Green Tomato Extract Prevents Bone Loss in Ovariectomized Rats, a Model of Osteoporosis

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Samples
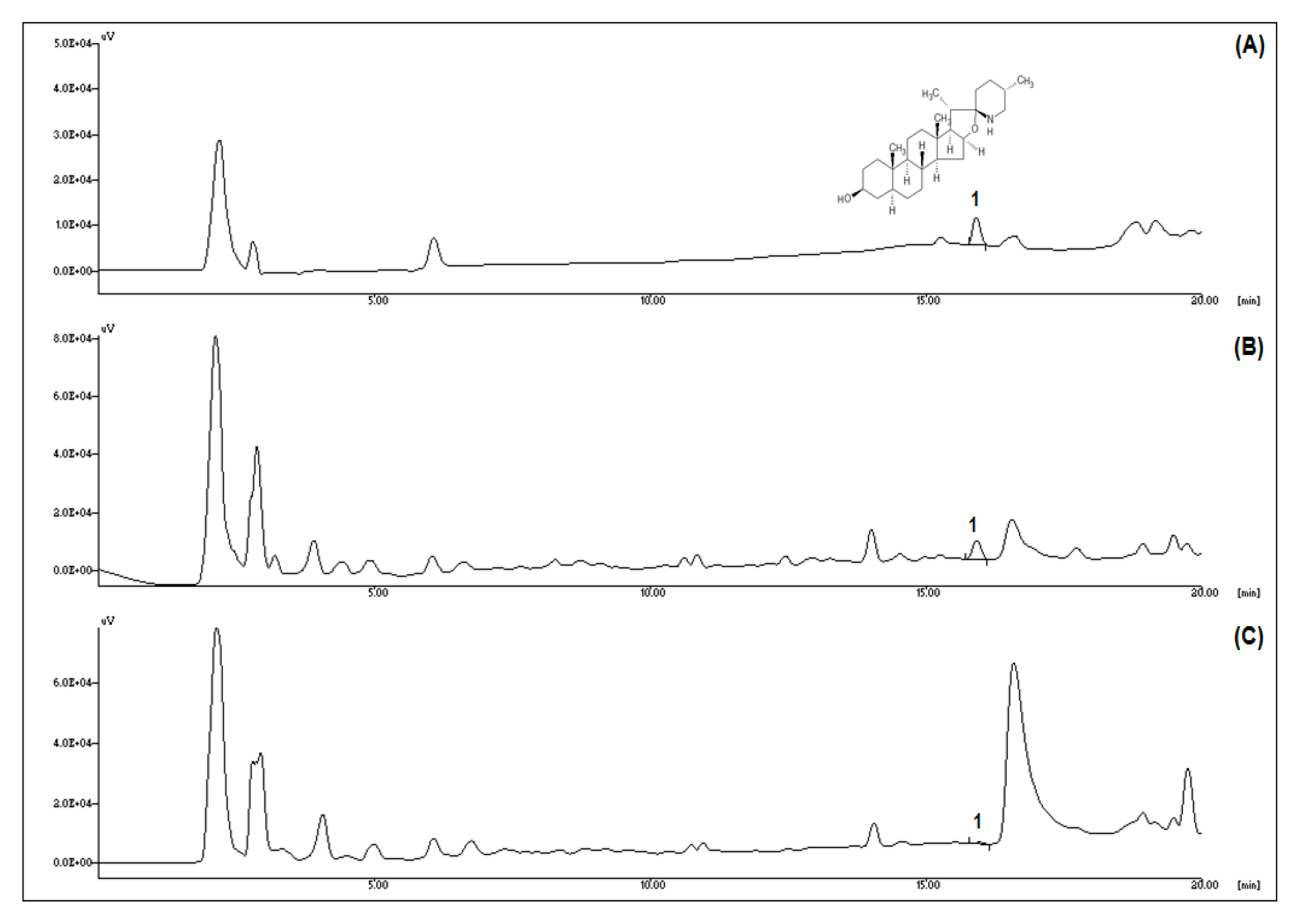
2.2. Tomatidine Analysis by High-Performance Liquid Chromatography (HPLC)
2.3. Animal Works
2.4. Micro-Computed Tomography (Micro-CT) Analysis
2.5. Serum and Urine Biochemical Assays
2.6. Quantitative Real-Time PCR (qRT-PCR) Analysis
2.7. Statistical Analysis
3. Results
3.1. Green Tomatoes Contain a Higher Content of Tomatidine than Ripened Red Tomatoes
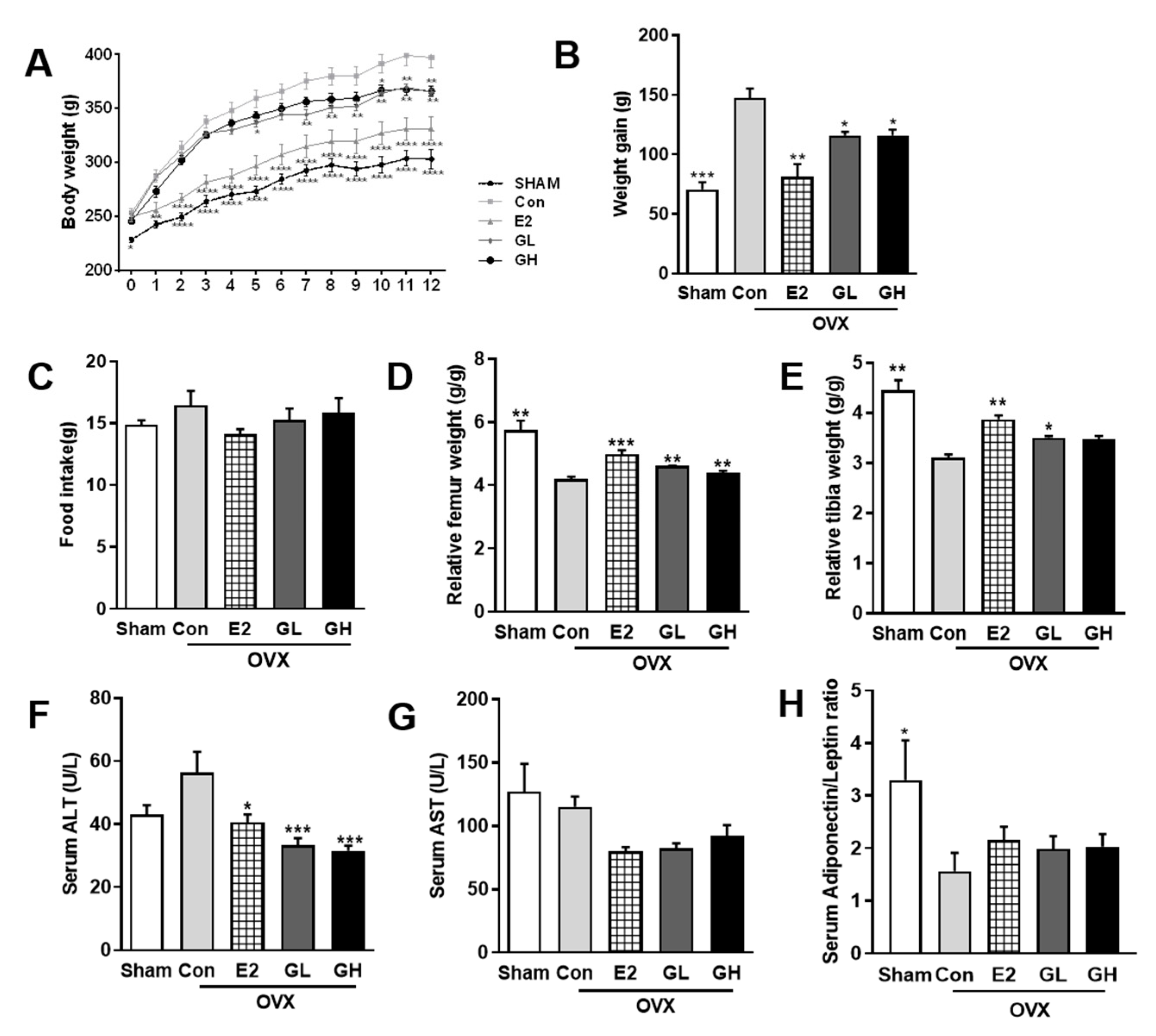
3.2. GTE Prevents Weight Gain and Increases Bone Mass in OVX Rats
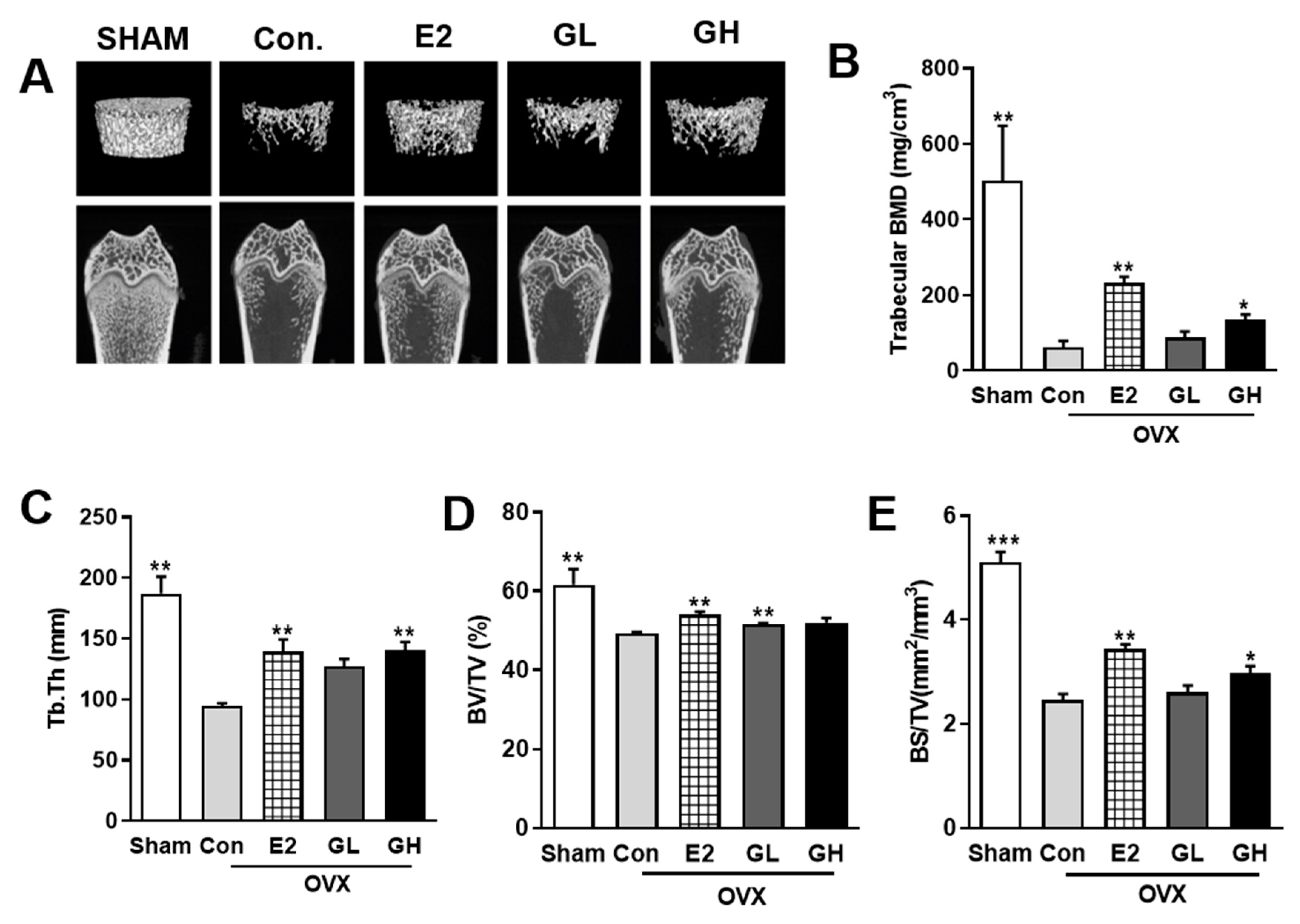
3.3. GTE Increases BMD in OVX Rats
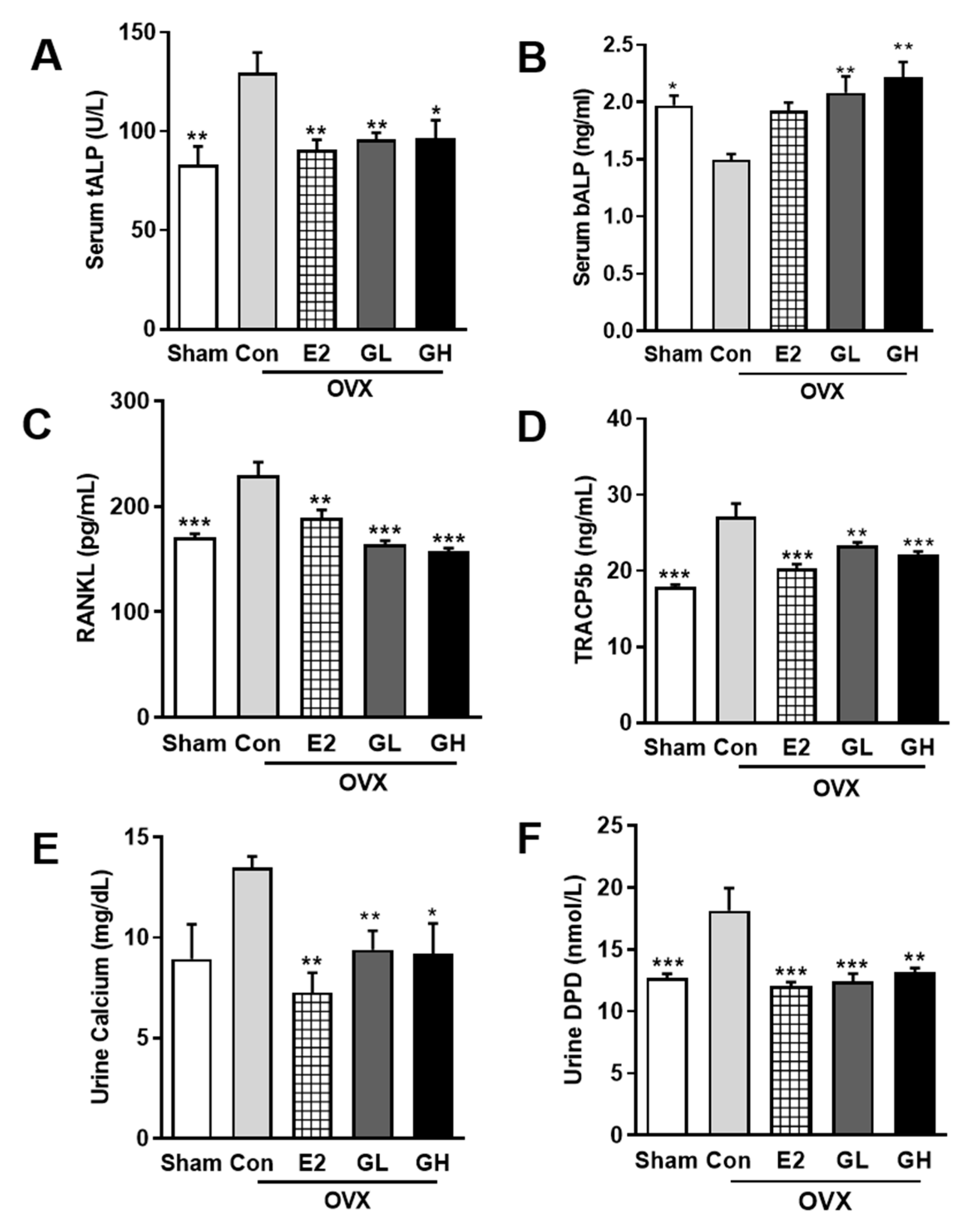
3.4. GTE Improves Bone Formation in OVX Rats
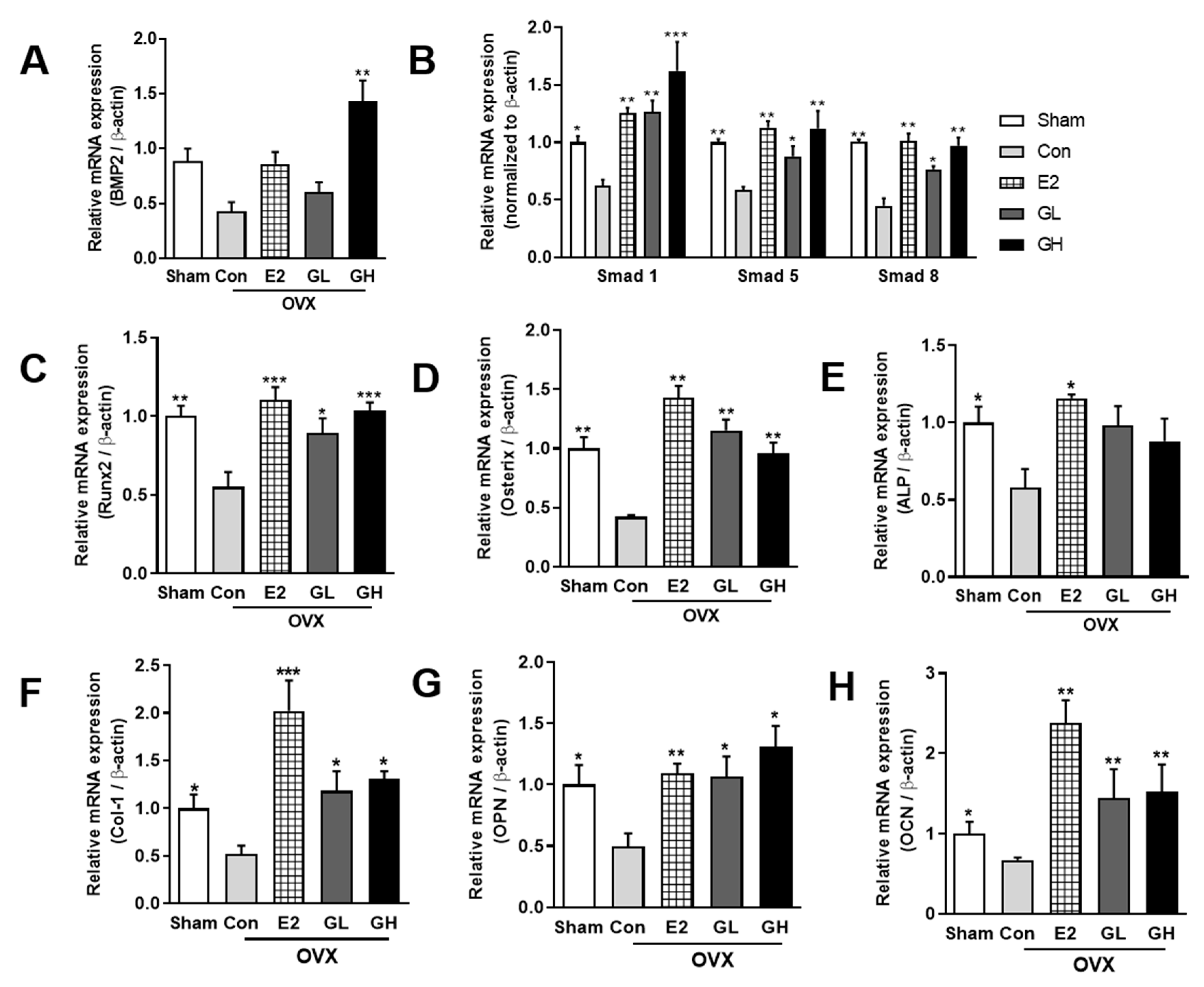
3.5. GTE Increases Bone Formation by Regulating Bmp2-Smads 1/5/8-Runx2 Signaling
3.6. GTE Decreases Bone Resorption through the RANKL/OPG Pathway
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Zaidi, M.; Yuen, T.; Sun, L.; Rosen, C.J. Regulation of Skeletal Homeostasis. Endocr. Rev. 2018, 39, 701–718. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Zhou, C.; Li, J.; Liu, R.; Shi, B.; Yuan, Q.; Zou, S. Autophagy in bone homeostasis and the onset of osteoporosis. Bone Res. 2019, 7, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watts, N.B. Postmenopausal Osteoporosis: A Clinical Review. J. Women’s Health. 2018, 27, 1093–1096. [Google Scholar] [CrossRef] [PubMed]
- Pouresmaeili, F.; Kamalidehghan, B.; Kamarehei, M.; Goh, Y.M. A comprehensive overview on osteoporosis and its risk factors. Ther. Clin. Risk Manag. 2018, 14, 2029–2049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sözen, T.; Özışık, L.; Başaran, N.Ç. An overview and management of osteoporosis. Eur. J. Rheumatol. 2017, 4, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Zizola, C.; Kennel, P.J.; Akashi, H.; Ji, R.; Castillero, E.; George, I.; Homma, S.; Schulze, P.C. Activation of PPARδ signaling improves skeletal muscle oxidative metabolism and endurance function in an animal model of ischemic left ventricular dysfunction. J. Physiol. Heart Circ. Physiol. 2015, 308, H1078–H1085. [Google Scholar] [CrossRef] [PubMed]
- Eastell, R.; O’Neill, T.W.; Hofbauer, L.C.; Langdahl, B.; Reid, I.R.; Gold, D.T.; Cummings, S.R. Postmenopausal osteoporosis. Nat. Rev. Dis. Primers 2016, 2, 16069. [Google Scholar] [CrossRef]
- Kling, J.M.; Clarke, B.L.; Sandhu, N.P. Osteoporosis prevention, screening, and treatment: A review. J. Womens Health 2014, 23, 563–572. [Google Scholar] [CrossRef]
- Chen, J.H.; Chen, Y.C.; Tsai, M.K.; Chiou, J.M.; Lee, W.C.; Tsao, C.K.; Tsai, K.S.; Chie, W.C. Predicting the risk of osteopenia for women aged 40–55 years. JFMA 2017, 116, 888–896. [Google Scholar] [CrossRef]
- Aggarwal, S.; Gahlot, N.; Saini, U.; Dhillon, M. Bisphosphonates in Orthopedics: Evidence-based Review of Indications and Adverse Effects. JPMER 2016, 50, 75–85. [Google Scholar] [CrossRef]
- Bodenner, D.; Redman, C.; Riggs, A. Teriparatide in the management of osteoporosis. Clin. Interv. Aging 2007, 2, 499–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gambacciani, M.; Levancini, M. Hormone replacement therapy and the prevention of postmenopausal osteoporosis. Prz. Menopauzalny 2014, 13, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Wallace, T.C. Dried Plums, Prunes and Bone Health: A Comprehensive Review. Nutrients 2017, 9, 401. [Google Scholar] [CrossRef] [PubMed]
- Matheson, E.M.; Mainous, A.G., 3rd; Carnemolla, M.A. The association between onion consumption and bone density in perimenopausal and postmenopausal non-Hispanic white women 50 years and older. Menopause 2009, 16, 756–759. [Google Scholar] [CrossRef] [PubMed]
- Hatefi, M.; Ahmadi, M.R.H.; Rahmani, A.; Dastjerdi, M.M.; Asadollahi, K. Effects of Curcumin on Bone Loss and Biochemical Markers of Bone Turnover in Patients with Spinal Cord Injury. World Neurosurg. 2018, 114, e785–e791. [Google Scholar] [CrossRef]
- Jeon, E.J.; Lee, D.H.; Kim, Y.J.; Ahn, J.; Kim, M.J.; Hwang, J.-T.; Hur, J.; Kim, M.; Jang, Y.-J.; Ha, T.-Y.; et al. Effects of yuja peel extract and its flavanones on osteopenia in ovariectomized rats and osteoblast differentiation. MNFR 2016, 60, 2587–2601. [Google Scholar] [CrossRef]
- Kim, J.S.; Lee, H.; Nirmala, F.S.; Jung, C.H.; Kim, M.J.; Jang, Y.J.; Ha, T.Y.; Ahn, J. Dihydrodaidzein and 6-hydroxydaidzein mediate the fermentation-induced increase of antiosteoporotic effect of soybeans in ovariectomized mice. FASEB J. 2018, 33, 3252–3263. [Google Scholar] [CrossRef]
- Wu, X.; Xie, C.Q.; Zhu, Q.Q.; Wang, M.Y.; Sun, B.; Huang, Y.P.; Shen, C.; An, M.F.; Zhao, Y.L.; Wang, X.J.; et al. Green tea (Camellia sinensis) aqueous extract alleviates postmenopausal osteoporosis in ovariectomized rats and prevents RANKL-induced osteoclastogenesis in vitro. Food Nutr. Res. 2018, 62. [Google Scholar] [CrossRef] [Green Version]
- Reimers, K.J.; Keast, D.R. Tomato Consumption in the United States and Its Relationship to the US Department of Agriculture Food Pattern: Results from what we Eat in America 2005–2010. Food Health 2016, 51, 198–205. [Google Scholar] [CrossRef]
- Friedman, M.; Levin, C.E.; Lee, S.-U.; Kim, H.-J.; Lee, I.-S.; Byun, J.-O.; Kozukue, N. Tomatine-Containing Green Tomato Extracts Inhibit Growth of Human Breast, Colon, Liver, and Stomach Cancer Cells. JAFC 2009, 57, 5727–5733. [Google Scholar] [CrossRef]
- Hu, B.; Sun, X.; Yang, Y.; Ying, Z.; Meng, J.; Zhou, C.; Jiang, G.; Li, S.; Wu, F.; Zhao, X.; et al. Tomatidine suppresses osteoclastogenesis and mitigates estrogen deficiency-induced bone mass loss by modulating TRAF6-mediated signaling. FASEB J. 2018, 33, 2574–2586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, E.F.; Waltz, T.B.; Kassahun, H.; Lu, Q.; Kerr, J.S.; Morevati, M.; Fivenson, E.M.; Wollman, B.N.; Marosi, K.; Wilson, M.A.; et al. Tomatidine enhances lifespan and healthspan in C. elegans through mitophagy induction via the SKN-1/Nrf2 pathway. Sci. Rep. 2017, 7, 46208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheng, B.; Chu, Y.P.; Wong, W.T.; Yau, E.K.C.; Chen, S.P.L.; Luk, W.H. Improvement of bone mineral density after enzyme replacement therapy in Chinese late-onset Pompe disease patients. BMC Res. Notes 2017, 10, 351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahn, J.; Lee, H.; Jung, C.H.; Jeon, T.I.; Ha, T.Y. MicroRNA-146b promotes adipogenesis by suppressing the SIRT1-FOXO1 cascade. EMBO Mol. Med. 2013, 5, 1602–1612. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, Y.; Kiyota, N.; Tsurushima, K.; Yoshitomi, M.; Horlad, H.; Ikeda, T.; Nohara, T.; Takeya, M.; Nagai, R. Tomatidine, a Tomato Sapogenol, Ameliorates Hyperlipidemia and Atherosclerosis in ApoE-Deficient Mice by Inhibiting Acyl-CoA:cholesterol Acyl-transferase (ACAT). JAFC 2012, 60, 2472–2479. [Google Scholar] [CrossRef]
- Nakahama, K.I. Cellular communications in bone homeostasis and repair. Cell. Mol. Life Sci. 2010, 67, 4001–4009. [Google Scholar] [CrossRef]
- Sun, J.; Li, J.; Li, C.; Yu, Y. Role of bone morphogenetic protein-2 in osteogenic differentiation of mesenchymal stem cells. Mol. Med. Rep. 2015, 12, 4230–4237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, H.; Tang, Y.; Lozito, T.P.; Oyster, N.; Wang, B.; Tuan, R.S. Efficient in vivo bone formation by BMP-2 engineered human mesenchymal stem cells encapsulated in a projection stereolithographically fabricated hydrogel scaffold. Stem Cell Res. Ther. 2019, 10, 254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, D.B.; Sui, C.; Wu, T.T.; Wu, L.Z.; Zhu, Y.Y.; Ren, Z.H. Association of Bone Morphogenetic Protein (BMP)/Smad Signaling Pathway with Fracture Healing and Osteogenic Ability in Senile Osteoporotic Fracture in Humans and Rats. Med. Sci. Monit. 2018, 24, 4363–4371. [Google Scholar] [CrossRef] [PubMed]
- Shu, B.; Zhang, M.; Xie, R.; Wang, M.; Jin, H.; Hou, W.; Tang, D.; Harris, S.E.; Mishina, Y.; O’Keefe, R.J.; et al. BMP2, but not BMP4, is crucial for chondrocyte proliferation and maturation during endochondral bone development. J. Cell Sci. 2011, 124, 3428–3440. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Oyajobi, B.O.; Harris, S.E.; Chen, D.; Tsao, C.; Deng, H.-W.; Zhao, M. Wnt/β-catenin signaling activates bone morphogenetic protein 2 expression in osteoblasts. Bone 2013, 52, 145–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukherjee, A.; Rotwein, P. Akt promotes BMP2-mediated osteoblast differentiation and bone development. J. Cell Sci. 2009, 122, 716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshiya, T.; Shingo, N.; Yosuke, O. Osteoblasts and Osteoclasts in Bone Remodeling and Inflammation. Curr. Drug Targets Inflamm. Allergy 2005, 4, 325–328. [Google Scholar] [CrossRef]
- Go, E.M.; Oh, J.H.; Park, J.H.; Lee, S.Y.; Lee, N.K. Spi-C positively regulates RANKL-mediated osteoclast differentiation and function. Exp. Mol. Med. 2020, 52, 691–701. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene. | Sequence (5′→3′) |
|---|---|
| BMP2 | F: TGA GGA TTA GCA GGT CTT TG |
| R: CAC AAC CAT GTC CTG ATA AT | |
| Smad1 | F: TCAATAGAGGAGATGTTCAAGCAGT |
| R: AAACCATCCACCAACACGCT | |
| Smad5 | F: CCCTATCCTCCTTCCCCTGC |
| R: GAGGGGTATCAGCTGGGAGTT | |
| Smad8 | F: ACT TCC GGC CAG TTT GCT AC |
| R: TGG GGA TCT TGC AGA CAG TG | |
| Runx2 | F: GGC GTC AAA CAG CCT CTT CA |
| R: GCT CGG ATC CCA AAA GAA GTT | |
| Osterix | F: TGG CCA TGC TGA CTG CAG CC |
| R: TGG GTA GGC GTC CCC CAT GG | |
| ALP | F:GCACAACATCAAGGACATCG |
| R:TGGCCTTCTCATCCAGTTCA | |
| Col-1 | F: GCA TGG CCA AGA AGA CAT CC |
| R: CCT CGG GTT TCC ACG TCT C | |
| OPN | F: GAT GAA TCT GAC GAA TCT CAC C |
| R: CTC AGA AGC TGG GCA ACA GGG AT | |
| OCN | F: CAG ACA AGT CCC ACA CAG |
| Rankl | F:TGAAGACACACTACCTGACTCCTG |
| R:CCACAATGTGTTGCAGTTCC | |
| OPG | F:GTTTCCCGAAGGACCACAAT |
| R:CCATTCAATGATGTCCAGGAG | |
| NOX4 | F: CTG CAT CTG TCC TGA ACC TCA A |
| R: TCT CCT GCT AGG GAC CTT CTG T | |
| β-actin | F:TCTTCCAGCCTTCCTTCCTG |
| R:TAGAGCCACCAATCCACACA |
| Tomato Samples | Tomatidine Content (mg/100 g Dry Weight) | p-Value |
|---|---|---|
| Green | 1.06 ± 0.11 | p < 0.001 |
| Red | 0.13 ± 0.01 |
| Weight (g) | Sham | Con | E2 | GL | GH |
|---|---|---|---|---|---|
| Liver | 9.76 ± 0.34 | 10.48 ± 0.18 | 8.30 ± 0.35 ** | 8.91 ± 0.24 *** | 8.20 ± 0.28 *** |
| WAT (Retroperitoneal) | 11.03 ± 0.04 ** | 17.94 ± 0.33 | 8.12 ± 0.99 *** | 12.98 ± 0.73 ** | 12.89 ± 0.93 ** |
| Spleen | 0.58 ± 0.02 | 0.63 ± 0.02 | 0.58 ± 0.02 | 0.59 ± 0.01 | 0.61 ± 0.02 |
| Kidney | 1.74 ± 0.08 | 1.66 ± 0.04 | 1.64 ± 0.05 | 1.64 ± 0.04 | 1.63 ± 0.04 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nirmala, F.S.; Lee, H.; Kim, J.-S.; Ha, T.; Jung, C.H.; Ahn, J. Green Tomato Extract Prevents Bone Loss in Ovariectomized Rats, a Model of Osteoporosis. Nutrients 2020, 12, 3210. https://doi.org/10.3390/nu12103210
Nirmala FS, Lee H, Kim J-S, Ha T, Jung CH, Ahn J. Green Tomato Extract Prevents Bone Loss in Ovariectomized Rats, a Model of Osteoporosis. Nutrients. 2020; 12(10):3210. https://doi.org/10.3390/nu12103210
Chicago/Turabian StyleNirmala, Farida S., Hyunjung Lee, Ji-Sun Kim, Taeyoul Ha, Chang Hwa Jung, and Jiyun Ahn. 2020. "Green Tomato Extract Prevents Bone Loss in Ovariectomized Rats, a Model of Osteoporosis" Nutrients 12, no. 10: 3210. https://doi.org/10.3390/nu12103210
APA StyleNirmala, F. S., Lee, H., Kim, J. -S., Ha, T., Jung, C. H., & Ahn, J. (2020). Green Tomato Extract Prevents Bone Loss in Ovariectomized Rats, a Model of Osteoporosis. Nutrients, 12(10), 3210. https://doi.org/10.3390/nu12103210