Imbalance of Pro- and Anti-Angiogenic Factors Due to Maternal Vitamin D Deficiency Causes Renal Microvasculature Alterations Affecting the Adult Kidney Function

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
2.2. Measurement of Systolic Blood Pressure
2.3. Renal Function Studies
2.4. Determination of the Plasma Angiotensin (ANGII) Level
2.5. Serum Levels of 25(OH)D, Parathyroid Hormone (PTH) and Calcium
2.6. Determination of Renal and Plasma Nitric Oxide (NO) Levels
2.7. Light Microscopy and Morphometric Studies
2.8. Immunohistochemical Studies
2.9. Western Blot Studies
2.10. Statistical Analysis
3. Results
3.1. Body Weight, Fluid Intake, and Food Consumption
3.2. Quantification of 25(OH) D, Parathyroid Hormone (PTH), and Calcium
3.3. Renal Function, Systolic Blood Pressure (SBP), and Urinary Albumin Excretion
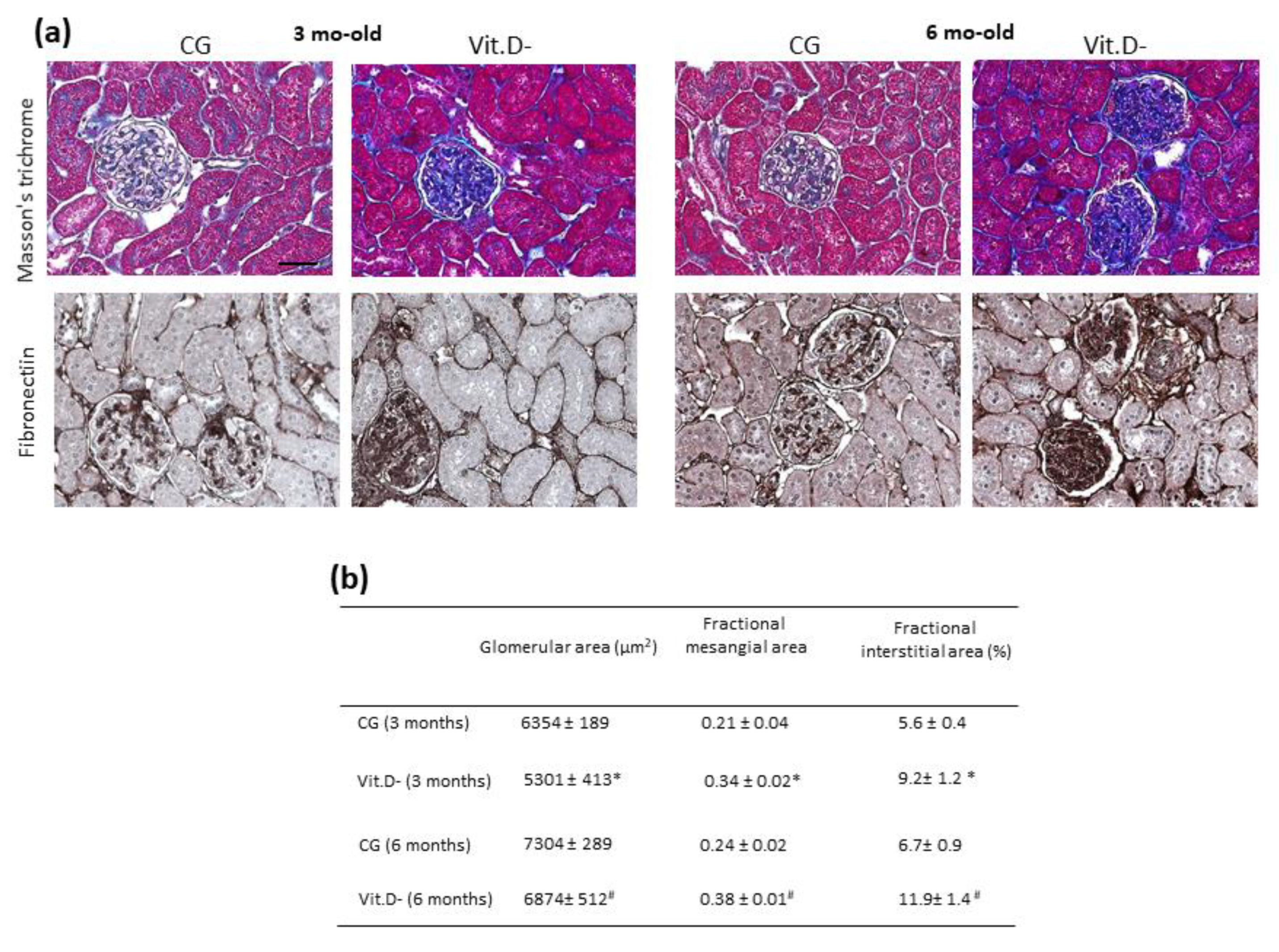
3.4. Histologic and Morphometric Analyses
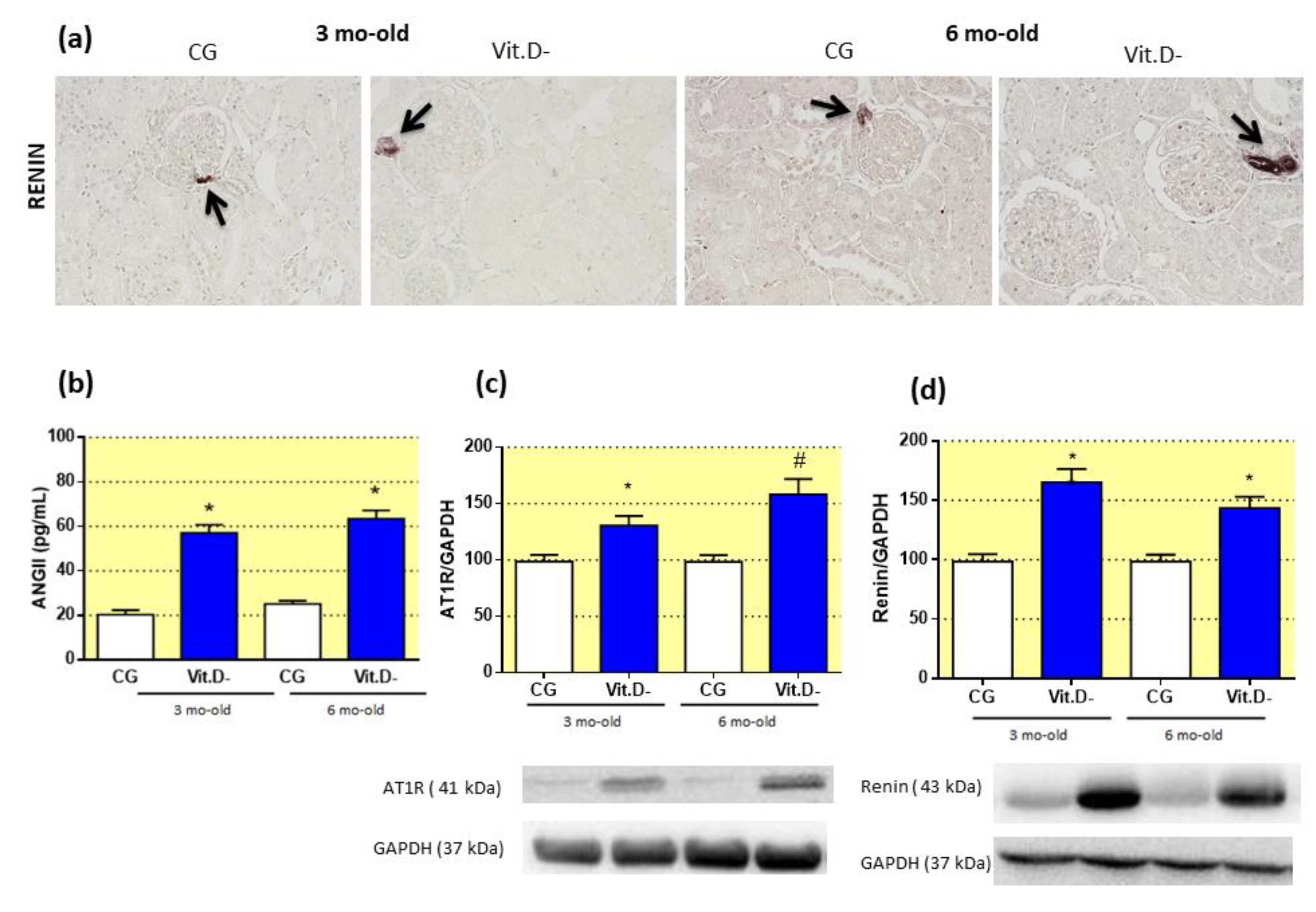
3.5. The Effect of Vitamin D (Vit.D) Deficiency on the Renin-Angiotensin System (RAS)
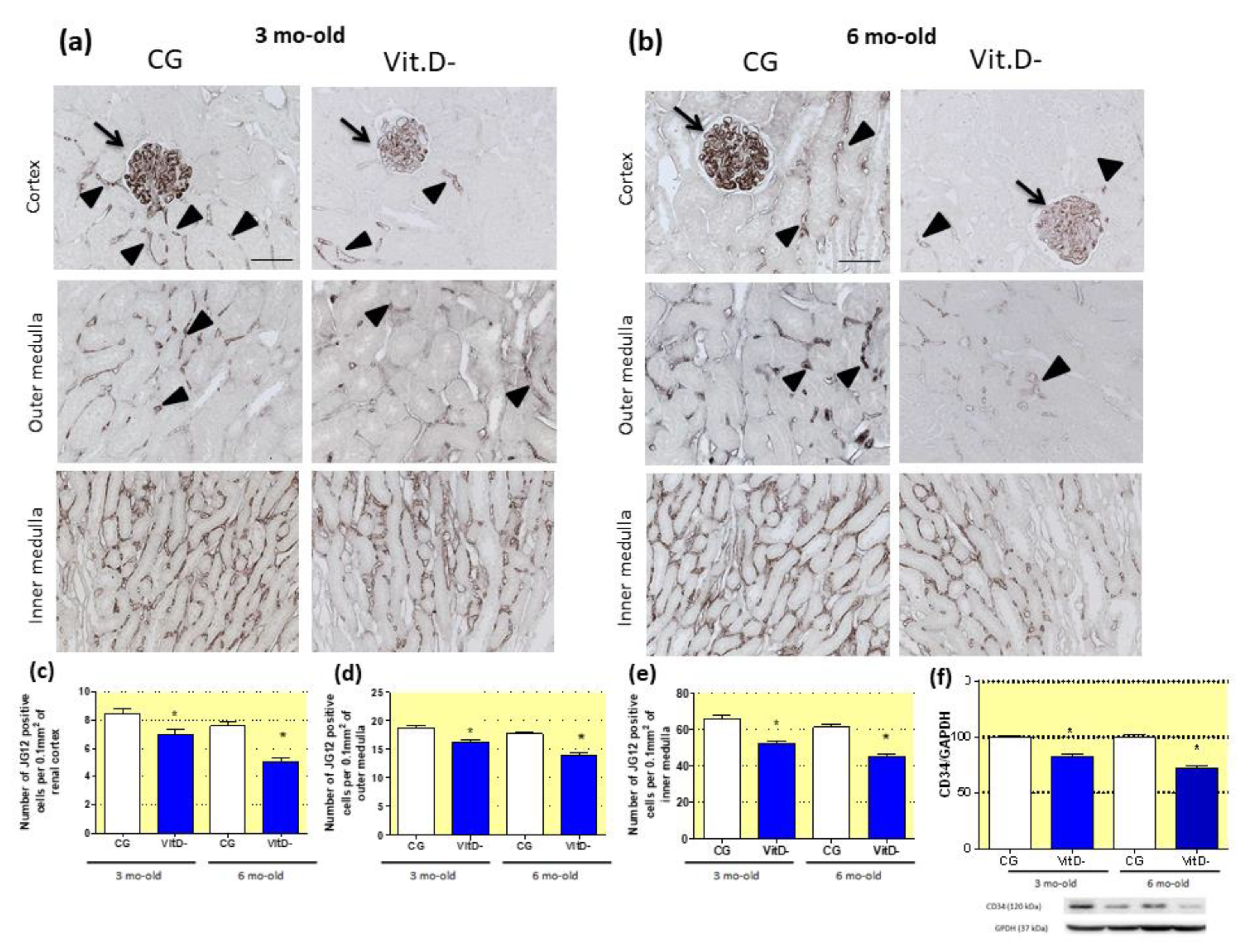
3.6. The Role of Vitamin D (Vit.D) in the Differentiation of the Renal Microvasculature
3.7. Vitamin D (Vit.D) Deficiency and Impairment of the Endothelial Nitric Oxide Synthase-Nitric Oxide (eNOS-NO) System in Adulthood
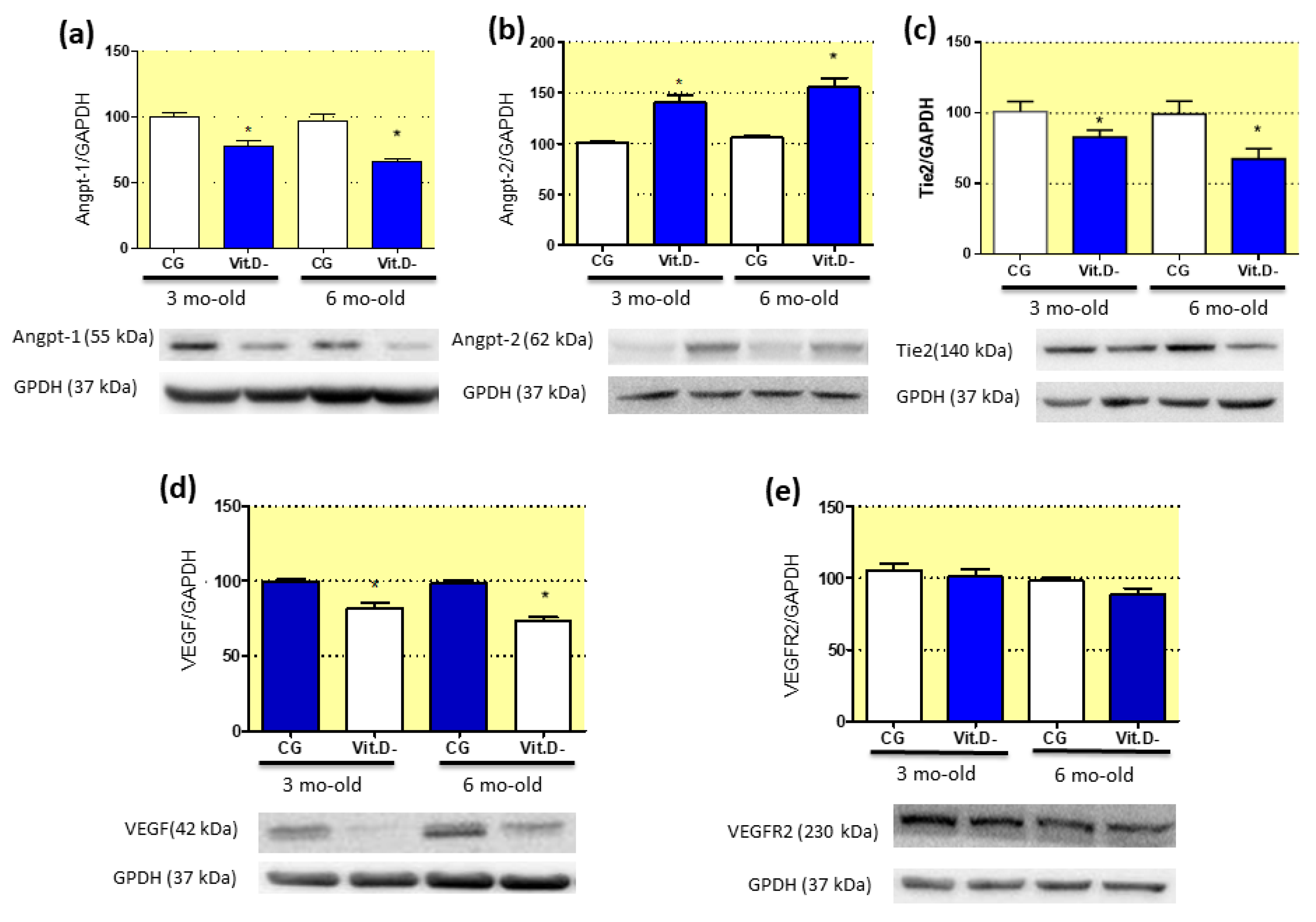
3.8. Imbalanced Pro- and Anti-Angiogenic Factors in Response to Vitamin D (Vit.D) Deficiency
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tare, M.; Emmett, S.J.; Coleman, H.A.; Skordilis, C.; Eyles, D.W.; Morley, R.; Parkington, H.C. Vitamin D insufficiency is associated with impaired vascular endothelial and smooth muscle function and hypertension in young rats. J. Physiol. 2011, 589, 4777–4786. [Google Scholar] [CrossRef] [PubMed]
- Argeri, R.; Nishi, E.E.; Volpini, R.A.; Palma, B.D.; Tufik, S.; Gomes, G.N. Sleep restriction during pregnancy and its effects on blood pressure and renal function among female offspring. Physiol. Rep. 2016, 4, e12888. [Google Scholar] [CrossRef] [PubMed]
- de Almeida, L.F.; Francescato, H.D.C.; da Silva, C.G.A.; Costa, R.S.; Coimbra, T.M. Calcitriol reduces kidney development disorders in rats provoked by losartan administration during lactation. Sci. Rep. 2017, 7, 11472. [Google Scholar] [CrossRef] [PubMed]
- Wen, J.; Hong, Q.; Wang, X.; Zhu, L.; Wu, T.; Xu, P.; Fu, Z.; You, L.; Wang, X.; Ji, C.; et al. The effect of maternal vitamin D deficiency during pregnancy on body fat and adipogenesis in rat offspring. Sci. Rep. 2018, 8, 365. [Google Scholar] [CrossRef] [PubMed]
- Hales, C.N.; Barker, D.J.P. Type 2 (non-insulin-dependent) diabetes mellitus: The thrifty phenotype hypothesis. Diabetologia 1992, 35, 590–601. [Google Scholar] [CrossRef]
- Simmons, R. Developmental origins of adult metabolic disease: Concepts and controversies. Trends Endocrinol. Metab. 2005, 16, 390–394. [Google Scholar] [CrossRef] [PubMed]
- Manios, Y.; Moschonis, G.; Lambrinou, C.P.; Tsoutsoulopoulou, K.; Binou, P.; Karachaliou, A.; Breidenassel, C.; Gonzalez-Gross, M.; Kiely, M.; Cashman, K.D. A systemic review of vitamin D status in southern European countries. Eur. J. Nutr. 2018, 57, 2001–2036. [Google Scholar] [CrossRef]
- Holick, M.F. The vitamin D deficiency pandemic: Approaches for diagnosis, treatment and prevention. Rev. Endocr. Metab. Disord. 2017, 18, 153–165. [Google Scholar] [CrossRef]
- Mulligan, M.L.; Felton, S.K.; Riek, A.E.; Bernal-Mizrachi, C. Implications of vitamin D deficiency in pregnancy and lactation. Am. J. Obstet. Gynecol. 2010, 202, 1–9. [Google Scholar] [CrossRef]
- de Almeida, L.F.; Coimbra, T.M. When Less or More Isn’t Enough: Renal Maldevelopment Arising From Disequilibrium in the Renin-Angiotensin System. Front Pediatr 2019, 7, 296. [Google Scholar] [CrossRef]
- Freundlich, M.; Quiroz, Y.; Zhang, Z.; Zhang, Y.; Bravo, Y.; Weisinger, J.R.; Li, Y.C. Rodriguez-Iturbe, B. Suppression of renin–angiotensin gene expression in the kidney by paricalcitol. Kidney Int. 2008, 74, 1394–1402. [Google Scholar] [CrossRef] [PubMed]
- Ramagopalan, S.V.; Heger, A.; Berlanga, A.J.; Maugeri, N.J.; Lincoln, M.R.; Burrell, A.; Handunnetthi, L.; Handel, A.E.; Disanto, G.; Orton, S.M.; et al. A ChIP-seq defined genome-wide map of vitamin D receptor binding: Associations with disease and evolution. Genome Res. 2010, 20, 1352–1360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Yang, R.; Qiao, W.; Zhang, W.; Chen, J.; Mao, L.; Goltzman, D.; Miao, D. 1,25-Dihydroxyvitamin D exerts an antiaging role by activation of Nrf2-antioxidant signaling and inactivation of p16/p53-senescence signaling. Aging Cell 2019, 18, e12951. [Google Scholar] [CrossRef] [PubMed]
- Maka, N.; Makrakis, J.; Parkington, H.C.; Tare, M.; Morley, R.; Black, M.J. Vitamin D deficiency during pregnancy and lactation stimulates nephrogenesis in rat offspring. Pediatric Nephrol. 2007, 23, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Boyce, A.C.; Palmer-Aronsten, B.J.; Kim, M.Y.; Gibson, K.J. Maternal vitamin D deficiency programmes adult renal renin gene expression and renal function. J. Dev. Orig. Health Dis. 2013, 4, 368–376. [Google Scholar] [CrossRef] [PubMed]
- Andersen, L.B.; Przybyl, L.; Haase, N.; von Versen-Höynck, F.; Qadri, F.; Jørgensen, J.S.; Sorensen, G.L.; Fruekilde, P.; Poglitsch, M.; Szijarto, I.; et al. Vitamin D depletion aggravates hypertension and target-organ damage. J. Am. Heart Assoc. 2015, 4, e001417. [Google Scholar] [CrossRef] [PubMed]
- Machado, F.G.; Poppi, E.P.B.; Fanelli, C.; Malheiros, D.M.A.C.; Zatz, R.; Fujihara, C.K. AT1blockade during lactation as a model of chronic nephropathy: Mechanisms of renal injury. Am. J. Physiol. Ren. Physiol. 2008, 294, F1345–F1353. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, F.A.M.; Ceciliano, T.C.; Aguila, M.B.; Mandarim-de-Lacerda, C.A. Maternal vitamin D deficiency delays glomerular maturity in F1 and F2 offspring. PLoS ONE 2012, 7, e41740. [Google Scholar] [CrossRef]
- Marin, E.C.S.; Francescato, H.D.C.; Costa, R.S.; da Silva, C.G.A.; Coimbra, T.M. The role of oxidative stress in renal injury induced in rats by losartan exposure during lactation. J. Renin Angiotensin Aldosterone Syst. 2013, 15, 362–377. [Google Scholar] [CrossRef]
- Almeida, L.F.; Francescato, H.D.C.; Silva, R.S.; Silva, C.G.A.; Antunes-Rodrigues, J.; de Paula, F.J.A.; Coimbra, T.M. Renal developmental disturbances and their long-term consequences in female pups from vitamin D-deficient mothers: Involved mechanisms. J. Dev. Orig. Health Dis. 2019, 6, 1–5. [Google Scholar] [CrossRef]
- Lobov, I.B.; Brooks, P.C.; Lang, R.A. Angiopoietin-2 displays VEGF-dependent modulation of capillary structure and endothelial cell survival in vivo. Proc. Natl. Acad. Sci. USA 2002, 99, 11205–11210. [Google Scholar] [CrossRef] [PubMed]
- Yancopoulos, G.D.; Davis, S.; Gale, N.W.; Rudge, J.S.; Wiegand, S.J.H. Vascular-specific growth factors and blood vessel formation. Nature 2000, 407, 242–248. [Google Scholar] [CrossRef] [PubMed]
- Potente, M.; Gerhardt, H.; Carmeliet, P. Basic and therapeutic aspects of angiogenesis. Cell 2011, 146, 873–887. [Google Scholar] [CrossRef] [PubMed]
- Augustin, H.G.; Koh, G.Y.; Thurston, G.; Alitalo, K. Control of vascular morphogenesis and homeostasis through the angiopoietin–Tie system. Nat. Rev. Mol. Cell Biol. 2009, 10, 165–177. [Google Scholar] [CrossRef] [PubMed]
- Otani, A.; Takagi, H.; Oh, H.; Koyama, S.; Honda, Y. Angiotensin II induces expression of the Tie2 receptor ligand, angiopoietin-2, in bovine retinal endothelial cells. Diabetes 2001, 50, 867–875. [Google Scholar] [CrossRef]
- Marcondes, F.K.; Bianchi, F.J.; Tanno, A.P. Determination of the estrous cycle phases of rats: Some helpful considerations. Braz. J. Biol. 2002, 62, 609–614. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, V.; Quiroz, Y.; Nava, M.; Pons, H.; Rodríguez-Iturbe, B. Overload proteinuria is followed by salt-sensitive hypertension caused by renal infiltration of immune cells. Am. J. Physiol. Ren. Physiol. 2002, 283, F1132–F1141. [Google Scholar] [CrossRef] [Green Version]
- Laurell, C.B. Electroimmuno assay. Scand. J. Clin. Lab. Investig. 1972, 29, 21–37. [Google Scholar] [CrossRef]
- Botelho, L.M.O.; Block, C.H.; Khosla, M.C.; Santos, R.A.S. Plasma angiotensin (1–7) immunoreactivity is increased by salt load, water deprivation, and hemorrhage. Peptides 1994, 15, 723–729. [Google Scholar] [CrossRef]
- Bradford, M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Baroni, E.A.; Costa, R.S.; Volpini, R.; Coimbra, T.M. Sodium bicarbonate treatment reduces renal injury, renal production of transforming growth factor-β and urinary transforming growth factor-β excretion in rats with doxorubicin-induced nephropathy. Am. J. Kidney Dis. 1999, 34, 328–337. [Google Scholar] [CrossRef]
- Coimbra, T.M.; Janssen, U.; Gröne, H.J.; Ostendorf, T.; Kunter, U.; Schmidt, H.; Brabant, G.; Floege, J. Early events leading to renal injury in obese zucker (fatty) rats with type II diabetes. Kidney Int. 2000, 57, 167–182. [Google Scholar] [CrossRef] [PubMed]
- Balbi, A.P.C.; Marin, E.C.S.; Francescato, H.D.C.; Costa, R.S.; Coimbra, T.M. MAPK and angiotensin II receptor in kidney of newborn rats from losartan-treated dams. Pediatric Nephrol. 2008, 23, 1433–1444. [Google Scholar] [CrossRef] [PubMed]
- Tarcin, O.; Yavuz, D.G.; Ozben, B.; Telli, A.; Ogunc, A.V.; Yuksel, M.; Toprak, A.; Yazici, D.; Sancak, S.; Deyneli, O.; et al. Effect of vitamin D deficiency and replacement on endothelial function in asymptomatic subjects. J. Clin. Endocrinol. Metab. 2009, 94, 4023–4030. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Yadav, A.K.; Lal, A.; Kumar, V.; Singhal, M.; Billot, L.; Gupta, K.L.; Banerjee, D.; Jha, V. A randomized trial of vitamin D supplementation on vascular function in CKD. J. Am. Soc. Nephrol. 2017, 28, 3100–3108. [Google Scholar] [CrossRef]
- Faleiros, C.M.; Francescato, H.D.C.; Papoti, M.; Chaves, L.; Silva, C.G.A.; Costa, R.S.; Coimbra, T.M. Effects of previous physical training on adriamycin nephropathy and its relationship with endothelial lesions and angiogenesis in the renal cortex. Life Sci. 2017, 169, 43–51. [Google Scholar] [CrossRef]
- Zakharova, I.; Klimov, L.; Kuryaninova, V.; Nikitina, I.; Malyavskaya, S.; Dolbnya, S.; Kasyanova, A.; Atanesyan, R.; Stoyan, M.; Todieva, A.; et al. Vitamin D insufficiency in overweight and obese children and adolescents. Front. Endocrinol. 2019, 10, 103. [Google Scholar] [CrossRef]
- Morley, R.; Carlin, J.B.; Pasco, J.A.; Wark, J.D. Maternal 25-hydroxyvitamin D and parathyroid hormone concentrations and offspring birth size. J. Clin. Endocrinol. Metab. 2006, 91, 906–912. [Google Scholar] [CrossRef]
- Skorecki, K.; Chertow, G.M.; Marsden, A.P.; Tall, M.W.; Yu, A.S.L. Brenner & Rector’s the Kidney; Elsevier: Philadelphia, PA, USA, 2016. [Google Scholar]
- Hackenthal, E.; Paul, M.; Ganten, D.; Taugner, R. Morphology, physiology and molecular biology of renin secretion. Physiol. Rev. 1990, 70, 1067–1116. [Google Scholar] [CrossRef]
- Fitzsimons, J.T. Angiotensin stimulation of the central nervous system. Rev. Physiol. Biochem. Pharmacol. 1980, 87, 117–167. [Google Scholar]
- Li, Y.C.; Kong, J.; Wei, M.; Chen, Z.F.; Liu, S.Q.; Cao, L.P. 1,25-Dihydroxyvitamin D3 is a negative endocrine regulator of the renin-angiotensin system. J. Clin. Investig. 2002, 110, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Lu, F.; Cao, K.; Xu, D.; Goltzman, D.; Miao, D. Calcium-independent and 1,25(OH)2D3-dependent regulation of the renin-angiotensin system in 1α-hydroxylase knockout mice. Kidney Int. 2008, 74, 170–179. [Google Scholar] [CrossRef] [PubMed]
- Madsen, K.; Tinning, A.R.; Marcussen, N.; Jensen, B.L. Postnatal development of the renal medulla; role of the renin-angiotensin system. Acta Physiol. 2013, 208, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Roman, R.J.; Kaldunski, M.L. Renal cortical and papillary blood flow in spontaneously hypertensive rats. Hypertension 1988, 11, 657–663. [Google Scholar] [CrossRef] [PubMed]
- Chitalia, N.; Ismail, T.; Tooth, L.; Boa, F.; Hampson, G.; Goldsmith, D.; Kaski, J.C.; Banerjee, D. Impact of vitamin D supplementation on arterial vasomotion, stiffness and endothelial biomarkers in chronic kidney disease patients. PLoS ONE 2014, 9, e91363. [Google Scholar] [CrossRef] [PubMed]
- Mollnau, H.; Wendt, M.; Szöcs, K.; Lassègue, B.; Schulz, E.; Oelze, M.; Li, H.; Bodenschatz, M.; August, M.; Kleschyov, A.L.; et al. Effects of angiotensin II infusion on the expression and function of NAD(P)H oxidase and components of nitric oxide/cGMP signaling. Circ. Res. 2002, 90, E58–E65. [Google Scholar] [CrossRef]
- Kida, Y.; Tchao, B.N.; Yamaguchi, I. Peritubular capillary rarefaction: A new therapeutic target in chronic kidney disease. Pediatric Nephrol. 2014, 29, 333–342. [Google Scholar] [CrossRef] [PubMed]
- Rizkalla, B. Increased renal vascular endothelial growth factor and angiopoietins by angiotensin II infusion is mediated by both AT1 and AT2 receptors. J. Am. Soc. Nephrol. 2003, 14, 3061–3071. [Google Scholar] [CrossRef]
- Hanahan, D.; Folkman, J. Patterns and emerging mechanisms of the angiogenic switch during tumorigenesis. Cell 1996, 86, 353–364. [Google Scholar] [CrossRef]
- Yoo, K.H.; Yim, H.E.; Bae, E.S.; Hong, Y.S. Capillary rarefaction and altered renal development: The imbalance between pro- and anti-angiogenic factors in response to angiotensin II inhibition in the developing rat kidney. J. Mol. Histol. 2018, 49, 219–228. [Google Scholar] [CrossRef]
- Biel, N.M.; Siemann, D.W. Targeting the angiopoietin-2/Tie-2 axis in conjunction with VEGF signal interference. Cancer Lett. 2016, 380, 525–533. [Google Scholar] [CrossRef] [PubMed]
- Zimanyi, M.A.; Bertram, J.F.; Black, M.J. Does a nephron deficit in rats predispose to salt-sensitive hypertension? Kidney Blood Press. Res. 2004, 27, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Zmijewski, M.A. Vitamin D and human health. Int. J. Mol. Sci. 2019, 20, 145. [Google Scholar] [CrossRef] [PubMed]
- Gaudio, A.; Fiore, V.; Rapisarda, R.; Sidoti, M.H.; Xourafa, A.; Catalano, A.; Tringali, G.; Zanoli, L.; Signorelli, S.S.; Fiore, C.E. Sclerostin is a possible candidate marker of arterial stiffness: Results from a cohort study in Catania. Mol. Med. Rep. 2017, 15, 3420–3424. [Google Scholar] [CrossRef] [PubMed]
- Evenepoel, P.; D’Haese, P.; Brandenburg, V. Sclerostin and DKK1: New players in renal bone and vascular disease. Kidney Int. 2015, 88, 235–240. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nutrient (g/kg) | Diet | |
|---|---|---|
| CG | Vit.D- | |
| Corn starch | 397.50 | 397.50 |
| Casein | 200.00 | 200.00 |
| Dextrinated starch | 132.00 | 132.00 |
| Sucrose | 100.00 | 100.00 |
| Soya bean oil | 70 | 70 |
| L- Cystine | 3.00 | 3.00 |
| Choline | 2.50 | 2.50 |
| Mineral mix | 35.00 | 35.00 |
| Calcium carbonate | 357.00 | 357.00 |
| Vitamin mix | 10.00 | 10.00 |
| Vitamin D3 | 0.25 | 0.00 |
| Fibre | 50.00 | 50.00 |
| 3 Months | 6 Months | |||
|---|---|---|---|---|
| Control | Vit.D- | Control | Vit.D- | |
| n = 7; m = 3 | n = 10; m = 3 | n = 7; m = 3 | n = 9; m = 3 | |
| Body weight (g) | 358 ± 18 | 401 ± 15 * | 448 ± 19 | 498 ± 13 * |
| Food intake (mg 100 g−1) | 8.6 ± 1.2 | 11.9 ± 1.2 * | 10.3 ± 1.2 | 13.6 ± 1.0 * |
| Water intake (mL 100 g−1) | 8.3 ± 1.2 | 11.3 ± 1.8 * | 9.8 ± 1.2 | 13.9 ± 1.2 * |
| V (mL 100 g−1) | 6.3 ± 1.8 | 8.5 ± 2.45 * | 7.2 ± 1.8 | 9.8 ± 2.3 * |
| U (mOsm kg H2O−1) | 1.458 ± 135 | 1.270 ± 108 * | 1.711 ± 145 | 1.250 ± 129 * |
| U Na+/Cr | 2.8 ± 0.8 | 3.4 ± 0.5 * | 2.9 ± 0.6 | 3.9 ± 0.7 * |
| U K+/Cr | 3.6 ± 0.3 | 4.3 ± 0.4* | 3.7 ± 0.3 | 4.5 ± 0.4* |
| SBP (mmHg) | 121 ± 1.3 | 133 ± 1.8 * | 123 ± 1.2 | 141 ± 1.6 * |
| ALB (µg 24 h−1) | 0.4 (0.3; 0.6) | 0.6 (0.3; 0.7) | 0.6 (0.4; 0.6) | 0.7 (0.5; 0.8) |
| 3 Months | 6 Months | |||
|---|---|---|---|---|
| Control | Vit.D- | Control | Vit.D- | |
| n = 7; m = 3 | n = 10; m = 3 | n = 7; m = 3 | n = 9; m = 3 | |
| Serum 25(OH)D3 (nmol L−1) | 98.4 ± 2.2 | 62.9 ± 2.5 * | 87.6 ± 1.9 | 57.9 ± 1.5 * |
| Serum Ca2+ (nmol L−1) | 2.37 ± 0.4 | 2.25 ± 0.3 | 2.33 ± 0.2 | 2.27 ± 0.3 |
| PTH (pg/mL) | 165.4 ± 28.59 | 178.8 ± 39.34 | 144.3 ± 33.11 | 197.8 ± 113.1 |
| Pcreat (mg/dL) | 0.69 ± (0.53; 0.72) | 0.72 ± (0.54; 0.74) | 0.79 ± (0.53; 0.82) | 0.82 ± (0.51; 0.84) |
| Na+ (mmol/L) | 147.1 ± 3.2 | 148.3 ± 2.5 | 146.7 ± 3.9 | 148.9 ± 3.5 |
| K+ (mmol/L) | 4.1 ± 0.5 | 4.6 ± 0.7 | 4.9 ± 0.8 | 4.7 ± 0.3 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferreira de Almeida, L.; Della Coletta Francescato, H.; Antunes-Rodrigues, J.; Jose Albuquerque de Paula, F.; Giovanni Alves da Silva, C.; Silva Costa, R.; Machado Coimbra, T. Imbalance of Pro- and Anti-Angiogenic Factors Due to Maternal Vitamin D Deficiency Causes Renal Microvasculature Alterations Affecting the Adult Kidney Function. Nutrients 2019, 11, 1929. https://doi.org/10.3390/nu11081929
Ferreira de Almeida L, Della Coletta Francescato H, Antunes-Rodrigues J, Jose Albuquerque de Paula F, Giovanni Alves da Silva C, Silva Costa R, Machado Coimbra T. Imbalance of Pro- and Anti-Angiogenic Factors Due to Maternal Vitamin D Deficiency Causes Renal Microvasculature Alterations Affecting the Adult Kidney Function. Nutrients. 2019; 11(8):1929. https://doi.org/10.3390/nu11081929
Chicago/Turabian StyleFerreira de Almeida, Lucas, Heloísa Della Coletta Francescato, Jose Antunes-Rodrigues, Francisco Jose Albuquerque de Paula, Cleonice Giovanni Alves da Silva, Roberto Silva Costa, and Terezila Machado Coimbra. 2019. "Imbalance of Pro- and Anti-Angiogenic Factors Due to Maternal Vitamin D Deficiency Causes Renal Microvasculature Alterations Affecting the Adult Kidney Function" Nutrients 11, no. 8: 1929. https://doi.org/10.3390/nu11081929
APA StyleFerreira de Almeida, L., Della Coletta Francescato, H., Antunes-Rodrigues, J., Jose Albuquerque de Paula, F., Giovanni Alves da Silva, C., Silva Costa, R., & Machado Coimbra, T. (2019). Imbalance of Pro- and Anti-Angiogenic Factors Due to Maternal Vitamin D Deficiency Causes Renal Microvasculature Alterations Affecting the Adult Kidney Function. Nutrients, 11(8), 1929. https://doi.org/10.3390/nu11081929