Gastrointestinal Sensing of Meal-Related Signals in Humans, and Dysregulations in Eating-Related Disorders



Abstract
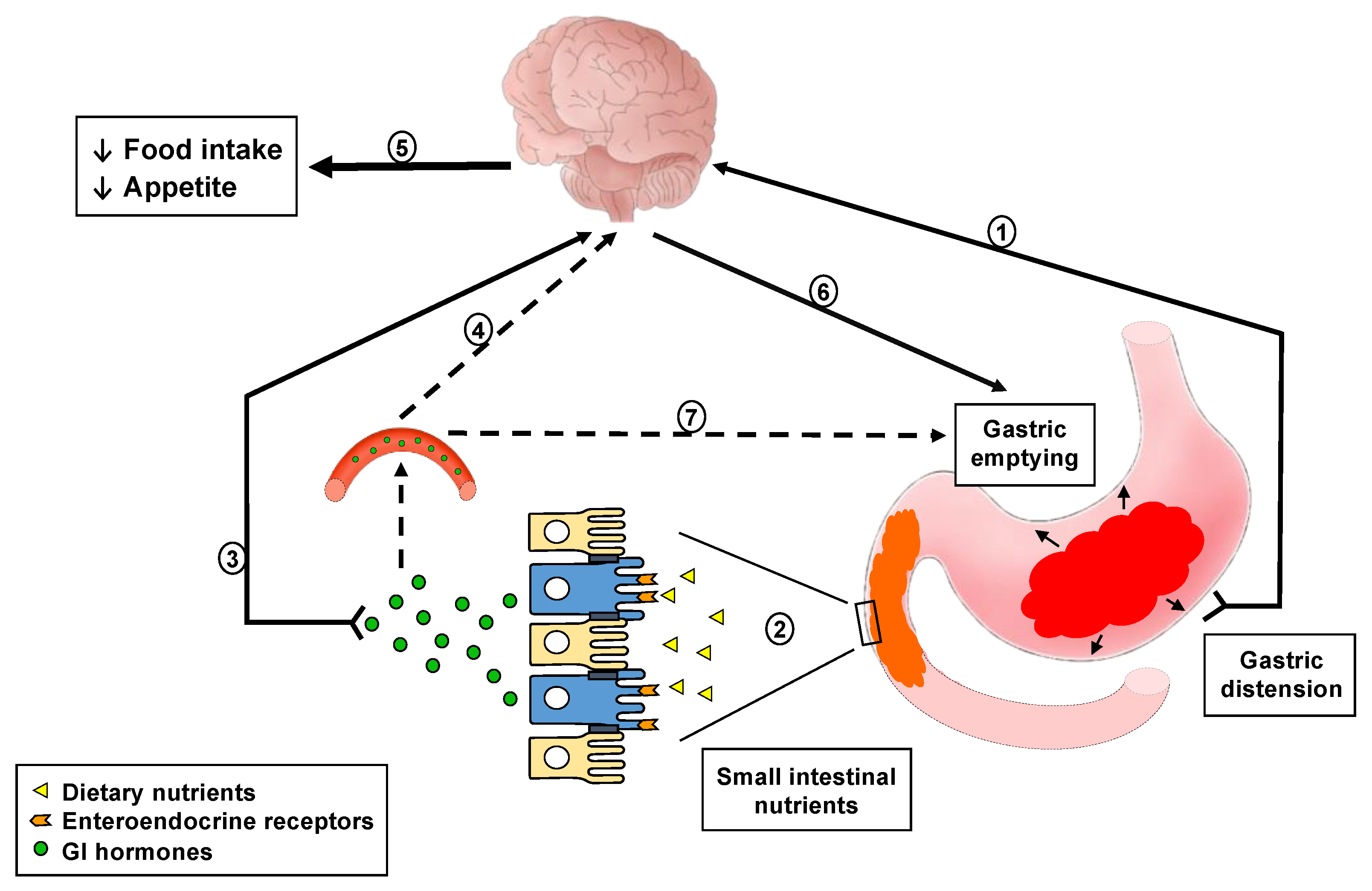
:1. Introduction
2. GI Sensing of Intraluminal Meal-Related Stimuli
2.1. Sensing of Gastric Distension
2.2. Effects of Nutrients in the Small Intestinal Lumen
2.2.1. Small Intestinal Sensing of Lipid
2.2.2. Small Intestinal Sensing of Protein
3. Altered GI Sensing of Meal-Related Stimuli in Eating-Related Disorders
3.1. Altered GI Sensing in Obesity
3.1.1. Sensitivity to Gastric Distension
3.1.2. Small Intestinal Sensing of Fat
3.1.3. Small Intestinal Sensing of Protein
3.2. Altered GI Sensing in Functional Dyspepsia
3.2.1. Sensitivity to Gastric Distension
3.2.2. Alterations in the Small Intestinal Sensing of Nutrients
3.3. Altered GI Sensing in Anorexia of Ageing
3.3.1. Sensitivity to Gastric Distension
3.3.2. Alterations in the Small Intestinal Sensing of Nutrients
4. Summary and Future Directions
Author Contributions
Funding
Conflicts of Interest
References
- Vella, A.; Camilleri, M. The gastrointestinal tract as an integrator of mechanical and hormonal response to nutrient ingestion. Diabetes 2017, 66, 2729–2737. [Google Scholar] [CrossRef]
- Latorre, R.; Sternini, C.; De Giorgio, R.; Greenwood-Van Meerveld, B. Enteroendocrine cells: A review of their role in brain-gut communication. Neurogastroenterol. Motil. 2016, 28, 620–630. [Google Scholar] [CrossRef]
- Depoortere, I. Taste receptors of the gut: Emerging roles in health and disease. Gut 2014, 63, 179–190. [Google Scholar] [CrossRef] [PubMed]
- De Lartigue, G.; Diepenbroek, C. Novel developments in vagal afferent nutrient sensing and its role in energy homeostasis. Curr. Opin. Pharmacol. 2016, 31, 38–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grundy, D. Signalling the state of the digestive tract. Auton. Neurosci. 2006, 125, 76–80. [Google Scholar] [CrossRef] [PubMed]
- Furness, J.B.; Rivera, L.R.; Cho, H.J.; Bravo, D.M.; Callaghan, B. The gut as a sensory organ. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 729–740. [Google Scholar] [CrossRef] [PubMed]
- Brookes, S.J.; Spencer, N.J.; Costa, M.; Zagorodnyuk, V.P. Extrinsic primary afferent signalling in the gut. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 286–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feinle, C.; Grundy, D.; Read, N.W. Effects of duodenal nutrients on sensory and motor responses of the human stomach to distension. Am. J. Physiol. 1997, 273, G721–G726. [Google Scholar] [CrossRef]
- Ezcurra, M.; Reimann, F.; Gribble, F.M.; Emery, E. Molecular mechanisms of incretin hormone secretion. Curr. Opin. Pharmacol. 2013, 13, 922–927. [Google Scholar] [CrossRef] [Green Version]
- Steinert, R.E.; Feinle-Bisset, C.; Asarian, L.; Horowitz, M.; Beglinger, C.; Geary, N. Ghrelin, CCK, GLP-1, and PYY(3-36): Secretory controls and physiological roles in eating and glycemia in health, obesity, and after RYGB. Physiol. Rev. 2017, 97, 411–463. [Google Scholar] [CrossRef]
- Enck, P.; Azpiroz, F.; Boeckxstaens, G.; Elsenbruch, S.; Feinle-Bisset, C.; Holtmann, G.; Lackner, J.M.; Ronkainen, J.; Schemann, M.; Stengel, A.; et al. Functional dyspepsia. Nat. Rev. Dis. Primers 2017, 3, 17081. [Google Scholar] [CrossRef] [PubMed]
- Deane, A.; Chapman, M.J.; Fraser, R.J.; Horowitz, M. Bench-to-bedside review: The gut as an endocrine organ in the critically ill. Crit. Care 2010, 14, 228. [Google Scholar] [CrossRef] [PubMed]
- Monteleone, P.; Castaldo, E.; Maj, M. Neuroendocrine dysregulation of food intake in eating disorders. Regul. Pept. 2008, 149, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Jayasena, C.N.; Bloom, S.R. Obesity and appetite control. Exp. Diabetes. Res. 2012, 2012, 824305. [Google Scholar] [CrossRef] [PubMed]
- Little, T.J.; Feinle-Bisset, C. Effects of dietary fat on appetite and energy intake in health and obesity--oral and gastrointestinal sensory contributions. Physiol. Behav. 2011, 104, 613–620. [Google Scholar] [CrossRef]
- Hetterich, L.; Mack, I.; Giel, K.E.; Zipfel, S.; Stengel, A. An update on gastrointestinal disturbances in eating disorders. Mol. Cell. Endocrinol. 2018. [Google Scholar] [CrossRef]
- Tong, J.; D’Alessio, D. Eating disorders and gastrointestinal peptides. Curr. Opin. Endocrinol. Diabetes Obes. 2011, 18, 42–49. [Google Scholar] [CrossRef]
- Feinle-Bisset, C.; Azpiroz, F. Dietary lipids and functional gastrointestinal disorders. Am. J. Gastroenterol. 2013, 108, 737–747. [Google Scholar] [CrossRef]
- Chapman, M.J.; Deane, A.M. Gastrointestinal dysfunction relating to the provision of nutrition in the critically ill. Curr. Opin. Clin. Nutr. Metab. Care 2015, 18, 207–212. [Google Scholar] [CrossRef]
- Parker, B.A.; Chapman, I.M. Food intake and ageing—The role of the gut. Mech. Ageing. Dev. 2004, 125, 859–866. [Google Scholar] [CrossRef]
- Kreuch, D.; Keating, D.J.; Wu, T.; Horowitz, M.; Rayner, C.K.; Young, R.L. Gut mechanisms linking intestinal sweet sensing to glycemic control. Front. Endocrinol. 2018, 9, 741. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.J.; Tomasi, D.; Backus, W.; Wang, R.; Telang, F.; Geliebter, A.; Korner, J.; Bauman, A.; Fowler, J.S.; Thanos, P.K.; et al. Gastric distention activates satiety circuitry in the human brain. Neuroimage 2008, 39, 1824–1831. [Google Scholar] [CrossRef] [PubMed]
- Distrutti, E.; Azpiroz, F.; Soldevilla, A.; Malagelada, J.R. Gastric wall tension determines perception of gastric distention. Gastroenterology 1999, 116, 1035–1042. [Google Scholar] [CrossRef]
- Kissileff, H.R.; Carretta, J.C.; Geliebter, A.; Pi-Sunyer, F.X. Cholecystokinin and stomach distension combine to reduce food intake in humans. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2003, 285, R992–R998. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.L.; Doran, S.M.; Hveem, K.; Bartholomeusz, F.D.; Morley, J.E.; Sun, W.M.; Chatterton, B.E.; Horowitz, M. Relation between postprandial satiation and antral area in normal subjects. Am. J. Clin. Nutr. 1997, 66, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Sturm, K.; Parker, B.; Wishart, J.; Feinle-Bisset, C.; Jones, K.L.; Chapman, I.; Horowitz, M. Energy intake and appetite are related to antral area in healthy young and older subjects. Am. J. Clin. Nutr. 2004, 80, 656–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heddle, R.; Collins, P.J.; Dent, J.; Horowitz, M.; Read, N.W.; Chatterton, B.; Houghton, L.A. Motor mechanisms associated with slowing of the gastric emptying of a solid meal by an intraduodenal lipid infusion. J. Gastroenterol. Hepatol. 1989, 4, 437–447. [Google Scholar] [CrossRef] [PubMed]
- Pilichiewicz, A.N.; Papadopoulos, P.; Brennan, I.M.; Little, T.J.; Meyer, J.H.; Wishart, J.M.; Horowitz, M.; Feinle-Bisset, C. Load-dependent effects of duodenal lipid on antropyloroduodenal motility, plasma CCK and PYY, and energy intake in healthy men. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 293, R2170–R2178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan, A.T.; Feinle-Bisset, C.; Kallas, A.; Wishart, J.M.; Clifton, P.M.; Horowitz, M.; Luscombe-Marsh, N.D. Intraduodenal protein modulates antropyloroduodenal motility, hormone release, glycemia, appetite, and energy intake in lean men. Am. J. Clin. Nutr. 2012, 96, 474–482. [Google Scholar] [CrossRef] [Green Version]
- Schober, G.; Lange, K.; Steinert, R.E.; Hutchison, A.T.; Luscombe-Marsh, N.D.; Landrock, M.F.; Horowitz, M.; Seimon, R.V.; Feinle-Bisset, C. Contributions of upper gut hormones and motility to the energy intake-suppressant effects of intraduodenal nutrients in healthy, lean men—A pooled-data analysis. Physiol. Rep. 2016, 4, e12943. [Google Scholar] [CrossRef]
- Seimon, R.V.; Lange, K.; Little, T.J.; Brennan, I.M.; Pilichiewicz, A.N.; Feltrin, K.L.; Smeets, A.J.; Horowitz, M.; Feinle-Bisset, C. Pooled-data analysis identifies pyloric pressures and plasma cholecystokinin concentrations as major determinants of acute energy intake in healthy, lean men. Am. J. Clin. Nutr. 2010, 92, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Ryan, A.T.; Luscombe-Marsh, N.D.; Saies, A.A.; Little, T.J.; Standfield, S.; Horowitz, M.; Feinle-Bisset, C. Effects of intraduodenal lipid and protein on gut motility and hormone release, glycemia, appetite, and energy intake in lean men. Am. J. Clin. Nutr. 2013, 98, 300–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feinle, C.; O’Donovan, D.; Doran, S.; Andrews, J.M.; Wishart, J.; Chapman, I.; Horowitz, M. Effects of fat digestion on appetite, APD motility, and gut hormones in response to duodenal fat infusion in humans. Am. J. Physiol. Gastrointest. Liver Physiol. 2003, 284, G798–G807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beglinger, S.; Drewe, J.; Schirra, J.; Goke, B.; D’Amato, M.; Beglinger, C. Role of fat hydrolysis in regulating glucagon-like peptide-1 secretion. J. Clin. Endocrinol. Metab. 2010, 95, 879–886. [Google Scholar] [CrossRef] [PubMed]
- Hunt, J.N.; Knox, M.T. A relation between the chain length of fatty acids and the slowing of gastric emptying. J. Physiol. 1968, 194, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Feltrin, K.L.; Little, T.J.; Meyer, J.H.; Horowitz, M.; Rades, T.; Wishart, J.; Feinle-Bisset, C. Comparative effects of intraduodenal infusions of lauric and oleic acids on antropyloroduodenal motility, plasma cholecystokinin and peptide YY, appetite, and energy intake in healthy men. Am. J. Clin. Nutr. 2008, 87, 1181–1187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matzinger, D.; Degen, L.; Drewe, J.; Meuli, J.; Duebendorfer, R.; Ruckstuhl, N.; D’Amato, M.; Rovati, L.; Beglinger, C. The role of long chain fatty acids in regulating food intake and cholecystokinin release in humans. Gut 2000, 46, 688–693. [Google Scholar] [CrossRef]
- Miyauchi, S.; Hirasawa, A.; Ichimura, A.; Hara, T.; Tsujimoto, G. New frontiers in gut nutrient sensor research: Free fatty acid sensing in the gastrointestinal tract. J. Pharmacol. Sci. 2010, 112, 19–24. [Google Scholar] [CrossRef]
- Cvijanovic, N.; Isaacs, N.J.; Rayner, C.K.; Feinle-Bisset, C.; Young, R.L.; Little, T.J. Lipid stimulation of fatty acid sensors in the human duodenum: Relationship with gastrointestinal hormones, BMI and diet. Int. J. Obes. 2017, 41, 233–239. [Google Scholar] [CrossRef]
- Feinle, C.; D’Amato, M.; Read, N.W. Cholecystokinin-A receptors modulate gastric sensory and motor responses to gastric distension and duodenal lipid. Gastroenterology 1996, 110, 1379–1385. [Google Scholar] [CrossRef]
- Lassman, D.J.; McKie, S.; Gregory, L.J.; Lal, S.; D’Amato, M.; Steele, I.; Varro, A.; Dockray, G.J.; Williams, S.C.; Thompson, D.G. Defining the role of cholecystokinin in the lipid-induced human brain activation matrix. Gastroenterology 2010, 138, 1514–1524. [Google Scholar] [CrossRef] [PubMed]
- Brennan, I.M.; Luscombe-Marsh, N.D.; Seimon, R.V.; Otto, B.; Horowitz, M.; Wishart, J.M.; Feinle-Bisset, C. Effects of fat, protein, and carbohydrate and protein load on appetite, plasma cholecystokinin, peptide YY, and ghrelin, and energy intake in lean and obese men. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 303, G129–G140. [Google Scholar] [CrossRef] [PubMed]
- Batterham, R.L.; Heffron, H.; Kapoor, S.; Chivers, J.E.; Chandarana, K.; Herzog, H.; Le Roux, C.W.; Thomas, E.L.; Bell, J.D.; Withers, D.J. Critical role for peptide YY in protein-mediated satiation and body-weight regulation. Cell. Metab. 2006, 4, 223–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, G.H.; Tecimer, S.N.; Shah, D.; Zafar, T.A. Protein source, quantity, and time of consumption determine the effect of proteins on short-term food intake in young men. J. Nutr. 2004, 134, 3011–3015. [Google Scholar] [CrossRef] [PubMed]
- Giezenaar, C.; Luscombe-Marsh, N.D.; Hutchison, A.T.; Standfield, S.; Feinle-Bisset, C.; Horowitz, M.; Chapman, I.; Soenen, S. Dose-dependent effects of randomized intraduodenal whey-protein loads on glucose, gut hormone, and amino acid concentrations in healthy older and younger men. Nutrients 2018, 10, 78. [Google Scholar] [CrossRef] [PubMed]
- Hall, W.L.; Millward, D.J.; Long, S.J.; Morgan, L.M. Casein and whey exert different effects on plasma amino acid profiles, gastrointestinal hormone secretion and appetite. Br. J. Nutr. 2003, 89, 239–248. [Google Scholar] [CrossRef]
- Thimister, P.W.; Hopman, W.P.; Sloots, C.E.; Rosenbusch, G.; Willems, H.L.; Trijbels, F.J.; Jansen, J.B. Role of intraduodenal proteases in plasma cholecystokinin and pancreaticobiliary responses to protein and amino acids. Gastroenterology 1996, 110, 567–575. [Google Scholar] [CrossRef] [Green Version]
- Luscombe-Marsh, N.D.; Hutchison, A.T.; Soenen, S.; Steinert, R.E.; Clifton, P.M.; Horowitz, M.; Feinle-Bisset, C. Plasma free amino acid responses to intraduodenal whey protein, and relationships with insulin, glucagon-like peptide-1 and energy intake in lean healthy men. Nutrients 2016, 8, 4. [Google Scholar] [CrossRef]
- Mellinkoff, S.M.; Frankland, M.; Boyle, D.; Greipel, M. Relationship between serum amino acid concentration and fluctuations in appetite. 1956. Obes. Res. 1997, 5, 381–384. [Google Scholar] [CrossRef]
- Steinert, R.E.; Ullrich, S.S.; Geary, N.; Asarian, L.; Bueter, M.; Horowitz, M.; Feinle-Bisset, C. Comparative effects of intraduodenal amino acid infusions on food intake and gut hormone release in healthy males. Physiol. Rep. 2017, 5, e13492. [Google Scholar] [CrossRef]
- Steinert, R.E.; Landrock, M.F.; Ullrich, S.S.; Standfield, S.; Otto, B.; Horowitz, M.; Feinle-Bisset, C. Effects of intraduodenal infusion of the branched-chain amino acid leucine on ad libitum eating, gut motor and hormone functions, and glycemia in healthy men. Am. J. Clin. Nutr. 2015, 102, 820–827. [Google Scholar] [CrossRef] [PubMed]
- Steinert, R.E.; Luscombe-Marsh, N.D.; Little, T.J.; Standfield, S.; Otto, B.; Horowitz, M.; Feinle-Bisset, C. Effects of intraduodenal infusion of L-tryptophan on ad libitum eating, antropyloroduodenal motility, glycemia, insulinemia, and gut peptide secretion in healthy men. J. Clin. Endocrinol. Metab. 2014, 99, 3275–3284. [Google Scholar] [CrossRef] [PubMed]
- Ballinger, A.B.; Clark, M.L. L-phenylalanine releases cholecystokinin (CCK) and is associated with reduced food intake in humans: Evidence for a physiological role of CCK in control of eating. Metabolism 1994, 43, 735–738. [Google Scholar] [CrossRef]
- Meyer-Gerspach, A.C.; Hafliger, S.; Meili, J.; Doody, A.; Rehfeld, J.F.; Drewe, J.; Beglinger, C.; Wolnerhanssen, B. Effect of L-tryptophan and L-leucine on gut hormone secretion, appetite feelings and gastric emptying rates in lean and non-diabetic obese participants: A randomized, double-blind, parallel-group trial. PLoS ONE 2016, 11, e0166758. [Google Scholar] [CrossRef] [PubMed]
- Carney, B.I.; Jones, K.L.; Horowitz, M.; Sun, W.M.; Hebbard, G.; Edelbroek, M.A. Stereospecific effects of tryptophan on gastric emptying and hunger in humans. J. Gastroenterol. Hepatol. 1994, 9, 557–563. [Google Scholar] [CrossRef] [PubMed]
- Potier, M.; Darcel, N.; Tome, D. Protein, amino acids and the control of food intake. Curr. Opin. Clin. Nutr. Metab. Care 2009, 12, 54–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, J.E.; Feinle-Bisset, C.; Keast, R.S. Fatty acid detection during food consumption and digestion: Associations with ingestive behavior and obesity. Prog. Lipid. Res. 2011, 50, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Acosta, A.; Camilleri, M.; Shin, A.; Vazquez-Roque, M.I.; Iturrino, J.; Burton, D.; O’Neill, J.; Eckert, D.; Zinsmeister, A.R. Quantitative gastrointestinal and psychological traits associated with obesity and response to weight-loss therapy. Gastroenterology 2015, 148, 537–546. [Google Scholar] [CrossRef]
- Geliebter, A.; Schachter, S.; Lohmann-Walter, C.; Feldman, H.; Hashim, S.A. Reduced stomach capacity in obese subjects after dieting. Am. J. Clin. Nutr. 1996, 63, 170–173. [Google Scholar] [CrossRef]
- Granstrom, L.; Backman, L. Stomach distension in extremely obese and in normal subjects. Acta. Chir. Scand. 1985, 151, 367–370. [Google Scholar]
- Geliebter, A. Gastric distension and gastric capacity in relation to food intake in humans. Physiol. Behav. 1988, 44, 665–668. [Google Scholar] [CrossRef]
- Park, M.I.; Camilleri, M. Gastric motor and sensory functions in obesity. Obes. Res. 2005, 13, 491–500. [Google Scholar] [CrossRef] [PubMed]
- Speechly, D.P.; Buffenstein, R. Appetite dysfunction in obese males: Evidence for role of hyperinsulinaemia in passive overconsumption with a high fat diet. Eur. J. Clin. Nutr. 2000, 54, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Rolls, B.J.; Kim-Harris, S.; Fischman, M.W.; Foltin, R.W.; Moran, T.H.; Stoner, S.A. Satiety after preloads with different amounts of fat and carbohydrate: Implications for obesity. Am. J. Clin. Nutr. 1994, 60, 476–487. [Google Scholar] [CrossRef] [PubMed]
- French, S.J.; Murray, B.; Rumsey, R.D.; Sepple, C.P.; Read, N.W. Preliminary studies on the gastrointestinal responses to fatty meals in obese people. Int. J. Obes. Relat. Metab. Disord. 1993, 17, 295–300. [Google Scholar] [PubMed]
- Stewart, J.E.; Seimon, R.V.; Otto, B.; Keast, R.S.; Clifton, P.M.; Feinle-Bisset, C. Marked differences in gustatory and gastrointestinal sensitivity to oleic acid between lean and obese men. Am. J. Clin. Nutr. 2011, 93, 703–711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heruc, G.A.; Little, T.J.; Kohn, M.; Madden, S.; Clarke, S.; Horowitz, M.; Feinle-Bisset, C. Appetite perceptions, gastrointestinal symptoms, ghrelin, peptide YY and state anxiety are disturbed in adolescent females with anorexia nervosa and only partially restored with short-term refeeding. Nutrients 2018, 11, 59. [Google Scholar] [CrossRef]
- Geliebter, A.; Melton, P.M.; McCray, R.S.; Gallagher, D.R.; Gage, D.; Hashim, S.A. Gastric capacity, gastric emptying, and test-meal intake in normal and bulimic women. Am. J. Clin. Nutr. 1992, 56, 656–661. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Brady, P. Gastroesophageal reflux disease: Pathophysiology, diagnosis, and treatment. Gastroenterol. Nurs. 2019, 42, 20–28. [Google Scholar] [CrossRef]
- Herregods, T.V.; Bredenoord, A.J.; Smout, A.J. Pathophysiology of gastroesophageal reflux disease: New understanding in a new era. Neurogastroenterol. Motil. 2015, 27, 1202–1213. [Google Scholar] [CrossRef]
- de Bortoli, N.; Tolone, S.; Frazzoni, M.; Martinucci, I.; Sgherri, G.; Albano, E.; Ceccarelli, L.; Stasi, C.; Bellini, M.; Savarino, V.; et al. Gastroesophageal reflux disease, functional dyspepsia and irritable bowel syndrome: Common overlapping gastrointestinal disorders. Ann. Gastroenterol. 2018, 31, 639–648. [Google Scholar] [CrossRef] [PubMed]
- Barbera, R.; Feinle, C.; Read, N.W. Abnormal sensitivity to duodenal lipid infusion in patients with functional dyspepsia. Eur. J. Gastroenterol. Hepatol. 1995, 7, 1051–1057. [Google Scholar] [CrossRef] [PubMed]
- Feinle-Bisset, C.; Azpiroz, F. Dietary and lifestyle factors in functional dyspepsia. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Pilichiewicz, A.N.; Horowitz, M.; Holtmann, G.J.; Talley, N.J.; Feinle-Bisset, C. Relationship between symptoms and dietary patterns in patients with functional dyspepsia. Clin. Gastroenterol. Hepatol. 2009, 7, 317–322. [Google Scholar] [CrossRef] [PubMed]
- Bradette, M.; Pare, P.; Douville, P.; Morin, A. Visceral perception in health and functional dyspepsia. Crossover study of gastric distension with placebo and domperidone. Dig. Dis. Sci. 1991, 36, 52–58. [Google Scholar] [CrossRef]
- Mearin, F.; Cucala, M.; Azpiroz, F.; Malagelada, J.R. The origin of symptoms on the brain-gut axis in functional dyspepsia. Gastroenterology 1991, 101, 999–1006. [Google Scholar] [CrossRef]
- Caldarella, M.P.; Azpiroz, F.; Malagelada, J.R. Antro-fundic dysfunctions in functional dyspepsia. Gastroenterology 2003, 124, 1220–1229. [Google Scholar] [CrossRef]
- Feinle-Bisset, C.; Meier, B.; Fried, M.; Beglinger, C. Role of cognitive factors in symptom induction following high and low fat meals in patients with functional dyspepsia. Gut 2003, 52, 1414–1418. [Google Scholar] [CrossRef] [PubMed]
- Pilichiewicz, A.N.; Feltrin, K.L.; Horowitz, M.; Holtmann, G.; Wishart, J.M.; Jones, K.L.; Talley, N.J.; Feinle-Bisset, C. Functional dyspepsia is associated with a greater symptomatic response to fat but not carbohydrate, increased fasting and postprandial CCK, and diminished PYY. Am. J. Gastroenterol. 2008, 103, 2613–2623. [Google Scholar] [CrossRef]
- Houghton, L.A.; Mangall, Y.F.; Dwivedi, A.; Read, N.W. Sensitivity to nutrients in patients with non-ulcer dyspepsia. Eur. J. Gastroenterol. Hepatol. 1993, 5, 109–114. [Google Scholar] [CrossRef]
- Barbera, R.; Feinle, C.; Read, N.W. Nutrient-specific modulation of gastric mechanosensitivity in patients with functional dyspepsia. Dig. Dis. Sci. 1995, 40, 1636–1641. [Google Scholar] [CrossRef] [PubMed]
- Feinle, C.; Meier, O.; Otto, B.; D’Amato, M.; Fried, M. Role of duodenal lipid and cholecystokinin A receptors in the pathophysiology of functional dyspepsia. Gut 2001, 48, 347–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chua, A.S.; Dinan, T.G.; Rovati, L.C.; Keeling, P.W. Cholecystokinin hyperresponsiveness in dysmotility-type nonulcer dyspepsia. Ann. N. Y. Acad. Sci. 1994, 713, 298–299. [Google Scholar] [CrossRef] [PubMed]
- Azpiroz, F.; Bouin, M.; Camilleri, M.; Mayer, E.A.; Poitras, P.; Serra, J.; Spiller, R.C. Mechanisms of hypersensitivity in IBS and functional disorders. Neurogastroenterol. Motil. 2007, 19, 62–88. [Google Scholar] [CrossRef] [PubMed]
- Chey, W.D.; Kurlander, J.; Eswaran, S. Irritable bowel syndrome: A clinical review. JAMA 2015, 313, 949–958. [Google Scholar] [CrossRef] [PubMed]
- Ukleja, A. Altered GI motility in critically Ill patients: Current understanding of pathophysiology, clinical impact, and diagnostic approach. Nutr. Clin. Pract. 2010, 25, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Giezenaar, C.; Trahair, L.G.; Rigda, R.; Hutchison, A.T.; Feinle-Bisset, C.; Luscombe-Marsh, N.D.; Hausken, T.; Jones, K.L.; Horowitz, M.; Chapman, I.; et al. Lesser suppression of energy intake by orally ingested whey protein in healthy older men compared with young controls. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2015, 309, R845–R854. [Google Scholar] [CrossRef] [PubMed]
- Giezenaar, C.; van der Burgh, Y.; Lange, K.; Hatzinikolas, S.; Hausken, T.; Jones, K.L.; Horowitz, M.; Chapman, I.; Soenen, S. Effects of substitution, and adding of carbohydrate and fat to whey-protein on energy intake, appetite, gastric emptying, glucose, insulin, ghrelin, CCK and GLP-1 in healthy older men-a randomized controlled trial. Nutrients 2018, 10, 113. [Google Scholar] [CrossRef] [PubMed]
- MacIntosh, C.G.; Morley, J.E.; Wishart, J.; Morris, H.; Jansen, J.B.; Horowitz, M.; Chapman, I.M. Effect of exogenous cholecystokinin (CCK)-8 on food intake and plasma CCK, leptin, and insulin concentrations in older and young adults: Evidence for increased CCK activity as a cause of the anorexia of aging. J. Clin. Endocrinol. Metab. 2001, 86, 5830–5837. [Google Scholar] [CrossRef] [PubMed]
- Di Francesco, V.; Zamboni, M.; Dioli, A.; Zoico, E.; Mazzali, G.; Omizzolo, F.; Bissoli, L.; Solerte, S.B.; Benini, L.; Bosello, O. Delayed postprandial gastric emptying and impaired gallbladder contraction together with elevated cholecystokinin and peptide YY serum levels sustain satiety and inhibit hunger in healthy elderly persons. J. Gerontol. A Biol. Sci. Med. Sci. 2005, 60, 1581–1585. [Google Scholar] [CrossRef]
- Horowitz, M.; Maddern, G.J.; Chatterton, B.E.; Collins, P.J.; Harding, P.E.; Shearman, D.J. Changes in gastric emptying rates with age. Clin. Sci. 1984, 67, 213–218. [Google Scholar] [CrossRef] [PubMed]
- Wegener, M.; Borsch, G.; Schaffstein, J.; Luth, I.; Rickels, R.; Ricken, D. Effect of ageing on the gastro-intestinal transit of a lactulose-supplemented mixed solid-liquid meal in humans. Digestion 1988, 39, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Clarkston, W.K.; Pantano, M.M.; Morley, J.E.; Horowitz, M.; Littlefield, J.M.; Burton, F.R. Evidence for the anorexia of aging: Gastrointestinal transit and hunger in healthy elderly vs. young adults. Am. J. Physiol. 1997, 272, R243–R248. [Google Scholar] [CrossRef] [PubMed]
- Rayner, C.K.; MacIntosh, C.G.; Chapman, I.M.; Morley, J.E.; Horowitz, M. Effects of age on proximal gastric motor and sensory function. Scand. J. Gastroenterol. 2000, 35, 1041–1047. [Google Scholar] [PubMed]
- Soenen, S.; Giezenaar, C.; Hutchison, A.T.; Horowitz, M.; Chapman, I.; Luscombe-Marsh, N.D. Effects of intraduodenal protein on appetite, energy intake, and antropyloroduodenal motility in healthy older compared with young men in a randomized trial. Am. J. Clin. Nutr. 2014, 100, 1108–1115. [Google Scholar] [CrossRef] [Green Version]
- Cook, C.G.; Andrews, J.M.; Jones, K.L.; Wittert, G.A.; Chapman, I.M.; Morley, J.E.; Horowitz, M. Effects of small intestinal nutrient infusion on appetite and pyloric motility are modified by age. Am. J. Physiol. 1997, 273, R755–R761. [Google Scholar] [CrossRef]
- MacIntosh, C.G.; Andrews, J.M.; Jones, K.L.; Wishart, J.M.; Morris, H.A.; Jansen, J.B.; Morley, J.E.; Horowitz, M.; Chapman, I.M. Effects of age on concentrations of plasma cholecystokinin, glucagon-like peptide 1, and peptide YY and their relation to appetite and pyloric motility. Am. J. Clin. Nutr. 1999, 69, 999–1006. [Google Scholar] [CrossRef]
- Sturm, K.; MacIntosh, C.G.; Parker, B.A.; Wishart, J.; Horowitz, M.; Chapman, I.M. Appetite, food intake, and plasma concentrations of cholecystokinin, ghrelin, and other gastrointestinal hormones in undernourished older women and well-nourished young and older women. J. Clin. Endocrinol. Metab. 2003, 88, 3747–3755. [Google Scholar] [CrossRef]
- Giezenaar, C.; Hutchison, A.T.; Luscombe-Marsh, N.D.; Chapman, I.; Horowitz, M.; Soenen, S. Effect of age on blood glucose and plasma insulin, glucagon, ghrelin, CCK, GIP, and GLP-1 responses to whey protein ingestion. Nutrients 2017, 10, 2. [Google Scholar] [CrossRef]
- Rigamonti, A.E.; Pincelli, A.I.; Corra, B.; Viarengo, R.; Bonomo, S.M.; Galimberti, D.; Scacchi, M.; Scarpini, E.; Cavagnini, F.; Muller, E.E. Plasma ghrelin concentrations in elderly subjects: Comparison with anorexic and obese patients. J. Endocrinol. 2002, 175, R1–R5. [Google Scholar] [CrossRef]
- Di Francesco, V.; Fantin, F.; Residori, L.; Bissoli, L.; Micciolo, R.; Zivelonghi, A.; Zoico, E.; Omizzolo, F.; Bosello, O.; Zamboni, M. Effect of age on the dynamics of acylated ghrelin in fasting conditions and in response to a meal. J. Am. Geriatr. Soc. 2008, 56, 1369–1370. [Google Scholar] [CrossRef]
- Di Francesco, V.; Zamboni, M.; Zoico, E.; Mazzali, G.; Dioli, A.; Omizzolo, F.; Bissoli, L.; Fantin, F.; Rizzotti, P.; Solerte, S.B.; et al. Unbalanced serum leptin and ghrelin dynamics prolong postprandial satiety and inhibit hunger in healthy elderly: Another reason for the “anorexia of aging”. Am. J. Clin. Nutr. 2006, 83, 1149–1152. [Google Scholar] [CrossRef]
- Wisen, O.; Johansson, C. Gastrointestinal function in obesity: Motility, secretion, and absorption following a liquid test meal. Metabolism 1992, 41, 390–395. [Google Scholar] [CrossRef]
- Castiglione, K.E.; Read, N.W.; French, S.J. Adaptation to high-fat diet accelerates emptying of fat but not carbohydrate test meals in humans. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2002, 282, R366–R371. [Google Scholar] [CrossRef]
- Park, M.I.; Camilleri, M.; O’Connor, H.; Oenning, L.; Burton, D.; Stephens, D.; Zinsmeister, A.R. Effect of different macronutrients in excess on gastric sensory and motor functions and appetite in normal-weight, overweight, and obese humans. Am. J. Clin. Nutr. 2007, 85, 411–418. [Google Scholar] [CrossRef]
- French, S.J.; Murray, B.; Rumsey, R.D.; Fadzlin, R.; Read, N.W. Adaptation to high-fat diets: Effects on eating behaviour and plasma cholecystokinin. Br. J. Nutr. 1995, 73, 179–189. [Google Scholar] [CrossRef]
- Boyd, K.A.; O’Donovan, D.G.; Doran, S.; Wishart, J.; Chapman, I.M.; Horowitz, M.; Feinle, C. High-fat diet effects on gut motility, hormone, and appetite responses to duodenal lipid in healthy men. Am. J. Physiol. Gastrointest. Liver Physiol. 2003, 284, G188–G196. [Google Scholar] [CrossRef] [Green Version]
- Brennan, I.M.; Seimon, R.V.; Luscombe-Marsh, N.D.; Otto, B.; Horowitz, M.; Feinle-Bisset, C. Effects of acute dietary restriction on gut motor, hormone and energy intake responses to duodenal fat in obese men. Int. J. Obes. 2011, 35, 448–456. [Google Scholar] [CrossRef]
- Seimon, R.V.; Taylor, P.; Little, T.J.; Noakes, M.; Standfield, S.; Clifton, P.M.; Horowitz, M.; Feinle-Bisset, C. Effects of acute and longer-term dietary restriction on upper gut motility, hormone, appetite, and energy-intake responses to duodenal lipid in lean and obese men. Am. J. Clin. Nutr. 2014, 99, 24–34. [Google Scholar] [CrossRef]
- Weigle, D.S.; Breen, P.A.; Matthys, C.C.; Callahan, H.S.; Meeuws, K.E.; Burden, V.R.; Purnell, J.Q. A high-protein diet induces sustained reductions in appetite, ad libitum caloric intake, and body weight despite compensatory changes in diurnal plasma leptin and ghrelin concentrations. Am. J. Clin. Nutr. 2005, 82, 41–48. [Google Scholar] [CrossRef]
- Leidy, H.J.; Clifton, P.M.; Astrup, A.; Wycherley, T.P.; Westerterp-Plantenga, M.S.; Luscombe-Marsh, N.D.; Woods, S.C.; Mattes, R.D. The role of protein in weight loss and maintenance. Am. J. Clin. Nutr. 2015, 101, 1320S–1329S. [Google Scholar] [CrossRef] [Green Version]
- Hutchison, A.T.; Feinle-Bisset, C.; Fitzgerald, P.C.; Standfield, S.; Horowitz, M.; Clifton, P.M.; Luscombe-Marsh, N.D. Comparative effects of intraduodenal whey protein hydrolysate on antropyloroduodenal motility, gut hormones, glycemia, appetite, and energy intake in lean and obese men. Am. J. Clin. Nutr. 2015, 102, 1323–1331. [Google Scholar] [CrossRef]
- Edelbroek, M.; Sun, W.M.; Horowitz, M.; Dent, J.; Smout, A.; Akkermans, L. Stereospecific effects of intraduodenal tryptophan on pyloric and duodenal motility in humans. Scand. J. Gastroenterol. 1994, 29, 1088–1095. [Google Scholar] [CrossRef]
- Ullrich, S.S.; Fitzgerald, P.C.E.; Giesbertz, P.; Steinert, R.E.; Horowitz, M.; Feinle-Bisset, C. Effects of intragastric administration of tryptophan on the blood glucose response to a nutrient drink and energy intake, in lean and obese men. Nutrients 2018, 10, 463. [Google Scholar] [CrossRef]
- Pepino, M.Y.; Finkbeiner, S.; Beauchamp, G.K.; Mennella, J.A. Obese women have lower monosodium glutamate taste sensitivity and prefer higher concentrations than do normal-weight women. Obesity 2010, 18, 959–965. [Google Scholar] [CrossRef]
- Tack, J.; Talley, N.J. Functional dyspepsia--symptoms, definitions and validity of the Rome III criteria. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 134–141. [Google Scholar] [CrossRef]
- Barbara, G.; Feinle-Bisset, C.; Ghoshal, U.C.; Quigley, E.M.; Santos, J.; Vanner, S.; Vergnolle, N.; Zoetendal, E.G. The intestinal microenvironment and functional gastrointestinal disorders. Gastroenterology 2016, 150, 1305–1318. [Google Scholar] [CrossRef]
- Bisschops, R.; Karamanolis, G.; Arts, J.; Caenepeel, P.; Verbeke, K.; Janssens, J.; Tack, J. Relationship between symptoms and ingestion of a meal in functional dyspepsia. Gut 2008, 57, 1495–1503. [Google Scholar] [CrossRef]
- Filipovic, B.F.; Randjelovic, T.; Kovacevic, N.; Milinic, N.; Markovic, O.; Gajic, M.; Filipovic, B.R. Laboratory parameters and nutritional status in patients with functional dyspepsia. Eur. J. Intern. Med. 2011, 22, 300–304. [Google Scholar] [CrossRef]
- Carvalho, R.V.; Lorena, S.L.; Almeida, J.R.; Mesquita, M.A. Food intolerance, diet composition, and eating patterns in functional dyspepsia patients. Dig. Dis. Sci. 2010, 55, 60–65. [Google Scholar] [CrossRef]
- Cuperus, P.; Keeling, P.W.; Gibney, M.J. Eating patterns in functional dyspepsia: A case control study. Eur. J. Clin. Nutr. 1996, 50, 520–523. [Google Scholar]
- Goktas, Z.; Koklu, S.; Dikmen, D.; Ozturk, O.; Yilmaz, B.; Asil, M.; Korkmaz, H.; Tuna, Y.; Kekilli, M.; Karamanoglu Aksoy, E.; et al. Nutritional habits in functional dyspepsia and its subgroups: A comparative study. Scand. J. Gastroenterol. 2016, 51, 903–907. [Google Scholar] [CrossRef]
- Gibson, P.R.; Shepherd, S.J. Food choice as a key management strategy for functional gastrointestinal symptoms. Am. J. Gastroenterol. 2012, 107, 657–666. [Google Scholar] [CrossRef]
- Mullan, A.; Kavanagh, P.; O’Mahony, P.; Joy, T.; Gleeson, F.; Gibney, M.J. Food and nutrient intakes and eating patterns in functional and organic dyspepsia. Eur. J. Clin. Nutr. 1994, 48, 97–105. [Google Scholar]
- Kaess, H.; Kellermann, M.; Castro, A. Food intolerance in duodenal ulcer patients, non ulcer dyspeptic patients and healthy subjects. A prospective study. Klin. Wochenschr. 1988, 66, 208–211. [Google Scholar] [CrossRef]
- Elli, L.; Tomba, C.; Branchi, F.; Roncoroni, L.; Lombardo, V.; Bardella, M.T.; Ferretti, F.; Conte, D.; Valiante, F.; Fini, L.; et al. Evidence for the presence of non-celiac gluten sensitivity in patients with functional gastrointestinal symptoms: Results from a multicenter randomized double-blind placebo-controlled gluten challenge. Nutrients 2016, 8, 84. [Google Scholar] [CrossRef]
- Santolaria, S.; Alcedo, J.; Cuartero, B.; Diez, I.; Abascal, M.; Garcia-Prats, M.D.; Marigil, M.; Vera, J.; Ferrer, M.; Montoro, M. Spectrum of gluten-sensitive enteropathy in patients with dysmotility-like dyspepsia. Gastroenterol. Hepatol. 2013, 36, 11–20. [Google Scholar] [CrossRef]
- Saito, Y.A.; Locke, G.R., 3rd; Weaver, A.L.; Zinsmeister, A.R.; Talley, N.J. Diet and functional gastrointestinal disorders: A population-based case-control study. Am. J. Gastroenterol. 2005, 100, 2743–2748. [Google Scholar] [CrossRef]
- De Castro, J.M. Age-related changes in spontaneous food intake and hunger in humans. Appetite 1993, 21, 255–272. [Google Scholar] [CrossRef]
- Bauer, J.; Biolo, G.; Cederholm, T.; Cesari, M.; Cruz-Jentoft, A.J.; Morley, J.E.; Phillips, S.; Sieber, C.; Stehle, P.; Teta, D.; et al. Evidence-based recommendations for optimal dietary protein intake in older people: A position paper from the PROT-AGE Study Group. J. Am. Med. Dir. Assoc. 2013, 14, 542–559. [Google Scholar] [CrossRef]
- Ryan, M.; Salle, A.; Favreau, A.M.; Simard, G.; Dumas, J.F.; Malthiery, Y.; Berrut, G.; Ritz, P. Oral supplements differing in fat and carbohydrate content: Effect on the appetite and food intake of undernourished elderly patients. Clin. Nutr. 2004, 23, 683–689. [Google Scholar] [CrossRef]
- Kupfer, R.M.; Heppell, M.; Haggith, J.W.; Bateman, D.N. Gastric emptying and small-bowel transit rate in the elderly. J. Am. Geriatr. Soc. 1985, 33, 340–343. [Google Scholar] [CrossRef]
- Rolls, B.J.; Dimeo, K.A.; Shide, D.J. Age-related impairments in the regulation of food intake. Am. J. Clin. Nutr. 1995, 62, 923–931. [Google Scholar] [CrossRef]
- Roberts, S.B.; Fuss, P.; Heyman, M.B.; Evans, W.J.; Tsay, R.; Rasmussen, H.; Fiatarone, M.; Cortiella, J.; Dallal, G.E.; Young, V.R. Control of food intake in older men. JAMA 1994, 272, 1601–1606. [Google Scholar] [CrossRef]
- Milan, A.M.; Cameron-Smith, D. Digestion and postprandial metabolism in the elderly. Adv. Food Nutr. Res. 2015, 76, 79–124. [Google Scholar]
- Poppitt, S.D.; McCormack, D.; Buffenstein, R. Short-term effects of macronutrient preloads on appetite and energy intake in lean women. Physiol. Behav. 1998, 64, 279–285. [Google Scholar] [CrossRef]
- Tylner, S.; Cederholm, T.; Faxen-Irving, G. Effects on weight, blood lipids, serum fatty acid profile and coagulation by an energy-dense formula to older care residents: A randomized controlled crossover trial. J. Am. Med. Dir. Assoc. 2016, 17, 275. [Google Scholar] [CrossRef]
- Spiller, R.C.; Trotman, I.F.; Adrian, T.E.; Bloom, S.R.; Misiewicz, J.J.; Silk, D.B. Further characterisation of the ’ileal brake’ reflex in man--effect of ileal infusion of partial digests of fat, protein, and starch on jejunal motility and release of neurotensin, enteroglucagon, and peptide YY. Gut 1988, 29, 1042–1051. [Google Scholar] [CrossRef]


{kind=link}
| GI Sensory Disturbances | References | |
|---|---|---|
| Obesity | ↓ Sensitivity to gastric distension ↑ Gastric capacity ↓ Sensitivity to small intestinal lipid ?↓ Sensitivity to CCK, PYY, ghrelin ↑ ↓ ↔ CCK secretion ↑ ↓ ↔ Gastric emptying | [42,43,57,58,59,60,61,62,63,64,65,66] |
| Psychiatric eating disorders | ||
| Anorexia nervosa | ↑ Sensitivity to gastric distension ↑ Sensitivity to small intestinal nutrients ↑ ↔ CCK secretion ?↑ Sensitivity to CCK ↑ Ghrelin, PYY secretion ↓ Gastric emptying ↓ Proximal gastric accommodation ↑ Antral filling | [13,16,17,67] |
| Bulimia nervosa | ↑ Sensitivity to gastric distension ↑ Gastric capacity ↓ CCK secretion ↓ ↔ Gastric emptying ↓ Proximal gastric accommodation | [16,17,68] |
| GI disorders | ||
| Gastroesophagealreflux disease | ↑ Sensitivity to gastric distension ↑ Sensitivity to small intestinal lipid ↓ Gastric emptying ↓ Gastric accommodation | [18,69,70,71] |
| Functional dyspepsia | ↑ Sensitivity to gastric distension ↑ Sensitivity to small intestinal lipid ?↑ Sensitivity to CCK ↓ Gastric emptying ↓ Gastric accommodation ↑ Antral distension | [11,18,72,73,74,75,76,77,78,79,80,81,82,83] |
| Irritable bowelsyndrome | ↑ Sensitivity to gastric distension ↑ Sensitivity to small intestinal lipid ↓ Gastric emptying ↑ ↓ Gut motility | [18,84,85] |
| Critical illness | ↓ Sensitivity to gastric distension ↑ Sensitivity to small intestinal nutrients ↑ CCK, PYY secretion ↓ Ghrelin, motilin secretion ↓ Gastric emptying | [12,19,86] |
| Anorexia of ageing | ↓ Sensitivity to gastric distension ↓ Sensitivity to small intestinal nutrients ↑ CCK secretion ↑ ↓ ↔ PYY, GLP-1 and ghrelin secretion ?↓ Sensitivity to CCK, PYY and GLP-1 ↓ ↔ Gastric emptying ↑ Proximal gastric retention | [26,87,88,89,90,91,92,93,94,95,96,97,98,99,100,101,102] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hajishafiee, M.; Bitarafan, V.; Feinle-Bisset, C. Gastrointestinal Sensing of Meal-Related Signals in Humans, and Dysregulations in Eating-Related Disorders. Nutrients 2019, 11, 1298. https://doi.org/10.3390/nu11061298
Hajishafiee M, Bitarafan V, Feinle-Bisset C. Gastrointestinal Sensing of Meal-Related Signals in Humans, and Dysregulations in Eating-Related Disorders. Nutrients. 2019; 11(6):1298. https://doi.org/10.3390/nu11061298
Chicago/Turabian StyleHajishafiee, Maryam, Vida Bitarafan, and Christine Feinle-Bisset. 2019. "Gastrointestinal Sensing of Meal-Related Signals in Humans, and Dysregulations in Eating-Related Disorders" Nutrients 11, no. 6: 1298. https://doi.org/10.3390/nu11061298
APA StyleHajishafiee, M., Bitarafan, V., & Feinle-Bisset, C. (2019). Gastrointestinal Sensing of Meal-Related Signals in Humans, and Dysregulations in Eating-Related Disorders. Nutrients, 11(6), 1298. https://doi.org/10.3390/nu11061298