Iron and Zinc Homeostasis and Interactions: Does Enteric Zinc Excretion Cross-Talk with Intestinal Iron Absorption?

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Iron Homeostasis
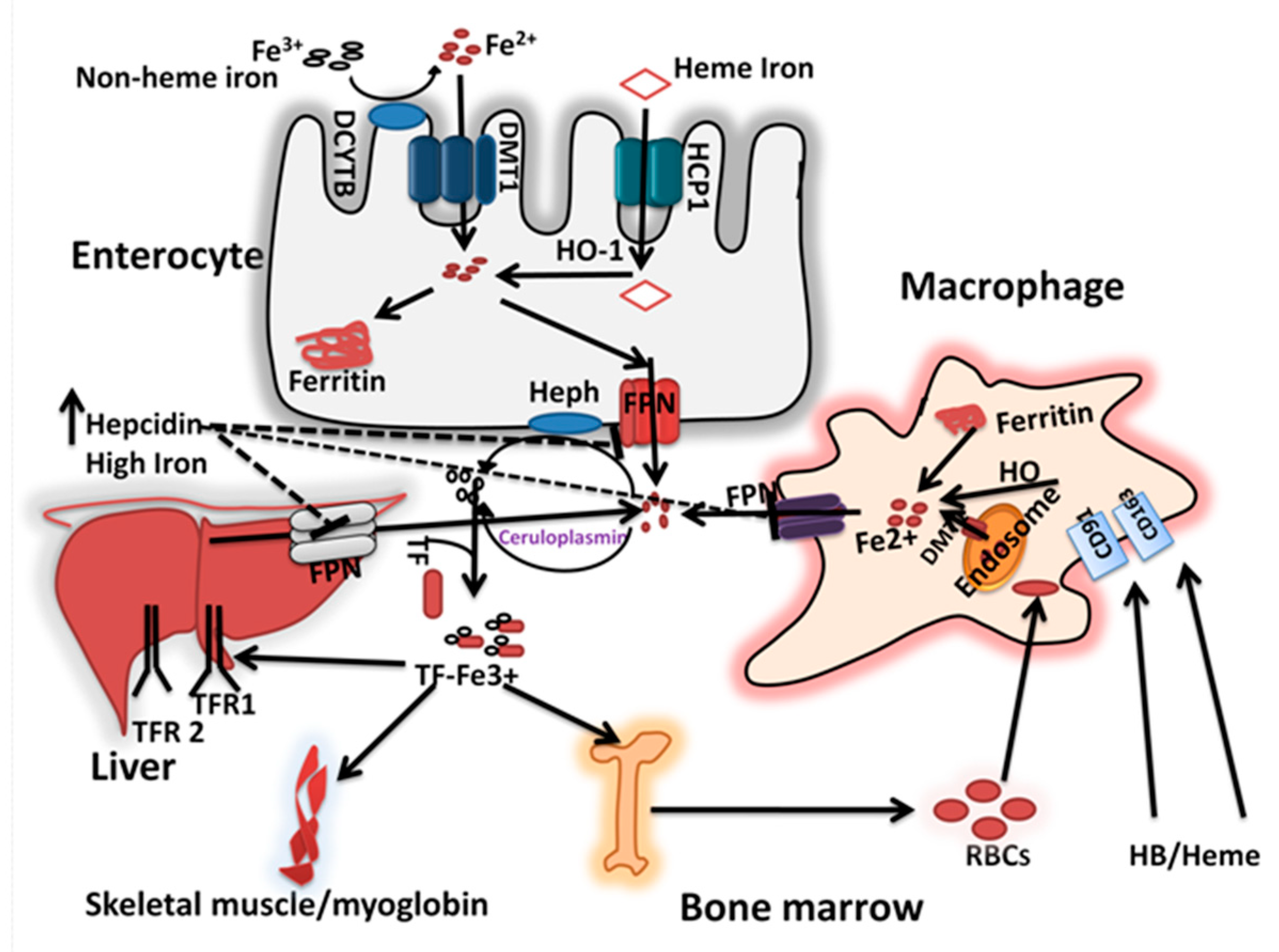
2.1. Iron Absorption and Recycling
2.2. Regulation of Cellular and Systemtic Iron Homeostasis
3. Zinc Homeostasis
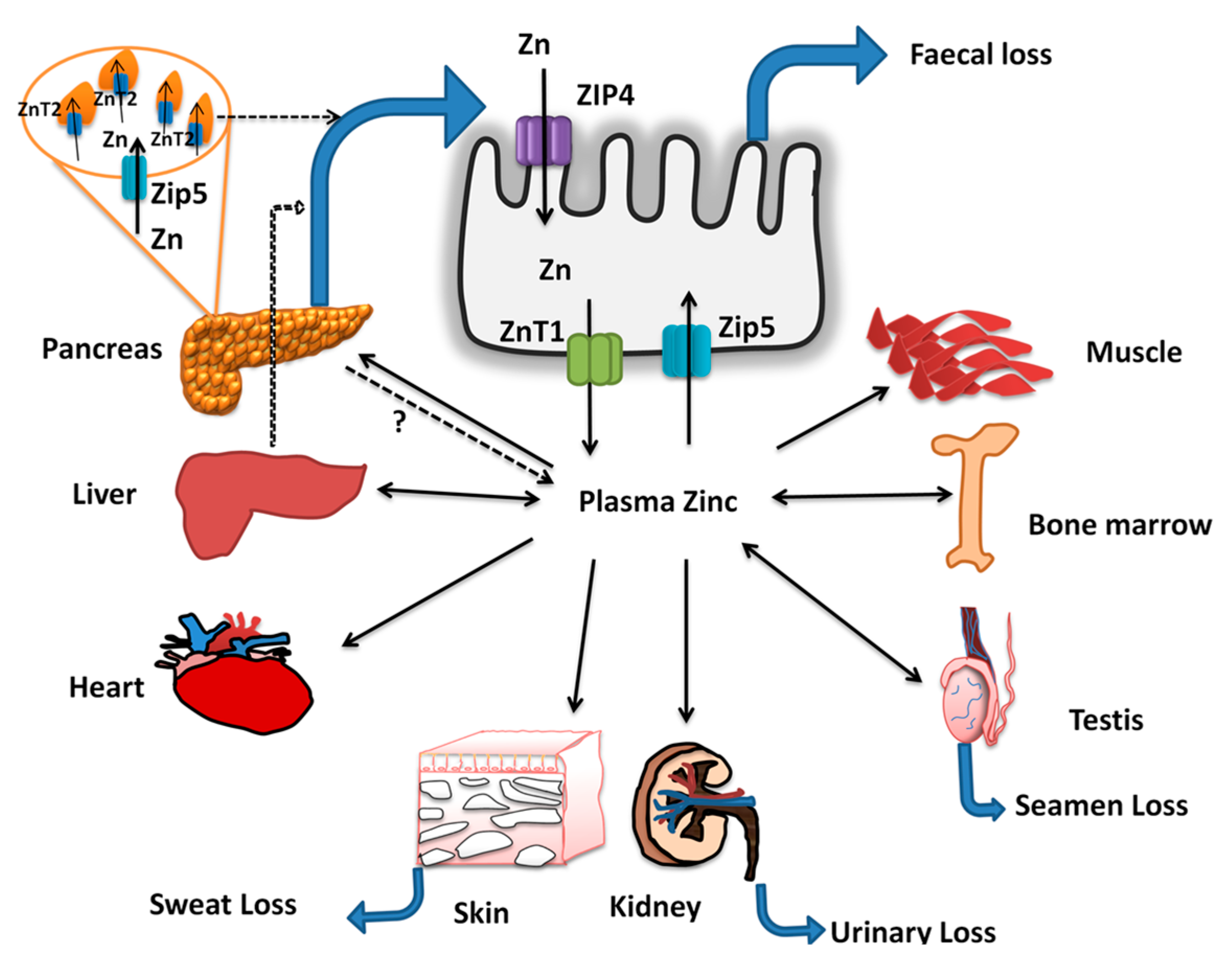
Zinc Absorption and Excretion
4. Impact of Zinc Deficiency on Iron Status
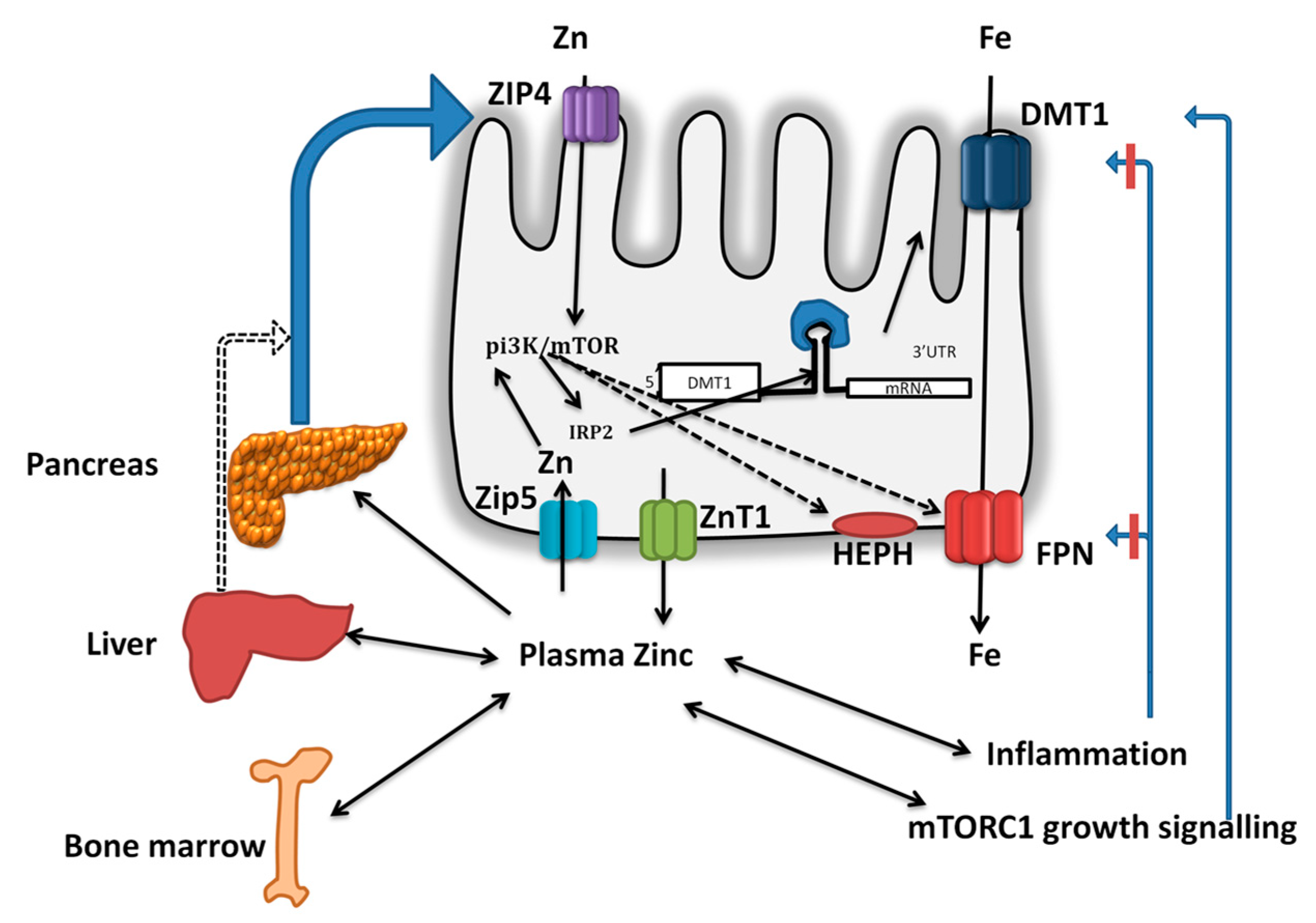
5. Iron and Zinc Interactions During Absorption
6. Does Enteric Zinc-DMT1/FPN1 Axis Play a Role in Intestinal Iron Absorption?
7. Physiological Advantages of Zinc Modulating Iron Homeostasis
8. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Golden, M.H. Specific deficiencies versus growth failure: Type I and type II nutrients. SCN News 1995, 12, 10–14. [Google Scholar] [CrossRef]
- Brown, K.H.; Rivera, J.A.; Bhutta, Z.; Gibson, R.S.; King, J.C.; Lonnerdal, B.; Ruel, M.T.; Sandtrom, B.; Wasantwisut, E.; Hotz, C. International Zinc Nutrition Consultative Group (IZiNCG) technical document #1. Assessment of the risk of zinc deficiency in populations and options for its control. Food. Nutr. Bull. 2004, 25, S99–S203. [Google Scholar] [PubMed]
- Nair, K.M.; Iyengar, V. Iron content, bioavailability & factors affecting iron status of Indians. Indian J. Med. Res. 2009, 130, 634–645. [Google Scholar] [PubMed]
- Gibson, R.S.; Raboy, V.; King, J.C. Implications of phytate in plant-based foods for iron and zinc bioavailability, setting dietary requirements, and formulating programs and policies. Nutr. Rev. 2018, 76, 793–804. [Google Scholar] [CrossRef] [PubMed]
- Lonnerdal, B. Dietary factors influencing zinc absorption. J. Nutr. 2000, 130, 1378S–1383S. [Google Scholar] [CrossRef] [PubMed]
- Whittaker, P. Iron and zinc interactions in humans. Am. J. Clin. Nutr. 1998, 68, 442S–446S. [Google Scholar] [CrossRef] [PubMed]
- Fischer Walker, C.; Kordas, K.; Stoltzfus, R.J.; Black, R.E. Interactive effects of iron and zinc on biochemical and functional outcomes in supplementation trials. Am. J. Clin. Nutr. 2005, 82, 5–12. [Google Scholar] [CrossRef]
- El Hendy, H.A.; Yousef, M.I.; Abo El-Naga, N.I. Effect of dietary zinc deficiency on hematological and biochemical parameters and concentrations of zinc, copper, and iron in growing rats. Toxicology 2001, 167, 163–170. [Google Scholar] [CrossRef]
- Geiser, J.; Venken, K.J.T.; De Lisle, R.C.; Andrews, G.K. A Mouse Model of Acrodermatitis Enteropathica: Loss of Intestine Zinc Transporter ZIP4 (Slc39a4) Disrupts the Stem Cell Niche and Intestine Integrity. PLoS Gen. 2012, 8, e1002766. [Google Scholar] [CrossRef]
- Prasad, A.S.; Oberleas, D.; Wolf, P.; Horwitz, H.P. Studies on zinc deficiency: Changes in trace elements and enzyme activities in tissues of zinc-deficient rats. J. Clin. Investig. 1967, 46, 549–557. [Google Scholar] [CrossRef]
- Niles, B.J.; Clegg, M.S.; Hanna, L.A.; Chou, S.S.; Momma, T.Y.; Hong, H.; Keen, C.L. Zinc deficiency-induced iron accumulation, a consequence of alterations in iron regulatory protein-binding activity, iron transporters, and iron storage proteins. J. Biol. Chem. 2008, 283, 5168–5177. [Google Scholar] [CrossRef] [PubMed]
- Ergul, A.B.; Turanoglu, C.; Karakukcu, C.; Karaman, S.; Torun, Y.A. Increased Iron Deficiency and Iron Deficiency Anemia in Children with Zinc Deficiency. Eurasian J. Med. 2018, 50, 34–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, A.G.; Chen, X.C.; Xu, R.X.; Zheng, M.C.; Wang, Y.; Li, J.S. Comparison of serum levels of iron, zinc and copper in anaemic and non-anaemic pregnant women in China. Asia Pac. J. Clin. Nutr. 2004, 13, 348–352. [Google Scholar] [PubMed]
- Houghton, L.A.; Parnell, W.R.; Thomson, C.D.; Green, T.J.; Gibson, R.S.; Timothy, J. Serum Zinc Is a Major Predictor of Anemia and Mediates the Effect of Selenium on Hemoglobin in School-Aged Children in a Nationally Representative Survey in New Zealand, 2. J. Nutr. 2016, 146, 1670–1676. [Google Scholar] [CrossRef] [PubMed]
- Gibson, R.S.; Abebe, Y.; Stabler, S.; Allen, R.H.; Westcott, J.E.; Stoecker, B.J.; Krebs, N.F.; Hambidge, K.M. Zinc, gravida, infection, and iron, but not vitamin B-12 or folate status, predict hemoglobin during pregnancy in Southern Ethiopia. J. Nutr. 2008, 138, 581–586. [Google Scholar] [CrossRef] [PubMed]
- Yamaji, S.; Tennant, J.; Tandy, S.; Williams, M.; Singh Srai, S.K.; Sharp, P. Zinc regulates the function and expression of the iron transporters DMT1 and IREG1 in human intestinal Caco-2 cells. FEBS Lett. 2001, 507, 137–141. [Google Scholar] [CrossRef]
- Iyengar, V.; Pullakhandam, R.; Nair, K.M. Coordinate expression and localization of iron and zinc transporters explain iron-zinc interactions during uptake in Caco-2 cells: Implications for iron uptake at the enterocyte. J. Nutr. Biochem. 2012, 23, 1146–1154. [Google Scholar] [CrossRef] [PubMed]
- Kelleher, S.L.; Lonnerdal, B. Zinc supplementation reduces iron absorption through age-dependent changes in small intestine iron transporter expression in suckling rat pups. J. Nutr. 2006, 136, 1185–1191. [Google Scholar] [CrossRef]
- Iyengar, V.; Pullakhandam, R.; Nair, K.M. Iron-zinc interaction during uptake in human intestinal Caco-2 cell line: Kinetic analyses and possible mechanism. Indian J. Biochem. Biophys. 2009, 46, 299–306. [Google Scholar]
- Kondaiah, P.; Aslam, M.F.; Mashurabad, P.; Sharp, P.A.; Pullakhandam, R. Zinc induces iron uptake and DMT1 expression in Caco-2 cells via a PI3K/IRP2 dependent mechanism. Biochem. J. 2019, 476, 1573–1583. [Google Scholar] [CrossRef]
- Gulec, S.; Anderson, G.J.; Collins, J.F. Mechanistic and regulatory aspects of intestinal iron absorption. Am. J. Physiol Gastrointest. Liver Physiol. 2014, 307, G397–G409. [Google Scholar] [CrossRef] [Green Version]
- Anderson, G.J.; Frazer, D.M.; McKie, A.T.; Vulpe, C.D.; Smith, A. Mechanisms of haem and non-haem iron absorption: Lessons from inherited disorders of iron metabolism. Biometals 2005, 18, 339–348. [Google Scholar] [CrossRef]
- Hallberg, L. Bioavailability of dietary iron in man. Annu. Rev. Nutr. 1981, 1, 123–147. [Google Scholar] [CrossRef]
- Sharp, P.; Srai, S.K. Molecular mechanisms involved in intestinal iron absorption. World J. Gastroenterol. 2007, 13, 4716–4724. [Google Scholar] [CrossRef] [PubMed]
- McKie, A.T.; Barrow, D.; Latunde-Dada, G.O.; Rolfs, A.; Sager, G.; Mudaly, E.; Mudaly, M.; Richardson, C.; Barlow, D.; Bomford, A.; et al. An iron-regulated ferric reductase associated with the absorption of dietary iron. Science 2001, 291, 1755–1759. [Google Scholar] [CrossRef]
- Gunshin, H.; Mackenzie, B.; Berger, U.V.; Gunshin, Y.; Romero, M.F.; Boron, W.F.; Nussberger, S.; Gollan, J.L.; Hediger, M.A. Cloning and characterization of a mammalian proton-coupled metal-ion transporter. Nature 1997, 388, 482–488. [Google Scholar] [CrossRef] [PubMed]
- Mancias, J.D.; Pontano Vaites, L.; Nissim, S.; Biancur, D.E.; Kim, A.J.; Wang, X.; Liu, Y.; Goessling, W.; Kimmelman, A.C.; Harper, J.W. Ferritinophagy via NCOA4 is required for erythropoiesis and is regulated by iron dependent HERC2-mediated proteolysis. eLife 2015, 4, e10308. [Google Scholar] [CrossRef]
- McKie, A.T.; Marciani, P.; Rolfs, A.; Brennan, K.; Wehr, K.; Barrow, D.; Miret, S.; Bomford, A.; Peters, T.J.; Farzaneh, F.; et al. A novel duodenal iron-regulated transporter, IREG1, implicated in the basolateral transfer of iron to the circulation. Mol. Cell 2000, 5, 299–309. [Google Scholar] [CrossRef]
- Anderson, G.J.; Frazer, D.M.; McKie, A.T.; Vulpe, C.D. The ceruloplasmin homolog hephaestin and the control of intestinal iron absorption. Blood Cells Mol. Dis. 2002, 29, 367–375. [Google Scholar] [CrossRef]
- Donovan, A.; Brownlie, A.; Zhou, Y.; Shepard, J.; Pratt, S.J.; Moynihan, J.; Paw, B.H.; Drejer, A.; Barut, B.; Zapata, A.; et al. Positional cloning of zebrafish ferroportin1 identifies a conserved vertebrate iron exporter. Nature 2000, 403, 776–781. [Google Scholar] [CrossRef]
- Drakesmith, H.; Nemeth, E.; Ganz, T. Ironing out Ferroportin. Cell Metab. 2015, 22, 777–787. [Google Scholar] [CrossRef] [Green Version]
- Harding, C.; Heuser, J.; Stahl, P. Receptor-mediated endocytosis of transferrin and recycling of the transferrin receptor in rat reticulocytes. J. Cell Biol. 1983, 97, 329–339. [Google Scholar] [CrossRef]
- MacKenzie, E.L.; Iwasaki, K.; Tsuji, Y. Intracellular iron transport and storage: From molecular mechanisms to health implications. Antioxid. Redox. Signal. 2008, 10, 997–1030. [Google Scholar] [CrossRef]
- Muckenthaler, M.U.; Rivella, S.; Hentze, M.W.; Galy, B. A Red Carpet for Iron Metabolism. Cell 2017, 168, 344–361. [Google Scholar] [CrossRef] [Green Version]
- Casey, J.L.; Hentze, M.W.; Koeller, D.M.; Caughman, S.W.; Rouault, T.A.; Klausner, R.D.; Harford, J.B. Iron-responsive elements: Regulatory RNA sequences that control mRNA levels and translation. Science 1988, 240, 924–928. [Google Scholar] [CrossRef]
- Collins, J.F.; Wessling-Resnick, M.; Knutson, M.D. Hepcidin regulation of iron transport. J. Nutr. 2008, 138, 2284–2288. [Google Scholar] [CrossRef]
- Ganz, T. Hepcidin, a key regulator of iron metabolism and mediator of anemia of inflammation. Blood 2003, 102, 783–788. [Google Scholar] [CrossRef] [Green Version]
- Qiao, B.; Sugianto, P.; Fung, E.; Del-Castillo-Rueda, A.; Moran-Jimenez, M.J.; Ganz, T.; Nemeth, E. Hepcidin-induced endocytosis of ferroportin is dependent on ferroportin ubiquitination. Cell Metab. 2012, 15, 918–924. [Google Scholar] [CrossRef]
- Nemeth, E.; Tuttle, M.S.; Powelson, J.; Vaughn, M.B.; Donovan, A.; Ward, D.M.; Ganz, T.; Kaplan, J. Hepcidin regulates cellular iron efflux by binding to ferroportin and inducing its internalization. Science 2004, 306, 2090–2093. [Google Scholar] [CrossRef]
- Vallee, B.L.; Falchuk, K.H. The biochemical basis of zinc physiology. Physiol. Rev. 1993, 73, 79–118. [Google Scholar] [CrossRef]
- King, J.C.; Shames, D.M.; Woodhouse, L.R. Zinc homeostasis in humans. J. Nutr. 2000, 130, 1360S–1366S. [Google Scholar] [CrossRef]
- Condomina, J.; Zornoza-Sabina, T.; Granero, L.; Polache, A. Kinetics of zinc transport in vitro in rat small intestine and colon: Interaction with copper. Eur. J. Pharm. Sci. 2002, 16, 289–295. [Google Scholar] [CrossRef]
- Woodhouse, L.R.; Lowe, N.M.; Randolph, J.M.; Sutherland, B.; Turnlund, J.R.; Jackson, M.J.; King, J.C. Zinc homeostasis during dietary zinc depletion and repletion in humans. Faseb. J. 1997, 11, 1128. [Google Scholar]
- Liuzzi, J.P.; Bobo, J.A.; Lichten, L.A.; Samuelson, D.A.; Cousins, R.J. Responsive transporter genes within the murine intestinal-pancreatic axis form a basis of zinc homeostasis. Proc. Natl. Acad. Sci. USA 2004, 101, 14355–14360. [Google Scholar] [CrossRef] [Green Version]
- Kambe, T.; Yamaguchi-Iwai, Y.; Sasaki, R.; Nagao, M. Overview of mammalian zinc transporters. Cell Mol. Life Sci. 2004, 61, 49–68. [Google Scholar] [CrossRef]
- Lichten, L.A.; Cousins, R.J. Mammalian zinc transporters: Nutritional and physiologic regulation. Annu. Rev. Nutr. 2009, 29, 153–176. [Google Scholar] [CrossRef]
- Liuzzi, J.P.; Cousins, R.J. Mammalian zinc transporters. Annu. Rev. Nutr. 2004, 24, 151–172. [Google Scholar] [CrossRef]
- Wang, F.D.; Kim, B.E.; Dufner-Beattie, J.; Petris, M.J.; Andrews, G.; Eide, D.J. Acrodermatitis enteropathica mutations affect transport activity, localization and zinc-responsive trafficking of the mouse ZIP4 zinc transporter. Hum. Mol. Gen. 2004, 13, 563–571. [Google Scholar] [CrossRef] [Green Version]
- Kambe, T.; Andrews, G.K. Novel Proteolytic Processing of the Ectodomain of the Zinc Transporter ZIP4 (SLC39A4) during Zinc Deficiency Is Inhibited by Acrodermatitis Enteropathica Mutations. Mol. Cell. Biol. 2009, 29, 129–139. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Kim, B.E.; Petris, M.J.; Eide, D.J. The mammalian Zip5 protein is a zinc transporter that localizes to the basolateral surface of polarized cells. J. Biol Chem 2004, 279, 51433–51441. [Google Scholar] [CrossRef]
- Dufner-Beattie, J.; Kuo, Y.M.; Gitschier, J.; Andrews, G.K. The adaptive response to dietary zinc in mice involves the differential cellular localization and zinc regulation of the zinc transporters ZIP4 and ZIP5. J. Biol. Chem. 2004, 279, 49082–49090. [Google Scholar] [CrossRef]
- Palmiter, R.D. Protection against zinc toxicity by metallothionein and zinc transporter 1. Proc. Natl. Acad. Sci. USA 2004, 101, 4918–4923. [Google Scholar] [CrossRef] [Green Version]
- Palmiter, R.D.; Findley, S.D. Cloning and functional characterization of a mammalian zinc transporter that confers resistance to zinc. EMBO J. 1995, 14, 639–649. [Google Scholar] [CrossRef]
- McMahon, R.J.; Cousins, R.J. Regulation of the zinc transporter ZnT-1 by dietary zinc. Proc. Natl. Acad. Sci. USA 1998, 95, 4841–4846. [Google Scholar] [CrossRef] [Green Version]
- Wastney, M.E.; House, W.A.; Barnes, R.M.; Subramanian, K.N. Kinetics of zinc metabolism: Variation with diet, genetics and disease. J. Nutr. 2000, 130, 1355S–1359S. [Google Scholar] [CrossRef]
- Hambidge, K.M.; Miller, L.V.; Westcott, J.E.; Sheng, X.; Krebs, N.F. Zinc bioavailability and homeostasis. Am. J. Clin. Nutr. 2010, 91, 1478S–1483S. [Google Scholar] [CrossRef] [Green Version]
- Williams, R.B.; Mills, C.F. The experimental production of zinc deficiency in the rat. Br. J. Nutr. 1970, 24, 989–1003. [Google Scholar] [CrossRef] [Green Version]
- Guo, L.; Lichten, L.A.; Ryu, M.S.; Liuzzi, J.P.; Wang, F.; Cousins, R.J. STAT5-glucocorticoid receptor interaction and MTF-1 regulate the expression of ZnT2 (Slc30a2) in pancreatic acinar cells. Proc. Natl. Acad. Sci. USA 2010, 107, 2818–2823. [Google Scholar] [CrossRef] [Green Version]
- Mostafa, W.Z.; Al-Zayer, A.A. Acrodermatitis enteropathica in Saudi Arabia. Int. J. Dermatol. 1990, 29, 134–138. [Google Scholar] [CrossRef]
- Sreedhar, B.; Nair, K.M. Modulation of aconitase, metallothionein, and oxidative stress in zinc-deficient rat intestine during zinc and iron repletion. Free Radic. Biol. Med. 2005, 39, 999–1008. [Google Scholar] [CrossRef]
- Rogers, J.M.; Lonnerdal, B.; Hurley, L.S.; Keen, C.L. Iron and zinc concentrations and 59Fe retention in developing fetuses of zinc-deficient rats. J. Nutr. 1987, 117, 1875–1882. [Google Scholar] [CrossRef]
- Zheng, D.; Kille, P.; Feeney, G.P.; Cunningham, P.; Handy, R.D.; Hogstrand, C. Dynamic transcriptomic profiles of zebrafish gills in response to zinc supplementation. BMC Genom. 2010, 11, 553. [Google Scholar] [CrossRef]
- Graham, R.D.; Knez, M.; Welch, R.M. How much nutritional iron deficiency in humans globally is due to an underlying zinc deficiency? In Advances in Agronomy; Elsevier: Amsterdam, The Netherlands, 2012; Volume 115, pp. 1–40. [Google Scholar]
- Knez, M.; Graham, R.D.; Welch, R.M.; Stangoulis, J.C. New perspectives on the regulation of iron absorption via cellular zinc concentrations in humans. Crit. Rev. Food Sci. Nutr. 2017, 57, 2128–2143. [Google Scholar] [CrossRef]
- Solomons, N.W.; Jacob, R.A. Studies on the bioavailability of zinc in humans: Effects of heme and nonheme iron on the absorption of zinc. Am. J. Clin. Nutr. 1981, 34, 475–482. [Google Scholar] [CrossRef]
- Solomons, N.W.; Pineda, O.; Viteri, F.; Sandstead, H.H. Studies on the bioavailability of zinc in humans: Mechanism of the intestinal interaction of nonheme iron and zinc. J. Nutr. 1983, 113, 337–349. [Google Scholar] [CrossRef]
- Sandstrom, B.; Davidsson, L.; Cederblad, A.; Lonnerdal, B. Oral iron, dietary ligands and zinc absorption. J. Nutr. 1985, 115, 411–414. [Google Scholar] [CrossRef]
- Fairweather-Tait, S.J. Iron-zinc and calcium-Fe interactions in relation to Zn and Fe absorption. Proc. Nutr. Soc. 1995, 54, 465–473. [Google Scholar] [CrossRef]
- Davidsson, L.; Almgren, A.; Sandstrom, B.; Hurrell, R.F. Zinc absorption in adult humans: The effect of iron fortification. Br. J. Nutr. 1995, 74, 417–425. [Google Scholar] [CrossRef]
- Harvey, L.J.; Dainty, J.R.; Hollands, W.J.; Bull, V.J.; Hoogewerff, J.A.; Foxall, R.J.; McAnena, L.; Strain, J.J.; Fairweather-Tait, S.J. Effect of high-dose iron supplements on fractional zinc absorption and status in pregnant women. Am. J. Clin. Nutr. 2007, 85, 131–136. [Google Scholar] [CrossRef]
- Rossander-Hulten, L.; Brune, M.; Sandstrom, B.; Lonnerdal, B.; Hallberg, L. Competitive inhibition of iron absorption by manganese and zinc in humans. Am. J. Clin. Nutr. 1991, 54, 152–156. [Google Scholar] [CrossRef]
- Alarcon, K.; Kolsteren, P.W.; Prada, A.M.; Chian, A.M.; Velarde, R.E.; Pecho, I.L.; Hoeree, T.F. Effects of separate delivery of zinc or zinc and vitamin A on hemoglobin response, growth, and diarrhea in young Peruvian children receiving iron therapy for anemia. Am. J. Clin. Nutr. 2004, 80, 1276–1282. [Google Scholar] [CrossRef] [Green Version]
- Dekker, L.H.; Villamor, E. Zinc supplementation in children is not associated with decreases in hemoglobin concentrations. J. Nutr. 2010, 140, 1035–1040. [Google Scholar] [CrossRef]
- Caulfield, L.E.; Zavaleta, N.; Chen, P.; Colombo, J.; Kannass, K. Mineral status of non-anemic Peruvian infants taking an iron and copper syrup with or without zinc from 6 to 18 months of age: A randomized controlled trial. Nutrition 2013, 29, 1336–1341. [Google Scholar] [CrossRef] [Green Version]
- Black, M.M.; Baqui, A.H.; Zaman, K.; Ake Persson, L.; El Arifeen, S.; Le, K.; McNary, S.W.; Parveen, M.; Hamadani, J.D.; Black, R.E. Iron and zinc supplementation promote motor development and exploratory behavior among Bangladeshi infants. Am. J. Clin. Nutr. 2004, 80, 903–910. [Google Scholar] [CrossRef] [Green Version]
- Gunshin, H.; DiRenzo, C.; Fujiwara, Y.; Andrews, N.C. Iron metabolism in the mice with targeted mutations of DMT1 gene in small intestine and erythroid precursors. Gastroenterology 2004, 126, A23. [Google Scholar]
- Tandy, S.; Williams, M.; Leggett, A.; Lopez-Jimenez, M.; Dedes, M.; Ramesh, B.; Srai, S.K.; Sharp, P. Nramp2 expression is associated with pH-dependent iron uptake across the apical membrane of human intestinal Caco-2 cells. J. Biol. Chem. 2000, 275, 1023–1029. [Google Scholar] [CrossRef]
- Troadec, M.B.; Ward, D.M.; Lo, E.; Kaplan, J.; De Domenico, I. Induction of FPN1 transcription by MTF-1 reveals a role for ferroportin in transition metal efflux. Blood 2010, 116, 4657–4664. [Google Scholar] [CrossRef] [Green Version]
- Pinilla-Tenas, J.J.; Sparkman, B.K.; Shawki, A.; Illing, A.C.; Mitchell, C.J.; Zhao, N.; Liuzzi, J.P.; Cousins, R.J.; Knutson, M.D.; Mackenzie, B. Zip14 is a complex broad-scope metal-ion transporter whose functional properties support roles in the cellular uptake of zinc and nontransferrin-bound iron. Am. J. Physiol. Cell. Physiol. 2011, 301, C862–C871. [Google Scholar] [CrossRef] [Green Version]
- Methfessel, A.H.; Spencer, H. Zinc metabolism in the rat. II. Secretion of zinc into intestine. J. Appl. Physiol. 1973, 34, 63–67. [Google Scholar] [CrossRef]
- Sullivan, J.F.; Williams, R.V.; Wisecarver, J.; Etzel, K.; Jetton, M.M.; Magee, D.F. The zinc content of bile an pancreatic juice in zinc-deficient swine. Proc. Soc. Exp. Biol. Med. 1981, 166, 39–43. [Google Scholar] [CrossRef]
- Montgomery, M.L.; Sheline, G.E.; Chaikoff, I.L. The elimination of administered zinc in pancreatic juice, duodenal juice and bile of the Dog as measured by its radioactive isotope. J. Exp. Med. 1943, 78, 151–159. [Google Scholar] [CrossRef]
- Sullivan, J.F.; Burch, R.E.; Quigley, J.H.; Magee, D.F. Zinc deficiency and decreased pancreatic secretory response. Am. J. Physiol. 1974, 227, 105–108. [Google Scholar] [CrossRef]
- Krebs, N.E.; Hambidge, K.M. Zinc metabolism and homeostasis: The application of tracer techniques to human zinc physiology. Biometals 2001, 14, 397–412. [Google Scholar] [CrossRef]
- Pandol, S.J. The Exocrine Pancreas. Colloquium Series on Integrated Systems Physiology: From Molecule to Function; Morgan & Claypool: San Rafael, CA, USA, 2011; Volume 3, pp. 1–64. [Google Scholar]
- Brown, K.H.; Wuehler, S.E.; Peerson, J.M.J.F.; Bulletin, N. The importance of zinc in human nutrition and estimation of the global prevalence of zinc deficiency. Food Nutr. Bull. 2001, 22, 113–125. [Google Scholar] [CrossRef]
- Kilari, S.; Pullakhandam, R.; Nair, K.M. Zinc inhibits oxidative stress-induced iron signaling and apoptosis in Caco-2 cells. Free Radic. Biol. Med. 2010, 48, 961–968. [Google Scholar] [CrossRef]
- King, J.C. Zinc: An essential but elusive nutrient. Am. J. Clin. Nutr. 2011, 94, 679S–684S. [Google Scholar] [CrossRef]
- Lynch, C.J.; Patson, B.J.; Goodman, S.A.; Trapolsi, D.; Kimball, S.R. Zinc stimulates the activity of the insulin-and nutrient-regulated protein kinase mTOR. Am. J. Physiol. Endocrinol. Metab. 2001, 281, E25–E34. [Google Scholar] [CrossRef]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kondaiah, P.; Yaduvanshi, P.S.; Sharp, P.A.; Pullakhandam, R. Iron and Zinc Homeostasis and Interactions: Does Enteric Zinc Excretion Cross-Talk with Intestinal Iron Absorption? Nutrients 2019, 11, 1885. https://doi.org/10.3390/nu11081885
Kondaiah P, Yaduvanshi PS, Sharp PA, Pullakhandam R. Iron and Zinc Homeostasis and Interactions: Does Enteric Zinc Excretion Cross-Talk with Intestinal Iron Absorption? Nutrients. 2019; 11(8):1885. https://doi.org/10.3390/nu11081885
Chicago/Turabian StyleKondaiah, Palsa, Puneeta Singh Yaduvanshi, Paul A Sharp, and Raghu Pullakhandam. 2019. "Iron and Zinc Homeostasis and Interactions: Does Enteric Zinc Excretion Cross-Talk with Intestinal Iron Absorption?" Nutrients 11, no. 8: 1885. https://doi.org/10.3390/nu11081885
APA StyleKondaiah, P., Yaduvanshi, P. S., Sharp, P. A., & Pullakhandam, R. (2019). Iron and Zinc Homeostasis and Interactions: Does Enteric Zinc Excretion Cross-Talk with Intestinal Iron Absorption? Nutrients, 11(8), 1885. https://doi.org/10.3390/nu11081885