
Spatial and Temporal Changes in Vegetation Phenology at Middle and High Latitudes of the Northern Hemisphere over the Past Three Decades



Abstract
:
1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
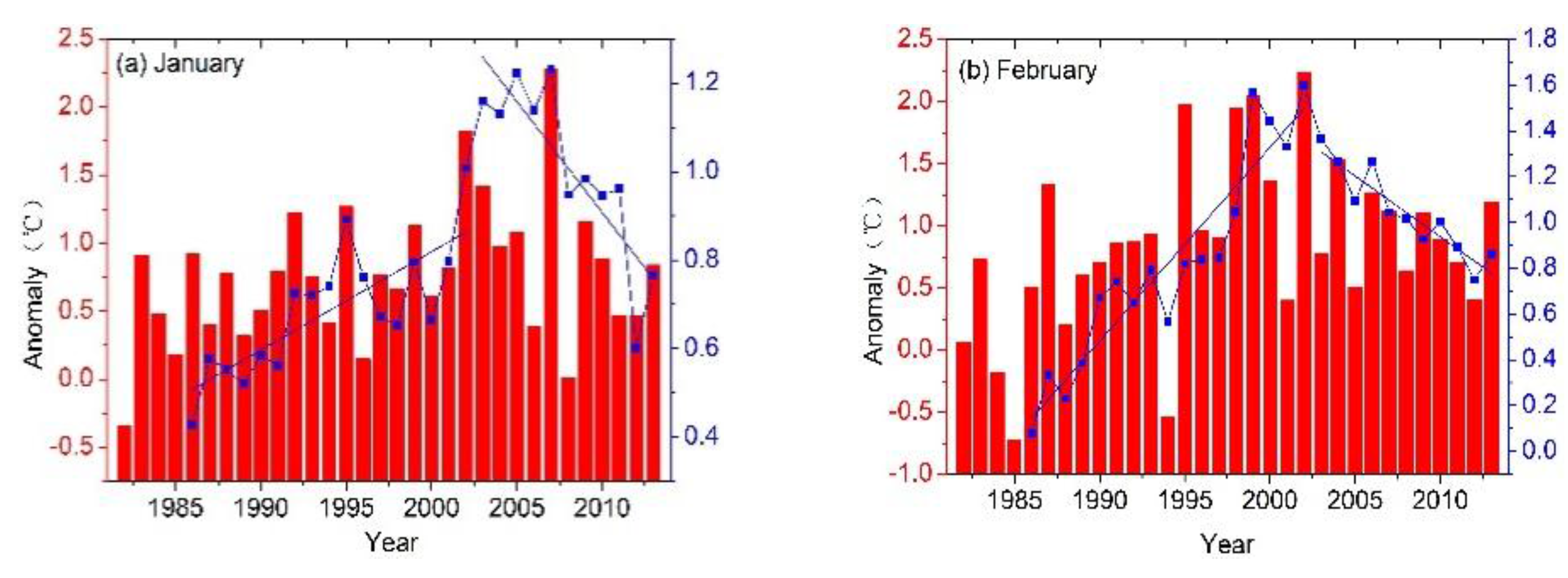{kind=link}
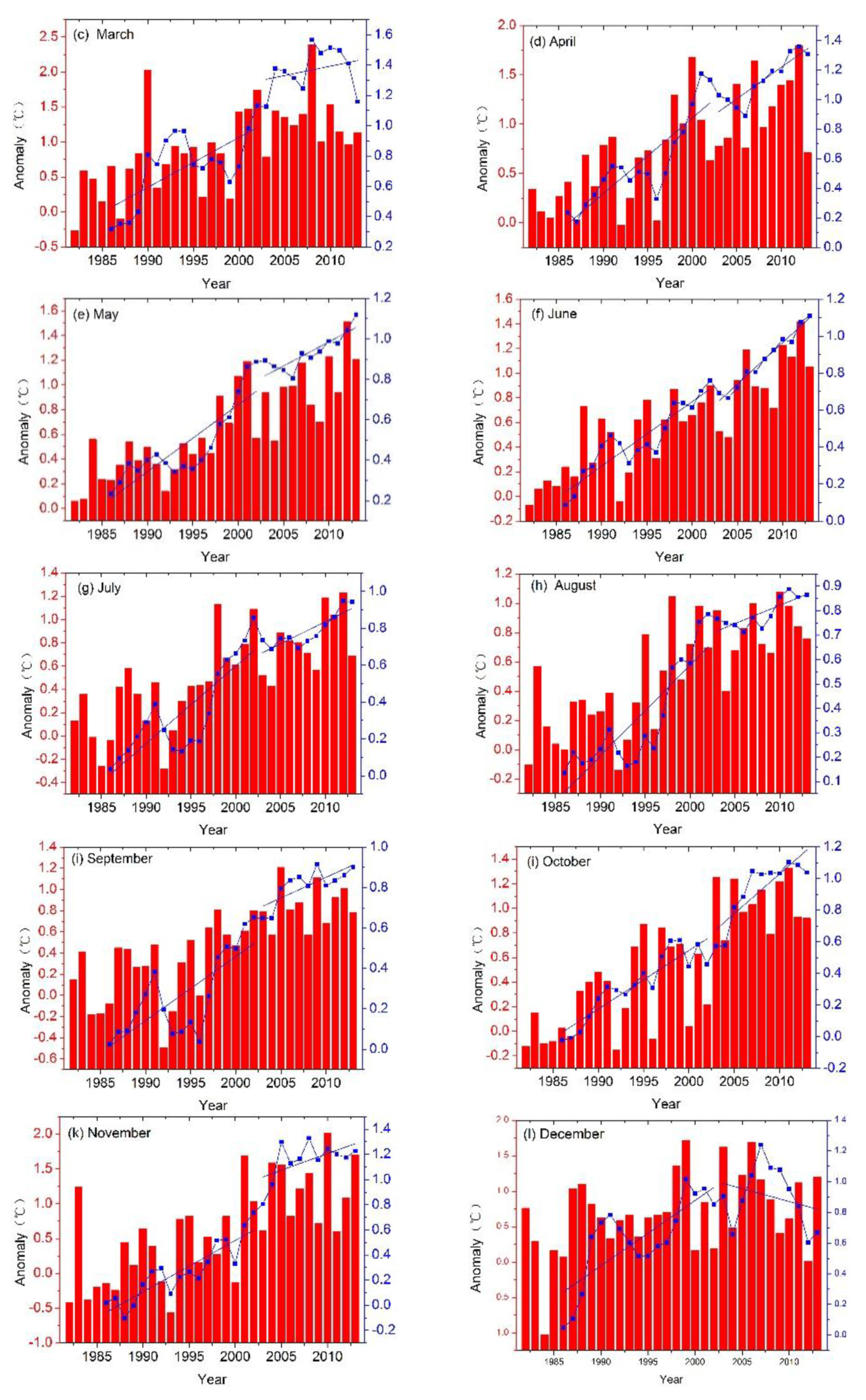{kind=link}
| Reference | Period | Type | Region | SOS | EOS | LOS |
|---|---|---|---|---|---|---|
| Myneni et al. (1997) [19] | 1982–1991 | AVHRR | ≥40°N | −8 | 4 | 12 |
| Tuker et al. (2001) [20] | 1982–1991 | AVHRR | 45°N–75°N | −6.2 | 4.4 | |
| Tuker et al. (2001) [20] | 1992–1999 | AVHRR | 45°N–75°N | −2.4 | 0.6 | |
| Zeng et al. (2011) [6] | 2000–2008 | AVHRR | Arctic | −0.2 | 2 | 2.2 |
| Piao et al. (2006) [21] | 1982–1999 | AVHRR | China | −7.9 | 3.7 | 11.6 |
| Stockli et al. (2004) [22] | 1982–2000 | AVHRR | Europe | −6 | 4.7 | 10.7 |
| Zhou et al. (2001) [23] | 1982–1999 | AVHRR | Eurasia | −3.3 | 6.1 | 13.3 |
| de Beurs et al. (2005) [24] | 1985–2000 | AVHRR | Eurasia | −4.5 | ||
| Zeng et al. (2011) [6] | 2000–2008 | AVHRR | Eurasia | −0.3 | 2.6 | 2.9 |
| Zhou et al. (2001) [23] | 1982–1999 | AVHRR | 40°N–70°N North America | −4.3 | 2 | 6.3 |
| de Beurs et al. (2005) [24] | 1985–1999 | AVHRR | North America | −6.6 | ||
| Zhu et al. (2012) [25] | 1982–2006 | AVHRR | North America | −1.3 | 5.5 | 6.8 |
| Zeng et al. (2011) [6] | 2000–2008 | AVHRR | North America | −0.1 | 1.1 | 1.2 |
| Jeong et al. (2011) [26] | 1982–1999 | AVHRR | Northern Hemisphere | −3.1 | 2.5 | 5.6 |
| Jeong et al. (2011) [26] | 2000–2008 | AVHRR | Northern Hemisphere | −0.2 | 2.6 | 2.8 |
| Wang et al. (2015) [27] | 1982–2011 | AVHRR | 30°N–75°N | −1.4 |
2. Materials and Methods
2.1. GIMMS NDVI3g Dataset
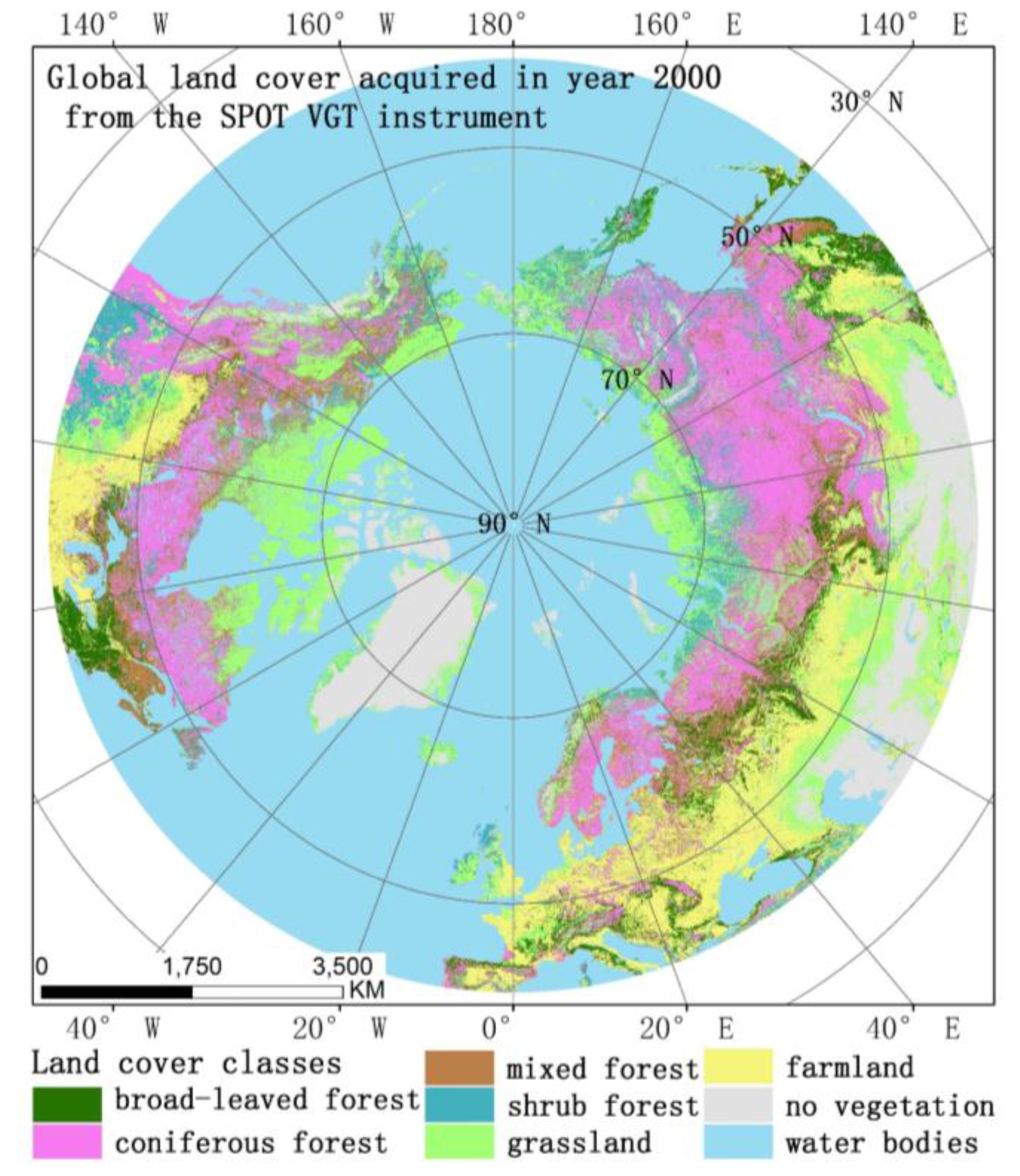
2.2. Land Cover Data
2.3. Climate Data
2.4. Phenology Metrics

2.5. Statistical Analysis
3. Results and Discussion
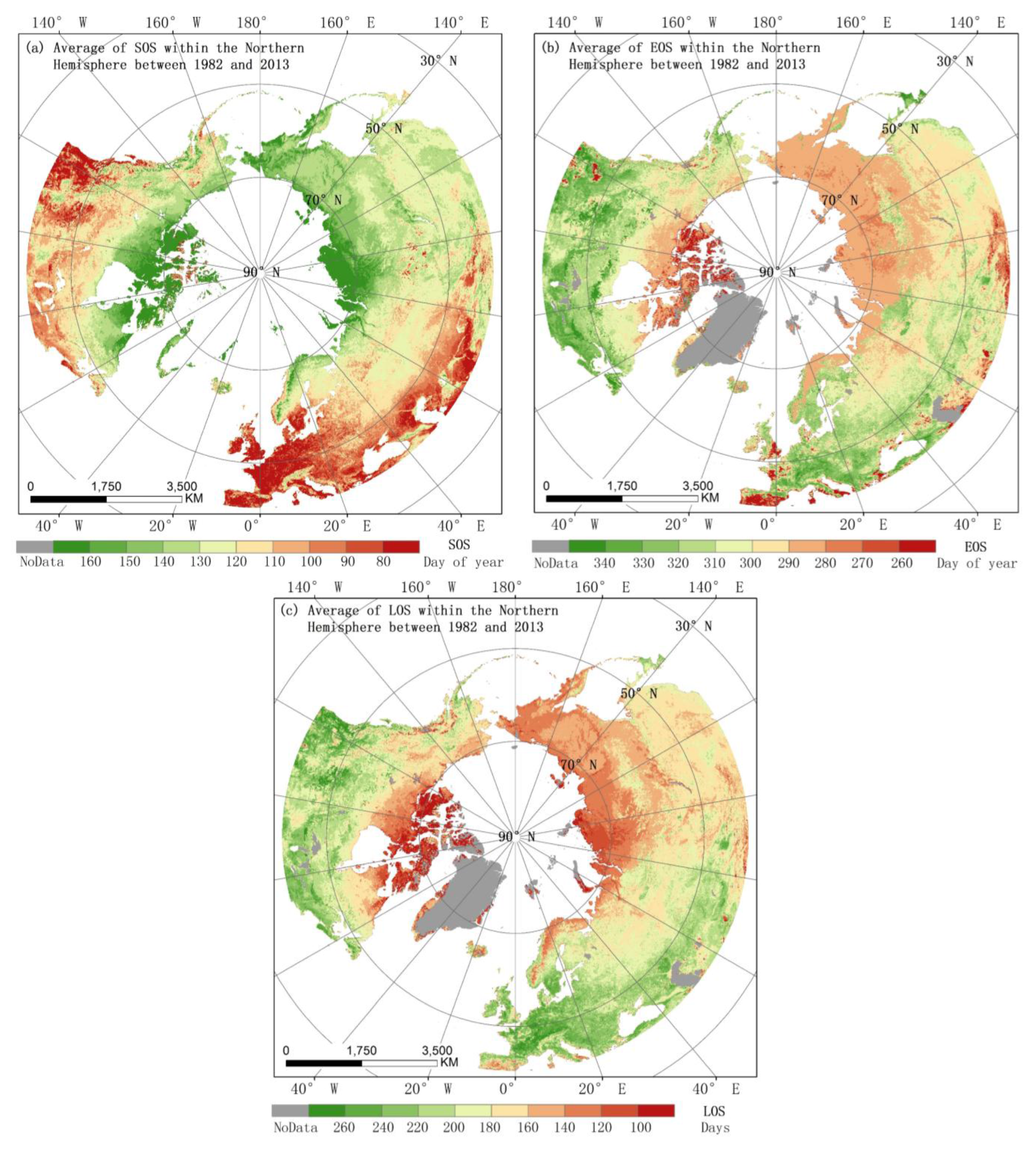
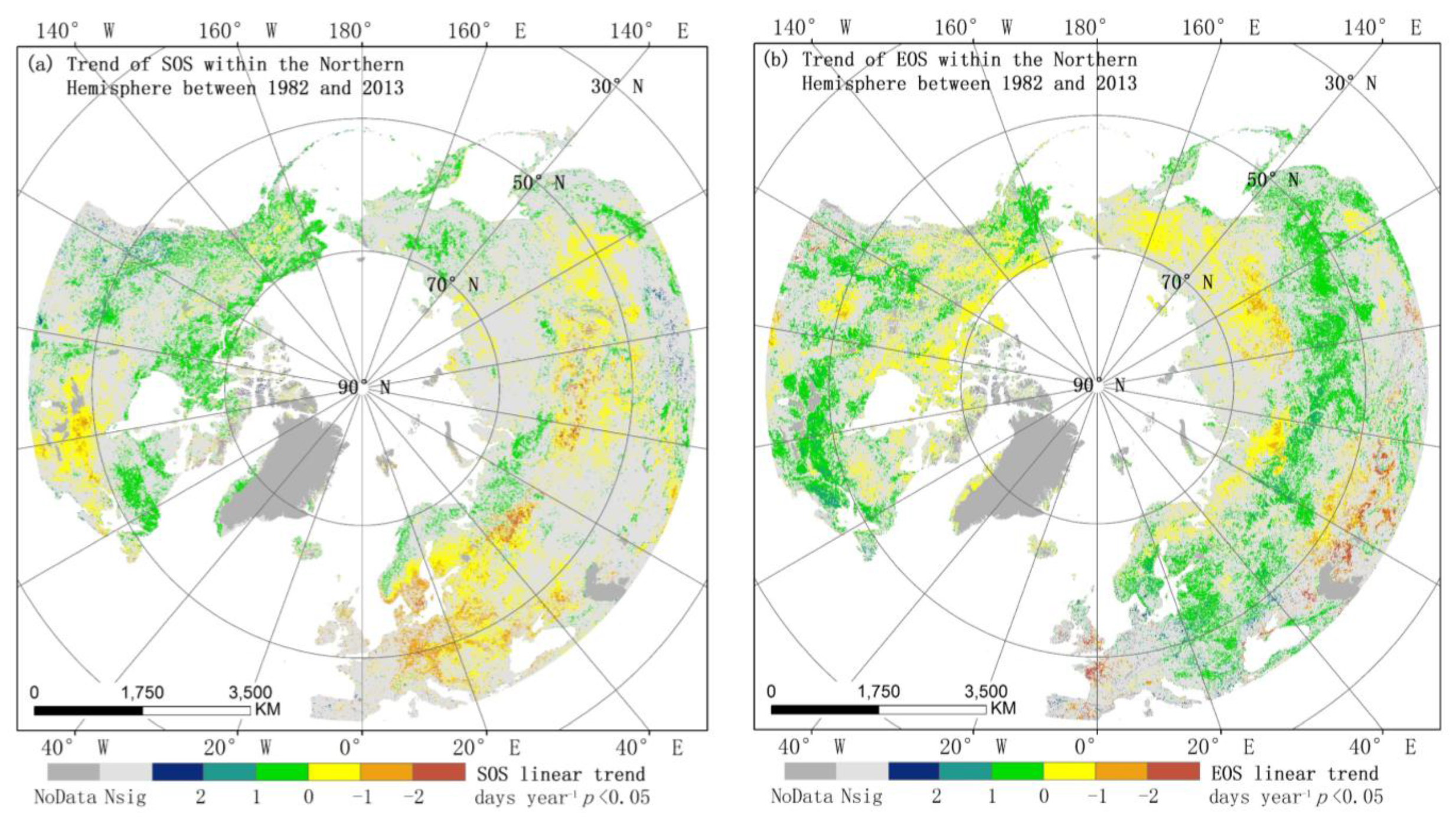
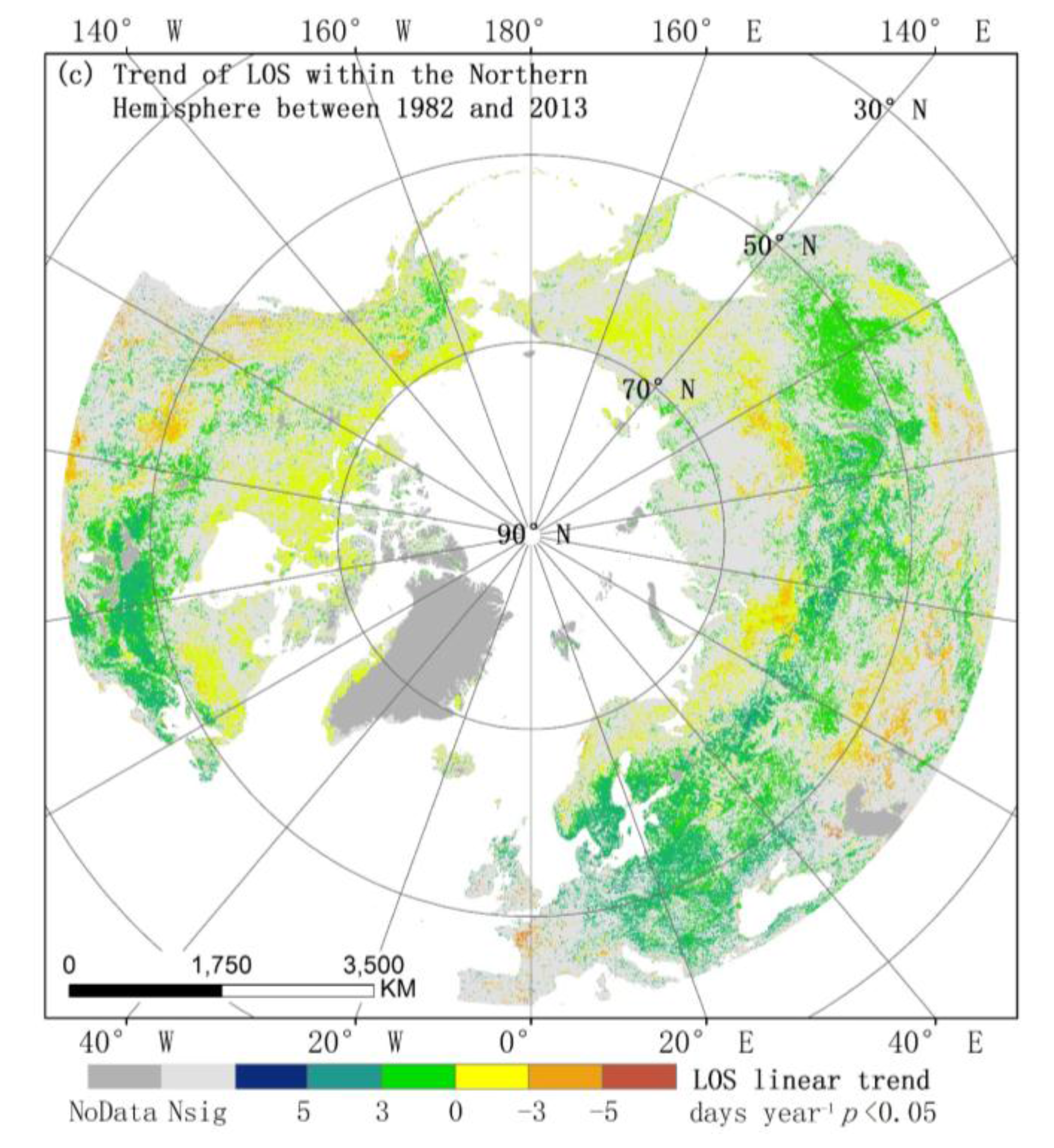
3.1. Spatial Patterns of Vegetation Phenology

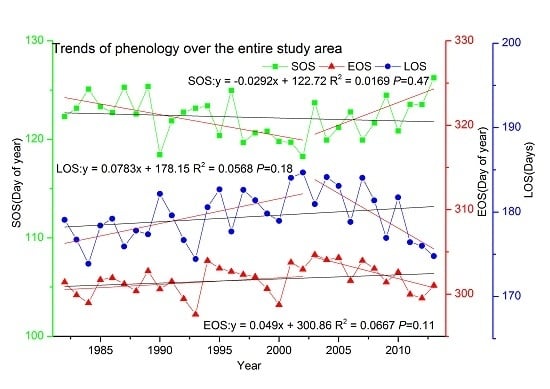
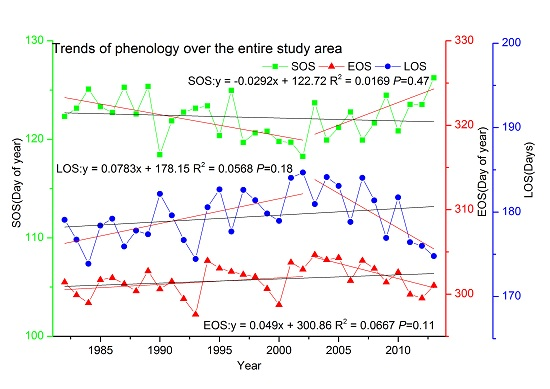
3.2. Trends in Phenology
3.2.1. Spatial Patterns of Phenological Trends


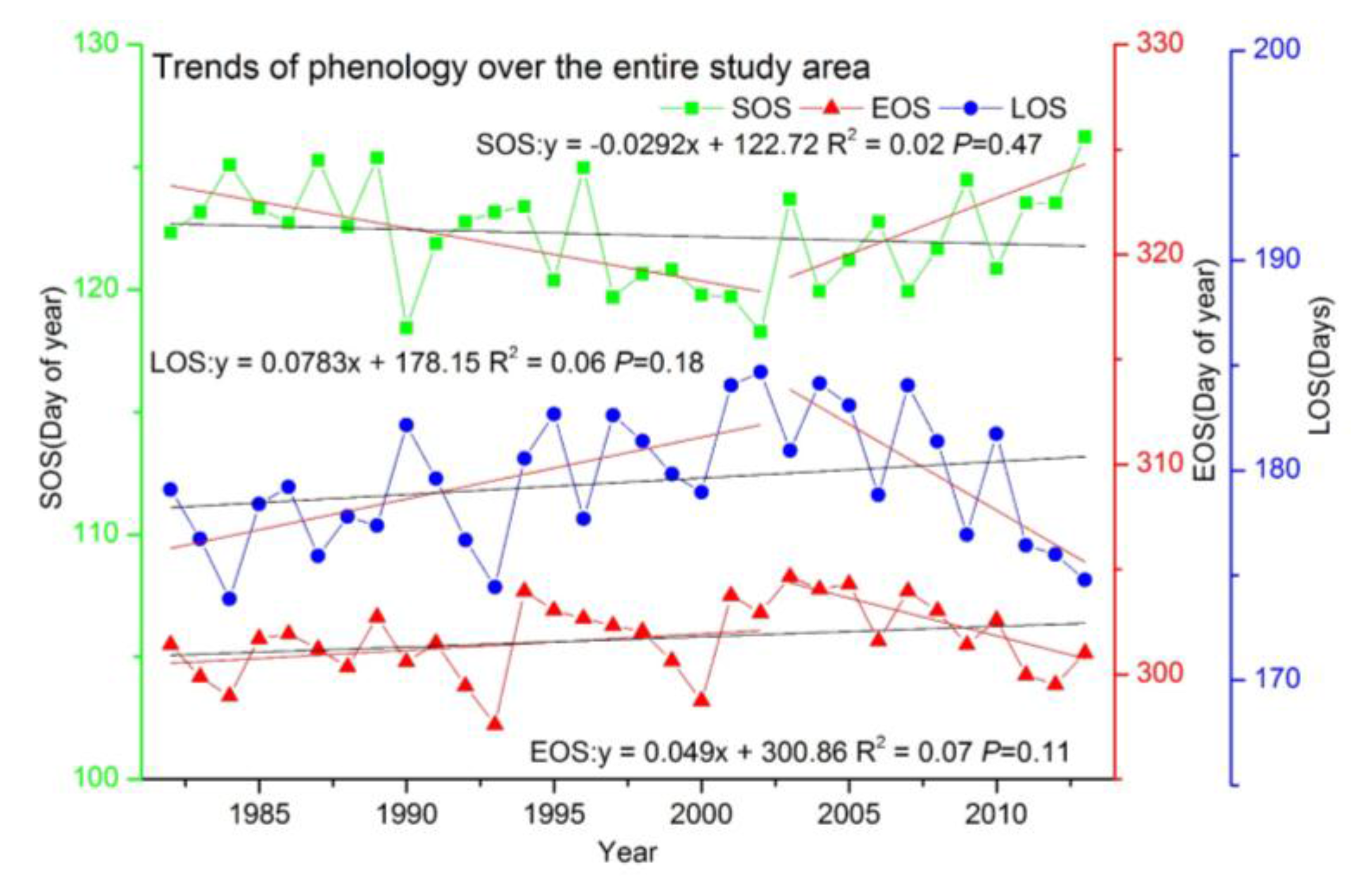
3.2.2. Trends over the Entire Study Area

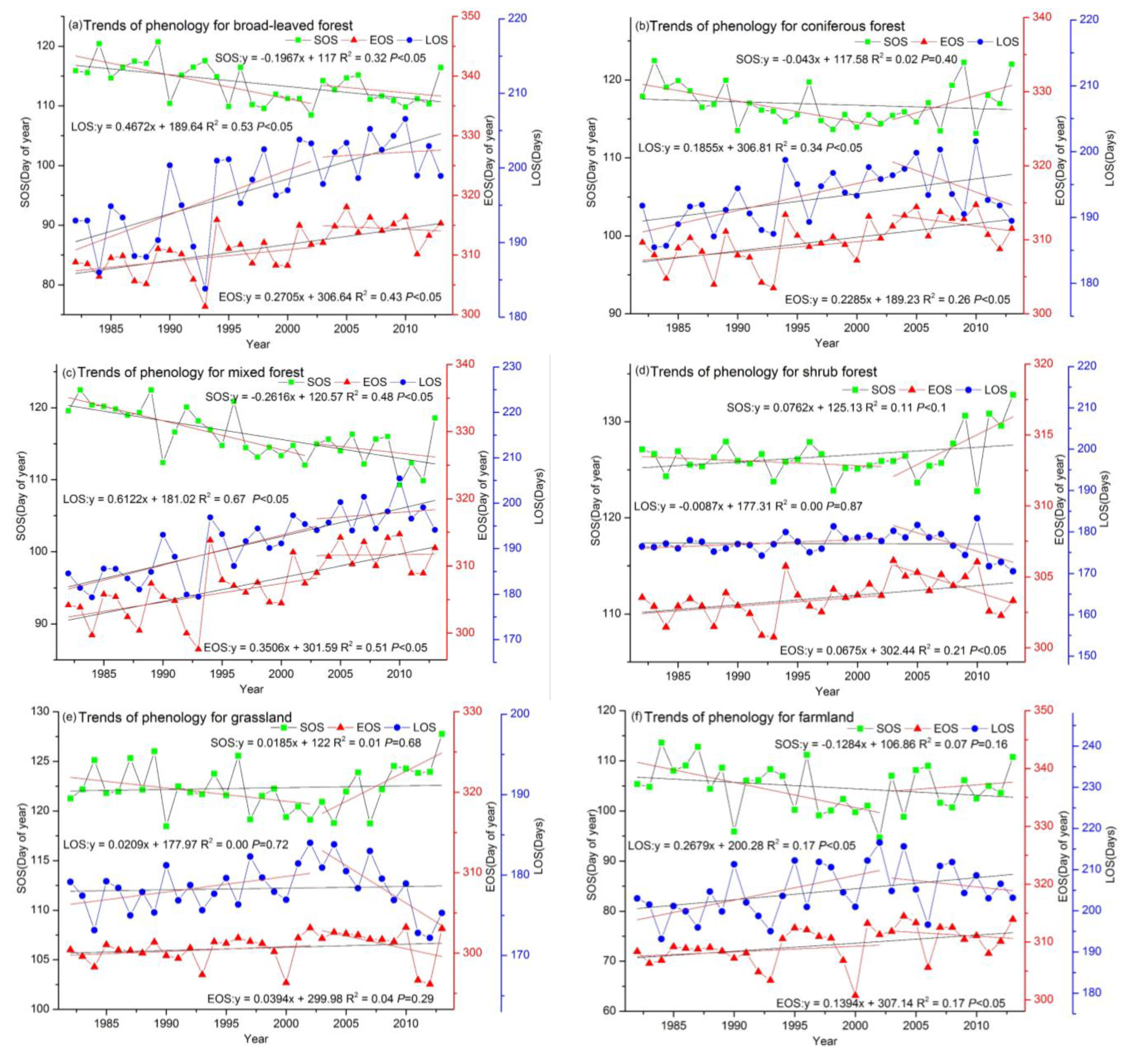
3.2.3. Phenological Trends for Different Land Cover Types

| 1982–2002 | 2003–2013 | 1982–2013 | ||
|---|---|---|---|---|
| SOS | a | −0.37 ‡ | 0.018 | −0.20 ‡ |
| b | −0.28 ‡ | 0.44 * | −0.04 | |
| c | −0.38 ‡ | −0.06 | −0.26 ‡ | |
| d | −0.06 | 0.54 † | 0.08 * | |
| e | −0.11 | 0.61 ‡ | 0.02 | |
| f | −0.42 † | 0.52 | −0.13 | |
| EOS | a | 0.19 | 0.03 | 0.27 ‡ |
| b | 0.14 | −0.08 | 0.19 ‡ | |
| c | 0.27 * | 0.18 | 0.35 ‡ | |
| d | 0.06 | −0.19 * | 0.068 † | |
| e | 0.02 | −0.32 | 0.040 | |
| f | 0.07 | −0.10 | 0.14 † | |
| LOS | a | 0.56 ‡ | 0.02 | 0.47 ‡ |
| b | 0.41 ‡ | −0.53 | 0.23 ‡ | |
| c | 0.65 ‡ | 0.24 | 0.61 ‡ | |
| d | 0.12 * | −0.71 † | −0.01 | |
| e | 0.13 | −0.93 ‡ | 0.02 | |
| f | 0.49 † | −0.63 | 0.26 † | |
| * p < 0.1, † p < 0.05, ‡ p < 0.01 | ||||
4. Discussion
4.1. Trends in Phenology
4.2. Climate Change and Phenology


| Jan | Feb | Mar | Apr | May | Jun | Jul | Aug | Sep | Oct | Nov | Dec | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| a | SOS | −0.35 | −0.45 ‡ | −0.54 ‡ | −0.70 ‡ | −0.46 ‡ | −0.62 ‡ | −0.60 ‡ | −0.65 ‡ | −0.58 ‡ | −0.51 ‡ | −0.51 ‡ | −0.14 |
| EOS | 0.2 | 0.03 | 0.49 ‡ | 0.55 ‡ | 0.61 ‡ | 0.62 ‡ | 0.54 ‡ | 0.58 ‡ | 0.64 ‡ | 0.67 ‡ | 0.77 ‡ | 0.2 | |
| LOS | 0.32 * | 0.27 | 0.61 ‡ | 0.73 ‡ | 0.64 ‡ | 0.73 ‡ | 0.67 ‡ | 0.73 ‡ | 0.73 ‡ | 0.70 ‡ | 0.77 ‡ | 0.21 | |
| b | SOS | −0.34 * | −0.28 | −0.31 * | −0.43 † | −0.27 | −0.30 * | −0.29 | −0.28 | −0.17 | −0.23 | −0.11 | −0.21 |
| EOS | 0.19 | 0.095 | 0.41 † | 0.49 ‡ | 0.47 ‡ | 0.49 ‡ | 0.50 ‡ | 0.57 ‡ | 0.62 ‡ | 0.68 ‡ | 0.70 ‡ | 0.31 * | |
| LOS | 0.35 * | 0.24 | 0.48 ‡ | 0.62 ‡ | 0.51 ‡ | 0.53 ‡ | 0.54 ‡ | 0.58 ‡ | 0.55 ‡ | 0.63 ‡ | 0.57 ‡ | 0.36 † | |
| c | SOS | −0.36 † | −0.37 † | −0.59 ‡ | −0.84 ‡ | −0.70 ‡ | −0.79 ‡ | −0.72 ‡ | −0.72 ‡ | −0.66 ‡ | −0.66 ‡ | −0.51 ‡ | −0.19 |
| EOS | 0.22 | 0.06 | 0.48 ‡ | 0.59 ‡ | 0.62 ‡ | 0.65 ‡ | 0.57 ‡ | 0.64 ‡ | 0.69v | 0.73 ‡ | 0.77 ‡ | 0.27 | |
| LOS | 0.32 * | 0.22 | 0.61 ‡ | 0.81 ‡ | 0.76 ‡ | 0.82 ‡ | 0.74 ‡ | 0.78 ‡ | 0.79 ‡ | 0.81 ‡ | 0.76 ‡ | 0.27 | |
| d | SOS | −0.15 | −0.09 | 0 | 0.03 | 0.11 | 0.16 | 0.044 | 0.02 | 0.19 | 0.15 | 0.134 | 0 |
| EOS | 0.24 | 0.11 | 0.35 † | 0.45 ‡ | 0.42 † | 0.37 † | 0.42 † | 0.54 ‡ | 0.56 ‡ | 0.58 ‡ | 0.58 ‡ | 0.32 * | |
| LOS | 0.25 | 0.13 | 0.18 | 0.21 | 0.12 | 0.06 | 0.18 | 0.26 | 0.13 | 0.18 | 0.19 | 0.15 | |
| e | SOS | −0.38 † | −0.19 | −0.35 † | −0.25 | 0.1 | 0.05 | 0.04 | −0.07 | −0.04 | 0 | −0.05 | 0 |
| EOS | 0.27 | 0.19 | 0.19 | −0.12 | 0.03 | 0.1 | 0.12 | 0.16 | 0.2 | 0.25 | 0.49 ‡ | 0.35 ‡ | |
| LOS | 0.47 ‡ | 0.27 | 0.40 † | 0.11 | −0.06 | 0.03 | 0.045 | 0.16 | 0.15 | 0.17 | 0.36 † | 0.23 | |
| f | SOS | −0.33 ‡ | −0.43 ‡ | −0.60 ‡ | −0.49 ‡ | −0.17 | −0.36 † | −0.36 † | −0.38 † | −0.32 * | −0.27 | −0.39 † | −0.06 |
| EOS | 0.23 | 0.05 | 0.23 | 0.2 | 0.32 † | 0.36 † | 0.36 † | 0.37 † | 0.46 ‡ | 0.50 ‡ | 0.63 ‡ | 0.2 | |
| LOS | 0.37 † | 0.36 † | 0.59 ‡ | 0.49 ‡ | 0.30 ‡ | 0.47 ‡ | 0.48 ‡ | 0.50 ‡ | 0.49 ‡ | 0.47 ‡ | 0.64 ‡ | 0.15 | |
| * p < 0.1, † p < 0.05, ‡ p < 0.01 | |||||||||||||
4.3. The Influence of Other Factors on Vegetation Phenology
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Parry, M.L.; Canziani, O.F.; Palutikof, J.P.; van der Linden, P.J.; Hanson, C.E. IPCC, 2007: Climate Change 2007: Impacts, Adaptation and Vulnerability; Contribution of Working Group II to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Lucht, W.; Prentice, I.C.; Myneni, R.B.; Sitch, S.; Friedlingstein, P.; Cramer, W.; Bousquet, P.; Buermann, W.; Smith, B. Climatic Control of the High-Latitude Vegetation Greening Trend and Pinatubo Effect. Science 2002, 296, 1687–1689. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.-J.; Ho, C.-H.; Kim, B.-M.; Feng, S.; Medvigy, D. Non-linear response of vegetation to coherent warming over northern high latitudes. Remote Sens. Lett. 2013, 4, 123–130. [Google Scholar] [CrossRef]
- Barichivich, J.; Briffa, K.R.; Myneni, R.B.; Osborn, T.J.; Melvin, T.M.; Ciais, P.; Piao, S.; Tucker, C. Large-scale variations in the vegetation growing season and annual cycle of atmospheric CO2 at high northern latitudes from 1950 to 2011. Global Chang. Biol. 2013, 19, 3167–3183. [Google Scholar] [CrossRef] [PubMed]
- Berner, L.T.; Beck, P.S.; Bunn, A.G.; Lloyd, A.H.; Goetz, S.J. High-latitude tree growth and satellite vegetation indices: Correlations and trends in Russia and Canada (1982–2008). J. Geophys. Res. Biogeosci. 2011. [Google Scholar] [CrossRef]
- Zeng, H.; Jia, G.; Epstein, H. Recent changes in phenology over the northern high latitudes detected from multi-satellite data. Environ. Res. Lett. 2011, 6, 045508. [Google Scholar] [CrossRef]
- Badeck, F.-W.; Bondeau, A.; Böttcher, K.; Doktor, D.; Lucht, W.; Schaber, J.; Sitch, S. Responses of spring phenology to climate change. New Phytol. 2004, 162, 295–309. [Google Scholar] [CrossRef]
- Hurlbert, A.H.; Haskell, J.P. The effect of energy and seasonality on avian species richness and community composition. Am. Nat. 2003, 161, 83–97. [Google Scholar] [CrossRef] [PubMed]
- Hebblewhite, M.; Merrill, E.; McDermid, G. A multi-scale test of the forage maturation hypothesis in a partially migratory ungulate population. Ecol. Monogr. 2008, 78, 141–166. [Google Scholar] [CrossRef]
- Jönsson, A.M.; Eklundh, L.; Hellström, M.; Bärring, L.; Jönsson, P. Annual changes in MODIS vegetation indices of Swedish coniferous forests in relation to snow dynamics and tree phenology. Remote Sens. Environ. 2010, 114, 2719–2730. [Google Scholar] [CrossRef]
- Van Leeuwen, W.J.D. Monitoring the effects of forest restoration treatments on post-fire vegetation recovery with MODIS multitemporal data. Sensors 2008, 8, 2017–2042. [Google Scholar] [CrossRef]
- Sakamoto, T.; Yokozawa, M.; Toritani, H.; Shibayama, M.; Ishitsuka, N.; Ohno, H. A crop phenology detection method using time-series MODIS data. Remote Sens. Environ. 2005, 96, 366–374. [Google Scholar] [CrossRef]
- Heumann, B.W.; Seaquist, J.W.; Eklundh, L.; Jönsson, P. AVHRR derived phenological change in the Sahel and Soudan, Africa, 1982–2005. Remote Sens. Environ. 2007, 108, 385–392. [Google Scholar] [CrossRef]
- Cleland, E.E.; Chuine, I.; Menzel, A.; Mooney, H.A.; Schwartz, M.D. Shifting plant phenology in response to global change. Trends Ecol. Evol. 2007, 22, 357–365. [Google Scholar] [CrossRef] [PubMed]
- Hmimina, G.; Dufrêne, E.; Pontailler, J.-Y.; Delpierre, N.; Aubinet, M.; Caquet, B.; de Grandcourt, A.; Burban, B.; Flechard, C.; Granier, A. Evaluation of the potential of MODIS satellite data to predict vegetation phenology in different biomes: An investigation using ground-based NDVI measurements. Remote Sens. Environ. 2013, 132, 145–158. [Google Scholar] [CrossRef]
- Hermance, J.F.; Jacob, R.W.; Bradley, B.A.; Mustard, J.F. Extracting phenological signals from multiyear AVHRR NDVI time series: Framework for applying high-order annual splines with roughness damping. IEEE Trans. Geosci. Remote Sens. 2007, 45, 3264–3276. [Google Scholar] [CrossRef]
- Chen, X.; Tan, Z.; Schwartz, M.D.; Xu, C. Determining the growing season of land vegetation on the basis of plant phenology and satellite data in Northern China. Int. J. Biometeorol. 2000, 44, 97–101. [Google Scholar] [CrossRef] [PubMed]
- Wagenseil, H.; Samimi, C. Assessing spatio-temporal variations in plant phenology using Fourier analysis on NDVI time series: Results from a dry savannah environment in Namibia. Int. J. Remote Sens. 2006, 27, 3455–3471. [Google Scholar] [CrossRef]
- Myneni, R.B.; Keeling, C.D.; Tucker, C.J.; Asrar, G.; Nemani, R.R. Increased plant growth in the northern high latitudes from 1981 to 1991. Nature 1997, 386, 698–702. [Google Scholar] [CrossRef]
- Tucker, C.J.; Slayback, D.A.; Pinzon, J.E.; Los, S.O.; Myneni, R.B.; Taylor, M.G. Higher northern latitude normalized difference vegetation index and growing season trends from 1982 to 1999. Int. J. Biometeorol. 2001, 45, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Piao, S.; Fang, J.; Zhou, L.; Ciais, P.; Zhu, B. Variations in satellite-derived phenology in China’s temperate vegetation. Global Chang. Biol. 2006, 12, 672–685. [Google Scholar] [CrossRef]
- Stöckli, R.; Vidale, P.L. European plant phenology and climate as seen in a 20-year AVHRR land-surface parameter dataset. Int. J. Remote Sens. 2004, 25, 3303–3330. [Google Scholar] [CrossRef]
- Zhou, L.; Tucker, C.J.; Kaufmann, R.K.; Slayback, D.; Shabanov, N.V.; Myneni, R.B. Variations in northern vegetation activity inferred from satellite data of vegetation index during 1981 to 1999. J. Geophys. Res. Atmos. 2001, 106, 20069–20083. [Google Scholar] [CrossRef]
- De Beurs, K.M.; Henebry, G.M. Land surface phenology and temperature variation in the International Geosphere–Biosphere Program high-latitude transects. Global Chang. Biol. 2005, 11, 779–790. [Google Scholar] [CrossRef]
- Zhu, W.; Tian, H.; Xu, X.; Pan, Y.; Chen, G.; Lin, W. Extension of the growing season due to delayed autumn over mid and high latitudes in North America during 1982–2006. Global Ecol. Biogeogr. 2012, 21, 260–271. [Google Scholar] [CrossRef]
- JEONG, S.-J.; HO, C.-H.; GIM, H.-J.; Brown, M.E. Phenology shifts at start vs. end of growing season in temperate vegetation over the Northern Hemisphere for the period 1982–2008. Global Chang. Biol. 2011, 17, 2385–2399. [Google Scholar] [CrossRef]
- Wang, X.; Piao, S.; Xu, X.; Ciais, P.; MacBean, N.; Myneni, R.B.; Li, L. Has the advancing onset of spring vegetation green-up slowed down or changed abruptly over the last three decades? Global Ecol. Biogeogr. 2015, 24, 621–631. [Google Scholar] [CrossRef]
- Zhu, Z.; Bi, J.; Pan, Y.; Ganguly, S.; Anav, A.; Xu, L.; Samanta, A.; Piao, S.; Nemani, R.R.; Myneni, R.B. Global data sets of vegetation leaf area index (LAI) 3g and Fraction of Photosynthetically Active Radiation (FPAR) 3g derived from Global Inventory Modeling and Mapping Studies (GIMMS) Normalized Difference Vegetation Index (NDVI3g) for the period 1981 to 2011. Remote Sens. 2013, 5, 927–948. [Google Scholar]
- Dardel, C.; Kergoat, L.; Hiernaux, P.; Mougin, E.; Grippa, M.; Tucker, C.J. Re-greening Sahel: 30 years of remote sensing data and field observations (Mali, Niger). Remote Sens. Environ. 2014, 140, 350–364. [Google Scholar] [CrossRef]
- Pinzon, J.E.; Tucker, C.J. A non-stationary 1981–2012 AVHRR NDVI3g time series. Remote Sens. 2014, 6, 6929–6960. [Google Scholar] [CrossRef]
- NASA Ames Ecological Forecasting Lab. Available online: http://ecocast.arc.nasa.gov/data/pub/gimms/3g.v0/ (accessed on 15 September 2014).
- Bartholomé, E.; Belward, A.S. GLC2000: A new approach to global land cover mapping from earth observation data. Int. J. Remote Sens. 2005, 26, 1959–1977. [Google Scholar] [CrossRef]
- NOAA’s National Centers. Available online: http://www.ncdc.noaa.gov/cag/time-series/global/ (accessed on 1 May 2015).
- Smith, T.M.; Reynolds, R.W.; Peterson, T.C.; Lawrimore, J. Improvements to NOAA’s Historical Merged Land–Ocean Surface Temperature Analysis (1880–2006). J. Clim. 2008, 21, 2283. [Google Scholar] [CrossRef]
- Viovy, N.; Arino, O.; Belward, A.S. The Best Index Slope Extraction (BISE): A method for reducing noise in NDVI time-series. Int. J. Remote Sens. 1992, 13, 1585–1590. [Google Scholar] [CrossRef]
- Jiang, N.; Zhu, W.; Mou, M.; Wang, L.; Zhang, J. A phenology-preserving filtering method to reduce noise in NDVI time series. In Proceedings of the 2012 IEEE International Geoscience and Remote Sensing Symposium (IGARSS), Munich, Germany, 22–27 July 2012; pp. 2384–2387.
- Song, Y.; Chen, P.; Wan, Y.; Shen, S. Application of Hybrid Classification Method Based on Fourier Transform to Time-Series NDVI Images. In Proceedings of the Congress on Image and Signal Processing, Sanya, China, 27–30 May 2008; pp. 634–638.
- Chen, J.; Jönsson, P.; Tamura, M.; Gu, Z.; Matsushita, B.; Eklundh, L. A simple method for reconstructing a high-quality NDVI time-series data set based on the Savitzky–Golay filter. Remote Sens. Environ. 2004, 91, 332–344. [Google Scholar] [CrossRef]
- White, M.A.; de Beurs, K.M.; Didan, K.; Inouye, D.W.; Richardson, A.D.; Jensen, O.P.; O’Keefe, J.; Zhang, G.; Nemani, R.R.; van Leeuwen, W.J.D. Intercomparison, interpretation, and assessment of spring phenology in North America estimated from remote sensing for 1982–2006. Global Chang. Biol. 2009, 15, 2335–2359. [Google Scholar] [CrossRef]
- Brown, M.E.; Beurs, K.D.; Vrieling, A. The response of African land surface phenology to large scale climate oscillations. Remote Sens. Environ. 2010, 114, 2286–2296. [Google Scholar] [CrossRef]
- Jonsson, P.; Eklundh, L. TIMESAT—A program for analyzing time-series of satellite sensor data. Comput. Geosci. 2004, 30, 833–845. [Google Scholar] [CrossRef]
- Martínez, B.; Gilabert, M.A. Vegetation dynamics from NDVI time series analysis using the wavelet transform. Remote Sens. Environ. 2009, 113, 1823–1842. [Google Scholar] [CrossRef]
- Metsämäki, S.; Vepsäläinen, J.; Pulliainen, J.; Sucksdorff, Y. Improved linear interpolation method for the estimation of snow-covered area from optical data. Remote Sens. Environ. 2002, 82, 64–78. [Google Scholar] [CrossRef]
- Jonsson, P.; Eklundh, L. Seasonality extraction by function fitting to time-series of satellite sensor data. IEEE Trans. Geosci. Remote Sens. 2002, 40, 1824–1832. [Google Scholar] [CrossRef]
- Bachoo, A.; Archibald, S. Influence of Using Date-Specific Values when Extracting Phenological Metrics from 8-day Composite NDVI Data. In Proceedings of the International Workshop on the Analysis of Multi-temporal Remote Sensing Images, Leuven, Belgium, 18–20 July 2007; pp. 1–4.
- Hird, J.N.; McDermid, G.J. Noise reduction of NDVI time series: An empirical comparison of selected techniques. Remote Sens. Environ. 2009, 113, 248–258. [Google Scholar] [CrossRef]
- Peppin, D.; Fulé, P.Z.; Sieg, C.H.; Beyers, J.L.; Hunter, M.E. Post-wildfire seeding in forests of the western United States: An evidence-based review. For. Ecol. Manag. 2010, 260, 573–586. [Google Scholar] [CrossRef]
- Steenkamp, K.; Wessels, K.; Archibald, S.; von Maltitz, G. Long-Term Phenology and Variability of Southern African Vegetation. In Proceedings of the IEEE International Geoscience and Remote Sensing Symposium, Boston, MA, USA, 7–11 July 2008; Volume 3, pp. 816–819.
- Zhao, J.; Wang, Y.; Hashimoto, H.; Melton, F.S.; Hiatt, S.H.; Zhang, H.; Nemani, R.R. The variation of land surface phenology from 1982 to 2006 along the Appalachian trail. IEEE Trans. Geosci. Remote Sens. 2013, 51, 2087–2095. [Google Scholar] [CrossRef]
- Cohen, J.L.; Furtado, J.C.; Barlow, M.; Alexeev, V.A.; Cherry, J.E. Asymmetric seasonal temperature trends. Geophys. Res. Lett. 2012, 39, 54–62. [Google Scholar] [CrossRef]
- Siebert, S.; Ewert, F. Spatio-temporal patterns of phenological development in Germany in relation to temperature and day length. Agric. For. Meteorol. 2012, 152, 44–57. [Google Scholar] [CrossRef]
- Menzel, A.; Sparks, T.H.; Estrella, N.; Koch, E.; Aasa, A.; Ahas, R.; Alm-Kübler, K.; Bissolli, P.; Braslavská, O.; Briede, A. European phenological response to climate change matches the warming pattern. Global Chang. Biol. 2006, 12, 1969–1976. [Google Scholar] [CrossRef]
- Estrella, N.; Sparks, T.H.; Menzel, A. Trends and temperature response in the phenology of crops in Germany. Global Chang. Biol. 2007, 13, 1737–1747. [Google Scholar] [CrossRef]
- Estrella, N.; Sparks, T.H.; Menzel, A.; Sparks, T. Effects of temperature, phase type and timing, location, and human density on plant phenological responses in Europe. Clim. Res. 2009, 39, 235–248. [Google Scholar] [CrossRef]
- Menzel, A.; Fabian, P. Growing season extended in Europe. Nature 1999, 397, 659. [Google Scholar] [CrossRef]
- Chmielewski, F.M.; Rotzer, T. Annual and spatial variability of the beginning of growing season in Europe in relation to air temperature changes. Clim. Res. 2002, 19, 257–264. [Google Scholar] [CrossRef]
- Maignan, F.; Bréon, F.M.; Bacour, C.; Demarty, J.; Poirson, A. Interannual vegetation phenology estimates from global AVHRR measurements : Comparison with in situ data and applications. Remote Sens. Environ. 2008, 112, 496–505. [Google Scholar] [CrossRef]
- Chen, X.; Tung, K.-K. Climate. Varying planetary heat sink led to global-warming slowdown and acceleration. Science 2014, 345, 897–903. [Google Scholar] [CrossRef] [PubMed]
- Guemas, V.; Doblas, F.J. Retrospective prediction of the global warming slowdown in the past decade. Nat. Clim. Chang. 2013, 3, 649–653. [Google Scholar] [CrossRef]
- Roberts, C.D.; Palmer, M.D.; Mcneall, D.; Collins, M. Quantifying the likelihood of a continued hiatus in global warming. Nat. Clim. Chang. 2015, 5, 337–342. [Google Scholar] [CrossRef]
- Easterling, D.R.; Wehner, M.F. Is the climate warming or cooling? Geophys. Res. Lett. 2009, 36, 262–275. [Google Scholar] [CrossRef]
- Solomon, S.; Rosenlof, K.H.; Portmann, R.W.; Daniel, J.S.; Davis, S.M.; Sanford, T.J.; Plattner, G.K. Contributions of stratospheric water vapor to decadal changes in the rate of global warming. Science 2010, 327, 1219–1223. [Google Scholar] [CrossRef] [PubMed]
- Kaufmann, R.K. Reconciling anthropogenic climate change with observed temperature 1998–2008. Proc. Natl. Acad. Sci. USA 2011, 108, 11790–11793. [Google Scholar] [CrossRef] [PubMed]
- García-Mozo, H.; Galán, C.; Belmonte, J.; Bermejo, D.; Candau, P.; Guardia, C.D. D.L.; Elvira, B.; Gutiérrez, M.; Jato, V.; Silva, I. Predicting the start and peak dates of the Poaceae pollen season in Spain using process-based models. Agric. For. Meteorol. 2009, 149, 256–262. [Google Scholar] [CrossRef]
- Ye, L.; Xiong, W.; Li, Z.; Yang, P.; Wu, W.; Yang, G.; Fu, Y.; Zou, J.; Chen, Z.; van Ranst, E.; Tang, H. Climate change impact on China food security in 2050. Agron. Sustain. Dev. 2013, 33, 363–374. [Google Scholar] [CrossRef] [Green Version]
- Challinor, A.J.; Ewert, F.; Arnold, S.; Simelton, E.; Fraser, E. Crops and climate change: progress, trends, and challenges in simulating impacts and informing adaptation. J. Exp. Bot. 2009, 60, 2775–2789. [Google Scholar] [CrossRef] [PubMed]
- Xiao, D.; Tao, F.; Liu, Y.; Shi, W.; Wang, M.; Liu, F.; Zhang, S.; Zhu, Z. Observed changes in winter wheat phenology in the North China Plain for 1981–2009. Int. J. Biometeorol. 2013, 57, 275–285. [Google Scholar] [CrossRef] [PubMed]
- Bhandari, S.; Phinn, S.; Gill, T. Assessing viewing and illumination geometry effects on the MODIS vegetation index (MOD13Q1) time series: implications for monitoring phenology and disturbances in forest communities in Queensland, Australia. Int. J. Remote Sens. 2011, 32, 7513–7538. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, J.; Zhang, H.; Zhang, Z.; Guo, X.; Li, X.; Chen, C. Spatial and Temporal Changes in Vegetation Phenology at Middle and High Latitudes of the Northern Hemisphere over the Past Three Decades. Remote Sens. 2015, 7, 10973-10995. https://doi.org/10.3390/rs70810973
Zhao J, Zhang H, Zhang Z, Guo X, Li X, Chen C. Spatial and Temporal Changes in Vegetation Phenology at Middle and High Latitudes of the Northern Hemisphere over the Past Three Decades. Remote Sensing. 2015; 7(8):10973-10995. https://doi.org/10.3390/rs70810973
Chicago/Turabian StyleZhao, Jianjun, Hongyan Zhang, Zhengxiang Zhang, Xiaoyi Guo, Xuedong Li, and Chun Chen. 2015. "Spatial and Temporal Changes in Vegetation Phenology at Middle and High Latitudes of the Northern Hemisphere over the Past Three Decades" Remote Sensing 7, no. 8: 10973-10995. https://doi.org/10.3390/rs70810973
APA StyleZhao, J., Zhang, H., Zhang, Z., Guo, X., Li, X., & Chen, C. (2015). Spatial and Temporal Changes in Vegetation Phenology at Middle and High Latitudes of the Northern Hemisphere over the Past Three Decades. Remote Sensing, 7(8), 10973-10995. https://doi.org/10.3390/rs70810973