Human Milk Omega-3 Fatty Acid Composition Is Associated with Infant Temperament

Abstract
:1. Introduction
2. Methods
2.1. Participants
2.2. Determination of Milk Fatty Acid Levels
2.3. Infant Temperament
2.4. Demographic and Health Information
2.5. Statistical Analysis Strategy
3. Results
3.1. Preliminary and Descriptive Analyses
3.2. Primary Analysis
3.3. Moderation Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Prado, E.L.; Dewey, K.G. Nutrition and brain development in early life. Nutr. Rev. 2014, 72, 267–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keunen, K.; Van Elburg, R.M.; Van Bel, F.; Benders, M.J. Impact of nutrition on brain development and its neuroprotective implications following preterm birth. Pediatr. Res. 2015, 77, 148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cusick, S.E.; Georgieff, M.K. The role of nutrition in brain development: The golden opportunity of the “first 1000 days”. J. Pediatr. 2016, 175, 16–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belfort, M.B.; Anderson, P.J.; Nowak, V.A.; Lee, K.J.; Molesworth, C.; Thompson, D.K.; Doyle, L.W.; Inder, T.E. Breast milk feeding, brain development, and neurocognitive outcomes: A 7-year longitudinal study in infants born at less than 30 weeks’ gestation. J. Pediatr. 2016, 177, 133–139. [Google Scholar] [CrossRef] [Green Version]
- Martin, C.; Ling, P.R.; Blackburn, G. Review of infant feeding: Key features of breast milk and infant formula. Nutrients 2016, 8, 279. [Google Scholar] [CrossRef] [Green Version]
- Innis, S.M.; Gilley, J.; Werker, J. Are human milk long-chain polyunsaturated fatty acids related to visual and neural development in breast-fed term infants? J. Pediatr. 2001, 139, 532–538. [Google Scholar] [CrossRef]
- Eidelman, A.I.; Schanler, R.J. Breastfeeding and the use of human milk. Pediatrics 2012, 5, 323–324. [Google Scholar]
- Birch, D.G.; Birch, E.E.; Hoffman, D.R.; Uauy, R.D. Retinal development in very-low-birth-weight infants fed diets differing in omega-3 fatty acids. Investig. Ophthalmol. Vis. Sci. 1992, 33, 2365–2376. [Google Scholar]
- Innis, S.M. Dietary omega 3 fatty acids and the developing brain. Brain. Res. 2008, 1237, 35–43. [Google Scholar] [CrossRef]
- Jensen, R.G. Lipids in human milk. Lipids 1999, 34, 1243–1271. [Google Scholar] [CrossRef]
- Brenna, J.T.; Salem, N., Jr.; Sinclair, A.J.; Cunnane, S.C. α-Linolenic acid supplementation and conversion to n-3 long-chain polyunsaturated fatty acids in humans. Prostaglandins Leukot. Essent. Fat. Acids 2009, 80, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Giusto, N.; Salvador, G.; Castagnet, P.; Pasquare, S.; de Boschero, M.I. Age-associated changes in central nervous system glycerolipid composition and metabolism. Neurochem. Res. 2002, 27, 1513–1523. [Google Scholar] [CrossRef] [PubMed]
- Meydani, S.N. Effect of (n-3) polyunsaturated fatty acidson cytokine production and their biologic function. Nutrition 1996, 12, S8–S14. [Google Scholar] [CrossRef]
- Endres, S.; Ghorbani, R.; Kelley, V.E.; Georgilis, K.; Lonnemann, G.; Van Der Meer, J.W.; Cannon, J.G.; Rogers, T.S.; Klempner, M.S.; Weber, P.C. The effect of dietary supplementation with n—3 polyunsaturated fatty acids on the synthesis of interleukin-1 and tumor necrosis factor by mononuclear cells. N. Engl. J. Med. 1989, 320, 265–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- U.S Department of Health and Human Services. 2015–2020 Dietary Guidelines for Americans, 8th ed.; U.S Department of Health and Human Services: Washington, DC, USA, 2015.
- Decsi, T.; Kelemen, B.; Minda, H.; Burus, I.; Kohn, G. Effect of type of early infant feeding on fatty acid composition of plasma lipid classes in full-term infants during the second 6 months of life. J. Pediatr. Gastroenterol. Nutr. 2000, 30, 547–551. [Google Scholar] [CrossRef] [PubMed]
- Cunnane, S.C.; Francescutti, V.; Brenna, J.T.; Crawford, M.A. Breast-fed infants achieve a higher rate of brain and whole body docosahexaenoate accumulation than formula-fed infants not consuming dietary docosahexaenoate. Lipids 2000, 35, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Su, K.P.; Matsuoka, Y.; Pae, C.U. Omega-3 polyunsaturated fatty acids in prevention of mood and anxiety disorders. Clin. Psychopharmacol. Neurosci. 2015, 13, 129. [Google Scholar] [CrossRef] [Green Version]
- Su, K.P. Biological Mechanism of Antidepressant Effect of Omega–3 Fatty Acids: How Does Fish Oil Act as a ‘Mind-Body Interface’? Neurosignals 2009, 17, 144–152. [Google Scholar] [CrossRef]
- Lin, P.Y.; Huang, S.Y.; Su, K.P. A meta-analytic review of polyunsaturated fatty acid compositions in patients with depression. Biol. Psychiatry 2010, 68, 140–147. [Google Scholar] [CrossRef]
- Stoll, A.L.; Severus, W.E.; Freeman, M.P.; Rueter, S.; Zboyan, H.A.; Diamond, E.; Cress, K.K.; Marangell, L.B. Omega 3 fatty acids in bipolar disorder: A preliminary double-blind, placebo-controlled trial. Arch. Gen. Psychiatry 1999, 56, 407–412. [Google Scholar] [CrossRef]
- Buydens-Branchey, L.; Branchey, M.; Hibbeln, J.R. Associations between increases in plasma n-3 polyunsaturated fatty acids following supplementation and decreases in anger and anxiety in substance abusers. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2008, 32, 568–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhatia, H.S.; Agrawal, R.; Sharma, S.; Huo, Y.-X.; Ying, Z.; Gomez-Pinilla, F. Omega-3 fatty acid deficiency during brain maturation reduces neuronal and behavioral plasticity in adulthood. PLoS ONE 2011, 6, e28451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brunst, K.J.; Enlow, M.B.; Kannan, S.; Carroll, K.N.; Coull, B.A.; Wright, R.J. Effects of prenatal social stress and maternal dietary fatty acid ratio on infant temperament: Does race matter? Epidemiology 2014, 4, 1000167. [Google Scholar] [PubMed]
- Clayton, E.H.; Hanstock, T.L.; Hirneth, S.J.; Kable, C.J.; Garg, M.L.; Hazell, P.L. Long-chain omega-3 polyunsaturated fatty acids in the blood of children and adolescents with juvenile bipolar disorder. Lipids 2008, 43, 1031–1038. [Google Scholar] [CrossRef] [PubMed]
- Grey, K.R.; Davis, E.P.; Sandman, C.A.; Glynn, L.M. Human milk cortisol is associated with infant temperament. Psychoneuroendocrinology 2013, 38, 1178–1185. [Google Scholar] [CrossRef] [Green Version]
- Hart, S.; Boylan, L.M.; Border, B.; Carroll, S.R.; McGunegle, D.; Lampe, R.M. Breast milk levels of cortisol and Secretory Immunoglobulin A (SIgA) differ with maternal mood and infant neuro-behavioral functioning. Infant Behav. Dev. 2004, 27, 101–106. [Google Scholar] [CrossRef]
- Nolvi, S.; Uusitupa, H.M.; Bridgett, D.J.; Pesonen, H.; Aatsinki, A.K.; Kataja, E.L.; Korja, R.; Karlsson, H.; Karlsson, L. Human milk cortisol concentration predicts experimentally induced infant fear reactivity: Moderation by infant sex. Dev. Sci. 2018, 21, e12625. [Google Scholar] [CrossRef]
- Hinde, K. Lactational programming of infant behavioral phenotype. In Building Babies; Springer: Berlin/Heidelberg, Germany, 2013; pp. 187–207. [Google Scholar]
- Dettmer, A.M.; Murphy, A.M.; Guitarra, D.; Slonecker, E.; Suomi, S.J.; Rosenberg, K.L.; Novak, M.A.; Meyer, J.S.; Hinde, K. Cortisol in neonatal mother’s milk predicts later infant social and cognitive functioning in rhesus monkeys. Child Dev. 2018, 89, 525–538. [Google Scholar] [CrossRef] [Green Version]
- Petrullo, L.; Hinde, K.; Lu, A. Steroid hormone concentrations in milk predict sex-specific offspring growth in a nonhuman primate. Am. J. Hum. Biol. 2019, 31, e23315. [Google Scholar] [CrossRef]
- Hinde, K.; Capitanio, J.P. Lactational programming? Mother’s milk energy predicts infant behavior and temperament in rhesus macaques (Macaca mulatta). Am. J. Primatol. Off. J. Am. Soc. Primatol. 2010, 72, 522–529. [Google Scholar] [CrossRef] [Green Version]
- Gartstein, M.A.; Rothbart, M.K. Studying infant temperament via the revised infant behavior questionnaire. Infant Behav. Dev. 2003, 26, 64–86. [Google Scholar] [CrossRef]
- Francois, C.A.; Connor, S.L.; Bolewicz, L.C.; Connor, W.E. Supplementing lactating women with flaxseed oil does not increase docosahexaenoic acid in their milk. Am. J. Clin. Nutr. 2003, 77, 226–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobson, J.L.; Jacobson, S.W.; Muckle, G.; Kaplan-Estrin, M.; Ayotte, P.; Dewailly, E. Beneficial effects of a polyunsaturated fatty acid on infant development: Evidence from the Inuit of Arctic Quebec. J. Pediatr. 2008, 152, 356–364. [Google Scholar] [CrossRef] [PubMed]
- Worobey, J.; Blajda, V.M. Temperament ratings at 2 weeks, 2 months, and 1 year: Differential stablity of activity and emotionality. Dev. Psychol. 1989, 25, 257. [Google Scholar] [CrossRef]
- World Health Organization. WHO Child Growth Standards: Length/Height-for-Age, Weight-for-Age, Weight-for-Length, Weight-for-Height and Body Mass Index-for-Age: Methods and Development; World Health Organization: Geneva, Switzerland, 2006. [Google Scholar]
- Cole, T.J.; Green, P.J. Smoothing reference centile curves: The LMS method and penalized likelihood. Stat. Med. 1992, 11, 1305–1319. [Google Scholar] [CrossRef]
- Gartstein, M.A.; Putnam, S.P.; Rothbart, M.K. Etiology of preschool behavior problems: Contributions of temperament attributes in early childhood. Infant Ment. Health J. 2012, 33, 197–211. [Google Scholar] [CrossRef]
- Rende, R.D. Longitudinal relations between temperament traits and behavioral syndromes in middle childhood. J. Am. Acad. Child Adolesc. Psychiatry 1993, 32, 287–290. [Google Scholar] [CrossRef]
- Auestad, N.; Halter, R.; Hall, R.T.; Blatter, M.; Bogle, M.L.; Burks, W.; Erickson, J.R.; Fitzgerald, K.M.; Dobson, V.; Innis, S.M. Growth and development in term infants fed long-chain polyunsaturated fatty acids: A double-masked, randomized, parallel, prospective, multivariate study. Pediatrics 2001, 108, 372–381. [Google Scholar] [CrossRef] [Green Version]
- Champoux, M.; Hibbeln, J.R.; Shannon, C.; Majchrzak, S.; Suomi, S.J.; Salem Jr, N.; Higley, J.D. Fatty acid formula supplementation and neuromotor development in rhesus monkey neonates. Pediatr. Res. 2002, 51, 273. [Google Scholar] [CrossRef] [Green Version]
- Carnielli, V.P.; Verlato, G.; Pederzini, F.; Luijendijk, I.; Boerlage, A.; Pedrotti, D.; Sauer, P. Intestinal absorption of long-chain polyunsaturated fatty acids in preterm infants fed breast milk or formula. Am. J. Clin. Nutr. 1998, 67, 97–103. [Google Scholar] [CrossRef]
- de Souza, C.O.; Leite, M.E.Q.; Lasekan, J.; Baggs, G.; Pinho, L.S.; Druzian, J.I.; Ribeiro, T.C.M.; Mattos, Â.P.; Menezes-Filho, J.A.; Costa-Ribeiro, H. Milk protein-based formulas containing different oils affect fatty acids balance in term infants: A randomized blinded crossover clinical trial. Lipids Health Dis. 2017, 16, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delion, S.; Chalon, S.; Hérault, J.; Guilloteau, D.; Besnard, J.C.; Durand, G. Chronic dietary α-linolenic acid deficiency alters dopaminergic and serotoninergic neurotransmission in rats. J. Nutr. 1994, 124, 2466–2476. [Google Scholar] [CrossRef] [PubMed]
- Meltzer, H. Serotonergic dysfunction in depression. Br. J. Psychiatry 1989, 155, 25–31. [Google Scholar] [CrossRef]
- Stahl, L.A.; Begg, D.P.; Weisinger, R.S.; Sinclair, A.J. The role of omega-3 fatty acids in mood disorders. Curr. Opin. Investig. Drugs 2008, 9, 57–64. [Google Scholar]
- Eisenberger, N.I.; Berkman, E.T.; Inagaki, T.K.; Rameson, L.T.; Mashal, N.M.; Irwin, M.R. Inflammation-induced anhedonia: Endotoxin reduces ventral striatum responses to reward. Biol. Psychiatry 2010, 68, 748–754. [Google Scholar] [CrossRef] [Green Version]
- Francois, C.A.; Connor, S.L.; Wander, R.C.; Connor, W.E. Acute effects of dietary fatty acids on the fatty acids of human milk. Am. J. Clin. Nutr. 1998, 67, 301–308. [Google Scholar] [CrossRef]
- Weseler, A.R.; Dirix, C.E.; Bruins, M.J.; Hornstra, G. Dietary arachidonic acid dose-dependently increases the arachidonic acid concentration in human milk. J. Nutr. 2008, 138, 2190–2197. [Google Scholar] [CrossRef] [Green Version]
- Makrides, M.; Neumann, M.; Gibson, R.A. Effect of maternal docosahexaenoic acid (DHA) supplementation on breast milk composition. Eur. J. Clin. Nutr. 1996, 50, 352–357. [Google Scholar]
- O’Neil, A.; Itsiopoulos, C.; Skouteris, H.; Opie, R.S.; McPhie, S.; Hill, B.; Jacka, F.N. Preventing mental health problems in offspring by targeting dietary intake of pregnant women. BMC Med. 2014, 12, 208. [Google Scholar] [CrossRef] [Green Version]
- Greer, F.R.; Sicherer, S.H.; Burks, A.W. Effects of early nutritional interventions on the development of atopic disease in infants and children: The role of maternal dietary restriction, breastfeeding, timing of introduction of complementary foods, and hydrolyzed formulas. Pediatrics 2008, 121, 183–191. [Google Scholar] [CrossRef] [Green Version]
- Hinde, K.; Skibiel, A.L.; Foster, A.B.; Del Rosso, L.; Mendoza, S.P.; Capitanio, J.P. Cortisol in mother’s milk across lactation reflects maternal life history and predicts infant temperament. Behav. Ecol. 2014, 26, 269–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Mean/% | Standard Deviation | Range | Omega-3 | Omega-6 | Omega-6/3 Ratio | Total Fat | |
|---|---|---|---|---|---|---|---|
| Stand. β | Stand. β | Stand. β | Stand. β | ||||
| Maternal Characteristics | |||||||
| Maternal age | 29.67 | 4.86 | 19.2–39.9 | 0.407 t | 0.285 | −0.066 | 0.254 |
| Education | 2.70 | 1.051 | 0−4 | 0.113 | −0.014 | −0.167 | −0.060 |
| Household Income | 69,489 | 33,980 | 25k–105k | 0.008 | −0.186 | −0.286 | −0.181 |
| Pre-Pregnancy BMI | 24.17 | 5.98 | 16.4–47.4 | −0.102 | −0.086 | −0.045 | −0.026 |
| Pregnancy Weight Gain (lbs) | 35.60 | 13.27 | 9.00–71.00 | 0.103 | 0.221 | 0.168 | 0.243 |
| Parity (% Primiparous) | 48.1% | 1−4 | 0.085 | −0.053 | −0.123 | −0.076 | |
| Exclusive Breastfeeding | 63.27% | 0.133 | 0.168 | 0.027 | 0.260 | ||
| % Married | 75% | −0.223 | 0.341 | 0.593 * | 0.223 | ||
| Race/Ethnicity | |||||||
| % White | 58.3% | ||||||
| % Latina | 18.8% | 0.006 | 0.118 | 0.117 | 0.220 | ||
| % Asian | 10.4% | 0.057 | −0.006 | −0.052 | 0.015 | ||
| % Multi-Ethnic/Other | 12.5% | −0.152 | −0.011 | 0.170 | −0.145 | ||
| Infant Characteristics | |||||||
| Birth Weight (grams) | 3465.49 | 386.874 | 2470–4220 | 0.104 | 0.405 | 0.449 t | 0.215 |
| Gestational Age at Birth (weeks) | 39.6822 | 1.12 | 37.1–42.3 | −0.093 | −0.070 | 0.090 | −0.147 |
| BMIP at Birth | 56.92% | 0.30 | 0.34–98% | −0.114 | −0.125 | −0.175 | −0.164 |
| BMIP at 3 mos | 46.25% | 0.27 | 0.02–97% | 0.164 | 0.101 | −0.034 | −0.013 |
| % Female Infants | 48% | 0.117 | 0.181 | 0.035 | 0.120 |
| IUPAC Name | Mean | Standard Deviation | Range | |
|---|---|---|---|---|
| (ug/mL) | (ug/mL) | (ug/mL) | ||
| Omega-3 Fatty Acids | ||||
| Linolenic acid (ALA) | (9Z,12Z,15Z)-octadeca-9,12,15-trienoic acid | 13.80 | 6.15 | 4.26–42.25 |
| Eicosatrienoic acid (ETE) | (11Z,14Z,17Z)-icosa-11,14,17-trienoic acid | 0.74 | 1.02 | 0.02–7.00 |
| Eicosapentaenoic acid (EPA) | (5Z,8Z,11Z,14Z,17Z)-icosa-5,8,11,14,17-pentaenoic acid | 1.09 | 1.16 | 0.06–7.00 |
| Docosahexaenoic acid (DHA) | (4Z,7Z,10Z,13Z,16Z,19Z)-docosa-4,7,10,13,16,19-hexaenoic acid | 2.72 | 1.91 | 0.77–10.71 |
| Omega-6 Fatty Acids | ||||
| Linolelaidic acid | (9E,12E)-octadeca-9,12-dienoic acid | 1.16 | 0.30 | 1.00–3.08 |
| Arachidic acid | (113C)icosanoic acid | 194.60 | 72.78 | 65.12–398.72 |
| Linoleic acid | (9Z,12Z)-octadeca-9,12-dienoic acid | 0.54 | .06 | 0.51–0.89 |
| Linolenic acid | (9Z,12Z,15Z)-octadeca-9,12,15-trienoic acid | 2.26 | 1.71 | 0.14–8.79 |
| Dihomo-gamma-linolenic acid (DGLA) | (8Z,11Z,14Z)-icosa-8,11,14-trienoic acid | 5.41 | 2.21 | 1.41–10.55 |
| Arachidonic acid (AA) | (5Z,8Z,11Z,14Z)-icosa-5,8,11,14-tetraenoic acid | 1.17 | 0.94 | 0.12–4.92 |
| Eicosadienoic Acid | (11E,14E)-icosa-11,14-dienoic acid | 3.72 | 1.78 | 1.04–9.01 |
| Composite Fatty Acid Variables | ||||
| Total Omega-3 | 18.36 | 7.98 | 6.66–55.64 | |
| Total Omega-6 | 208.85 | 77.88 | 70.11–433.86 | |
| Omega-6/Omega-3 Ratio | 12.12 | 4.36 | 4.86–27.14 | |
| Total Milk PUFAs | 227.21 | 83.57 | 83.58−489.50 | |
| Total Milk Fat | 942.51 | 346.76 | 398–2301 |
| Negative Affectivity | Orienting/Regulation | Surgency/Extraversion | |
|---|---|---|---|
| Standardized β (p-Value) | Standardized β (p-Value) | Standardized β (p-Value) | |
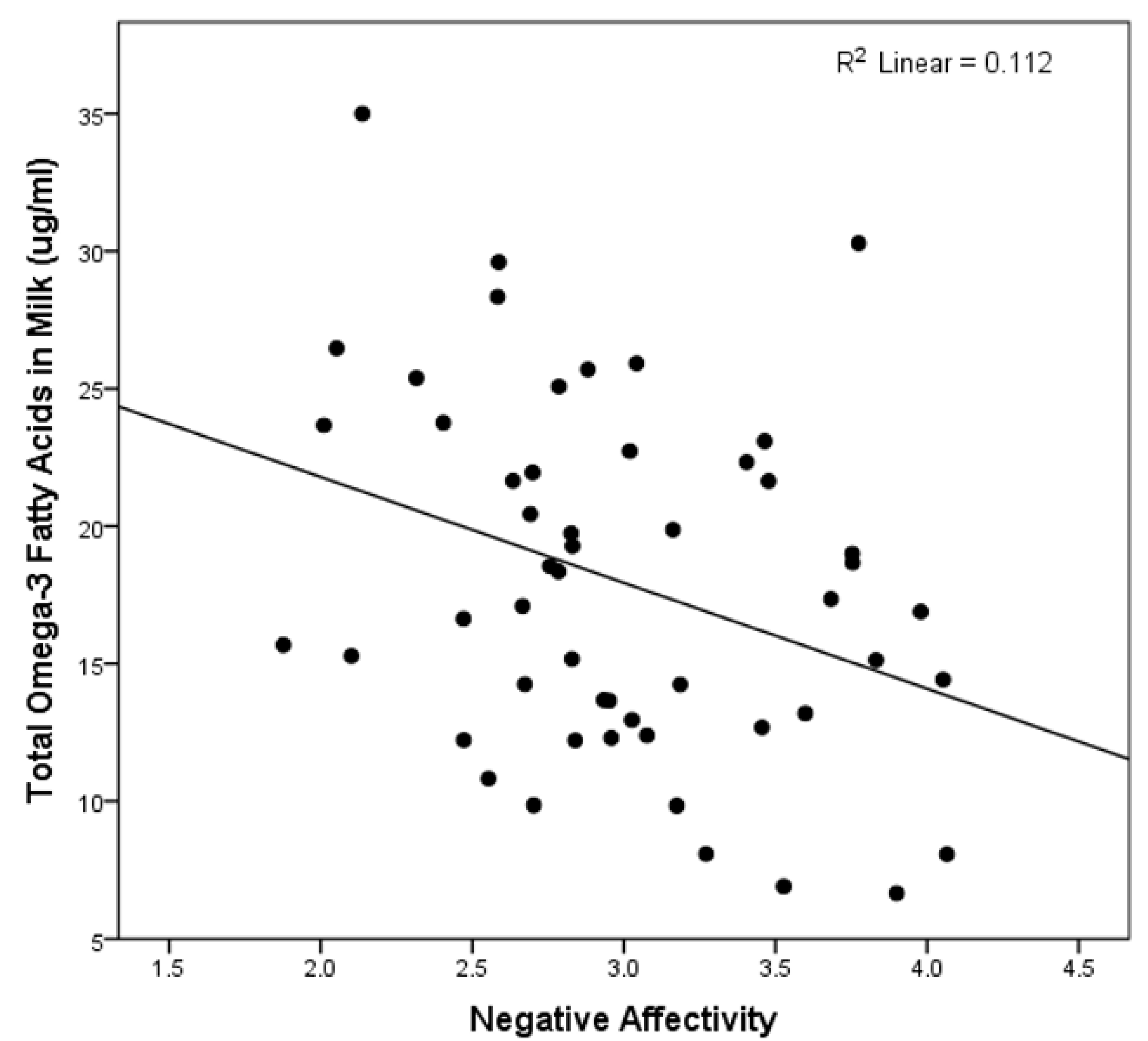
| Omega-3 | −0.352 (0.020) * | −0.014 (0.927) | −0.108 (0.479) |
| Omega-6 | −0.249 (0.106) | 0.131 (0.394) | 0.053 (0.727) |
| Omega-6/3 ratio | 0.134 (0.387) | 0.159 (0.297) | 0.179 (0.236) |
| Total PUFAs | −0.266 (0.083) | 0.127 (0.408) | 0.042 (0.784) |
| Total Milk Fat | −0.124 (0.401) | −0.004 (0.981) | −0.039 (0.791) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hahn-Holbrook, J.; Fish, A.; Glynn, L.M. Human Milk Omega-3 Fatty Acid Composition Is Associated with Infant Temperament. Nutrients 2019, 11, 2964. https://doi.org/10.3390/nu11122964
Hahn-Holbrook J, Fish A, Glynn LM. Human Milk Omega-3 Fatty Acid Composition Is Associated with Infant Temperament. Nutrients. 2019; 11(12):2964. https://doi.org/10.3390/nu11122964
Chicago/Turabian StyleHahn-Holbrook, Jennifer, Adi Fish, and Laura M. Glynn. 2019. "Human Milk Omega-3 Fatty Acid Composition Is Associated with Infant Temperament" Nutrients 11, no. 12: 2964. https://doi.org/10.3390/nu11122964
APA StyleHahn-Holbrook, J., Fish, A., & Glynn, L. M. (2019). Human Milk Omega-3 Fatty Acid Composition Is Associated with Infant Temperament. Nutrients, 11(12), 2964. https://doi.org/10.3390/nu11122964