Safety of Hyaluronan 35 in Healthy Human Subjects: A Pilot Study

 and
and
Abstract
:1. Introduction.
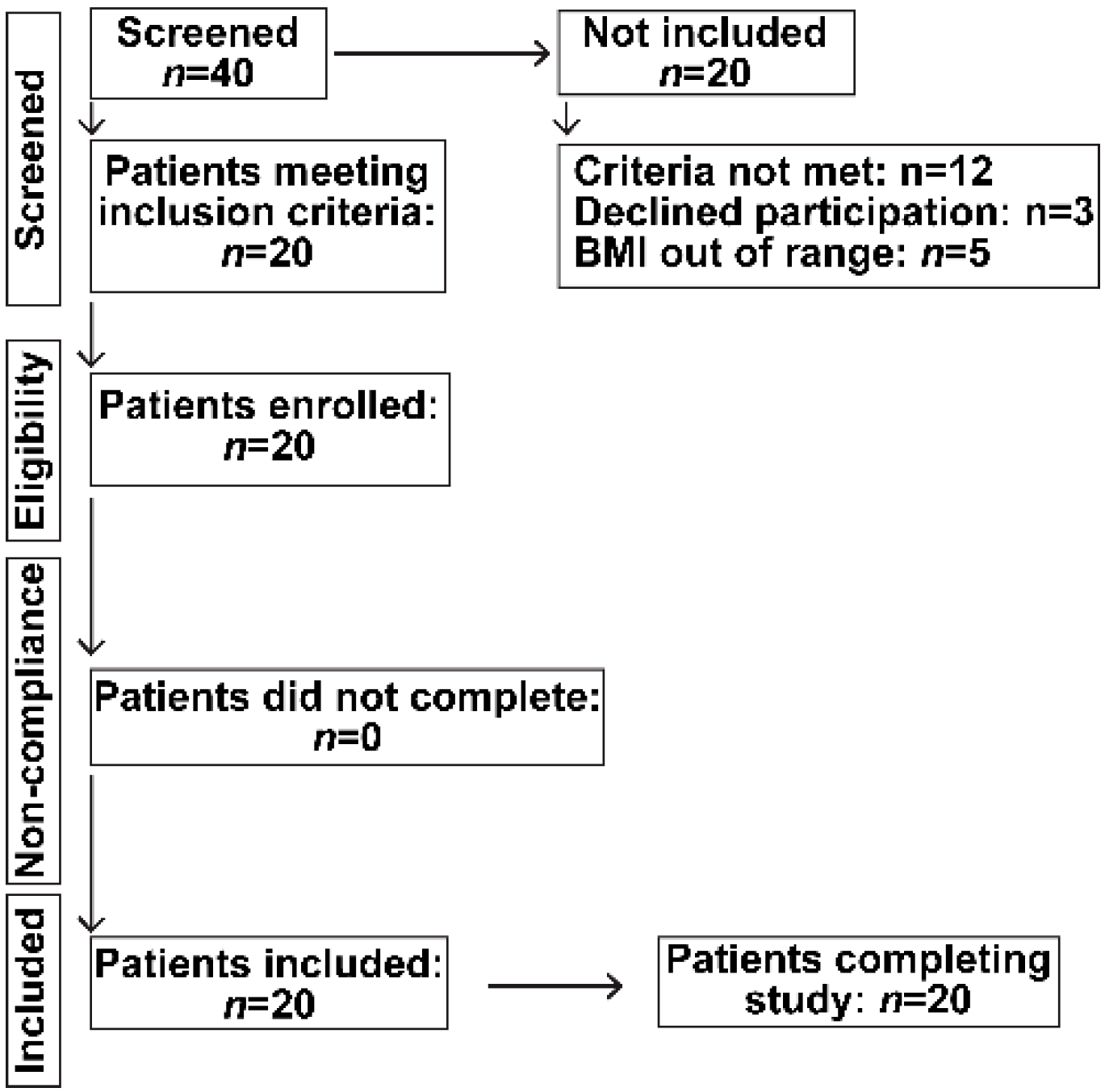
2. Patients and Methods
Study Subjects
3. Methods
3.1. Clinical Assessment
3.2. Clinical Laboratory Tests
3.3. Indirect Calorimetry
3.4. Stool Microbiome Diversity
3.5. Serum Cytokines
3.6. Serum HA Levels
3.7. Intestinal Permeability
3.8. Antimicrobial Peptides in Stool
3.9. Statistical Analyses
4. Results
4.1. Clinical and Demographics
4.2. Clinical Laboratory Parameters
4.3. Indirect Calorimetry Data
4.4. Serum Indicators of Inflammation and Injury
4.5. Fecal Peptides: Calprotectin and β-Defensin 2
4.6. Serum levels of HA and Intestinal Permeability
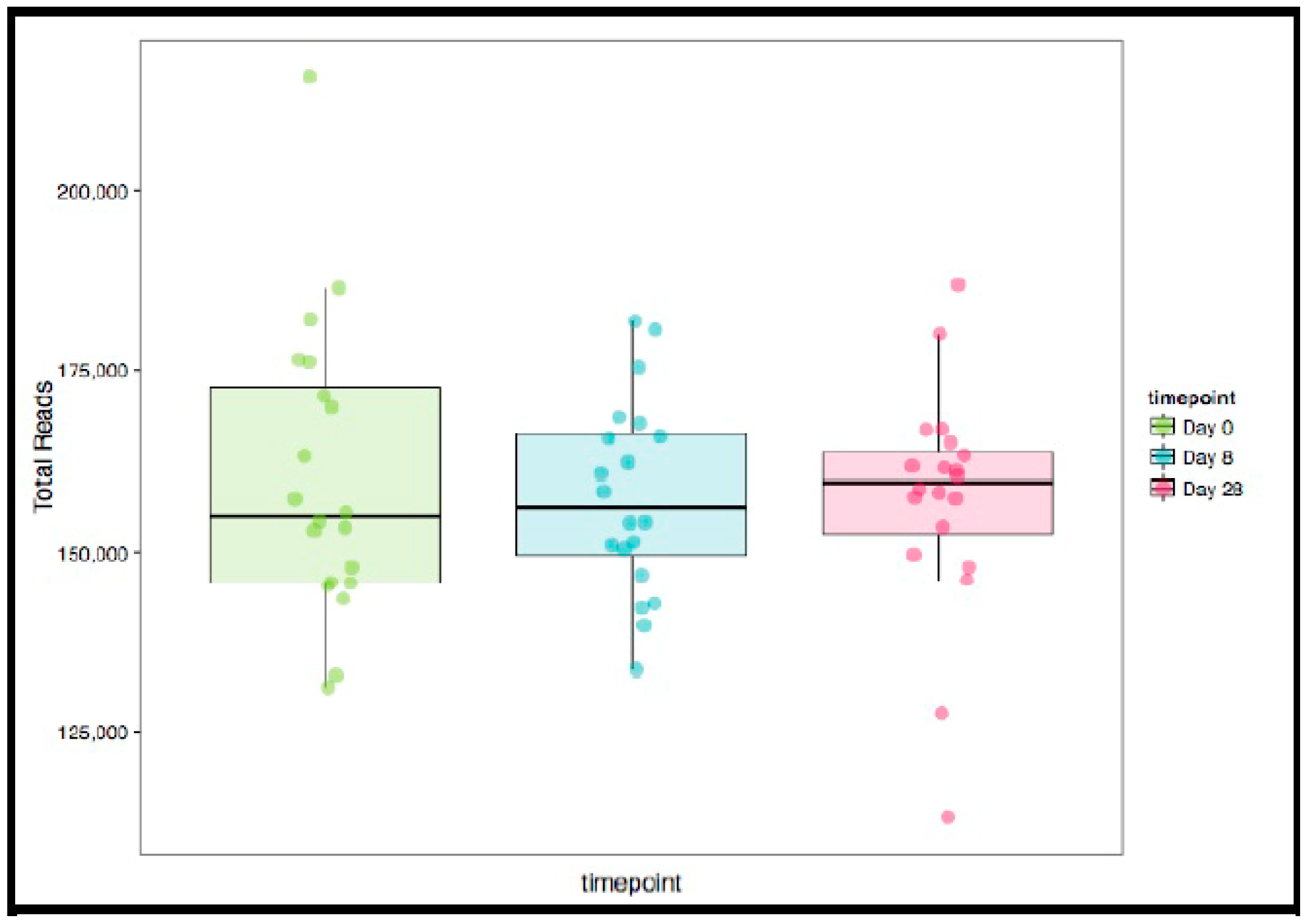
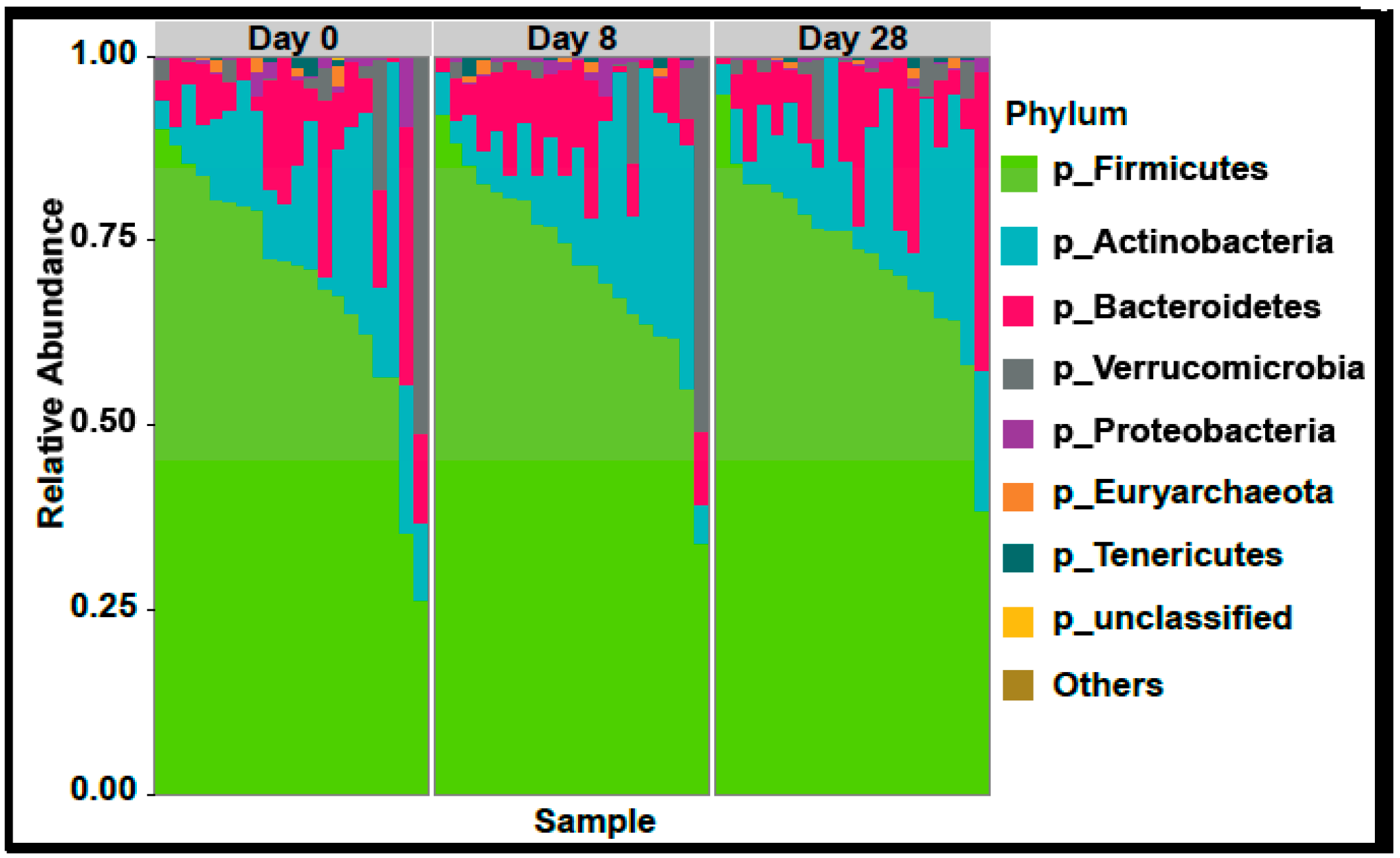
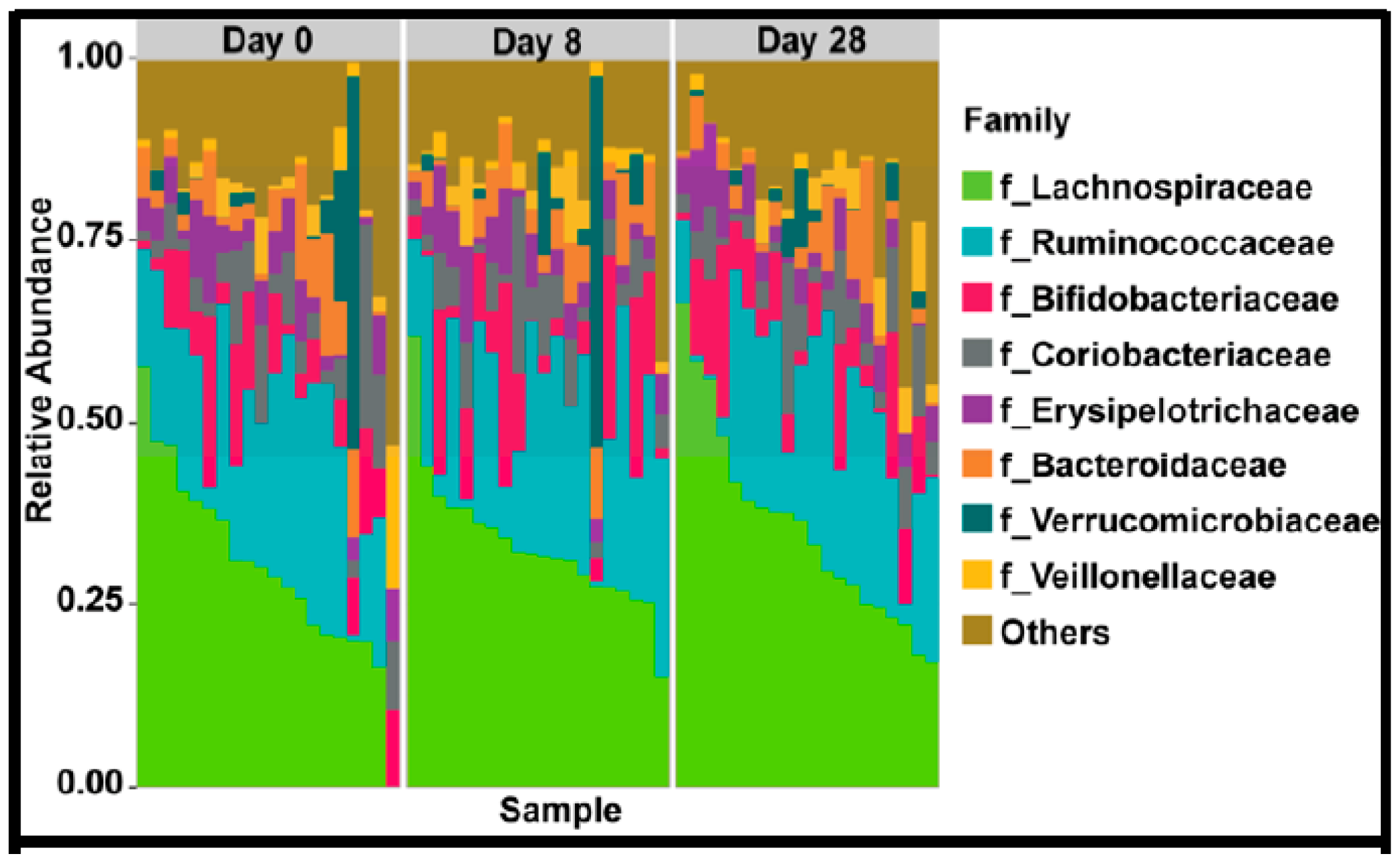
4.7. Gut Microbiome
4.8. GI Symptoms in Response to HA35
5. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Stern, R.; Asari, A.A.; Sugahara, K.N. Hyaluronan fragments: An information-rich system. Eur. J. Cell Biol. 2006, 85, 699–715. [Google Scholar] [CrossRef] [PubMed]
- Weigel, P.H.; Hascall, V.C.; Tammi, M. Hyaluronan synthases. J. Biol. Chem. 1997, 272, 13997–14000. [Google Scholar] [CrossRef] [PubMed]
- Petrey, A.C.; de la Motte, C.A. Hyaluronan, a crucial regulator of inflammation. Front. Immunol. 2014, 5, 101. [Google Scholar] [CrossRef]
- Penna, I.; Albanesi, E.; Bertorelli, R.; Bandiera, T.; Russo, D. Cytoprotective, anti-inflammatory, and antioxidant properties of high-molecular-weight hyaluronan enriched with red orange extract in human fibroblasts exposed to ultra violet light b irradiation. Biotechnol. Appl. Biochem. 2018. [Google Scholar] [CrossRef] [PubMed]
- Cyphert, J.M.; Trempus, C.S.; Garantziotis, S. Size matters: Molecular weight specificity of hyaluronan effects in cell biology. Int. J. Cell Biol. 2015, 2015, 563818. [Google Scholar] [CrossRef] [PubMed]
- Adamia, S.; Maxwell, C.A.; Pilarski, L.M. Hyaluronan and hyaluronan synthases: Potential therapeutic targets in cancer. Curr. Drug Targets Cardiovasc. Haematol. Disord. 2005, 5, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Homann, S.; Grandoch, M.; Kiene, L.S.; Podsvyadek, Y.; Feldmann, K.; Rabausch, B.; Nagy, N.; Lehr, S.; Kretschmer, I.; Oberhuber, A.; et al. Hyaluronan synthase 3 promotes plaque inflammation and atheroprogression. Matrix Biol. 2018, 66, 67–80. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Gao, F.; Han, Z.; Xu, X.; Underhill, C.B.; Zhang, L. Hyaluronan synthase 3 overexpression promotes the growth of tsu prostate cancer cells. Cancer Res. 2001, 61, 5207–5214. [Google Scholar]
- He, Y.; Wu, G.D.; Sadahiro, T.; Noh, S.I.; Wang, H.; Talavera, D.; Wang, H.; Vierling, J.M.; Klein, A.S. Interaction of cd44 and hyaluronic acid enhances biliary epithelial proliferation in cholestatic livers. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 295, G305–G312. [Google Scholar] [CrossRef]
- Wolny, P.M.; Banerji, S.; Gounou, C.; Brisson, A.R.; Day, A.J.; Jackson, D.G.; Richter, R.P. Analysis of cd44-hyaluronan interactions in an artificial membrane system: Insights into the distinct binding properties of high and low molecular weight hyaluronan. J. Biol. Chem. 2010, 285, 30170–30180. [Google Scholar] [CrossRef]
- Jiang, D.; Liang, J.; Fan, J.; Yu, S.; Chen, S.; Luo, Y.; Prestwich, G.D.; Mascarenhas, M.M.; Garg, H.G.; Quinn, D.A.; et al. Regulation of lung injury and repair by toll-like receptors and hyaluronan. Nat. Med. 2005, 11, 1173–1179. [Google Scholar] [CrossRef]
- Bono, P.; Rubin, K.; Higgins, J.M.; Hynes, R.O. Layilin, a novel integral membrane protein, is a hyaluronan receptor. Mol. Biol. Cell 2001, 12, 891–900. [Google Scholar] [CrossRef]
- Forteza, R.M.; Casalino-Matsuda, S.M.; Falcon, N.S.; Valencia Gattas, M.; Monzon, M.E. Hyaluronan and layilin mediate loss of airway epithelial barrier function induced by cigarette smoke by decreasing e-cadherin. J. Biol. Chem. 2012, 287, 42288–42298. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Kessler, S.P.; Obery, D.R.; Homer, C.R.; McDonald, C.; de la Motte, C.A. Hyaluronan 35kda treatment protects mice from citrobacter rodentium infection and induces epithelial tight junction protein zo-1 in vivo. Matrix Biol. 2017, 62, 28–39. [Google Scholar] [CrossRef]
- Saikia, P.; Roychowdhury, S.; Bellos, D.; Pollard, K.A.; McMullen, M.R.; McCullough, R.L.; McCullough, A.J.; Gholam, P.; de la Motte, C.; Nagy, L.E. Hyaluronic acid 35 normalizes tlr4 signaling in kupffer cells from ethanol-fed rats via regulation of microrna291b and its target tollip. Sci. Rep. 2017, 7, 15671. [Google Scholar] [CrossRef]
- Kim, Y.; West, G.A.; Ray, G.; Kessler, S.P.; Petrey, A.C.; Fiocchi, C.; McDonald, C.; Longworth, M.S.; Nagy, L.E.; de la Motte, C.A. Layilin is critical for mediating hyaluronan 35kda-induced intestinal epithelial tight junction protein zo-1 in vitro and in vivo. Matrix Biol. 2018, 66, 93–109. [Google Scholar] [CrossRef] [PubMed]
- Saikia, P.; Bellos, D.; McMullen, M.R.; Pollard, K.A.; de la Motte, C.; Nagy, L.E. Microrna 181b-3p and its target importin alpha5 regulate toll-like receptor 4 signaling in kupffer cells and liver injury in mice in response to ethanol. Hepatology 2017, 66, 602–615. [Google Scholar] [CrossRef] [PubMed]
- Kessler, S.P.; Obery, D.R.; Nickerson, K.P.; Petrey, A.C.; McDonald, C.; de la Motte, C.A. Multifunctional role of 35 kilodalton hyaluronan in promoting defense of the intestinal epithelium. J. Histochem. Cytochem. 2018, 66, 273–287. [Google Scholar] [CrossRef]
- Tibble, J.A.; Bjarnason, I. Fecal calprotectin as an index of intestinal inflammation. Drugs Today (Barc) 2001, 37, 85–96. [Google Scholar] [CrossRef]
- Robinson, K.; Deng, Z.; Hou, Y.; Zhang, G. Regulation of the intestinal barrier function by host defense peptides. Front Vet. Sci. 2015, 2, 57. [Google Scholar] [CrossRef]
- Diet*calc Analysis Program, Version 1.5.0. Available online: https://epi.grants.cancer (accessed on 21 May 2019).
- Glass, C.; Hipskind, P.; Tsien, C.; Malin, S.K.; Kasumov, T.; Shah, S.N.; Kirwan, J.P.; Dasarathy, S. Sarcopenia and a physiologically low respiratory quotient in patients with cirrhosis: A prospective controlled study. J. Appl. Physiol. (1985) 2013, 114, 559–565. [Google Scholar] [CrossRef]
- Kaczmarek, J.L.; Musaad, S.M.; Holscher, H.D. Time of day and eating behaviors are associated with the composition and function of the human gastrointestinal microbiota. Am. J. Clin. Nutr. 2017, 106, 1220–1231. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and clustering orders of magnitude faster than blast. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef]
- Edgar, R.C. Uparse: Highly accurate otu sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive bayesian classifier for rapid assignment of rrna sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef]
- DeSantis, T.Z.; Hugenholtz, P.; Larsen, N.; Rojas, M.; Brodie, E.L.; Keller, K.; Huber, T.; Dalevi, D.; Hu, P.; Andersen, G.L. Greengenes, a chimera-checked 16s rrna gene database and workbench compatible with arb. Appl. Environ. Microbiol. 2006, 72, 5069–5072. [Google Scholar] [CrossRef]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer Publishing Company, Incorporated: Salmon Tower Building, NY, USA, 2009; p. 216. [Google Scholar]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for rna-seq data with deseq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- McMurdie, P.J.; Holmes, S. Waste not, want not: Why rarefying microbiome data is inadmissible. PLoS Comput. Biol. 2014, 10, e1003531. [Google Scholar] [CrossRef]
- Gudowska, M.; Gruszewska, E.; Panasiuk, A.; Cylwik, B.; Flisiak, R.; Swiderska, M.; Szmitkowski, M.; Chrostek, L. Hyaluronic acid concentration in liver diseases. Clin. Exp. Med. 2016, 16, 523–528. [Google Scholar] [CrossRef]
- Tashiro, T.; Seino, S.; Sato, T.; Matsuoka, R.; Masuda, Y.; Fukui, N. Oral administration of polymer hyaluronic acid alleviates symptoms of knee osteoarthritis: A double-blind, placebo-controlled study over a 12-month period. Sci. World J. 2012, 2012, 167928. [Google Scholar] [CrossRef]
- Oe, M.; Tashiro, T.; Yoshida, H.; Nishiyama, H.; Masuda, Y.; Maruyama, K.; Koikeda, T.; Maruya, R.; Fukui, N. Oral hyaluronan relieves knee pain: A review. Nutr. J. 2016, 15, 11. [Google Scholar] [CrossRef] [PubMed]
- Faix, J.D. Biomarkers of sepsis. Crit. Rev. Clin. Lab. Sci. 2013, 50, 23–36. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Anthropometric Data | Day 0 | Day 8 | Day 28 |
|---|---|---|---|
| Number | 20 | 20 | 20 |
| Height (meters) | 1.7 ± 0.1 (1.5–1.9) | 1.7 ± 0.1 (1.5–1.9) | 1.7 ± 0.1 (1.5–1.9) |
| Weight (kilograms) | 79.0 ± 16.3 | 78.7 ± 16.4 | 78.9 ± 16.9 |
| SBP (mmHg) | 120.9 ± 15.3 | 121.3 ± 11.0 | 120.5 ± 11.8 |
| DBP (mmHg) | 74.4 ± 10.6 | 70.6 ± 11 | 71.5 ± 10.3 |
| BMI (kg/m2) | 27.4 ± 5.4 | 27.2 ± 5.5 | 27.2 ± 5.7 |
| HR (beats per minute) | 76.2 ± 13.6 | 73.9 ± 11.1 | 75.7 ± 10.2 |
| Parameter | Day 0 | Day 8 | Day 28 |
|---|---|---|---|
| Number | 20 | 20 | 20 |
| Leukocyte count k/μL (3.7–11) | 6.0 ± 1.6 | 6.1 ± 1.4 | 6.2 ± 1.5 |
| Hemoglobin g/dL (13–17) | 13.4 ± 1.5 | 13.4 ± 1.4 | 13.2 ± 1.5 |
| Platelet Count k/μL (150–400) | 263 ± 59 | 262 ± 63 | 271 ± 64 |
| Protein g/dL (6.0–8.4) | 7.4 ± 0.3 | 7.3 ± 0.4 | 7.3 ± 0.4 |
| Albumin g/dL (3.5–5.0) | 4.4 ± 0.3 | 4.4 ± 0.3 | 4.3 ± 0.3 |
| Plasma calcium mg/dL (8.5–10.5) | 9.2 ± 0.4 | 9.2 ± 0.2 | 9.1 ± 0.3 |
| Total bilirubin mg/dL (0.0–1.5) | 0.4 ± 0.3 | 0.4 ± 0.2 | 0.4 ± 0.3 |
| Alkaline phosphatase U/L (40–150) | 66 ± 16 | 66 ± 17 | 67 ± 17 |
| AST U/L (7–40) | 18 ± 6 | 22 ± 18 | 19 ± 7 |
| ALT U/L (5–50) | 16 ± 7 | 15 ± 6 | 18 ± 17 |
| Glucose mg/dL (60–100) | 92 ± 20 | 94 ± 16 | 89 ± 27 |
| BUN mg/dL (10–25) | 11 ± 3 | 11 ± 4 | 12 ± 3 |
| Creatinine mg/dL (70–1.40) | 0.85 ± 0.16 | 0.85 ± 0.16 | 0.83 ± 0.15 |
| Sodium mmol/L (135–146) | 140 ± 1 | 140 ± 2 | 140 ± 2 |
| Potassium mmol/L (3.5–5.0) | 4.0 ± 0.2 | 4.1 ± 0.3 | 4.0 ± 0.2 |
| Bicarbonate mmol/L (23–32) | 25 ± 2 | 24 ± 2 | 24 ± 2 |
| Measurement | Day 0 | Day 8 | Day 28 |
|---|---|---|---|
| Number | 20 | 20 | 20 |
| V02 (L/min) | 0.23 ± 0.04 | 0.22 ± 0.04 | 0.24 ± 0.05 |
| VC02 (L/min) | 0.18 ± 0.03 | 0.18 ± 0.04 | 0.20 ± 0.04 |
| Respiratory quotient | 0.81 ± 0.08 | 0.82 ± 0.06 | 0.83 ± 0.071 |
| Measured REE (Kcal/day) | 1556.3 ± 274.5 | 1562.8 ± 221.4 | 1670.6 ± 333.9 |
| Metabolic Rate (Kcal/day) | 1653.1 ± 268.7 | 1665.2 ± 258.8 | 1668.4 ± 259.8 |
| REE (% predicted) | 95.9 ± 9.5 | 95.7 ± 12.9 | 101.4 ± 10.4 |
| Protein | Day 0 | Day 8 | Day 28 |
|---|---|---|---|
| Number | 20 | 20 | 20 |
| TNF alpha (pg/mL) | 2.0 ± 5.1 | 2.4 ± 6.6 | 3.2 ± 6.9 |
| IL-6 (pg/mL) | 9.6 ± 14.6 | 11.7 ± 16.8 | 10.5 ± 14.6 |
| C- Reactive Protein (g/dL) | 10.2 ± 15.5 | 10.3 ±12.8 | 11.6 ± 13.7 |
| Fecal Proteins | Day 0 | Day 8 | Day 28 |
|---|---|---|---|
| Number | 20 | 20 | 20 |
| Total soluble fecal protein (μg/mL) | 22,883.8 ± 6425.7 | 22,365.3 ± 5872.8 | 22,726.3 ± 6305.2 |
| Calprotectin (ng/mL) | 139.3 ± 89.1 | 177.4 ± 133.1 | 195.00 ± 117.0 |
| Normalized calprotectin (ng/μg protein) | 0.006 ± 0.003 | 0.008 ± 0.006 | 0.009 ± 0.006 |
| Human β-defensin 2 (ng/mL) | 74.13 ± 109.0 | 92.06 ± 168.9 | 67.48 ± 116.5 |
| Normalized β-defensin 2 (ng/μg protein) | 0.004 ± 0.007 | 0.005 ± 0.0105 | 0.004 ± 0.008 |
| Measurement | Day 0 | Day 8 | Day 28 |
|---|---|---|---|
| Number | 20 | 20 | 20 |
| Serum HA (ng/mL) | 338.8 ± 82.8 | 329.0 ± 106.6 | 338.7 ± 73.9 |
| LPS (ng/mL) | 0.41 ± 1.35 | 2.68 ± 5.96 | 1.26 ± 4.03 |
| Symptom | Number of Patients Experiencing Symptoms (%) | Severity of Symptoms (%) | Frequency of Symptoms (%) |
|---|---|---|---|
| Bloating | 3 (15) | None: 17 (85) | 3 (15) several hours |
| Mild: 2 (10) | |||
| Moderate 1 (5) | |||
| Cramping | 8 (40) | None: 12 (60) | 8 (40) several hours |
| Mild: 2 (10) | |||
| Moderate 6 (30) | |||
| Nausea | 1 (5) | None: 19 (95) | 1 (5) several hours |
| Severe: 1 (5) | |||
| Increased Hunger | 1 (5) | None: 19 (95) | 1 (5) several hours |
| Mild: 1 (5) | |||
| Flatulence | 1 (5) | None: 19 (95) | 1 (5) more than one day |
| Severe: 1 (5) | |||
| Abdominal Pain | 1 (5) | None: 19 (95) | 1 (5) several hours |
| Moderate: 1 (5) | |||
| Constipation | 1 (5) | None: 19 (95) | 1 (5) several hours |
| Moderate: 1 (5) | |||
| Diarrhea | 1 (5) | None: 19 (95) | 1 (5) several hours |
| Moderate: 1 (5) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bellar, A.; Kessler, S.P.; Obery, D.R.; Sangwan, N.; Welch, N.; Nagy, L.E.; Dasarathy, S.; de la Motte, C. Safety of Hyaluronan 35 in Healthy Human Subjects: A Pilot Study. Nutrients 2019, 11, 1135. https://doi.org/10.3390/nu11051135
Bellar A, Kessler SP, Obery DR, Sangwan N, Welch N, Nagy LE, Dasarathy S, de la Motte C. Safety of Hyaluronan 35 in Healthy Human Subjects: A Pilot Study. Nutrients. 2019; 11(5):1135. https://doi.org/10.3390/nu11051135
Chicago/Turabian StyleBellar, Annette, Sean P. Kessler, Dana R. Obery, Naseer Sangwan, Nicole Welch, Laura E. Nagy, Srinivasan Dasarathy, and Carol de la Motte. 2019. "Safety of Hyaluronan 35 in Healthy Human Subjects: A Pilot Study" Nutrients 11, no. 5: 1135. https://doi.org/10.3390/nu11051135
APA StyleBellar, A., Kessler, S. P., Obery, D. R., Sangwan, N., Welch, N., Nagy, L. E., Dasarathy, S., & de la Motte, C. (2019). Safety of Hyaluronan 35 in Healthy Human Subjects: A Pilot Study. Nutrients, 11(5), 1135. https://doi.org/10.3390/nu11051135