Choline Intake as Supplement or as a Component of Eggs Increases Plasma Choline and Reduces Interleukin-6 without Modifying Plasma Cholesterol in Participants with Metabolic Syndrome


 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Dietary and Exercise Records
2.3. Anthropometric Measurements and Blood Pressure
2.4. Plasma Lipids, Glucose, C-Reactive Protein (CRP), Creatinine, and Liver Enzymes
2.5. Plasma Choline
2.6. Plasma Insulin and Insulin Resistance
2.7. Inflammatory Markers
2.8. Statistical Analysis
3. Results
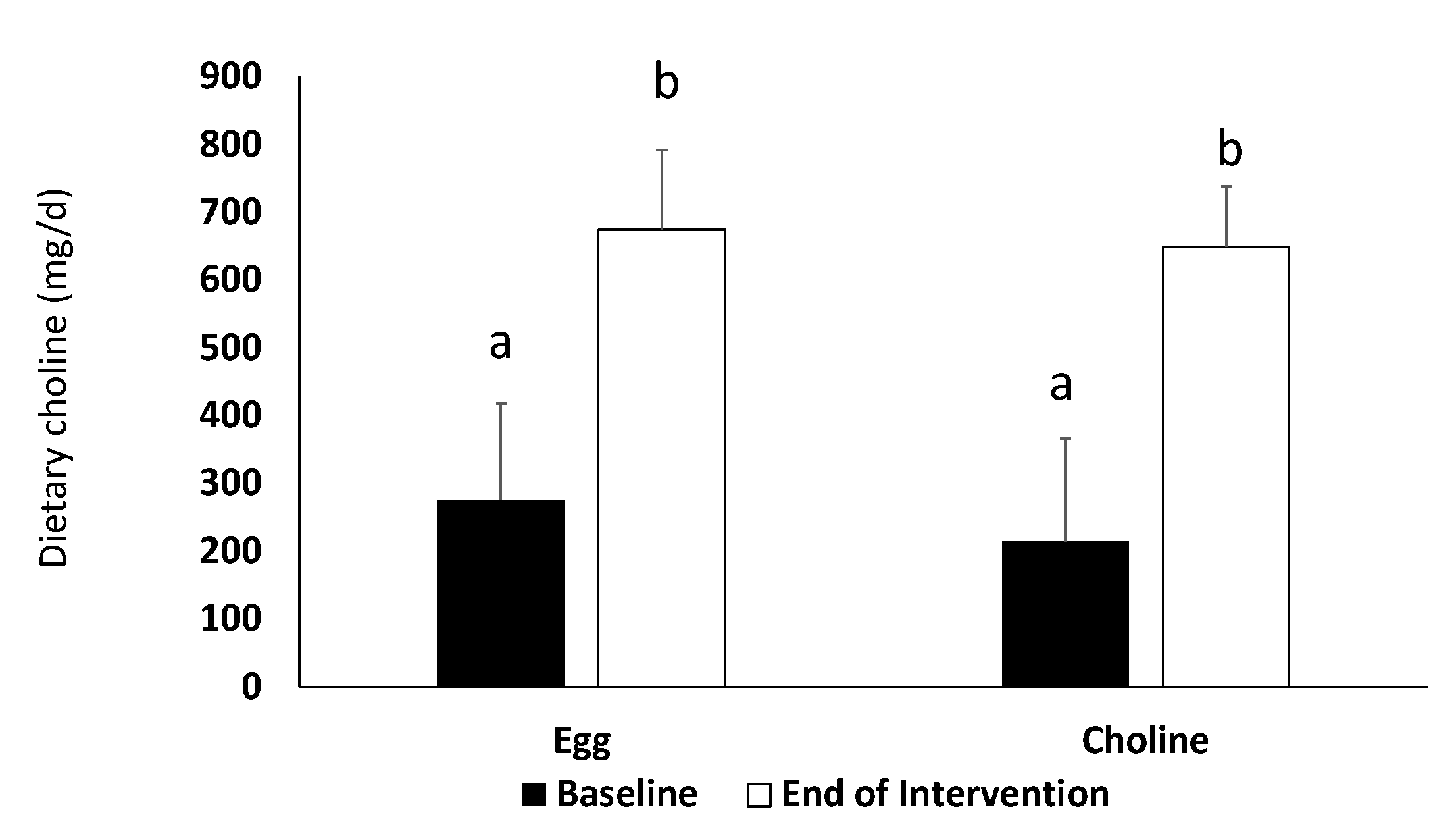
3.1. Dietary and Exercise Records
3.2. Initial Characteristics of Subjects
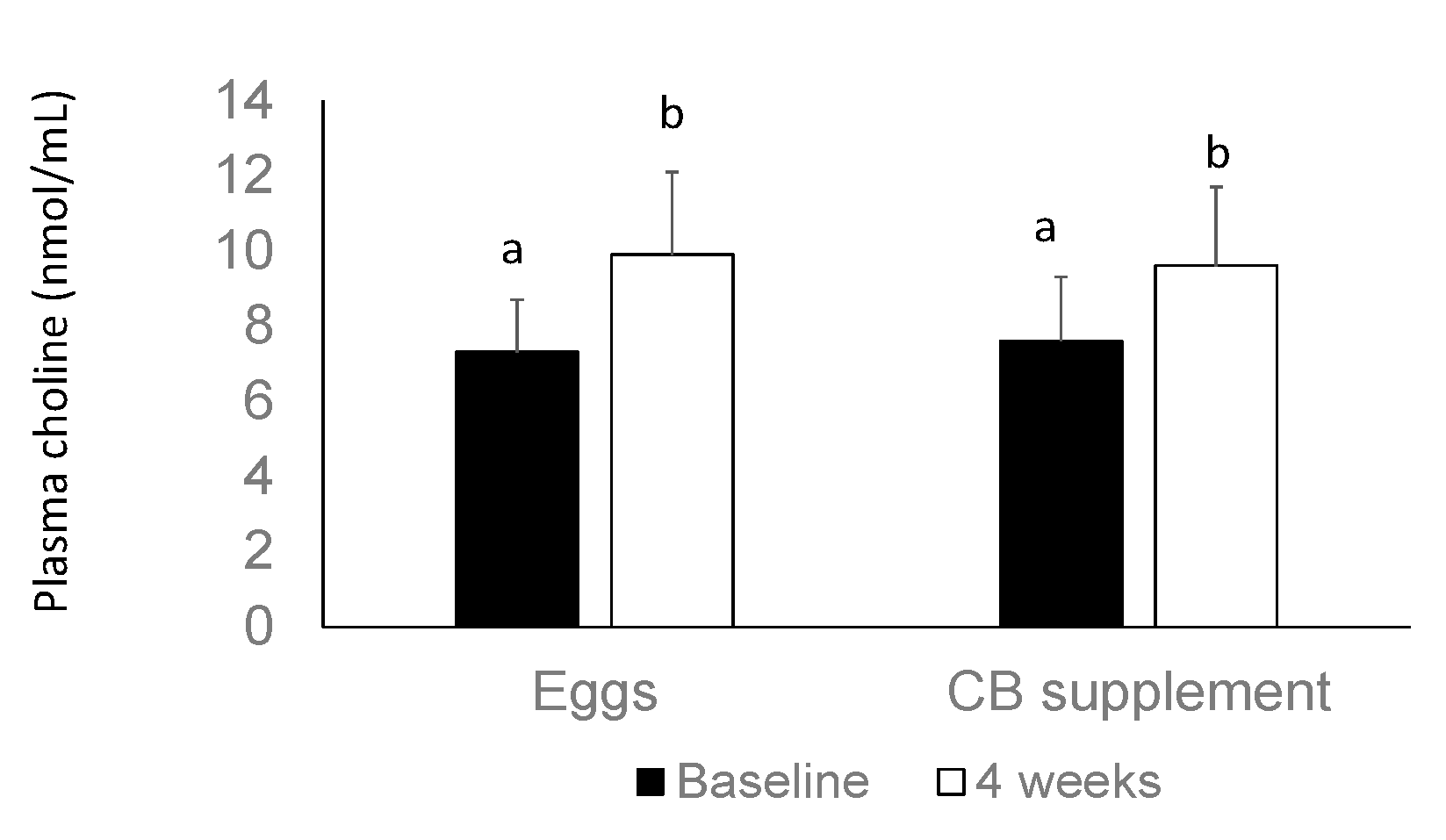
3.3. Plasma Choline Concentrations
3.4. Anthropometrics, CRP, and Liver Enzymes
3.5. Plasma Lipids and Glucose
3.6. Plasma Choline
3.7. Inflammatory Markers
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Grundy, S.M.; Williams, C.; Vega, G.L. Upper body fat predicts metabolic syndrome similarly in men and women. Eur. J. Clin. Investig. 2018, 48, e12941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drewnowski, A. The Nutrient Rich Foods Index helps to identify healthy, affordable foods. Am. J. Clin. Nutr. 2010, 91, 1095S–1101S. [Google Scholar] [CrossRef] [PubMed]
- Rehault, S.M.; Guyot, N.; Nys, Y. The Golden Egg: Nutritional Value, Bioactivities, and Emerging Benefits for Human Health. Nutrients 2019, 11, 684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wal, J.S.V.; Gupta, A.; Khosla, P.; Dhurandhar, N.V. Egg breakfast enhances weight loss. Int. J. Obes. (Lond.) 2008, 32, 1545–1551. [Google Scholar] [CrossRef] [Green Version]
- Volek, J.S.; Feinman, R.D. Carbohydrate restriction improves the features of Metabolic Syndrome. Metabolic Syndrome may be defined by the response to carbohydrate restriction. Nutr. Metab. 2005, 2, 31. [Google Scholar] [CrossRef] [Green Version]
- Zhong, V.W.; Van Horn, L.; Cornelis, M.C.; Wilkins, J.T.; Ning, H.; Carnethon, M.R.; Greenland, P.; Mentz, R.J.; Tucker, K.L.; Zhao, L.; et al. Associations of Dietary Cholesterol or Egg Consumption With Incident Cardiovascular Disease and Mortality. JAMA 2019, 321, 1081–1095. [Google Scholar] [CrossRef]
- Khawaja, O.; Singh, H.; Luni, F.; Kabour, A.; Ali, S.S.; Taleb, M.; Ahmed, H.; Gaziano, J.M.; Djoussé, L. Egg Consumption and Incidence of Heart Failure: A Meta-Analysis of Prospective Cohort Studies. Front. Nutr. 2017, 4, 10. [Google Scholar] [CrossRef] [Green Version]
- Blesso, C.N.; Andersen, C.J.; Barona, J.; Volek, J.S.; Fernandez, M.L. Whole egg consumption improves lipoprotein profiles and insulin sensitivity to a greater extent than yolk-free egg substitute in individuals with metabolic syndrome. Metab. Clin Exper. 2013, 62, 400–410. [Google Scholar] [CrossRef]
- DiMarco, D.M.; Missimer, A.; Murillo, A.G.; Lemos, B.S.; Malysheva, O.V.; Caudill, M.A.; Blesso, C.N.; Fernandez, M.L. Intake of up to 3 Eggs/Day Increases HDL Cholesterol and Plasma Choline While Plasma Trimethylamine-N-oxide is Unchanged in a Healthy Population. Lipids 2017, 52, 255–263. [Google Scholar] [CrossRef]
- Berger, S.; Raman, G.; Vishwanathan, R.; Jacques, P.F.; Johnson, E.J. Dietary cholesterol and cardiovascular disease: A systematic review and meta-analysis. Am. J. Clin. Nutr. 2015, 102, 276–294. [Google Scholar] [CrossRef] [Green Version]
- Vincent, M.J.; Allen, B.; Palacios, O.M.; Haber, L.T.; Maki, K.C. Meta-regression analysis of the effects of dietary cholesterol intake on LDL and HDL cholesterol. Am. J. Clin. Nutr. 2018, 109, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Kuang, H.; Yang, F.; Zhang, Y.; Wang, T.; Chen, G. The Impact of Egg Nutrient Composition and Its Consumption on Cholesterol Homeostasis. Cholesterol 2018, 2018, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Carson, J.A.S.; Lichtenstein, A.H.; Anderson, C.A.; Appel, L.J.; Kris-Etherton, P.M.; Meyer, K.A.; Petersen, K.; Polonsky, T.; Van Horn, L. Dietary Cholesterol and Cardiovascular Risk: A Science Advisory From the American Heart Association. Circulation 2020, 141, e39–e53. [Google Scholar] [CrossRef] [Green Version]
- Soliman, A.G. Dietary Cholesterol and the Lack of Evidence in Cardiovascular Disease. Nutrients 2018, 10, 780. [Google Scholar] [CrossRef] [Green Version]
- Mistry, P.; Miller, N.E.; Laker, M.; Hazzard, W.R.; Lewis, B. Individual Variation in the Effects of Dietary Cholesterol on Plasma Lipoproteins and Cellular Cholesterol Homeostasis in Man, Studies of low density lipoprotein receptor activity and 3-hydroxy-3-methylglutaryl coenzyme a reductase activity in blood mononuclear cells. J. Clin. Investig. 1981, 67, 493–502. [Google Scholar] [CrossRef]
- McNamara, D.J.; Kolb, R.; Parker, T.S.; Batwin, H.; Samuel, P.; Brown, C.D.; Ahrens, E.H. Heterogeneity of cholesterol homeostasis in man. Response to changes in dietary fat quality and cholesterol quantity. J. Clin. Investig. 1987, 79, 1729–1739. [Google Scholar] [CrossRef] [PubMed]
- Berg, J.M.; Tymoczko, J.L.; Stryer, L. The Complex Regulation of Cholesterol Biosynthesis Takes Place at Several Levels. In Biochemistry, 5th ed.; Section 26.3; W H Freeman: New York, NY, USA, 2002. [Google Scholar]
- Afonso, M.S.; Machado, R.M.; Lottenberg, A.M.; Quintão, E.C.R.; Moore, K.J.; Lottenberg, A.M. Molecular Pathways Underlying Cholesterol Homeostasis. Nutrients 2018, 10, 760. [Google Scholar] [CrossRef] [Green Version]
- Blesso, C.N. Egg Phospholipids and Cardiovascular Health. Nutrients 2015, 7, 2731–2747. [Google Scholar] [CrossRef] [Green Version]
- U.S. Department of Health and Human Services and U.S. Department of Agriculture. 2015–2020 Dietary Guidelines for Americans, 8th ed. December 2015. Available online: https://health.gov/our-work/food-and-nutrition/2015-2020-dietary-guidelines/ (accessed on 31 August 2020).
- Zeisel, S.H.; Da Costa, K.-A. Choline: An essential nutrient for public health. Nutr. Rev. 2009, 67, 615–623. [Google Scholar] [CrossRef] [Green Version]
- Detopoulou, P.; Panagiotakos, D.B.; Antonopoulou, S.; Pitsavos, C.; Stefanadis, C. Dietary choline and betaine intakes in relation to concentrations of inflammatory markers in healthy adults: The ATTICA study. Am. J. Clin. Nutr. 2008, 87, 424–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallace, T.C.; Fulgoni, V.L. Assessment of Total Choline Intakes in the United States. J. Am. Coll. Nutr. 2016, 35, 108–112. [Google Scholar] [CrossRef] [PubMed]
- Institute of Medicine Dietary Reference Intakes for Thiamin, Riboflavin, Niacin, Vitamin B6, Folate, Vitamin B12, Pantothenic Acid, Biotin, and Choline. Dietary Reference Intakes for Thiamin, Riboflavin, Niacin, Vitamin B6, Folate, Vitamin B12, Pantothenic Acid, Biotin, and Cholin; National Academic Press: Washington, DC, USA, 1998. [Google Scholar]
- Lemos, B.S.; Medina-Vera, I.; Malysheva, O.V.; Caudill, M.A.; Fernandez, M.L. Effects of Egg Consumption and Choline Supplementation on Plasma Choline and Trimethylamine-N-Oxide in a Young Population. J. Am. Coll. Nutr. 2018, 37, 716–723. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Wang, W.; Gregory, J.F.; Malysheva, O.; Brenna, J.T.; Stabler, S.P.; Allen, R.H.; A Caudill, M. MTHFR C677T genotype influences the isotopic enrichment of one-carbon metabolites in folate-compromised men consuming d9-choline. Am. J. Clin. Nutr. 2011, 93, 348–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and β-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef] [Green Version]
- Blesso, C.N.; Andersen, C.; Barona, J.; Volk, B.; Volek, J.S.; Fernandez, M.L. Effects of carbohydrate restriction and dietary cholesterol provided by eggs on clinical risk factors in metabolic syndrome. J. Clin. Lipidol. 2013, 7, 463–471. [Google Scholar] [CrossRef]
- Gao, X.; Wang, Y.; Sun, G. High dietary choline and betaine intake is associated with low insulin resistance in the Newfoundland population. Nutrition 2017, 33, 28–34. [Google Scholar] [CrossRef]
- Nakamura, T.; Fujiwara, R.; Ishiguro, N.; Oyabu, M.; Nakanishi, T.; Shirasaka, Y.; Maeda, T.; Tamai, I. Involvement of choline transporter-like proteins, CTL1 and CTL2, in glucocorticoid-induced acceleration of phosphatidylcholine synthesis via increased choline uptake. Biol. Pharm. Bull. 2010, 33, 691–696. [Google Scholar] [CrossRef] [Green Version]
- Wang, M. The role of glucocorticoid action in the pathophysiology of the Metabolic Syndrome. Nutr. Metab. 2005, 2, 3. [Google Scholar] [CrossRef] [Green Version]
- Pinelli, N.R.; Jaber, L.A.; Brown, M.B.; Herman, W.H. Serum 25-Hydroxy Vitamin D and Insulin Resistance, Metabolic Syndrome, and Glucose Intolerance Among Arab Americans. Diabetes Care 2010, 33, 1373–1375. [Google Scholar] [CrossRef] [Green Version]
- Farrokhian, A.; Bahmani, F.; Taghizadeh, M.; Mirhashemi, S.M.; Aarabi, M.H.; Raygan, F.; Aghadavod, E.; Asemi, Z. Selenium Supplementation Affects Insulin Resistance and Serum hs-CRP in Patients with Type 2 Diabetes and Coronary Heart Disease. Horm. Metab. Res. 2016, 48, 263–268. [Google Scholar] [CrossRef] [Green Version]
- Mäntyselkä, P.; Kautiainen, H.; Saltevo, J.; Würtz, P.; Soininen, P.; Kangas, A.J.; Ala-Korpela, M.; Vanhala, M. Weight change and lipoprotein particle concentration and particle size: A cohort study with 6.5-year follow-up. Atherosclerosis 2012, 223, 239–243. [Google Scholar] [CrossRef] [PubMed]
- Rosenkilde, M.; Rygaard, L.; Nordby, P.; Nielsen, L.B.; Stallknecht, B.M. Exercise and weight loss effects on cardiovascular risk factors in overweight men. J. Appl. Physiol. 2018, 125, 901–908. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Hobbs, D.A.; Cockcroft, J.R.; Elwood, P.C.; Pickering, J.E.; Lovegrove, J.A.; Givens, D.I. Association between egg consumption and cardiovascular disease events, diabetes and all-cause mortality. Eur. J. Nutr. 2017, 57, 2943–2952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pourafshar, S.; Akhavan, N.S.; George, K.S.; Foley, E.M.; Johnson, S.A.; Keshavarz, B.; Navaei, N.; Davoudi, A.; Clark, E.A.; Arjmandi, B.H. Egg consumption may improve factors associated with glycemic control and insulin sensitivity in adults with pre- and type II diabetes. Food Funct. 2018, 9, 4469–4479. [Google Scholar] [CrossRef] [PubMed]
- Djoussé, L.; Kamineni, A.; Nelson, T.L.; Carnethon, M.R.; Mozaffarian, D.; Siscovick, D.; Mukamal, K.J. Egg consumption and risk of type 2 diabetes in older adults. Am. J. Clin. Nutr. 2010, 92, 422–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, P. Inflammation and the Metabolic Syndrome. Indian J. Clin. Biochem. 2011, 26, 317–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devaraj, S.; Singh, U.; Jialal, I. Human C-reactive protein and the metabolic syndrome. Curr. Opin. Lipidol. 2009, 20, 182–189. [Google Scholar] [CrossRef] [Green Version]
- Ratliff, J.C.; Mutungi, G.; Puglisi, M.J.; Volek, J.S.; Fernandez, M.L. Eggs modulate the inflammatory response to carbohydrate restricted diets in overweight men. Nutr. Metab. 2008, 5, 6. [Google Scholar] [CrossRef] [Green Version]
- Mutungi, G.; Ratliff, J.; Puglisi, M.; Torres-Gonzalez, M.; Vaishnav, U.; Leite, J.O.; Quann, E.; Volek, J.S.; Fernandez, M.L. Dietary Cholesterol from Eggs Increases Plasma HDL Cholesterol in Overweight Men Consuming a Carbohydrate-Restricted Diet. J. Nutr. 2008, 138, 272–276. [Google Scholar] [CrossRef] [Green Version]
- Greene, C.M.; Zern, T.L.; Wood, R.J.; Shrestha, S.; Aggarwal, D.; Sharman, M.J.; Volek, J.S.; Fernandez, M.L. Maintenance of the LDL Cholesterol:HDL Cholesterol Ratio in an Elderly Population Given a Dietary Cholesterol Challenge. J. Nutr. 2005, 135, 2793–2798. [Google Scholar] [CrossRef] [Green Version]
- Ballesteros, M.N.; Cabrera, R.M.; Saucedo, M.D.S.; Fernandez, M.L. Dietary cholesterol does not increase biomarkers for chronic disease in a pediatric population from northern Mexico. Am. J. Clin. Nutr. 2004, 80, 855–861. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Parameter | Egg | CB Supplement |
|---|---|---|
| Energy (Kcal/day) 2 | 1860 ± 152 | 1675 ± 141 |
| Fat (% Energy) | 42.4 ± 7.6 a | 36.4 ± 6.1 b |
| Carbohydrate (% Energy) | 34.9 ± 10.0 a | 44.1 ± 9.2 b |
| Protein (% Energy) | 20.3 ± 5.2 a | 17.6 ± 6.0 b |
| Alcohol (% Energy) | 2.3 ± 4.2 | 1.9 ± 3.2 |
| SFA (g/d) | 30.8 ± 14.3 | 26.0 ± 13.6 |
| MUFA (g/d) | 31.2 ± 11.2 a | 24.3 ± 10.9 b |
| PUFA (g/d) | 17.2 ± 8.8 | 15.2 ± 7.5 |
| TFA (g/d) | 2.1 ± 1.1 | 2.0 ± 1.0 |
| Omega (g/d) | 2.0 ± 1.0 | 1.9 ± 1.1 |
| Added sugar (g/d) | 31.9 ± 32.8 | 41.1 ± 32.0 |
| Glycemic index | 59.0 ± 6.9 | 57.8 ± 5.4 |
| Glycemic load | 94.8 ± 55.8 | 100.7 ± 49.5 |
| Fiber (g/d) | 16.0 ± 6.8 | 17.6 ± 5.2 |
| Soluble Fiber (g/d) | 5.8 ± 2.4 | 6.5 ± 2.2 |
| Insoluble Fiber (g/d) | 10.1 ± 4.8 | 11.0 ± 3.8 |
| Cholesterol (mg/d) | 752.7 ± 123.6 | 201.8 ± 90.2 |
| Vitamin A | 1077 ± 457 | 1022 ± 551 |
| Vitamin E | 9.04 ± 3.67 | 7.60 ± 4.42 |
| Vitamin D | 7.1 ± 4.1 a | 3.8 ± 3.4 b |
| Selenium | 138.4 ± 54.8 a | 93.1 ± 41.1 b |
| α-carotene (mg/d) | 353 ± 448 | 480 ± 486 |
| β-carotene (mg/d) | 2782 ± 2092 | 3701 ± 3065 |
| Cryptoxanthin | 247 ± 596 | 143 ± 252 |
| Lutein + Zeaxanthin (mg/d) | 2226 ± 1835 | 2600 ± 4222 |
| Lycopene (mg/d) | 6000 ± 9099 | 6026 ± 4656 |
| Physical activity (min) | 68.3 ± 47.1 | 71.3 ± 50.8 |
| Parameter | Values |
|---|---|
| Age (years) | 55.2 ± 8.9 |
| Gender Females (%) | 65 |
| Waist Circumference (cm) | 113.3 ± 12.4 |
| Systolic Blood Pressure (mm Hg) | 129.3 ± 7.3 |
| Diastolic Blood Pressure (mm Hg) | 87.5 ± 4.8 |
| HDL cholesterol (mg/dL) | 47.7 ± 17.4 |
| Triglycerides (mg/dL) | 129.5 ± 57.6 |
| Glucose (mg/dL) | 102.6 ± 10.5 |
| Parameter | BL-Egg | p-Eggs | BL-CB | p-CB |
|---|---|---|---|---|
| Body weight (Kg) 2 | 94.1 ± 19.0 | 94.0 ± 19.2 | 94.1 ± 18.6 | 93.9 ± 18.9 |
| BMI (kg/m2) | 32.4 ± 4.7 | 32.3 ± 4.8 | 32.4 ± 4.6 | 32.3 ± 4.7 |
| Waist circumference (cm) | 112.3 ± 12.0 | 112.9 ± 11.9 | 112.7 ± 12.1 | 112.1 ± 12.3 |
| Systolic blood pressure (mm Hg) | 127.6 ± 8.2 | 127.8 ± 11.7 | 128.2 ± 7.6 | 129.5 ± 7.7 |
| Diastolic blood pressure (mm Hg) | 85.6 ± 16.3 | 85.4 ± 7.5 | 81.8 ± 16.8 | 85.9 ± 7.0 |
| CRP (mg/dL) | 0.48 ± 0.57 a | 0.32 ± 0.31 b | 0.36 ± 0.31 ab | 0.36 ± 0.34 ab |
| ALT (U/L) | 27.3 ± 13.2 | 27.3 ± 13.3 | 26.8 ± 12.9 | 24.8 ± 10.2 |
| AST (U/L) | 22.9 ± 6.1 | 23.9 ± 9.1 | 23.4 ± 7.4 | 23.9 ± 6.3 |
| Parameter | BL-Egg | p-Egg | BL-CB | p-CB |
|---|---|---|---|---|
| Total cholesterol (mg/dL) | 177.7 ± 27.3 | 185.9 ± 25.5 | 177.9 ± 19.1 | 185.3 ± 24.8 |
| LDL Cholesterol (mg/dL) | 105.4 ± 23.2 | 111.4 ± 20.5 | 103.2 ± 16.5 | 110.8 ± 21.4 |
| HDL Cholesterol (mg/dL) | 48.5 ± 16.9 | 50.2 ± 16.7 | 48.4 ± 14.3 | 48.5 ± 14.8 |
| Triglycerides (mg/dL) | 122.0 ± 46.9 | 122.4 ± 46.9 | 137.3 ± 56.6 | 129.3 ± 42.2 |
| Plasma Glucose (mg/dL) | 100.6 ± 10.8 | 101.9 ± 10.9 | 99.8 ± 12.5 | 101.8 ± 11.3 |
| LDL/HDL ratio | 2.43 ± 0.92 | 2.47 ± 0.97 | 2.33 ± 0.78 | 2.50 ± 0.99 |
| Parameter | Baseline | p-Egg | p-CB |
|---|---|---|---|
| Il-6 (pg/mL) 2 | 5.6 ± 0.9 a | 4.6 ± 1.7 b | 4.4 ± 1.3 b |
| TNF-α (pg/mL) | 5.1 ± 1.1 | 5.9 ± 1.6 | 5.9 ± 1.5 |
| MCP-1 (pg/mL) | 107.4 ± 46.1 | 110.9 ± 31.6 | 116.0 ± 31.8 |
| Insulin (pg/mL) | 152.4 ± 87.2 a | 97.7 ± 66.5 b | 111.3 ± 100 a,b |
| Insulin Resistance (HOMA) | 5.43 ± 0.38 a | 3.16 ± 0.26 b | 4.01 ± 0.40 a,b |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
DiBella, M.; Thomas, M.S.; Alyousef, H.; Millar, C.; Blesso, C.; Malysheva, O.; Caudill, M.A.; Fernandez, M.L. Choline Intake as Supplement or as a Component of Eggs Increases Plasma Choline and Reduces Interleukin-6 without Modifying Plasma Cholesterol in Participants with Metabolic Syndrome. Nutrients 2020, 12, 3120. https://doi.org/10.3390/nu12103120
DiBella M, Thomas MS, Alyousef H, Millar C, Blesso C, Malysheva O, Caudill MA, Fernandez ML. Choline Intake as Supplement or as a Component of Eggs Increases Plasma Choline and Reduces Interleukin-6 without Modifying Plasma Cholesterol in Participants with Metabolic Syndrome. Nutrients. 2020; 12(10):3120. https://doi.org/10.3390/nu12103120
Chicago/Turabian StyleDiBella, Marissa, Minu S. Thomas, Hana Alyousef, Courtney Millar, Christopher Blesso, Olga Malysheva, Marie A. Caudill, and Maria Luz Fernandez. 2020. "Choline Intake as Supplement or as a Component of Eggs Increases Plasma Choline and Reduces Interleukin-6 without Modifying Plasma Cholesterol in Participants with Metabolic Syndrome" Nutrients 12, no. 10: 3120. https://doi.org/10.3390/nu12103120
APA StyleDiBella, M., Thomas, M. S., Alyousef, H., Millar, C., Blesso, C., Malysheva, O., Caudill, M. A., & Fernandez, M. L. (2020). Choline Intake as Supplement or as a Component of Eggs Increases Plasma Choline and Reduces Interleukin-6 without Modifying Plasma Cholesterol in Participants with Metabolic Syndrome. Nutrients, 12(10), 3120. https://doi.org/10.3390/nu12103120