Low Sucrose, Omega-3 Enriched Diet Has Region-Specific Effects on Neuroinflammation and Synaptic Function Markers in a Mouse Model of Doxorubicin-Based Chemotherapy

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Treatments
2.2. Experimental Design
2.3. Blood and Tissue Collection
2.4. RNA Preparation and Quantitative Polymerase Chain Reaction
2.5. Cytokine and Chemokine Protein Assays
2.6. Fatty Acid Analysis
2.7. Lipid Peroxidation and Oxidative Stress Assays
2.8. Statistical Analysis
3. Results
3.1. Effects of Omega-3 Fatty Acids and Chemotherapy on Food Intake and Body Weight
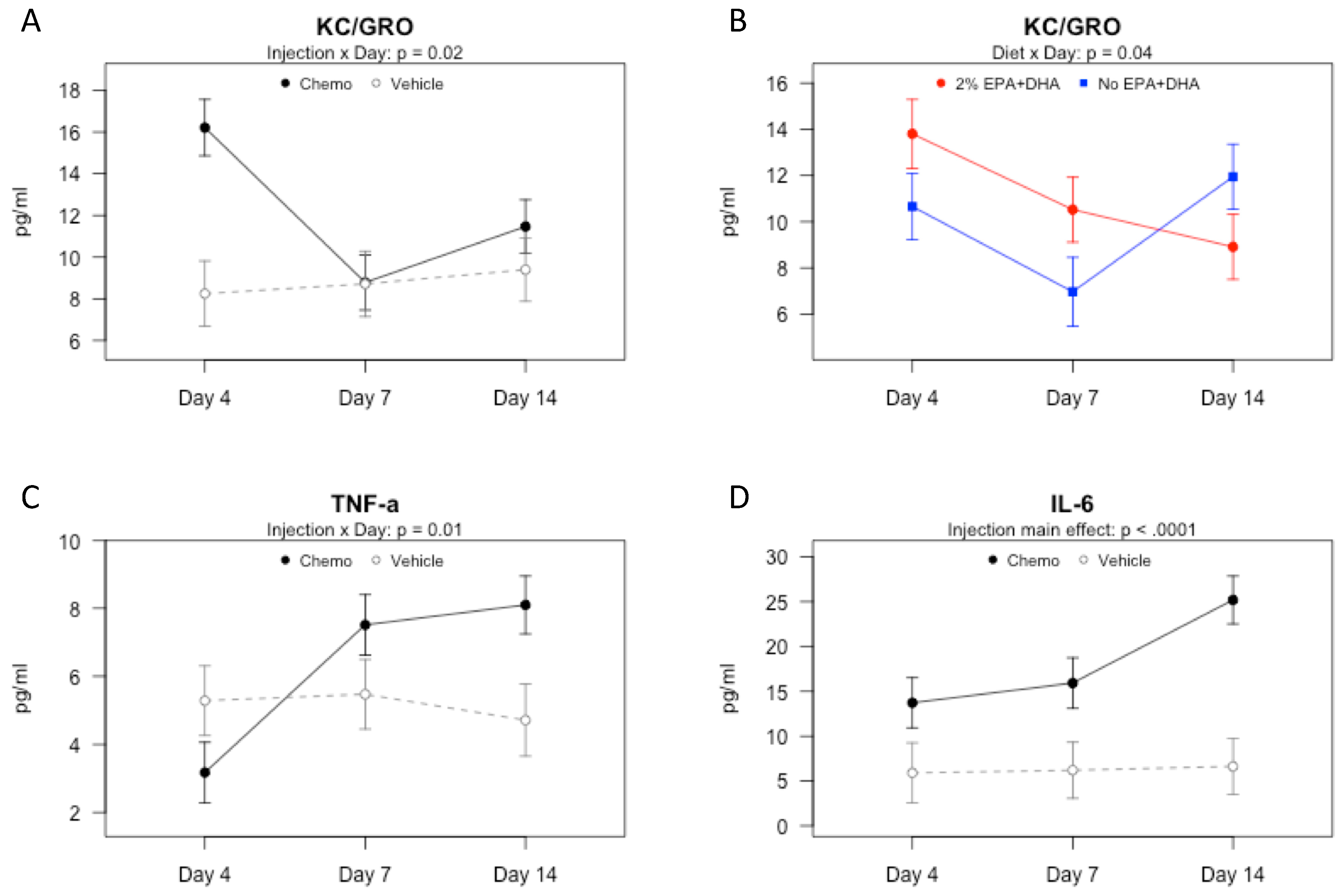
3.2. Effects of Omega-3 Fatty Acids and Chemotherapy on Serum Protein Concentrations of Cytokines and Chemokines over Time
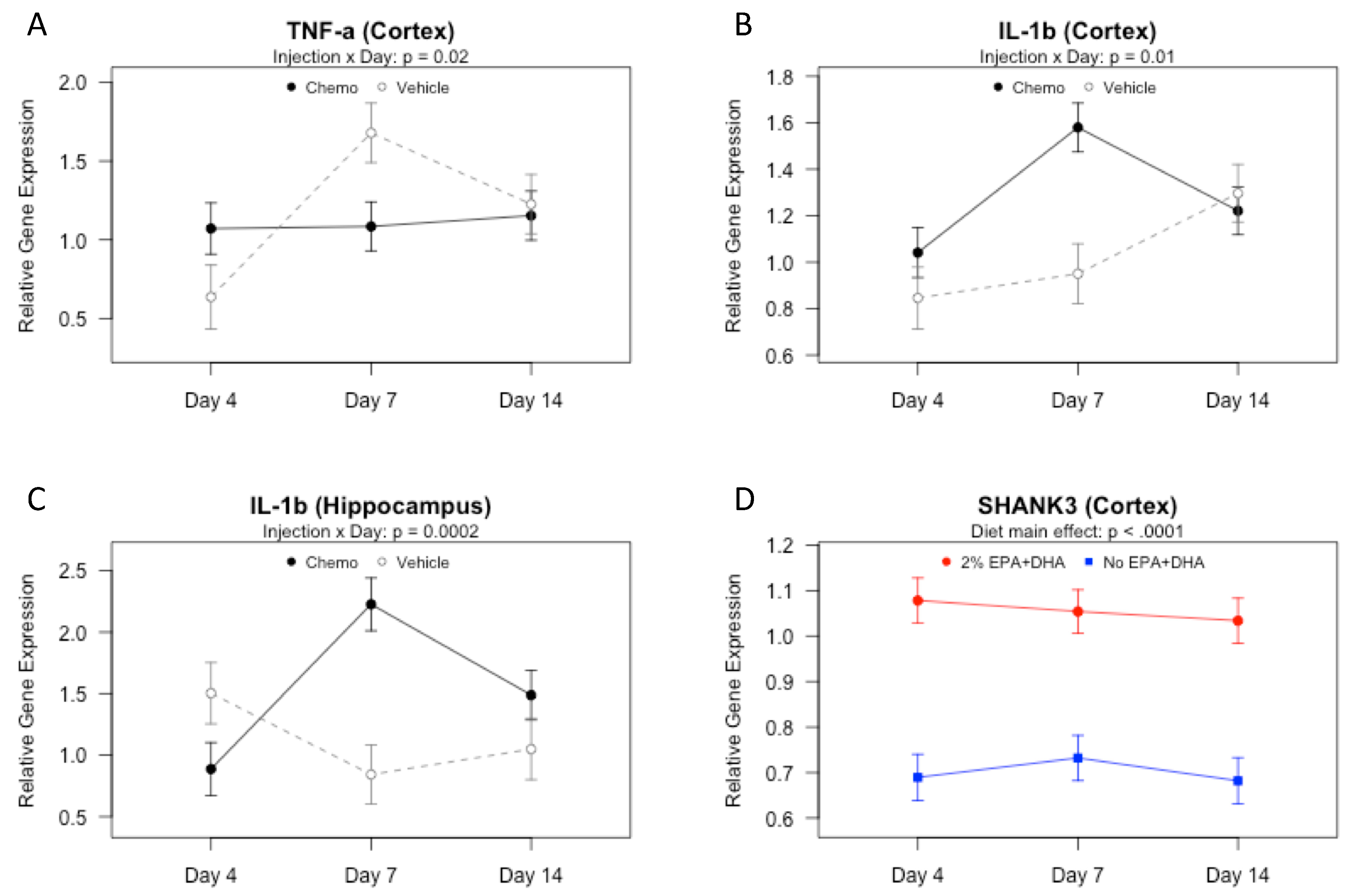
3.3. Effects of Omega-3 Fatty Acids and Chemotherapy on Gene Expression of Cytokines, Chemokines, and Shank in the Brain over Time
3.4. Effects of Sucrose, Omega-3 Fatty Acids, and Chemotherapy on Food Intake and Body Weight
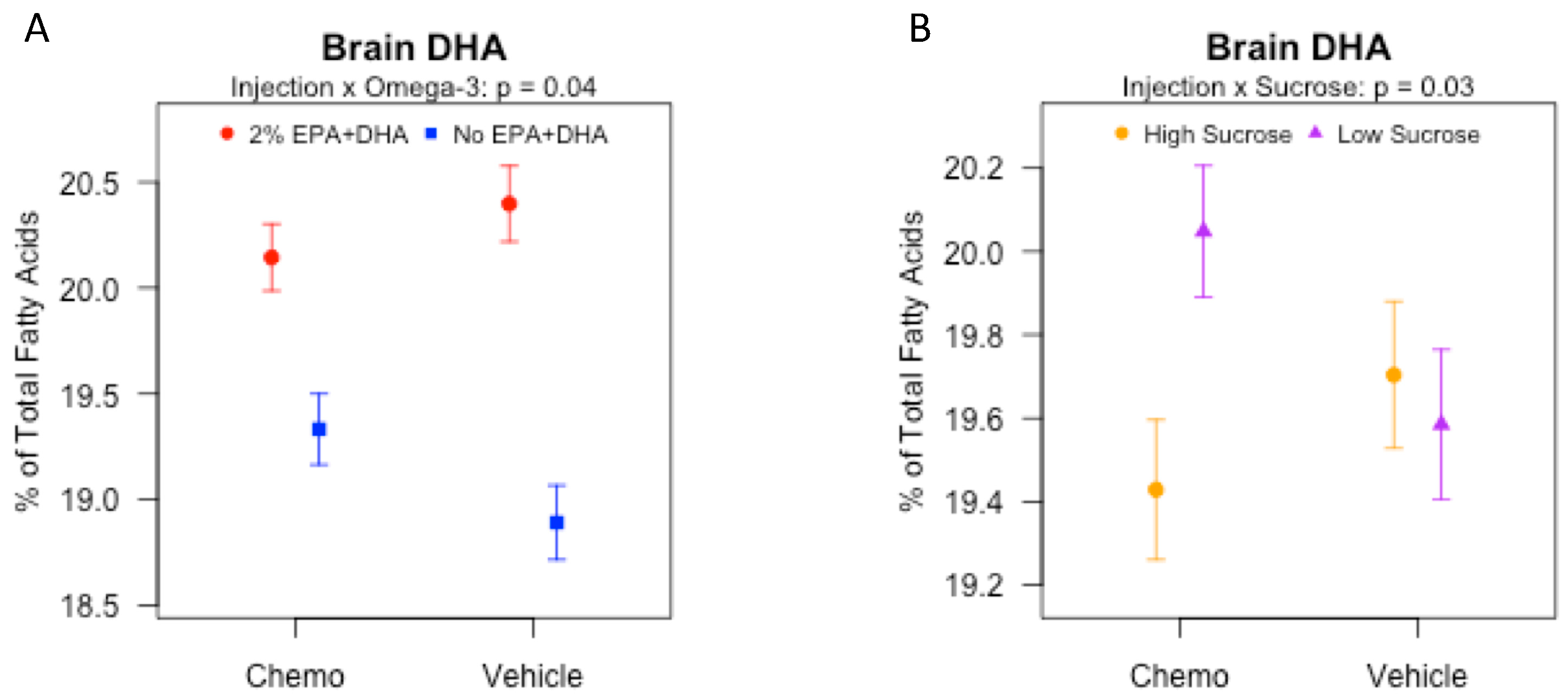
3.5. Effects of Sucrose, Omega-3 Fatty Acids, and Chemotherapy on Brain Fatty Acids
3.6. Effects of Sucrose, Omega-3 Fatty Acids, and Chemotherapy on Serum Protein Concentrations of Cytokines and Chemokines
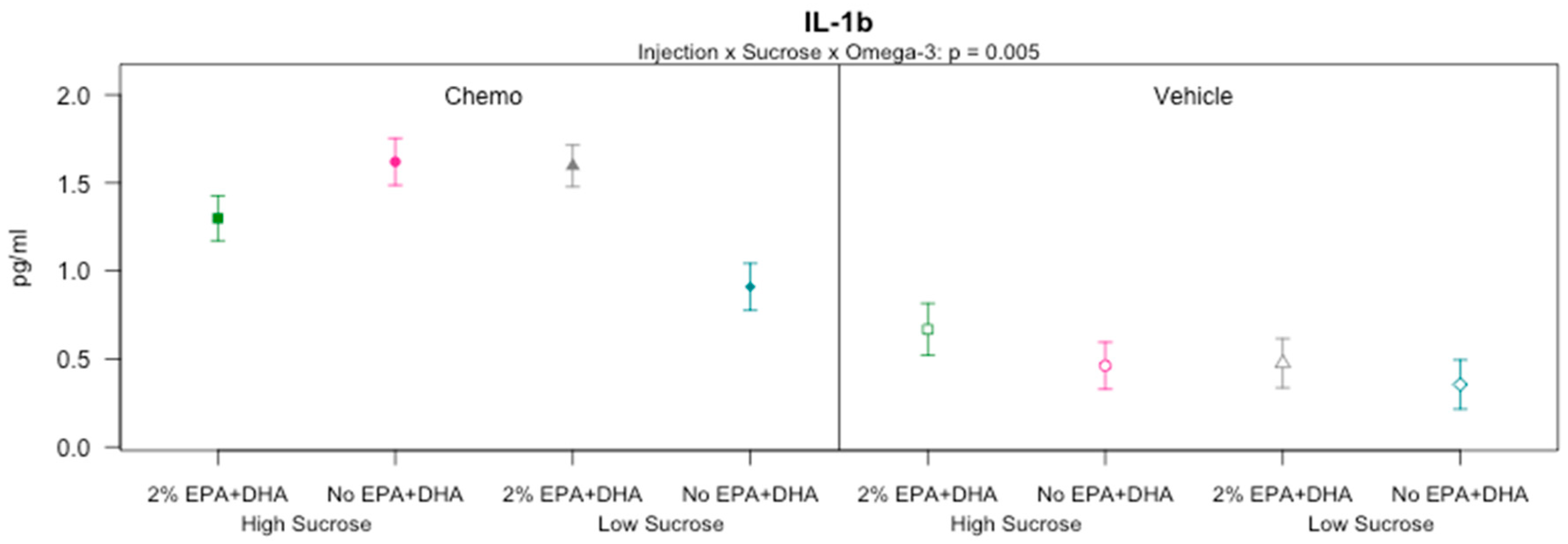
3.7. Effects of Sucrose, Omega-3 Fatty Acids, and Chemotherapy on Protein Concentrations of Cytokines and Chemokines in the Brain
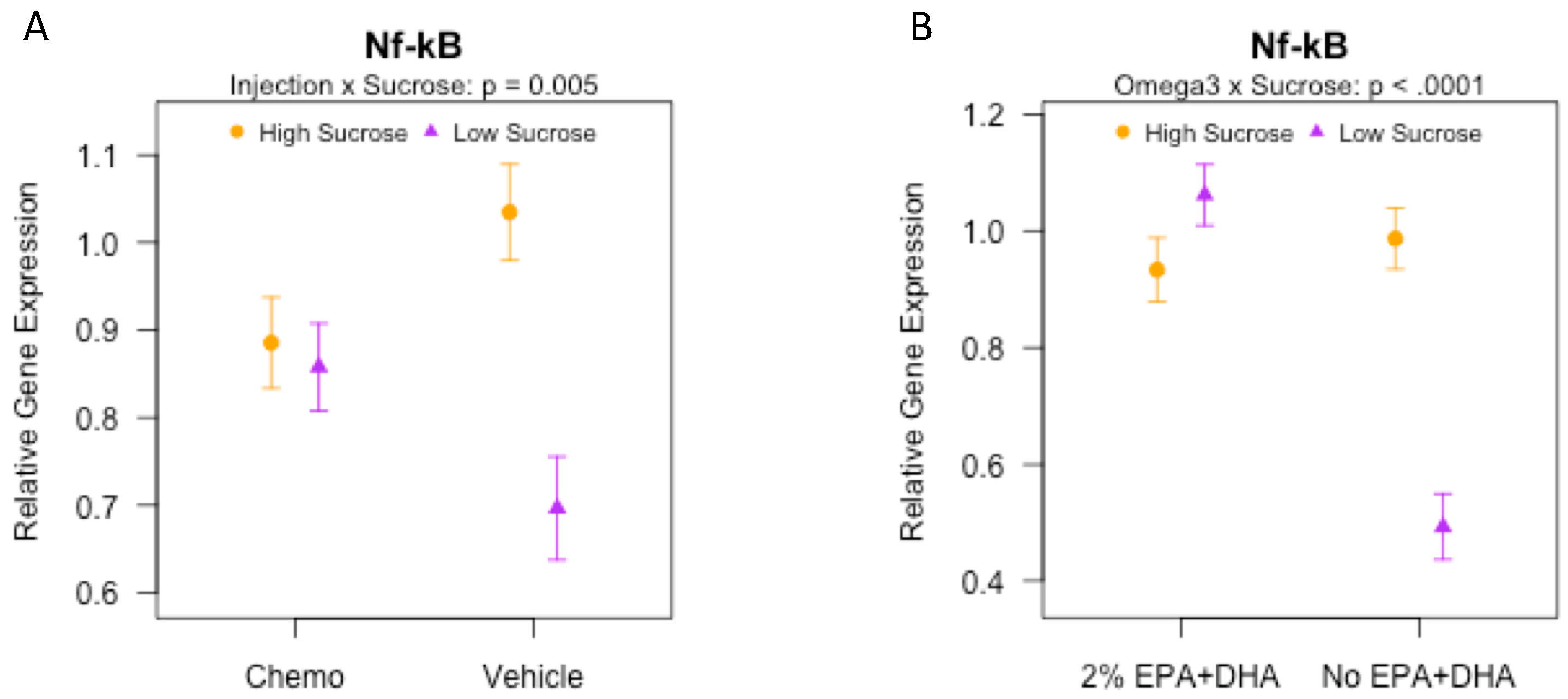
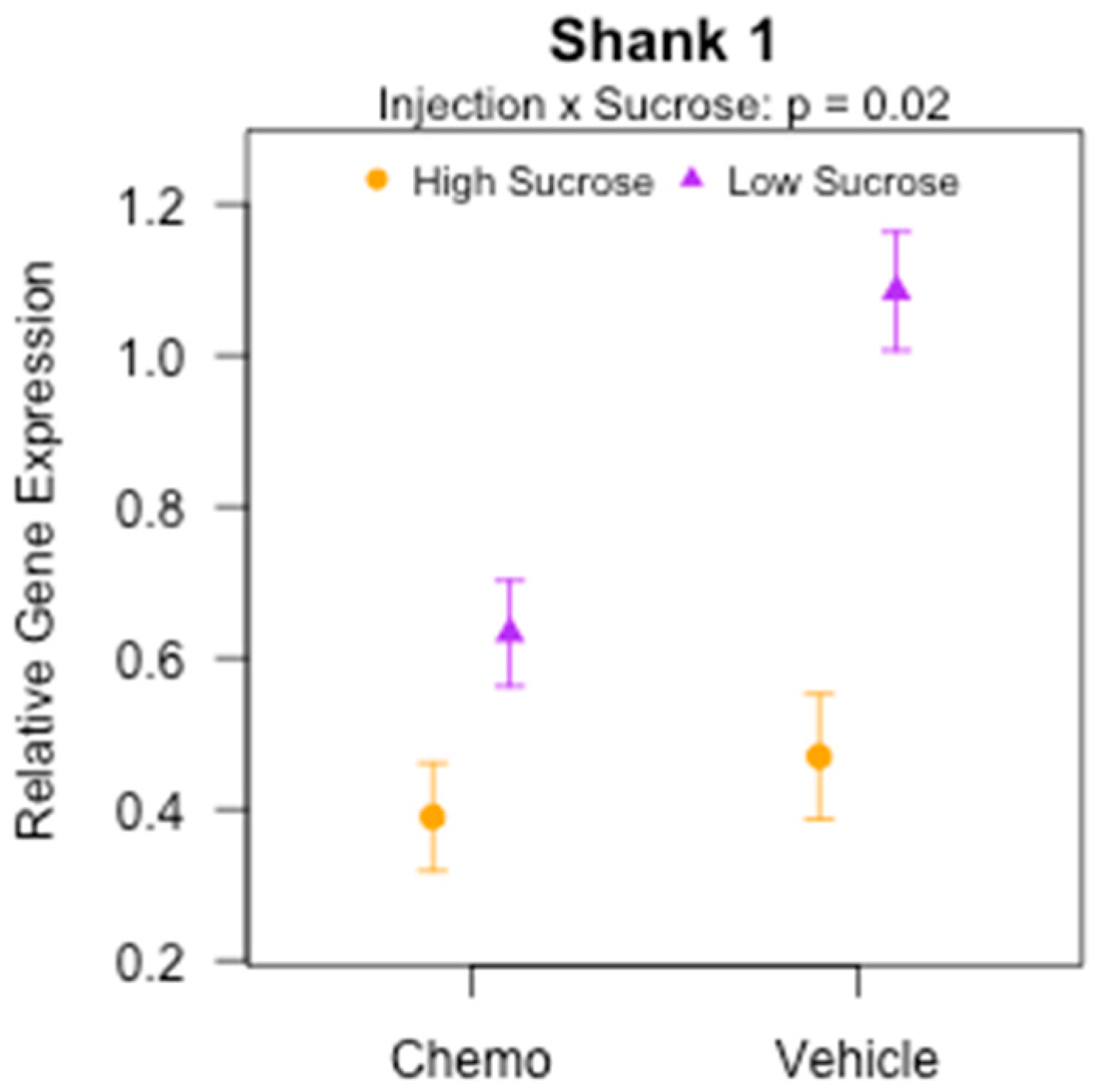
3.8. Effects of Sucrose, Omega-3 Fatty Acids, and Chemotherapy on Gene Expression of Inflammatory and Synaptic Markers in the Brain
3.9. Effects of Sucrose, Omega-3 Fatty Acids, and Chemotherapy on Oxidative Stress, Anti-Oxidant Defense and Lipid Peroxidation Markers in the Brain
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cancer Treatment and Survivorship Facts & Figures 2016-2017; American Cancer Society: Atlanta, GA, USA, 2016.
- Dumas, J.A.; Makarewicz, J.; Schaubhut, G.J.; Devins, R.; Albert, K.; Dittus, K.; Newhouse, P.A. Chemotherapy altered brain functional connectivity in women with breast cancer: A pilot study. Brain Imaging Behav. 2013, 7, 524–532. [Google Scholar] [CrossRef] [PubMed]
- McDonald, B.C.; Conroy, S.K.; Ahles, T.A.; West, J.D.; Saykin, A.J. Gray matter reduction associated with systemic chemotherapy for breast cancer: A prospective MRI study. Breast Cancer Res. Treat. 2010, 123, 819–828. [Google Scholar] [CrossRef] [PubMed]
- Silverman, D.H.; Dy, C.J.; Castellon, S.A.; Lai, J.; Pio, B.S.; Abraham, L.; Waddell, K.; Petersen, L.; Phelps, M.E.; Ganz, P.A. Altered frontocortical, cerebellar, and basal ganglia activity in adjuvant-treated breast cancer survivors 5-10 years after chemotherapy. Breast Cancer Res. Treat. 2007, 103, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Von Ah, D.; Habermann, B.; Carpenter, J.S.; Schneider, B.L. Impact of perceived cognitive impairment in breast cancer survivors. Eur. J. Oncol. Nurs. 2013, 17, 236–241. [Google Scholar] [CrossRef] [PubMed]
- Myers, J.S. Chemotherapy-related cognitive impairment: The breast cancer experience. Oncol. Nurs. Forum 2012, 39, E31–E40. [Google Scholar] [CrossRef] [PubMed]
- Ahles, T.A.; Saykin, A.J.; McDonald, B.C.; Li, Y.; Furstenberg, C.T.; Hanscom, B.S.; Mulrooney, T.J.; Schwartz, G.N.; Kaufman, P.A. Longitudinal assessment of cognitive changes associated with adjuvant treatment for breast cancer: Impact of age and cognitive reserve. J. Clin. Oncol. 2010, 28, 4434–4440. [Google Scholar] [CrossRef] [PubMed]
- Vichaya, E.G.; Chiu, G.S.; Krukowski, K.; Lacourt, T.E.; Kavelaars, A.; Dantzer, R.; Heijnen, C.J.; Walker, A.K. Mechanisms of chemotherapy-induced behavioral toxicities. Front. Neurosci. 2015, 9, 131. [Google Scholar] [CrossRef] [PubMed]
- Borniger, J.C.; Gaudier-Diaz, M.M.; Zhang, N.; Nelson, R.J.; DeVries, A.C. Cytotoxic chemotherapy increases sleep and sleep fragmentation in non-tumor-bearing mice. Brain Behav. Immun. 2015, 47, 218–227. [Google Scholar] [CrossRef] [PubMed]
- Bower, J.E.; Ganz, P.A.; Irwin, M.R.; Kwan, L.; Breen, E.C.; Cole, S.W. Inflammation and behavioral symptoms after breast cancer treatment: Do fatigue, depression, and sleep disturbance share a common underlying mechanism? J. Clin. Oncol. 2011, 29, 3517–3522. [Google Scholar] [CrossRef] [PubMed]
- Joshi, G.; Sultana, R.; Tangpong, J.; Cole, M.P.; St Clair, D.K.; Vore, M.; Estus, S.; Butterfield, D.A. Free radical mediated oxidative stress and toxic side effects in brain induced by the anti cancer drug adriamycin: Insight into chemobrain. Free Radic. Res. 2005, 39, 1147–1154. [Google Scholar] [CrossRef] [PubMed]
- Tangpong, J.; Cole, M.P.; Sultana, R.; Joshi, G.; Estus, S.; Vore, M.; St Clair, W.; Ratanachaiyavong, S.; St Clair, D.K.; Butterfield, D.A. Adriamycin-induced, TNF-alpha-mediated central nervous system toxicity. Neurobiol. Dis. 2006, 23, 127–139. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.H.; Ancoli-Israel, S.; Bower, J.E.; Capuron, L.; Irwin, M.R. Neuroendocrine-immune mechanisms of behavioral comorbidities in patients with cancer. J. Clin. Oncol. 2008, 26, 971–982. [Google Scholar] [CrossRef] [PubMed]
- Cheung, Y.T.; Ng, T.; Shwe, M.; Ho, H.K.; Foo, K.M.; Cham, M.T.; Lee, J.A.; Fan, G.; Tan, Y.P.; Yong, W.S.; et al. Association of proinflammatory cytokines and chemotherapy-associated cognitive impairment in breast cancer patients: A multi-centered, prospective, cohort study. Ann. Oncol. 2015, 26, 1446–1451. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. Marine omega-3 fatty acids and inflammatory processes: Effects, mechanisms and clinical relevance. Biochim. Biophys. Acta 2015, 1851, 469–484. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.Q.; Dang, R.L.; Tang, M.M.; Cai, H.L.; Li, H.D.; Liao, D.H.; He, X.; Cao, L.J.; Xue, Y.; Jiang, P. Long Chain Omega-3 Polyunsaturated Fatty Acid Supplementation Alleviates Doxorubicin-Induced Depressive-Like Behaviors and Neurotoxicity in Rats: Involvement of Oxidative Stress and Neuroinflammation. Nutrients 2016, 8, 243. [Google Scholar] [CrossRef] [PubMed]
- Finocchiaro, C.; Segre, O.; Fadda, M.; Monge, T.; Scigliano, M.; Schena, M.; Tinivella, M.; Tiozzo, E.; Catalano, M.G.; Pugliese, M.; et al. Effect of n-3 fatty acids on patients with advanced lung cancer: A double-blind, placebo-controlled study. Br. J. Nutr. 2012, 108, 327–333. [Google Scholar] [CrossRef] [PubMed]
- Orchard, T.S.; Andridge, R.R.; Yee, L.D.; Lustberg, M.B. Diet Quality, Inflammation, and Quality of Life in Breast Cancer Survivors: A Cross-Sectional Analysis of Pilot Study Data. J. Acad. Nutr. Diet. 2017. [Google Scholar] [CrossRef] [PubMed]
- Bougnoux, P.; Hajjaji, N.; Ferrasson, M.N.; Giraudeau, B.; Couet, C.; Le Floch, O. Improving outcome of chemotherapy of metastatic breast cancer by docosahexaenoic acid: A phase II trial. Br. J. Cancer 2009, 101, 1978–1985. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, R.; Gomez-Pinilla, F. ‘Metabolic syndrome’ in the brain: Deficiency in omega-3 fatty acid exacerbates dysfunctions in insulin receptor signalling and cognition. J. Physiol. 2012, 590, 2485–2499. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, L.B.; Raben, A.; Stender, S.; Astrup, A. Effect of sucrose on inflammatory markers in overweight humans. Am. J. Clin. Nutr. 2005, 82, 421–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, T.; Liaset, B.; Hao, Q.; Petersen, R.K.; Fjaere, E.; Ngo, H.T.; Lillefosse, H.H.; Ringholm, S.; Sonne, S.B.; Treebak, J.T.; et al. Sucrose counteracts the anti-inflammatory effect of fish oil in adipose tissue and increases obesity development in mice. PLoS ONE 2011, 6, e21647. [Google Scholar] [CrossRef] [PubMed]
- Taha, A.Y.; Gao, F.; Ramadan, E.; Cheon, Y.; Rapoport, S.I.; Kim, H.W. Upregulated expression of brain enzymatic markers of arachidonic and docosahexaenoic acid metabolism in a rat model of the metabolic syndrome. BMC Neurosci. 2012, 13, 131. [Google Scholar] [CrossRef] [PubMed]
- Reagan-Shaw, S.; Nihal, M.; Ahmad, N. Dose translation from animal to human studies revisited. FASEB J. 2008, 22, 659–661. [Google Scholar] [CrossRef] [PubMed]
- Folch, J.; Lees, M.; Stanley, G.H.S. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [PubMed]
- Stoffel, W.; Chu, F.; Ahrens, E. Analysis of long-chain fatty acids by gas-liquid chromatography. Anal. Chem. 1959, 31, 307–308. [Google Scholar] [CrossRef]
- Belury, M.A.; Cole, R.M.; Bailey, B.E.; Ke, J.Y.; Andridge, R.R.; Kiecolt-Glaser, J.K. Erythrocyte linoleic acid, but not oleic acid, is associated with improvements in body composition in men and women. Mol. Nutr. Food Res. 2016, 60, 1206–1212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andreasen, A.S.; Krabbe, K.S.; Krogh-Madsen, R.; Taudorf, S.; Pedersen, B.K.; Moller, K. Human endotoxemia as a model of systemic inflammation. Curr. Med. Chem. 2008, 15, 1697–1705. [Google Scholar] [CrossRef] [PubMed]
- Stolina, M.; Bolon, B.; Dwyer, D.; Middleton, S.; Duryea, D.; Kostenuik, P.J.; Feige, U.; Zack, D.J. The evolving systemic and local biomarker milieu at different stages of disease progression in rat collagen-induced arthritis. Biomarkers 2008, 13, 692–712. [Google Scholar] [CrossRef] [PubMed]
- Rittner, H.L.; Mousa, S.A.; Labuz, D.; Beschmann, K.; Schafer, M.; Stein, C.; Brack, A. Selective local PMN recruitment by CXCL1 or CXCL2/3 injection does not cause inflammatory pain. J. Leukoc. Biol. 2006, 79, 1022–1032. [Google Scholar] [CrossRef] [PubMed]
- Paz, M.; Gomes, A.L.J.; Islam, M.T.; Tabrez, S.; Jabir, N.R.; Alam, M.Z.; Machado, K.C.; de Alencar, M.; Ali, E.S.; Mishra, S.K.; et al. Assessment of chemotherapy on various biochemical markers in breast cancer patients. J. Cell. Biochem. 2018, 119, 2923–2928. [Google Scholar] [CrossRef] [PubMed]
- Hanada, T.; Yoshimura, A. Regulation of cytokine signaling and inflammation. Cytokine Growth Factor Rev. 2002, 13, 413–421. [Google Scholar] [CrossRef]
- Monteiro, P.; Feng, G. SHANK proteins: Roles at the synapse and in autism spectrum disorder. Nat. Rev. Neurosci. 2017, 18, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Sala, C.; Vicidomini, C.; Bigi, I.; Mossa, A.; Verpelli, C. Shank synaptic scaffold proteins: Keys to understanding the pathogenesis of autism and other synaptic disorders. J. Neurochem. 2015, 135, 849–858. [Google Scholar] [CrossRef] [PubMed]
- Peca, J.; Feliciano, C.; Ting, J.T.; Wang, W.; Wells, M.F.; Venkatraman, T.N.; Lascola, C.D.; Fu, Z.; Feng, G. Shank3 mutant mice display autistic-like behaviours and striatal dysfunction. Nature 2011, 472, 437–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, K.; Holder, J.L., Jr.; Schaaf, C.P.; Lu, H.; Chen, H.; Kang, H.; Tang, J.; Wu, Z.; Hao, S.; Cheung, S.W.; et al. SHANK3 overexpression causes manic-like behaviour with unique pharmacogenetic properties. Nature 2013, 503, 72–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bazinet, R.P.; Laye, S. Polyunsaturated fatty acids and their metabolites in brain function and disease. Nat. Rev. Neurosci. 2014, 15, 771–785. [Google Scholar] [CrossRef] [PubMed]
- Ismail, H.M.; Dorchies, O.M.; Perozzo, R.; Strosova, M.K.; Scapozza, L.; Ruegg, U.T. Inhibition of iPLA2 beta and of stretch-activated channels by doxorubicin alters dystrophic muscle function. Br. J. Pharmacol. 2013, 169, 1537–1550. [Google Scholar] [CrossRef] [PubMed]
- Grataroli, R.; Leonardi, J.; Chautan, M.; Lafont, H.; Nalbone, G. Effect of anthracyclines on phospholipase A2 activity and prostaglandin E2 production in rat gastric mucosa. Biochem. Pharmacol. 1993, 46, 349–355. [Google Scholar] [CrossRef]
- Swift, L.; McHowat, J.; Sarvazyan, N. Inhibition of membrane-associated calcium-independent phospholipase A2 as a potential culprit of anthracycline cardiotoxicity. Cancer Res. 2003, 63, 5992–5998. [Google Scholar] [PubMed]
- Janelsins, M.C.; Mustian, K.M.; Palesh, O.G.; Mohile, S.G.; Peppone, L.J.; Sprod, L.K.; Heckler, C.E.; Roscoe, J.A.; Katz, A.W.; Williams, J.P.; et al. Differential expression of cytokines in breast cancer patients receiving different chemotherapies: Implications for cognitive impairment research. Support Care Cancer 2012, 20, 831–839. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Li, L.; Cui, B.; Gai, Z.; Li, Q.; Wang, S.; Yan, J.; Lin, B.; Tian, L.; Liu, H.; et al. Early postnatal exposure to airborne fine particulate matter induces autism-like phenotypes in male rats. Toxicol. Sci. 2017. [Google Scholar] [CrossRef] [PubMed]
- Beilharz, J.E.; Maniam, J.; Morris, M.J. Short-term exposure to a diet high in fat and sugar, or liquid sugar, selectively impairs hippocampal-dependent memory, with differential impacts on inflammation. Behav. Brain Res. 2016, 306, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Grabert, K.; Michoel, T.; Karavolos, M.H.; Clohisey, S.; Baillie, J.K.; Stevens, M.P.; Freeman, T.C.; Summers, K.M.; McColl, B.W. Microglial brain region-dependent diversity and selective regional sensitivities to aging. Nat. Neurosci. 2016, 19, 504–516. [Google Scholar] [CrossRef] [PubMed]
- McNamara, R.K.; Able, J.; Jandacek, R.; Rider, T.; Tso, P. Inbred C57BL/6J and DBA/2J mouse strains exhibit constitutive differences in regional brain fatty acid composition. Lipids 2009, 44, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Etherton, T.D.; Martin, K.R.; Gillies, P.J.; West, S.G.; Kris-Etherton, P.M. Dietary alpha-linolenic acid inhibits proinflammatory cytokine production by peripheral blood mononuclear cells in hypercholesterolemic subjects. Am. J. Clin. Nutr. 2007, 85, 385–391. [Google Scholar] [CrossRef] [PubMed]
- Rapoport, S.I.; Igarashi, M. Can the rat liver maintain normal brain DHA metabolism in the absence of dietary DHA? Prostaglandins Leukot. Essent. Fatty Acids 2009, 81, 119–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lukiw, W.J.; Cui, J.G.; Marcheselli, V.L.; Bodker, M.; Botkjaer, A.; Gotlinger, K.; Serhan, C.N.; Bazan, N.G. A role for docosahexaenoic acid-derived neuroprotectin D1 in neural cell survival and Alzheimer disease. J. Clin. Investig. 2005, 115, 2774–2783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Zhu, M.; Hjorth, E.; Cortes-Toro, V.; Eyjolfsdottir, H.; Graff, C.; Nennesmo, I.; Palmblad, J.; Eriksdotter, M.; Sambamurti, K.; et al. Resolution of inflammation is altered in Alzheimer’s disease. Alzheimer’s Dement. 2015, 11, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Lustberg, M.B.; Orchard, T.S.; Reinbolt, R.; Andridge, R.; Pan, X.; Belury, M.; Cole, R.; Logan, A.; Layman, R.; Ramaswamy, B. Randomized placebo-controlled pilot trial of omega 3 fatty acids for prevention of aromatase inhibitor-induced musculoskeletal pain. Breast Cancer Res. Treat. 2018, 167, 709–718. [Google Scholar] [CrossRef] [PubMed]
- Yee, L.D.; Lester, J.L.; Cole, R.M.; Richardson, J.R.; Hsu, J.C.; Li, Y.; Lehman, A.; Belury, M.A.; Clinton, S.K. Omega-3 Fatty acid supplements in women at high risk of breast cancer have dose-dependent effects on breast adipose tissue fatty acid composition. Am. J. Clin. Nutr. 2010, 91, 1185–1194. [Google Scholar] [CrossRef] [PubMed]
- Lalancette-Hebert, M.; Julien, C.; Cordeau, P.; Bohacek, I.; Weng, Y.C.; Calon, F.; Kriz, J. Accumulation of dietary docosahexaenoic acid in the brain attenuates acute immune response and development of postischemic neuronal damage. Stroke 2011, 42, 2903–2909. [Google Scholar] [CrossRef] [PubMed]
- Arsenault, D.; Julien, C.; Tremblay, C.; Calon, F. DHA improves cognition and prevents dysfunction of entorhinal cortex neurons in 3xTg-AD mice. PLoS ONE 2011, 6, e17397. [Google Scholar] [CrossRef] [PubMed]
- Yee, L.D.; Young, D.C.; Rosol, T.J.; Vanbuskirk, A.M.; Clinton, S.K. Dietary (n-3) polyunsaturated fatty acids inhibit HER-2/neu-induced breast cancer in mice independently of the PPARgamma ligand rosiglitazone. J. Nutr. 2005, 135, 983–988. [Google Scholar] [CrossRef] [PubMed]
- U.S. Department of Agriculture and U.S. Department of Health and Human Services. Dietary Guidelines for Americans, 7th ed.; U.S. Government Printing Office: Washington, DC, USA, 2010.
- Institute of Medicine. Dietary Reference Intakes for Energy, Carbohydrate, Fiber, Fat, Fatty Acids, Cholesterol, Protein, and Amino Acids; The National Academies Press: Washington, DC, USA, 2005. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemotherapy | Vehicle | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2% EPA + DHA | No EPA + DHA | 2% EPA + DHA | No EPA + DHA | 2% EPA + DHA | No EPA + DHA | 2% EPA + DHA | No EPA + DHA | |||||||||
| Low Sucrose | Low Sucrose | High Sucrose | High Sucrose | Low Sucrose | Low Sucrose | High Sucrose | High Sucrose | |||||||||
| Serum | ||||||||||||||||
| IFN-γ # | 0.78 | (0.33) | 0.73 | 0.44 | 0.62 | 0.18 | 0.60 | 0.27 | 0.44 | 0.16 | 0.43 | 0.24 | 0.54 | 0.26 | 0.41 | 0.30 |
| IL-10 | 9.0 | (4.5) | 7.7 | (1.7) | 8.1 | (2.8) | 9.0 | (2.7) | 7.7 | (1.2) | 9.0 | (3.5) | 7.7 | (1.8) | 7.3 | (3.2) |
| IL-1b ^ | 1.6 | (0.56) | 0.91 | (0.62) | 1.3 | (0.50) | 1.6 | (0.52) | 0.48 | (0.24) | 0.36 | (0.14) | 0.67 | (0.21) | 0.46 | (0.33) |
| IL-2 | 1.3 | (1.6) | 1.5 | (0.88) | 1.1 | (0.55) | 1.0 | (0.38) | 0.60 | (0.38) | 0.87 | (0.50) | 0.52 | (0.24) | 0.73 | (0.26) |
| IL-5 # | 9.1 | (6.1) | 9.5 | (6.9) | 8.5 | (4.9) | 10.1 | (4.4) | 2.7 | (1.1) | 2.7 | (1.4) | 1.9 | (0.57) | 2.9 | (1.2) |
| IL-6 # | 16.7 | (7.4) | 21.1 | (16) | 16.7 | (7.1) | 19.2 | (5.5) | 4.0 | (2.6) | 4.5 | (2.9) | 3.6 | (2.0) | 2.1 | (2.0) |
| KC/GRO | 73.6 | (26) | 85.6 | (34) | 84.6 | (20) | 90.5 | (21) | 94.0 | (19) | 94.1 | (23) | 100.8 | (12) | 95.6 | (16) |
| TNF-a # | 10.3 | (2.2) | 11.8 | (5.5) | 11.4 | (3.6) | 11.1 | (2.2) | 7.2 | (2.4) | 7.5 | (2.5) | 8.5 | (2.1) | 9.4 | (2.5) |
| Hippocampus | ||||||||||||||||
| IL-10 | 1.5 | (0.55) | 1.5 | (0.59) | 0.98 | (0.51) | 1.3 | (0.75) | 0.86 | (0.53) | 1.09 | (0.76) | 1.1 | (0.57) | 0.74 | (0.85) |
| IL-5 #,Ω | 1.2 | (0.37) | 1.2 | (0.43) | 1.1 | (0.39) | 0.98 | (0.32) | 1.0 | (0.33) | 0.74 | (0.18) | 1.1 | (0.42) | 0.91 | (0.23) |
| IL-6 #,Ω | 19.0 | (4.2) | 18.7 | (5) | 18.5 | (5.5) | 16.1 | (3.8) | 17.4 | (4.1) | 12.9 | (2.5) | 17.2 | (5.1) | 15.1 | (3.0) |
| Cortex | ||||||||||||||||
| IFN-γ | 0.088 | (0.052) | 0.161 | (0.086) | 0.100 | (0.094) | 0.108 | (0.061) | 0.132 | (0.099) | 0.088 | (0.049) | 0.115 | (0.068) | 0.073 | (0.031) |
| IL-10 | 1.3 | (0.67) | 1.7 | (0.88) | 1.1 | (0.72) | 1.4 | (0.45) | 1.2 | (0.35) | 1.2 | (0.55) | 1.3 | (0.65) | 1.1 | (0.58) |
| IL-12 | 13 | (7.0) | 18 | (9.9) | 12 | (4.9) | 15 | (8.0) | 13 | (9.2) | 15 | (4.0) | 14 | (4.9) | 15 | (3.5) |
| IL-1b | 0.21 | (0.13) | 0.28 | (0.15) | 0.43 | (0.55) | 0.23 | (0.07) | 0.28 | (0.35) | 0.29 | (0.13) | 0.26 | (0.12) | 0.40 | (0.25) |
| IL-2 | 0.83 | (0.33) | 0.98 | (0.28) | 1.22 | (1.49) | 0.85 | (0.26) | 1.05 | (0.49) | 1.12 | (0.41) | 0.87 | (0.30) | 0.84 | (0.18) |
| IL-4 | 0.31 | (0.18) | 0.55 | (0.20) | 0.43 | (0.16) | 0.44 | (0.19) | 0.37 | (0.22) | 0.42 | (0.17) | 0.46 | (0.14) | 0.51 | (0.15) |
| IL-5 Ω | 1.2 | (0.23) | 1.4 | (0.27) | 1.1 | (0.40) | 1.3 | (0.28) | 1.1 | (0.34) | 1.4 | (0.23) | 1.3 | (0.30) | 1.5 | (0.29) |
| IL-6 Ω | 17 | (3.3) | 20 | (3.7) | 17 | (5.6) | 18 | (4.0) | 16 | (4.1) | 20 | (3.0) | 18 | (3.3) | 21 | (4.1) |
| KC/GRO Ω | 0.39 | (0.28) | 0.80 | (0.38) | 0.41 | (0.25) | 0.57 | (0.26) | 0.46 | (0.34) | 0.52 | (0.25) | 0.40 | (0.19) | 0.43 | (0.18) |
| TNF-a | 0.32 | (0.17) | 0.45 | (0.22) | 0.26 | (0.14) | 0.28 | (0.09) | 0.31 | (0.22) | 0.35 | (0.14) | 0.31 | (0.18) | 0.37 | (0.12) |
| Chemo | Vehicle | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2% EPA + DHA | No EPA + DHA | 2% EPA + DHA | No EPA + DHA | 2% EPA + DHA | No EPA + DHA | 2% EPA + DHA | No EPA + DHA | |||||||||
| Low Sucrose | Low Sucrose | High Sucrose | High Sucrose | Low Sucrose | Low Sucrose | High Sucrose | High Sucrose | |||||||||
| Hippocampus | ||||||||||||||||
| TNF-a | 2.0 | (1.4) | 1.3 | (0.82) | 0.82 | (0.6) | 2.0 | (1.7) | 1.2 | (1.3) | 0.65 | (0.51) | 1.3 | (1.3) | 1.2 | (0.93) |
| IL-1b | 2.4 | (1.9) | 1.0 | (1.0) | 0.66 | (0.4) | 2.5 | (3.1) | 1.1 | (0.85) | 0.4 | (0.31) | 2.1 | (2.9) | 1.8 | (1.8) |
| IL-6 | 1.0 | (0.53) | 1.12 | (0.45) | 1.0 | (0.47) | 1.4 | (0.90) | 1.22 | (0.83) | 1.11 | (0.60) | 0.99 | (0.47) | 0.99 | (0.36) |
| Nfk-b π,ω | 1.1 | (0.31) | 0.57 | (0.14) | 0.82 | (0.18) | 0.95 | (0.25) | 0.98 | (0.37) | 0.41 | (0.10) | 1.05 | (0.31) | 1.02 | (0.21) |
| Shank 3 | 1.0 | (0.72) | 1.0 | 0.72 | 0.73 | (0.47) | 0.84 | (0.46) | 1.2 | (0.66) | 1.1 | (0.58) | 1.2 | (0.76) | 1.2 | (0.65) |
| Cortex | ||||||||||||||||
| TNF-a | 1.1 | (0.49) | 0.66 | (0.36) | 1.2 | (0.49) | 0.73 | (0.33) | 1.0 | (0.58) | 0.88 | (0.34) | 1.1 | (0.56) | 1.1 | (0.42) |
| IL-1b | 1.3 | (0.66) | 0.89 | (0.30) | 1.4 | (0.49) | 1.3 | (0.77) | 1.1 | (0.55) | 1.1 | (0.39) | 1.4 | (0.85) | 1.1 | (0.36) |
| IL-6 | 1.1 | (0.59) | 0.82 | (0.43) | 1.4 | (0.66) | 1.1 | (0.48) | 1.2 | (0.69) | 1.1 | (0.28) | 1.0 | (0.36) | 1.1 | (0.32) |
| Nfk-b | 0.93 | (0.25) | 0.91 | (0.30) | 1.1 | (0.27) | 0.95 | (0.16) | 1.0 | (0.26) | 1.0 | (0.32) | 1.0 | (0.20) | 1.0 | (0.19) |
| Shank 3 #,Ω | 1.0 | (0.20) | 0.82 | (0.20) | 1.0 | (0.24) | 0.87 | (0.12) | 1.1 | (0.19) | 1.0 | (0.20) | 1.0 | (0.20) | 0.98 | (0.16) |
| Shank 1 π | 0.71 | (0.27) | 0.55 | (0.20) | 0.49 | (0.17) | 0.29 | (0.15) | 0.98 | (0.55) | 1.2 | (0.61) | 0.61 | (0.36) | 0.33 | (0.16) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Orchard, T.S.; Gaudier-Diaz, M.M.; Phuwamongkolwiwat-Chu, P.; Andridge, R.; Lustberg, M.B.; Bomser, J.; Cole, R.M.; Belury, M.A.; DeVries, A.C. Low Sucrose, Omega-3 Enriched Diet Has Region-Specific Effects on Neuroinflammation and Synaptic Function Markers in a Mouse Model of Doxorubicin-Based Chemotherapy. Nutrients 2018, 10, 2004. https://doi.org/10.3390/nu10122004
Orchard TS, Gaudier-Diaz MM, Phuwamongkolwiwat-Chu P, Andridge R, Lustberg MB, Bomser J, Cole RM, Belury MA, DeVries AC. Low Sucrose, Omega-3 Enriched Diet Has Region-Specific Effects on Neuroinflammation and Synaptic Function Markers in a Mouse Model of Doxorubicin-Based Chemotherapy. Nutrients. 2018; 10(12):2004. https://doi.org/10.3390/nu10122004
Chicago/Turabian StyleOrchard, Tonya S., Monica M. Gaudier-Diaz, Panchita Phuwamongkolwiwat-Chu, Rebecca Andridge, Maryam B. Lustberg, Joshua Bomser, Rachel M. Cole, Martha A. Belury, and A. Courtney DeVries. 2018. "Low Sucrose, Omega-3 Enriched Diet Has Region-Specific Effects on Neuroinflammation and Synaptic Function Markers in a Mouse Model of Doxorubicin-Based Chemotherapy" Nutrients 10, no. 12: 2004. https://doi.org/10.3390/nu10122004
APA StyleOrchard, T. S., Gaudier-Diaz, M. M., Phuwamongkolwiwat-Chu, P., Andridge, R., Lustberg, M. B., Bomser, J., Cole, R. M., Belury, M. A., & DeVries, A. C. (2018). Low Sucrose, Omega-3 Enriched Diet Has Region-Specific Effects on Neuroinflammation and Synaptic Function Markers in a Mouse Model of Doxorubicin-Based Chemotherapy. Nutrients, 10(12), 2004. https://doi.org/10.3390/nu10122004