Moderate Alcohol Consumption Inhibits Sodium-Dependent Glutamine Co-Transport in Rat Intestinal Epithelial Cells in Vitro and Ex Vivo

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Studies
2.2. Cell Culture
2.2.1. Calphostin C Treatments
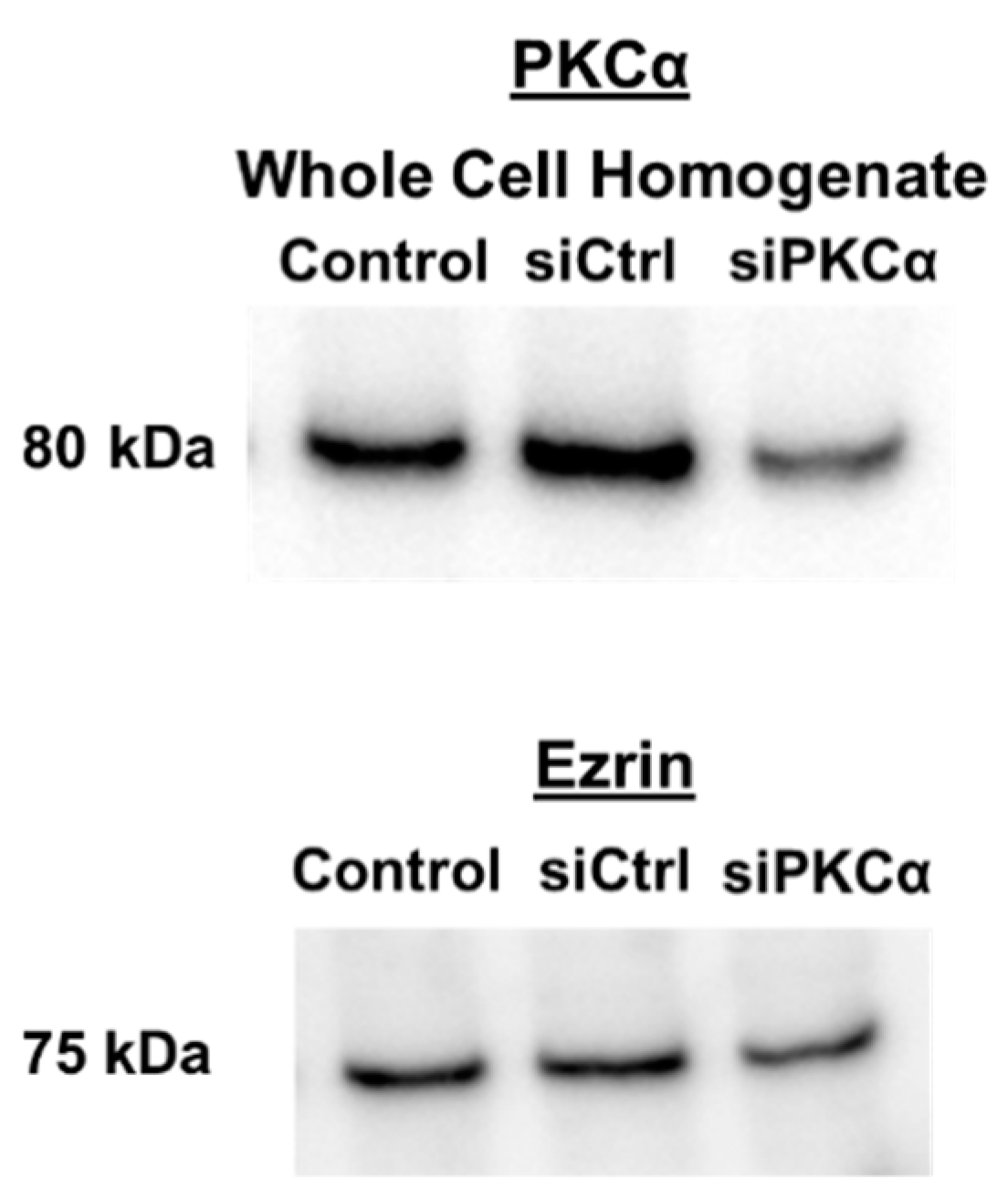
2.2.2. siRNA Transfections
2.3. Na-K-ATPase Measurement
2.4. Villus Cell Isolation
2.5. Protein Quantification
2.6. Protein Expression Studies
2.6.1. Western Blot Studies
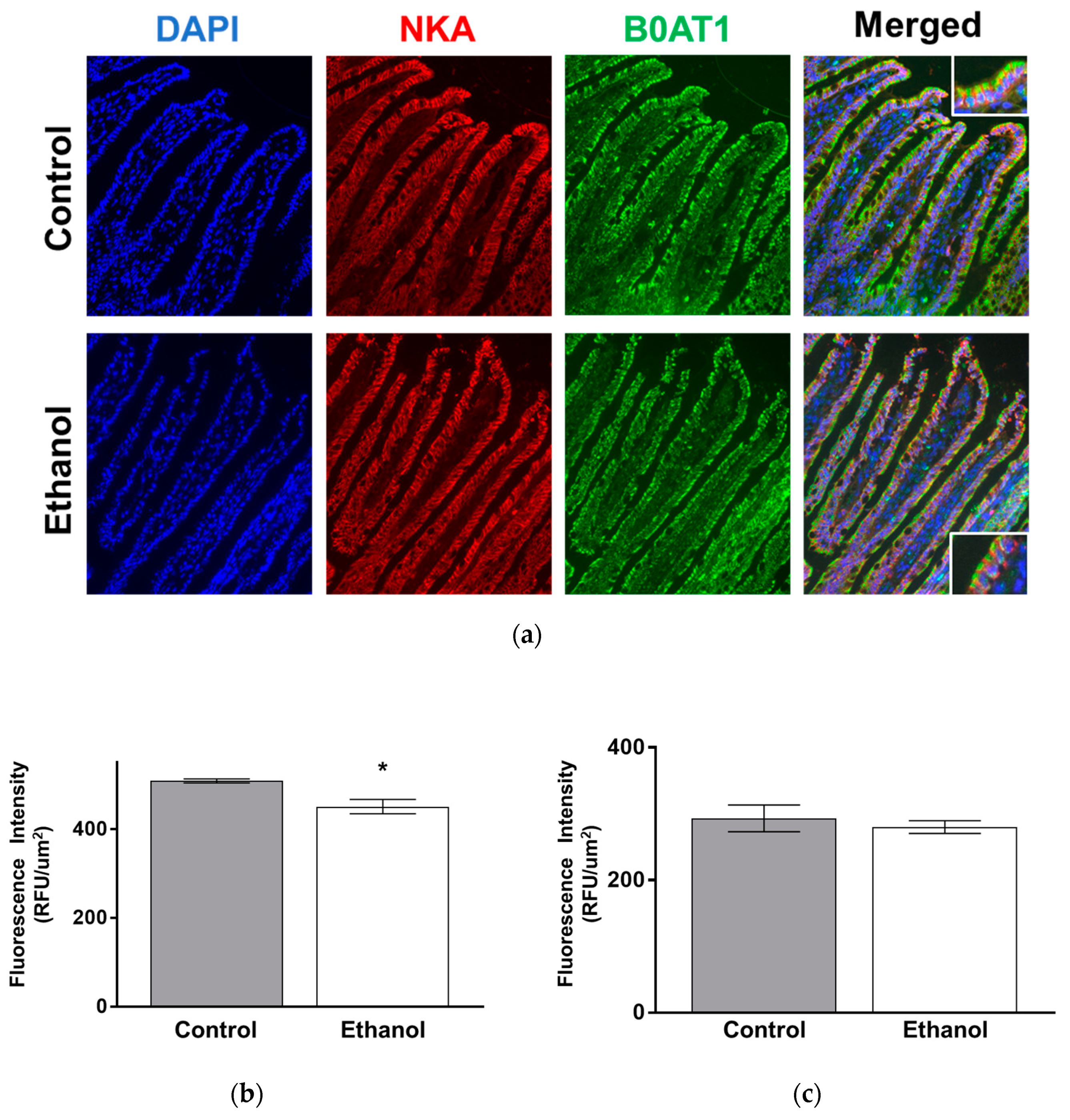
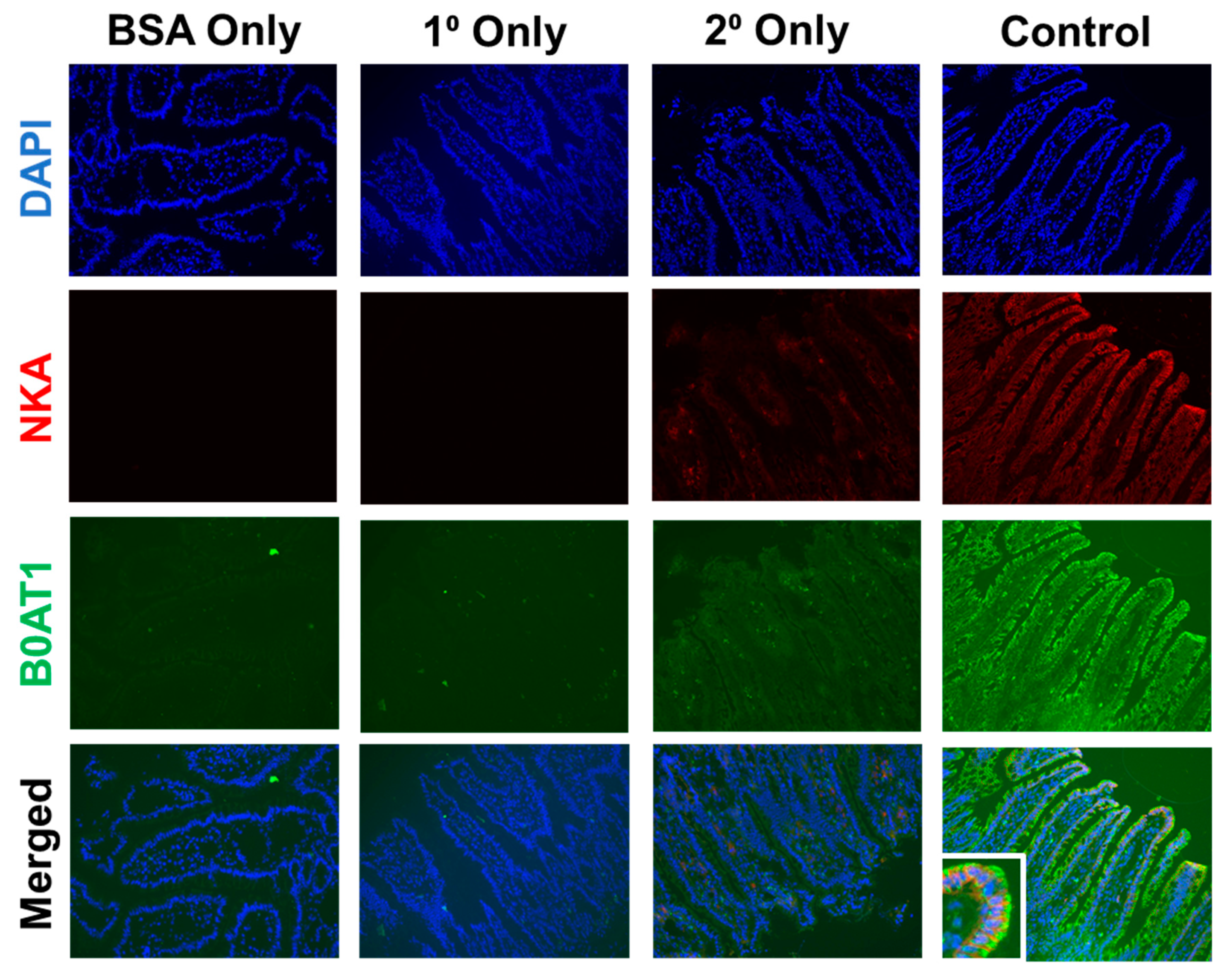
2.6.2. Immunohistochemistry (IHC)
2.6.3. Immunocytochemistry (ICC)
2.7. Uptake Studies
2.7.1. Brush Border Membrane Vesicle (BBMV) Preparation
2.7.2. Kinetics
2.8. Statistical Analysis
3. Results
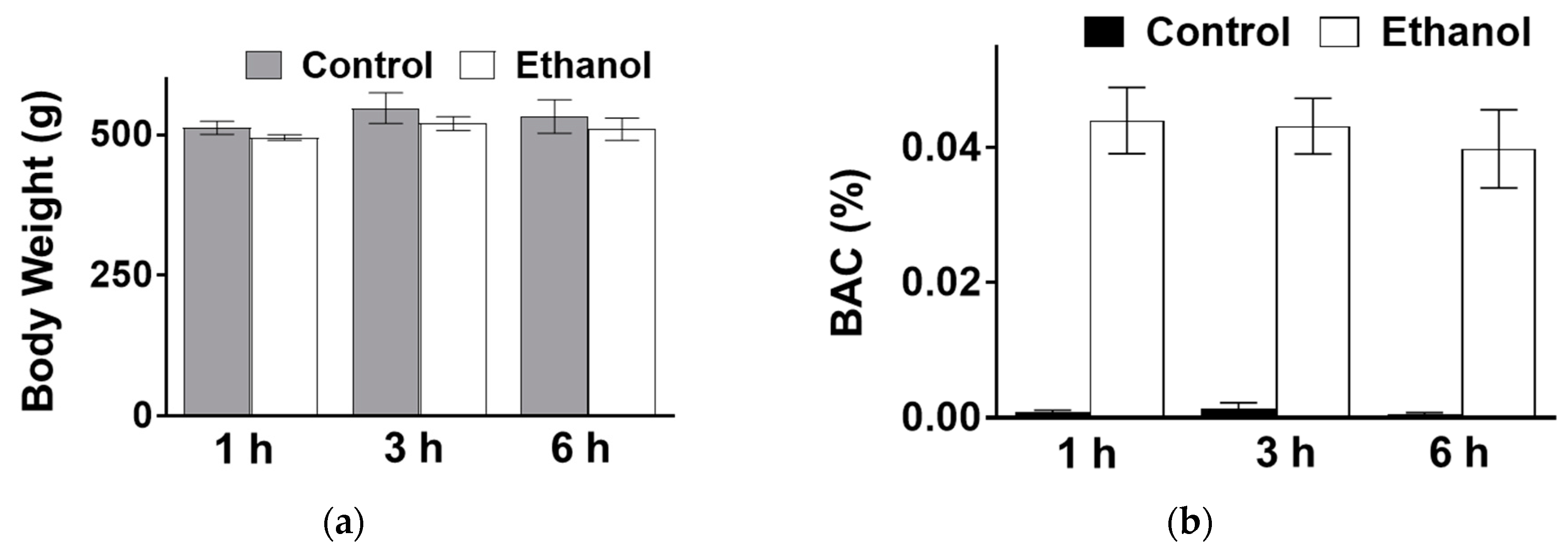
3.1. The Effect of Moderate Ethanol on Body Weight and BAC in SD Rats Administered Via Oral Gavage
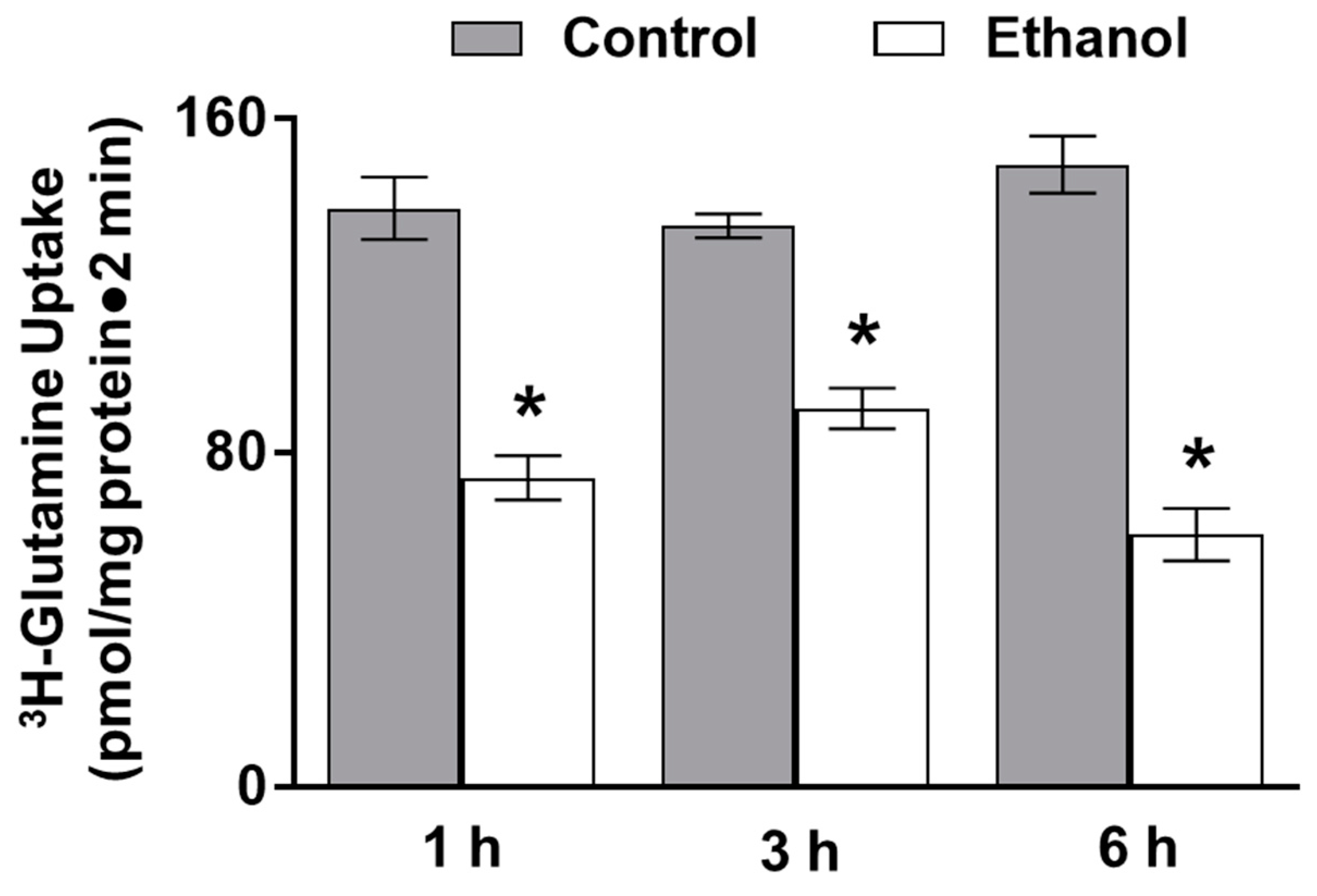
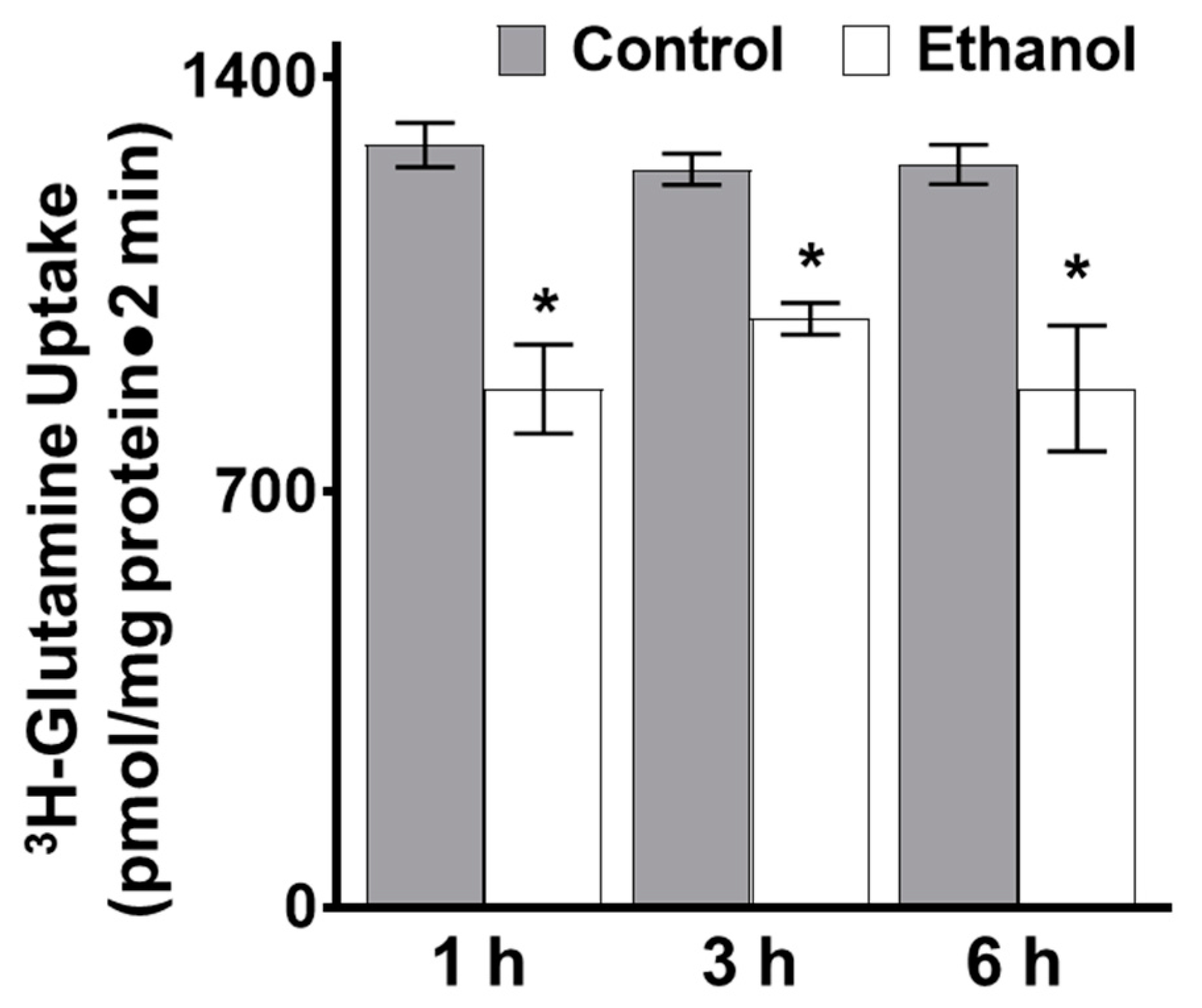
3.1.1. The Effect of Moderate Ethanol on Sodium-Dependent Glutamine Uptake in Ex Vivo Intestinal Epithelial Cells
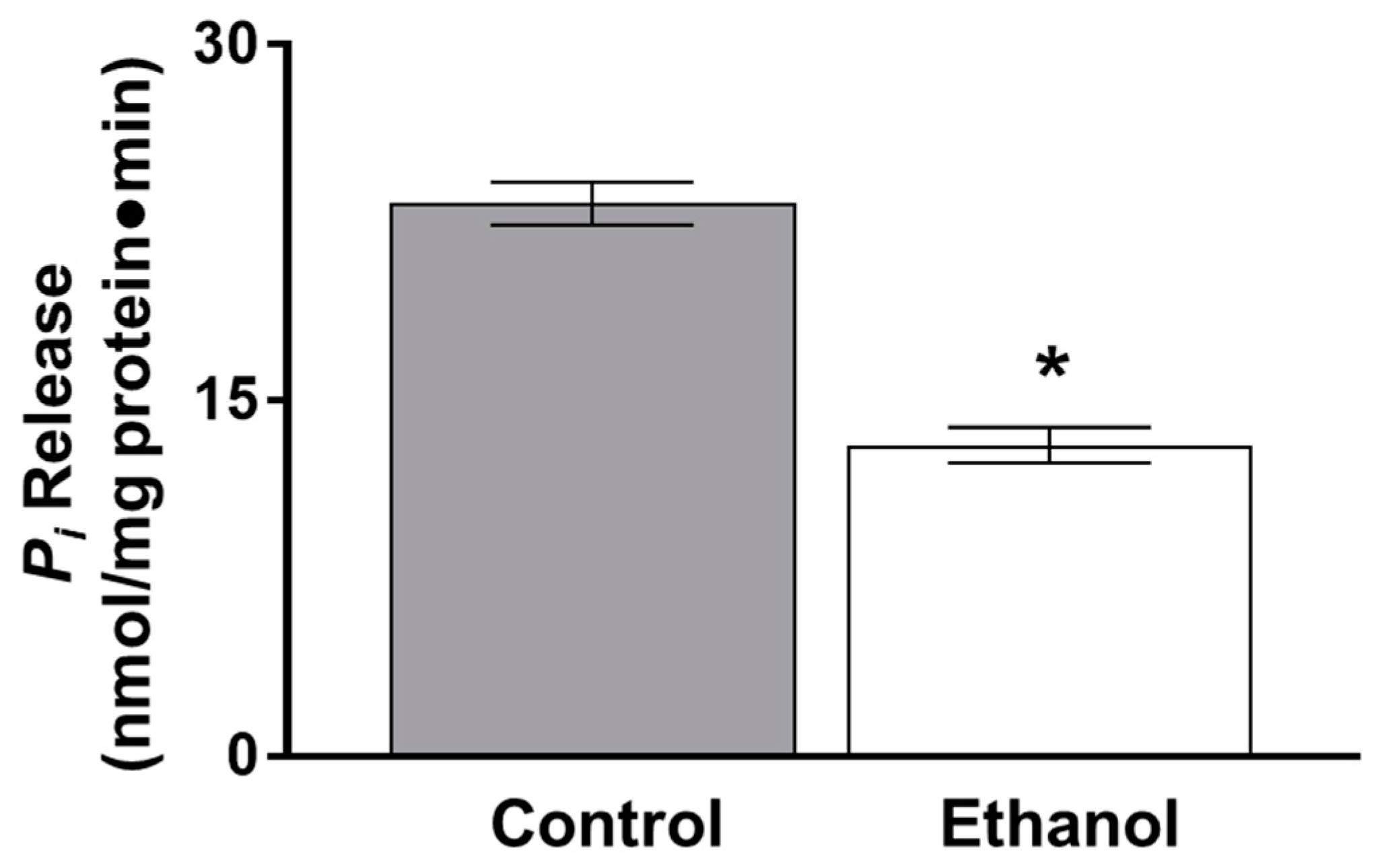
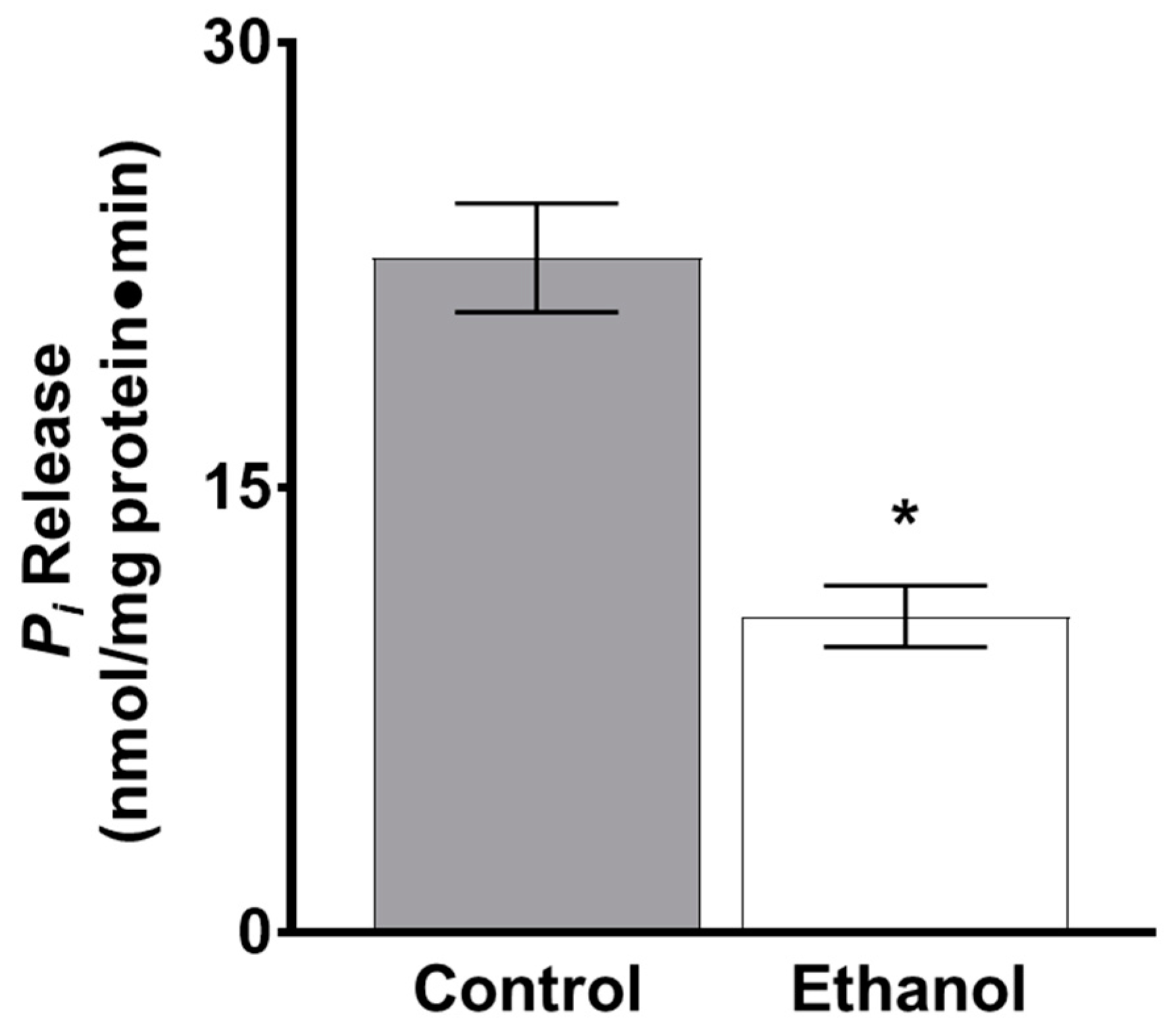
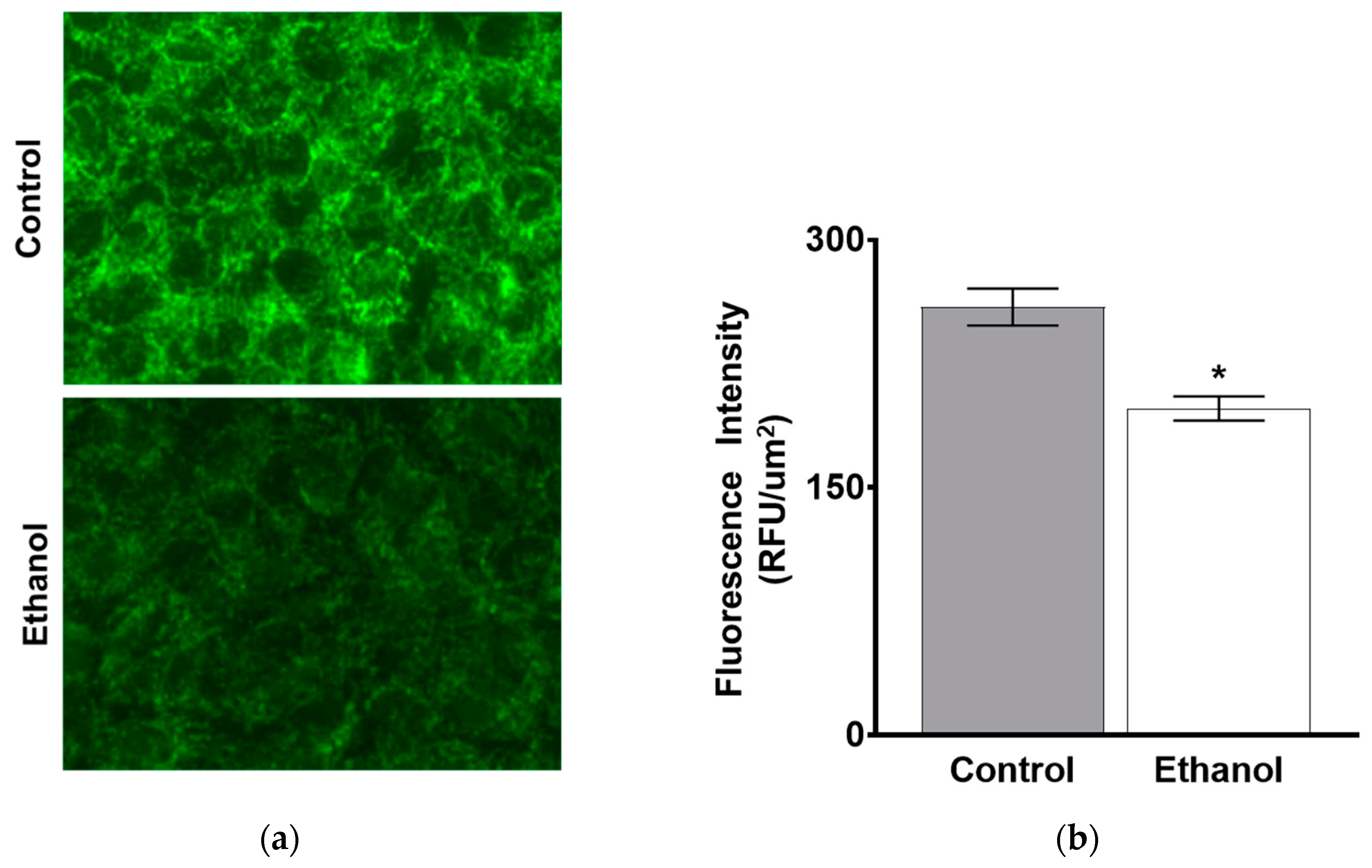
3.1.2. The Effect of Moderate Ethanol on Ex Vivo Na-K-ATPase Activity
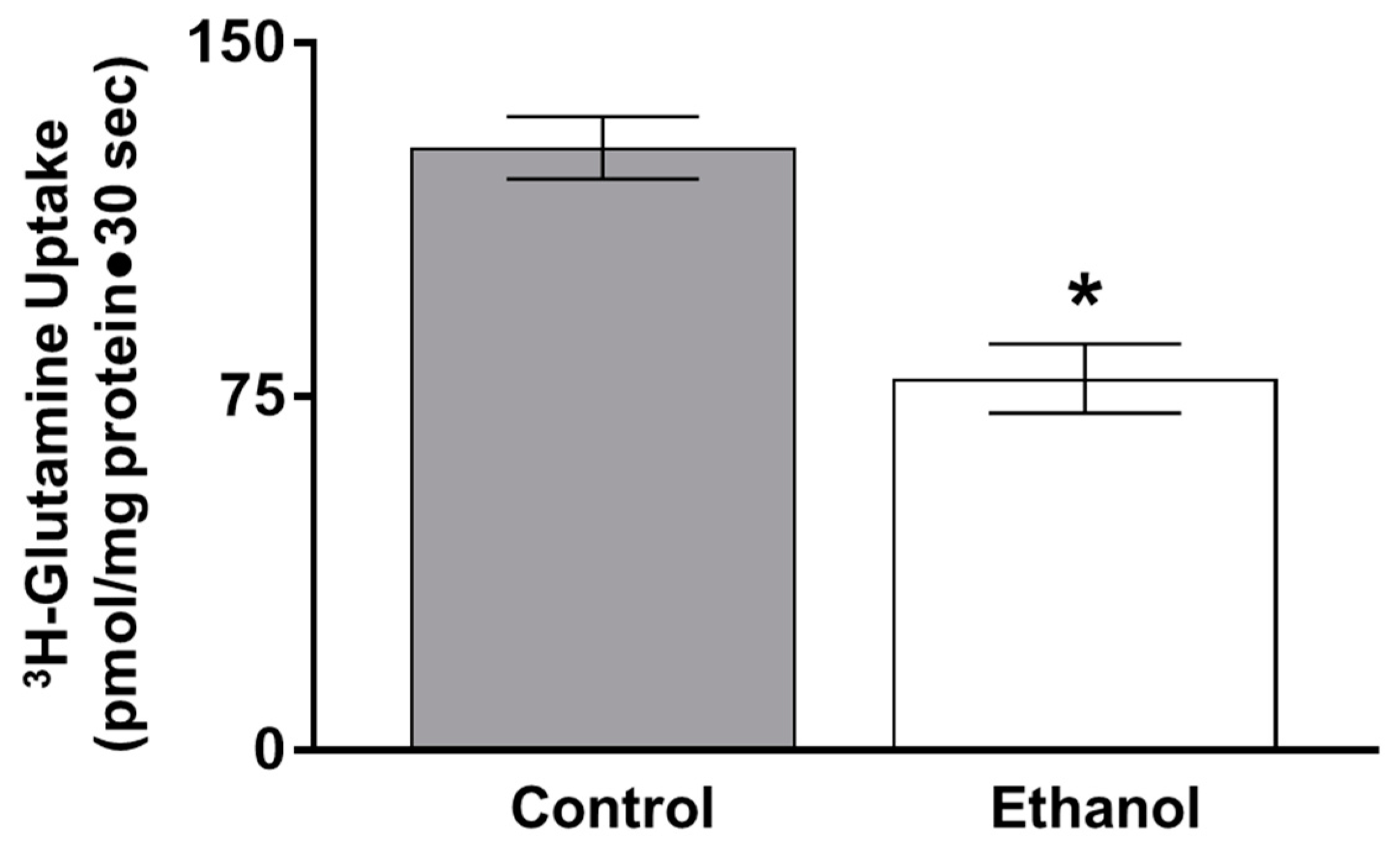
3.1.3. The Effect of Ethanol on Sodium-dependent Glutamine BBMV Uptakes in SD Rats
3.1.4. The Effect of Moderate Ethanol on the Kinetic Parameters of Sodium-Dependent Glutamine Co-Transport in SD Rats
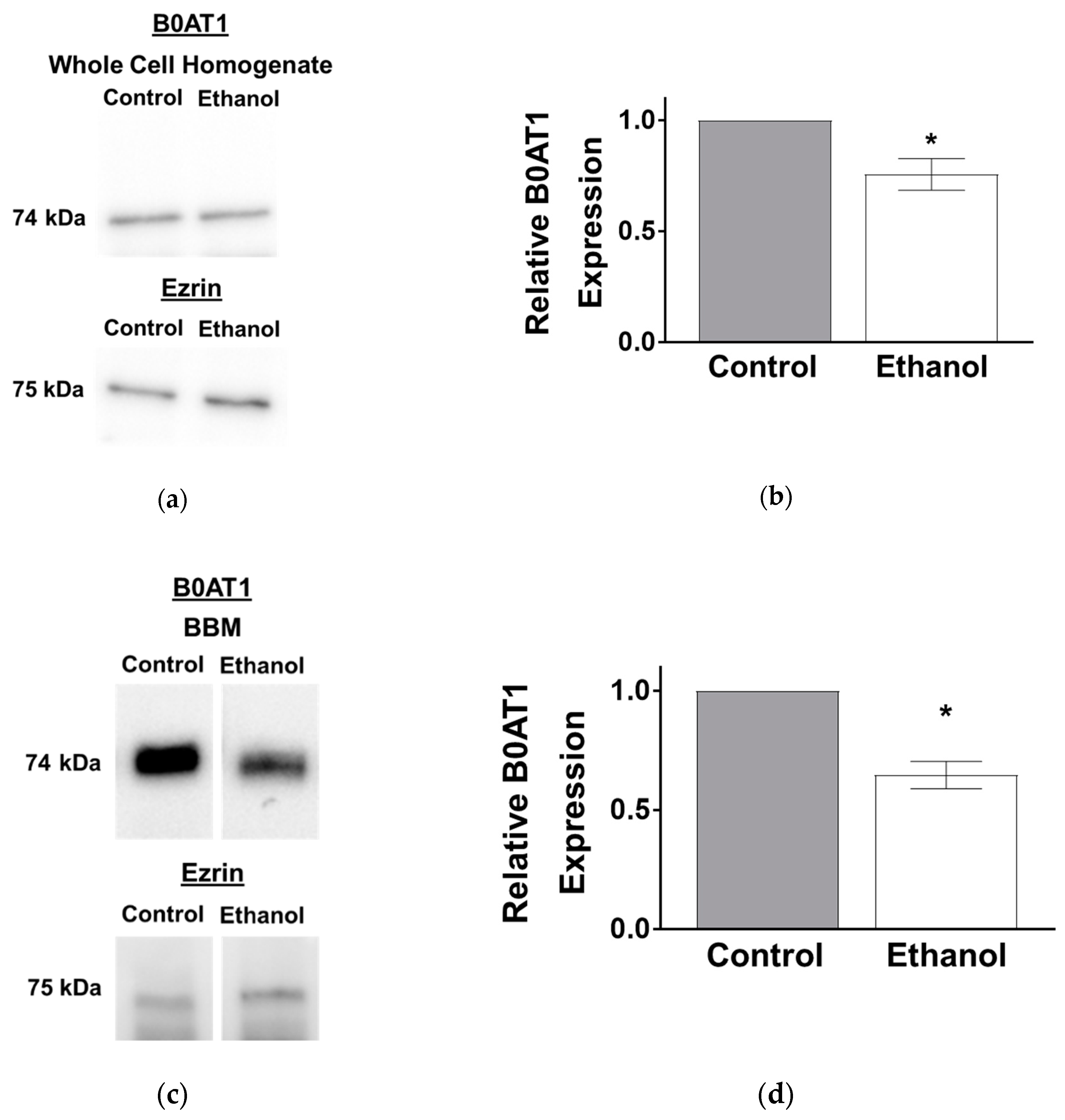
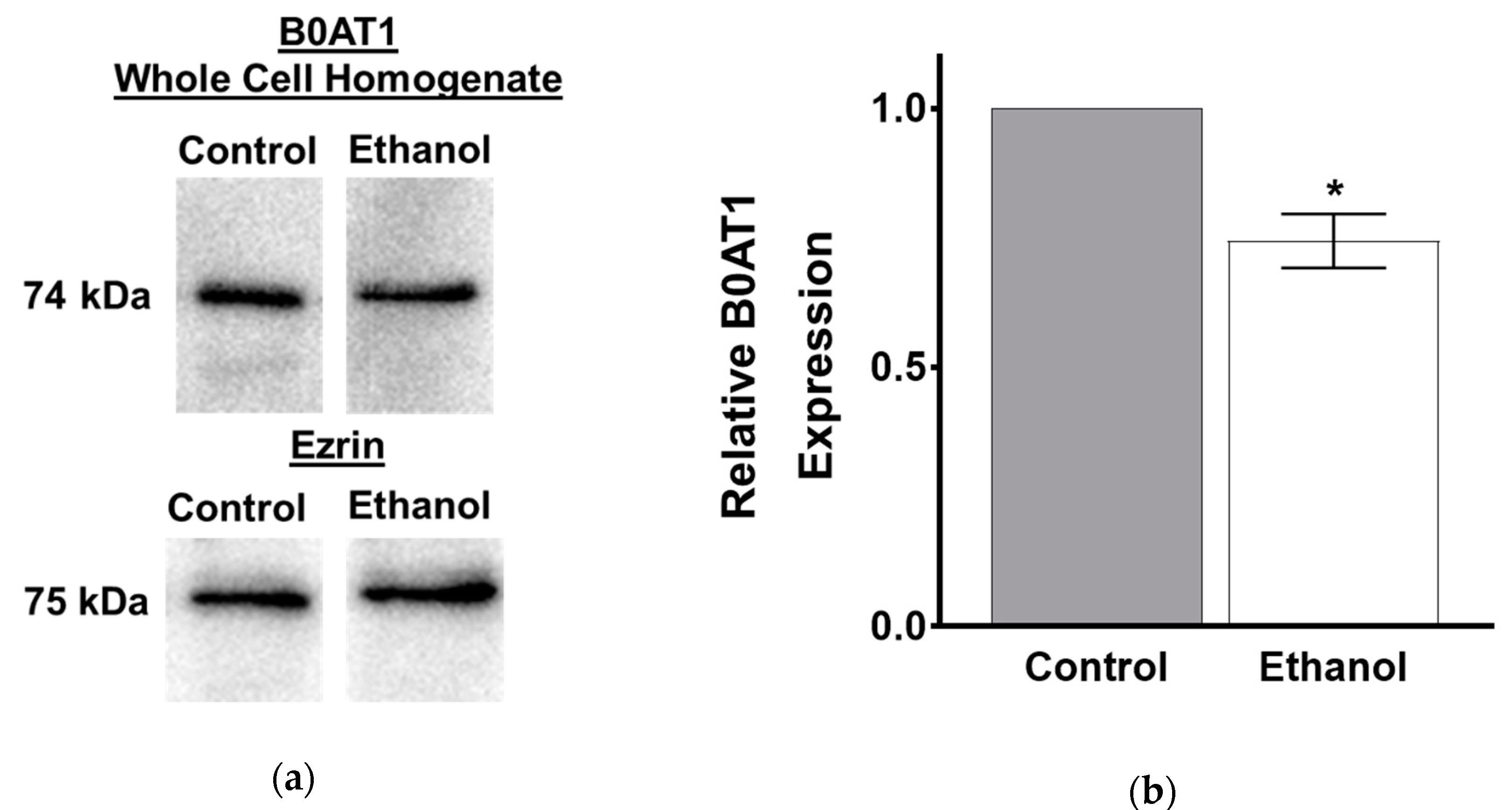
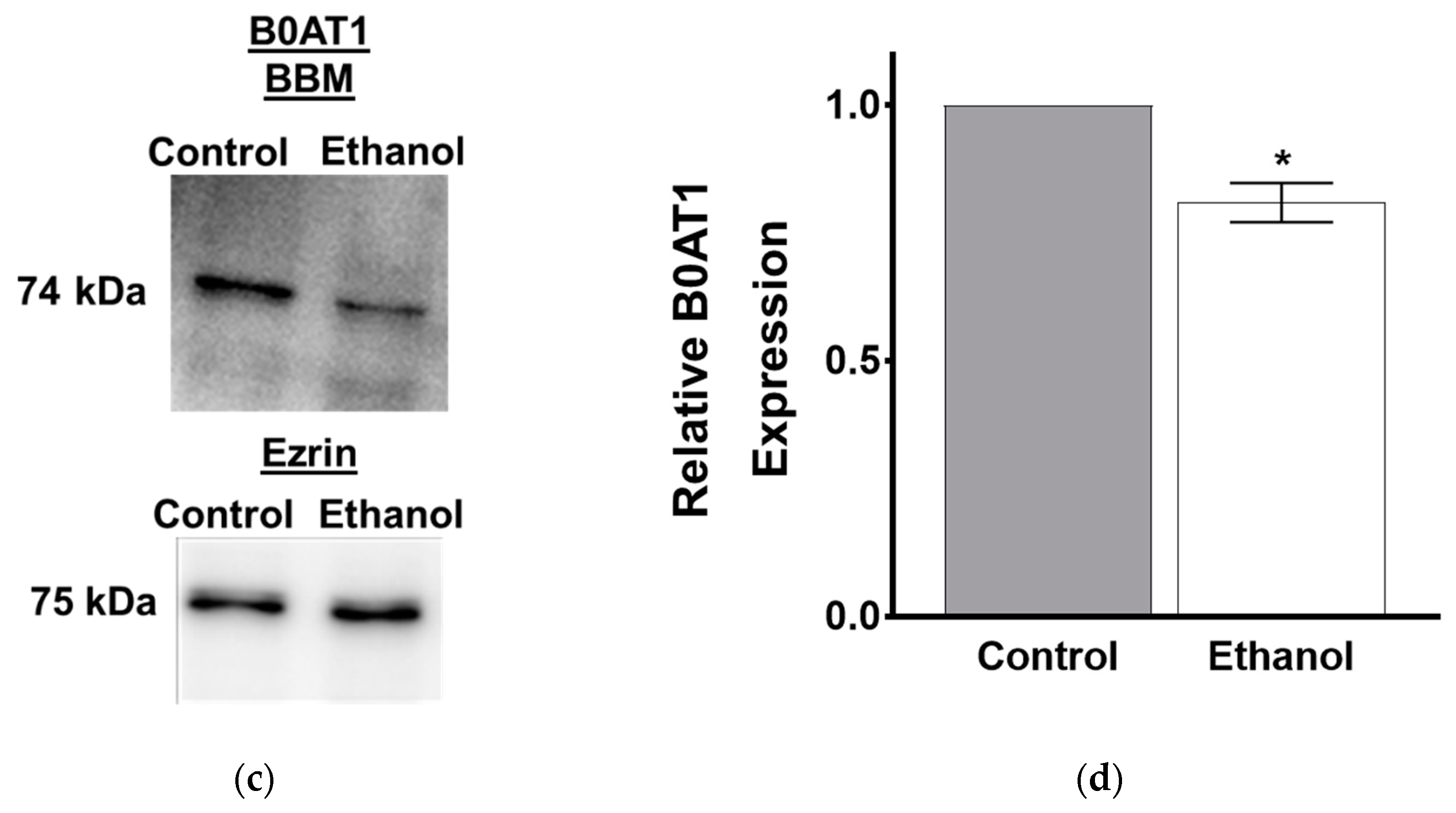
3.1.5. Protein Expression of B0AT1 in SD Rats Exposed to Moderate Ethanol
3.2. The Effect of Moderate Ethanol on Sodium-Dependent Glutamine Uptake in IEC-18 Cells
3.2.1. The Effect of Moderate Ethanol on Na-K-ATPase Activity in IEC-18 Cells
3.2.2. The Effect of Moderate Ethanol on the Kinetic Parameters of Sodium-Dependent Glutamine Co-Transport in IEC-18 Cells
3.2.3. Protein Expression of B0AT1 in IEC-18 Cells Exposed to Moderate Ethanol
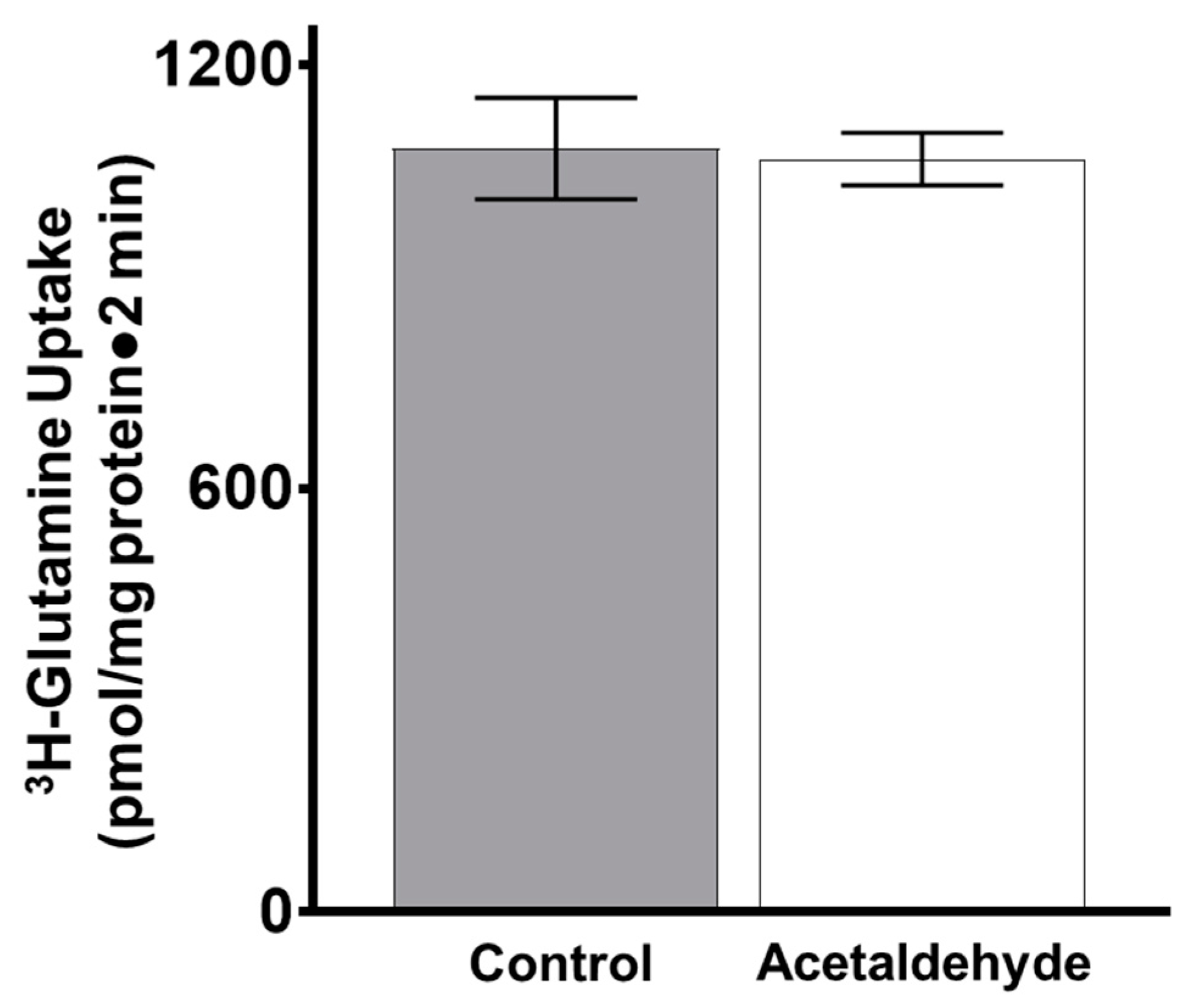
3.3. The Effect of Acetaldehyde on Glutamine Uptake in IEC-18 Cells
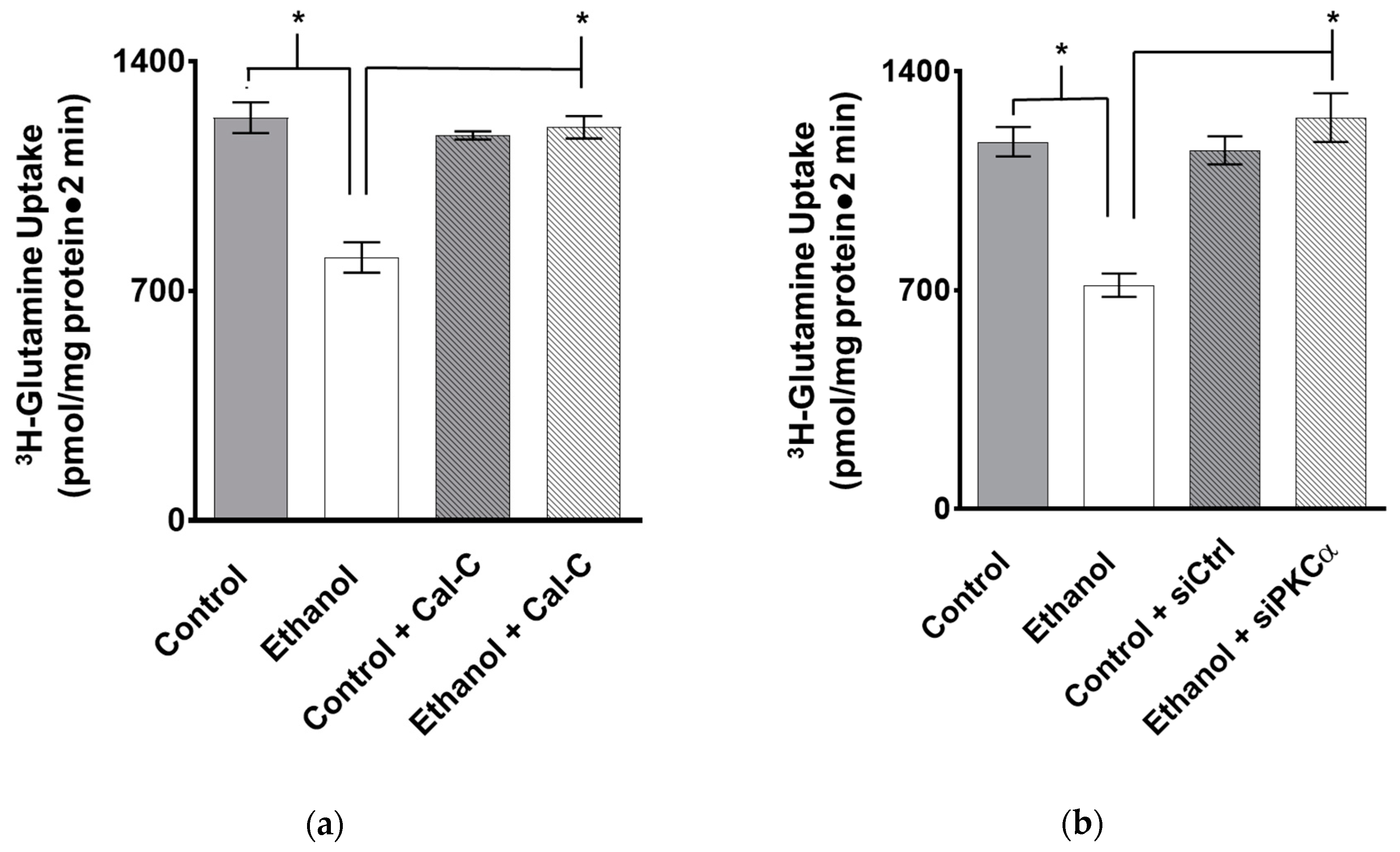
3.4. The Effect of PKC Inhibition on Glutamine Absorption in IEC-18 Cells Exposed to Ethanol
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A. Supplemental Materials and Methods
Appendix A.1. Acetaldehyde Measurement
Appendix A.2. PKCα Western Blots
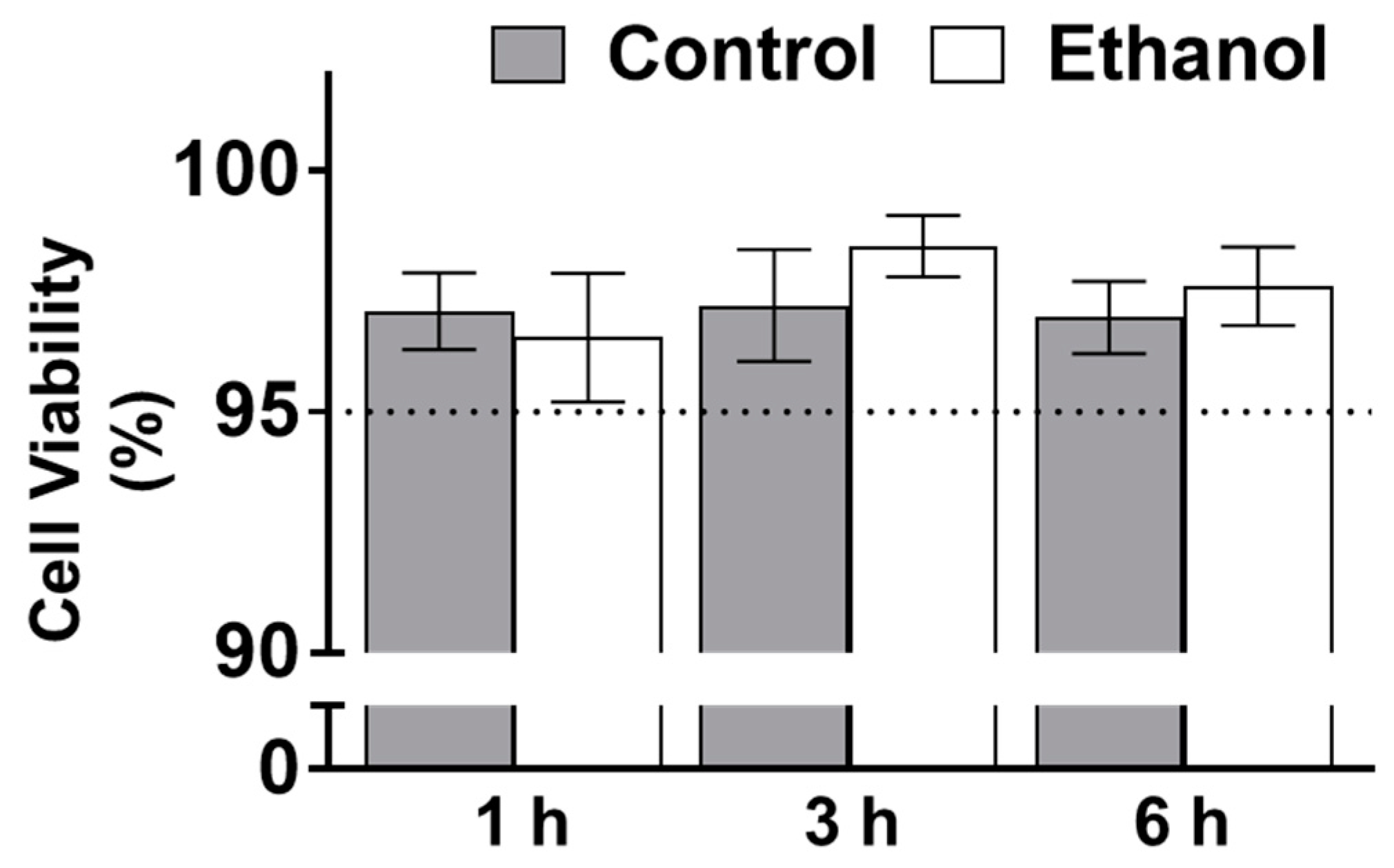
Appendix A.3. Trypan Blue Exclusions
Appendix B. Supplemental Results





References
- Rehm, J.; Baliunas, D.; Borges, G.L.; Graham, K.; Irving, H.; Kehoe, T.; Parry, C.D.; Patra, J.; Popova, S.; Poznyak, V.; et al. The relation between different dimensions of alcohol consumption and burden of disease: An overview. Addiction 2010, 105, 817–843. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Global Status Report on Alcohol and Health; World Health Organization: Geneva, Switzerland, 2018. [Google Scholar]
- McClain, C.J.; Barve, S.S.; Barve, A.; Marsano, L. Alcoholic liver disease and malnutrition. Alcohol. Clin. Exp. Res. 2011, 35, 815–820. [Google Scholar] [CrossRef] [PubMed]
- Bode, C.; Bode, J.C. Effect of alcohol consumption on the gut. Best Pract. Res. Clin. Gastroenterol. 2003, 17, 575–592. [Google Scholar] [CrossRef]
- Chang, T.; Lewis, J.; Glazko, A.J. Effect of ethanol and other alcohols on the transport of amino acids and glucose by everted sacs of rat small intestine. Biochim. Biophys. Acta 1967, 135, 1000–1007. [Google Scholar] [CrossRef]
- Dinda, P.K.; Beck, I.T. On the mechanism of the inhibitory effect of ethanol on intestinal glucose and water absorption. Am. J. Dig. Dis. 1977, 22, 529–533. [Google Scholar] [CrossRef]
- Dinda, P.K.; Beck, I.T. Ethanol-induced inhibition of glucose transport across the isolated brush-border membrane of hamster jejunum. Dig. Dis. Sci. 1981, 26, 23–32. [Google Scholar] [CrossRef]
- Dinda, P.K.; Beck, I.T.; Beck, M.; McElligott, T.F. Effect of ethanol on sodium-dependent glucose transport in the small intestine of the hamster. Gastroenterology. 1975, 68, 1517–1526. [Google Scholar]
- Pfeiffer, A.; Schmidt, T.; Vidon, N.; Pehl, C.; Kaess, H. Absorption of a nutrient solution in chronic alcoholics without nutrient deficiencies and liver cirrhosis. Scand. J. Gastroenterol. 1992, 27, 1023–1030. [Google Scholar] [CrossRef]
- Guo, X.; Wang, Y.; Shen, Y.; Gao, Y.; Chang, Y.; Duan, X. Gene expression profiles of sodium-dependent vitamin C transporters in mice after alcohol consumption. Acta Biochim. Biophys. Sin. 2013, 45, 912–920. [Google Scholar] [CrossRef] [Green Version]
- Subramanya, S.B.; Subramanian, V.S.; Said, H.M. Chronic alcohol consumption and intestinal thiamin absorption: Effects on physiological and molecular parameters of the uptake process. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 299, G23–G31. [Google Scholar] [CrossRef]
- Romanoff, R.L.; Ross, D.M.; McMartin, K.E. Acute ethanol exposure inhibits renal folate transport, but repeated exposure upregulates folate transport proteins in rats and human cells. J. Nutr. 2007, 137, 1260–1265. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, V.S.; Subramanya, S.B.; Ghosal, A.; Said, H.M. Chronic alcohol feeding inhibits physiological and molecular parameters of intestinal and renal riboflavin transport. Am. J. Physiol. Cell Physiol. 2013, 305, C539–C546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pochini, L.; Scalise, M.; Galluccio, M.; Indiveri, C. Membrane transporters for the special amino acid glutamine: Structure/function relationships and relevance to human health. Front. Chem. 2014, 2, 61. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Arthur, S.; Sundaram, U. Unique regulation of Na-glutamine cotransporter SN2/SNAT5 in rabbit intestinal crypt cells during chronic enteritis. J. Cell. Mol. Med. 2018, 22, 1443–1451. [Google Scholar] [CrossRef]
- Singh, S.; Arthur, S.; Talukder, J.; Palaniappan, B.; Coon, S.; Sundaram, U. Mast cell regulation of Na-glutamine co-transporters B0AT1 in villus and SN2 in crypt cells during chronic intestinal inflammation. BMC Gastroenterol. 2015, 15, 47. [Google Scholar] [CrossRef]
- Talukder, J.R.; Kekuda, R.; Saha, P.; Arthur, S.; Sundaram, U. Identification and characterization of rabbit small intestinal villus cell brush border membrane Na-glutamine cotransporter. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 295, G7–G15. [Google Scholar] [CrossRef] [Green Version]
- Curi, R.; Lagranha, C.J.; Doi, S.Q.; Sellitti, D.F.; Procopio, J.; Pithon-Curi, T.C.; Corless, M.; Newsholme, P. Molecular mechanisms of glutamine action. J. Cell. Physiol. 2005, 204, 392–401. [Google Scholar] [CrossRef]
- Livy, D.J.; Parnell, S.E.; West, J.R. Blood ethanol concentration profiles: A comparison between rats and mice. Alcohol 2003, 29, 165–171. [Google Scholar] [CrossRef]
- Arthur, S.; Sundaram, U. Protein kinase C-mediated phosphorylation of RKIP regulates inhibition of Na-alanine cotransport by leukotriene D (4) in intestinal epithelial cells. Am. J. Physiol. Cell Physiol. 2014, 307, C1010–C1016. [Google Scholar] [CrossRef]
- Forbush, B., 3rd. Assay of Na,K-ATPase in plasma membrane preparations: Increasing the permeability of membrane vesicles using sodium dodecyl sulfate buffered with bovine serum albumin. Anal. Biochem. 1983, 128, 159–163. [Google Scholar] [CrossRef]
- Sundaram, U.; Knickelbein, R.G.; Dobbins, J.W. pH regulation in ileum: Na(+)-H+ and Cl(-)-HCO3- exchange in isolated crypt and villus cells. Am. J. Physiol. 1991, 260, G440–G449. [Google Scholar] [CrossRef] [PubMed]
- Arthur, S.; Singh, S.; Sundaram, U. Cyclooxygenase pathway mediates the inhibition of Na-glutamine co-transporter B0AT1 in rabbit villus cells during chronic intestinal inflammation. PLoS ONE 2018, 13, 9–e0203552. [Google Scholar] [CrossRef] [PubMed]
- Arthur, S.; Coon, S.; Kekuda, R.; Sundaram, U. Regulation of sodium glucose co-transporter SGLT1 through altered glycosylation in the intestinal epithelial cells. Biochim. Biophys. Acta 2014, 1838, 1208–1214. [Google Scholar] [CrossRef] [Green Version]
- Fisher, S.J.; Swaan, P.W.; Eddington, N.D. The ethanol metabolite acetaldehyde increases paracellular drug permeability in vitro and oral bioavailability in vivo. J. Pharmacol. Exp. Ther. 2010, 332, 326–333. [Google Scholar] [CrossRef] [PubMed]
- Stubbs, C.D.; Slater, S.J. Ethanol and protein kinase C. Alcohol. Clin. Exp. Res. 1999, 23, 1552–1560. [Google Scholar] [CrossRef] [PubMed]
- Hoyumpa, A.M., Jr. Mechanisms of thiamin deficiency in chronic alcoholism. Am. J. Clin. Nutr. 1980, 33, 2750–2761. [Google Scholar] [CrossRef]
- Hoyumpa, A.M., Jr.; Breen, K.J.; Schenker, S.; Wilson, F.A. Thiamine transport across the rat intestine. II. Effect of ethanol. J. Lab. Clin. Med. 1975, 86, 803–816. [Google Scholar]
- Hoyumpa, A.M., Jr.; Patwardhan, R.; Antonson, D.; Nichols, S.; Gray, J.P. Effect of thiamin deficiency and acute ethanol ingestion on jejunal glucose transport in rats. Am. J. Clin. Nutr. 1981, 34, 14–19. [Google Scholar] [CrossRef]
- Hamid, A.; Wani, N.A.; Rana, S.; Vaiphei, K.; Mahmood, A.; Kaur, J. Down-regulation of reduced folate carrier may result in folate malabsorption across intestinal brush border membrane during experimental alcoholism. FEBS J. 2007, 274, 6317–6328. [Google Scholar] [CrossRef]
- Thakur, S.; Rahat, B.; More, D.; Kaur, J. Reduced SP1-mediated transcriptional activation decreases expression of intestinal folate transporters in response to ethanol exposure. Mol. Nutr. Food Res. 2015, 59, 1713–1724. [Google Scholar] [CrossRef]
- Ducroc, R.; Sakar, Y.; Fanjul, C.; Barber, A.; Bado, A.; Lostao, M.P. Luminal leptin inhibits L-glutamine transport in rat small intestine: Involvement of ASCT2 and B0AT1. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 299, G179–G185. [Google Scholar] [CrossRef] [PubMed]
- Otis, J.S.; Mitchell, P.O.; Kershaw, C.D.; Joshi, P.C.; Guidot, D.M. Na,K-ATPase expression is increased in the lungs of alcohol-fed rats. Alcohol. Clin. Exp. Res. 2008, 32, 699–705. [Google Scholar] [CrossRef] [PubMed]
- Rodrigo, R.; Thielemann, L. Effects of chronic and acute ethanol exposure on renal (Na + K)-ATPase in the rat. Gen. Pharmacol. 1997, 29, 719–723. [Google Scholar] [CrossRef]
- Rasic-Markovic, A.; Krstic, D.; Vujovic, Z.; Jakovljevic, V.; Stanojlovic, O.; Hrncic, D.; Djuric, D.; Loncar-Stevanovic, H. Modulations of rabbit erythrocyte ATPase activities induced by in vitro and in vivo exposure to ethanol. Mol. Cell. Biochem. 2008, 308, 111–116. [Google Scholar] [CrossRef]
- Atkinson, K.J.; Rao, R.K. Role of protein tyrosine phosphorylation in acetaldehyde-induced disruption of epithelial tight junctions. Am. J. Physiol. Gastrointest. Liver Physiol. 2001, 280, G1280–G1288. [Google Scholar] [CrossRef]
- Basuroy, S.; Sheth, P.; Mansbach, C.M.; Rao, R.K. Acetaldehyde disrupts tight junctions and adherens junctions in human colonic mucosa: Protection by EGF and l-glutamine. Am. J. Physiol.-Gastrointest. Liver Physiol. 2005, 289, G367–G375. [Google Scholar] [CrossRef]
- Seitz, H.K.; Stickel, F. Acetaldehyde as an underestimated risk factor for cancer development: Role of genetics in ethanol metabolism. Genes. Nutr. 2010, 5, 121–128. [Google Scholar] [CrossRef]
- Bibert, S.; Roy, S.; Schaer, D.; Horisberger, J.D.; Geering, K. Phosphorylation of phospholemman (FXYD1) by protein kinases A and C modulates distinct Na,K-ATPase isozymes. J. Boil. Chem. 2008, 283, 476–486. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vmax (nmol/mg Protein·6 sec) | Km (mM) | |
|---|---|---|
| Control | 3.2 ± 0.1 | 26 ± 1.4 |
| Ethanol | 2.6 ± 0.1 * | 26 ± 2.2 |
| Vmax (nmol/mg Protein·30 sec) | Km (mM) | |
|---|---|---|
| Control | 1.5 ± 0.03 | 0.1 ± 0.01 |
| Ethanol | 1.3 ± 0.03 * | 0.1 ± 0.01 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Butts, M.; Singh Paulraj, R.; Haynes, J.; Arthur, S.; Singh, S.; Sundaram, U. Moderate Alcohol Consumption Inhibits Sodium-Dependent Glutamine Co-Transport in Rat Intestinal Epithelial Cells in Vitro and Ex Vivo. Nutrients 2019, 11, 2516. https://doi.org/10.3390/nu11102516
Butts M, Singh Paulraj R, Haynes J, Arthur S, Singh S, Sundaram U. Moderate Alcohol Consumption Inhibits Sodium-Dependent Glutamine Co-Transport in Rat Intestinal Epithelial Cells in Vitro and Ex Vivo. Nutrients. 2019; 11(10):2516. https://doi.org/10.3390/nu11102516
Chicago/Turabian StyleButts, Molly, Raja Singh Paulraj, Jennifer Haynes, Subha Arthur, Soudamani Singh, and Uma Sundaram. 2019. "Moderate Alcohol Consumption Inhibits Sodium-Dependent Glutamine Co-Transport in Rat Intestinal Epithelial Cells in Vitro and Ex Vivo" Nutrients 11, no. 10: 2516. https://doi.org/10.3390/nu11102516
APA StyleButts, M., Singh Paulraj, R., Haynes, J., Arthur, S., Singh, S., & Sundaram, U. (2019). Moderate Alcohol Consumption Inhibits Sodium-Dependent Glutamine Co-Transport in Rat Intestinal Epithelial Cells in Vitro and Ex Vivo. Nutrients, 11(10), 2516. https://doi.org/10.3390/nu11102516