84 | 0.88 | |
| 8 | 27.2 | 6.1 | 3.5 | 63.2 | 0.91 | 0.95 | 90 | 0.93 | |
| 14 | 24.8 | 8.5 | 4.9 | 61.7 | 0.88 | 0.93 | 87 | 0.90 | |
| NBHIs | 3 | 17.5 | 15.9 | 7.2 | 59.5 | 0.79 | 0.89 | 77 | 0.84 |
| 4 | 22.1 | 11.2 | 6.9 | 59.7 | 0.84 | 0.90 | 82 | 0.87 | |
| 5 | 17.3 | 16.0 | 7.7 | 58.9 | 0.79 | 0.88 | 76 | 0.83 | |
| 6 | 22.4 | 10.9 | 11.3 | 55.3 | 0.84 | 0.83 | 78 | 0.83 | |
| 7 | 17.3 | 16.0 | 9.5 | 57.2 | 0.78 | 0.86 | 75 | 0.82 | |
| 8 | 12.3 | 21.1 | 11.6 | 55.1 | 0.72 | 0.83 | 67 | 0.77 | |
| 14 | 21.7 | 11.6 | 9.3 | 57.3 | 0.83 | 0.86 | 79 | 0.85 | |
| NBHIs + Thermal indices | 3 | 13.3 | 20.0 | 7.1 | 59.6 | 0.75 | 0.89 | 73 | 0.81 |
| 4 | 22.5 | 10.8 | 8.1 | 58.5 | 0.84 | 0.88 | 81 | 0.86 | |
| 5 | 16.0 | 17.3 | 7.1 | 59.6 | 0.77 | 0.89 | 76 | 0.83 | |
| 6 | 22.0 | 11.3 | 9.1 | 57.6 | 0.84 | 0.86 | 80 | 0.85 | |
| 7 | 25.5 | 7.9 | 3.6 | 63.1 | 0.89 | 0.95 | 89 | 0.92 | |
| 8 | 27.2 | 6.1 | 3.5 | 63.2 | 0.91 | 0.95 | 90 | 0.93 | |
| 14 | 30.0 | 3.3 | 5.7 | 60.9 | 0.95 | 0.91 | 91 | 0.93 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Watt, M.S.; Estarija, H.J.C.; Bartlett, M.; Main, R.; Pasquini, D.; Yorston, W.; McLay, E.; Zhulanov, M.; Dobbie, K.; Wardhaugh, K.; et al. Early Detection of Myrtle Rust on Pōhutukawa Using Indices Derived from Hyperspectral and Thermal Imagery. Remote Sens. 2024, 16, 1050. https://doi.org/10.3390/rs16061050
Watt MS, Estarija HJC, Bartlett M, Main R, Pasquini D, Yorston W, McLay E, Zhulanov M, Dobbie K, Wardhaugh K, et al. Early Detection of Myrtle Rust on Pōhutukawa Using Indices Derived from Hyperspectral and Thermal Imagery. Remote Sensing. 2024; 16(6):1050. https://doi.org/10.3390/rs16061050
Chicago/Turabian StyleWatt, Michael S., Honey Jane C. Estarija, Michael Bartlett, Russell Main, Dalila Pasquini, Warren Yorston, Emily McLay, Maria Zhulanov, Kiryn Dobbie, Katherine Wardhaugh, and et al. 2024. "Early Detection of Myrtle Rust on Pōhutukawa Using Indices Derived from Hyperspectral and Thermal Imagery" Remote Sensing 16, no. 6: 1050. https://doi.org/10.3390/rs16061050
APA StyleWatt, M. S., Estarija, H. J. C., Bartlett, M., Main, R., Pasquini, D., Yorston, W., McLay, E., Zhulanov, M., Dobbie, K., Wardhaugh, K., Hossain, Z., Fraser, S., & Buddenbaum, H. (2024). Early Detection of Myrtle Rust on Pōhutukawa Using Indices Derived from Hyperspectral and Thermal Imagery. Remote Sensing, 16(6), 1050. https://doi.org/10.3390/rs16061050