Immunostimulatory Effects of Live Lactobacillus sakei K040706 on the CYP-Induced Immunosuppression Mouse Model

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Live Lactobacillus sakei K040706 (Live K040706)
2.2. Animals
2.3. CYP-Induced Immunosuppression in Mice
2.4. Cell Preparation from Spleen and PPs
2.5. Determination of NK Cytotoxic Activity and Granzyme B Production
2.6. Lymphocyte Proliferation Assay
2.7. Determination of Cytokine Production
2.8. RNA Extraction and Quantitative Real-Time RT-PCR (qRT-PCR)
2.9. Analysis of Microbiota
2.9.1. DNA Extraction, Emulsion-Based PCR (emPCR), and Next-Generation Sequencing
2.9.2. Selection of 16S rRNAs and Taxonomic Assignment
2.10. Measurement of the Bone Marrow (BM) Cell Proliferation and Cytokine Production on Splenocytes and PP Cells
2.11. Statistical Analysis
3. Results
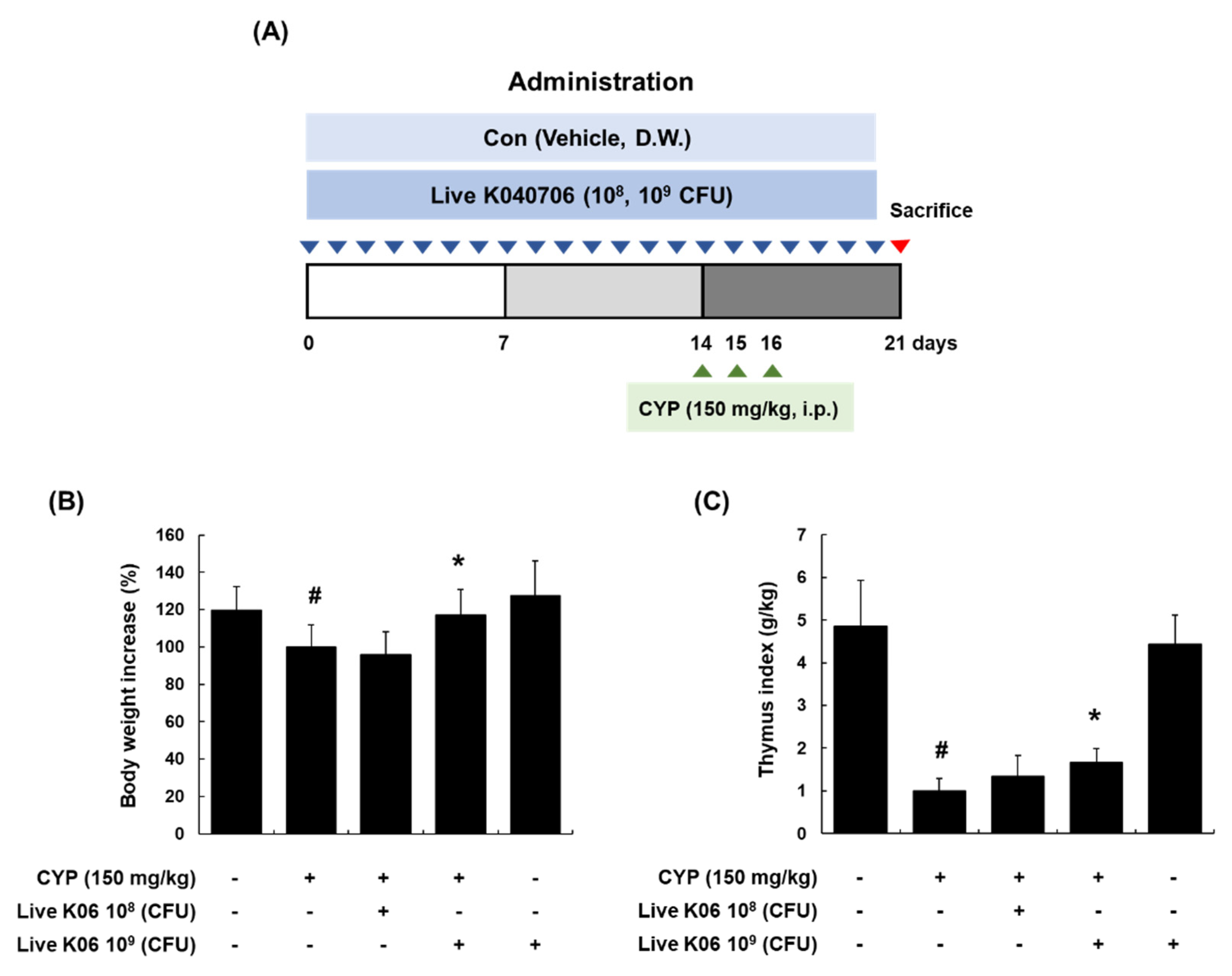
3.1. Amelioration of Changes in Body Weight and Thymus Index in CYP-Induced Mice by Live K040706
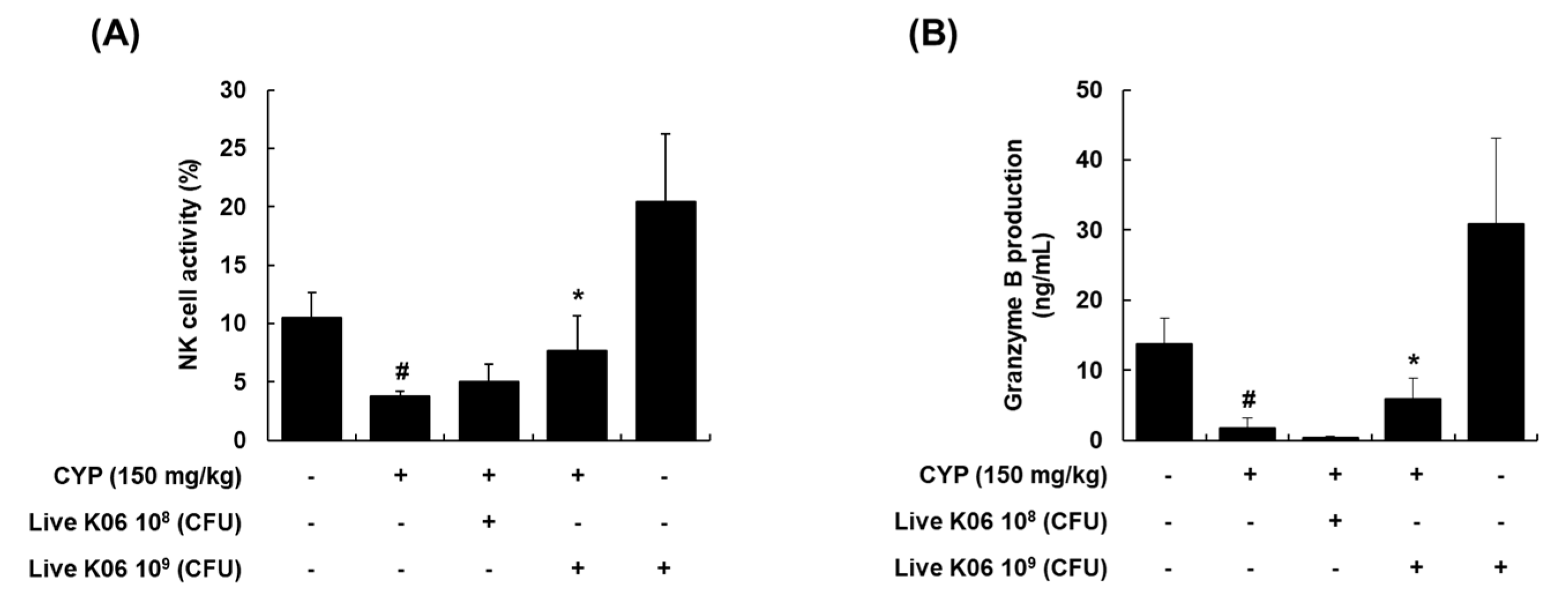
3.2. Live K040706–Mediated Restoration of NK Cell Cytotoxic Activity and Granzyme B Production in CYP-Induced Mice
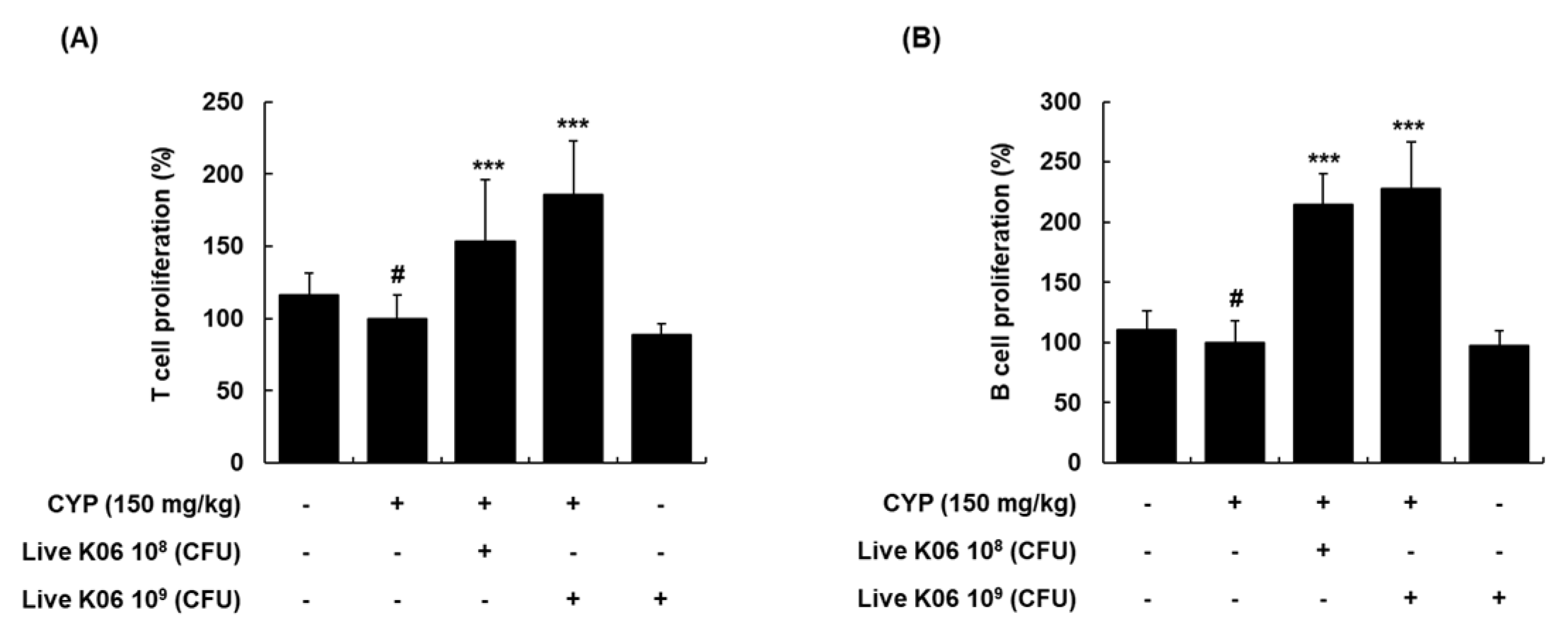
3.3. Live K040706–Mediated Increase in Splenocyte Proliferation of CYP-Induced Mice
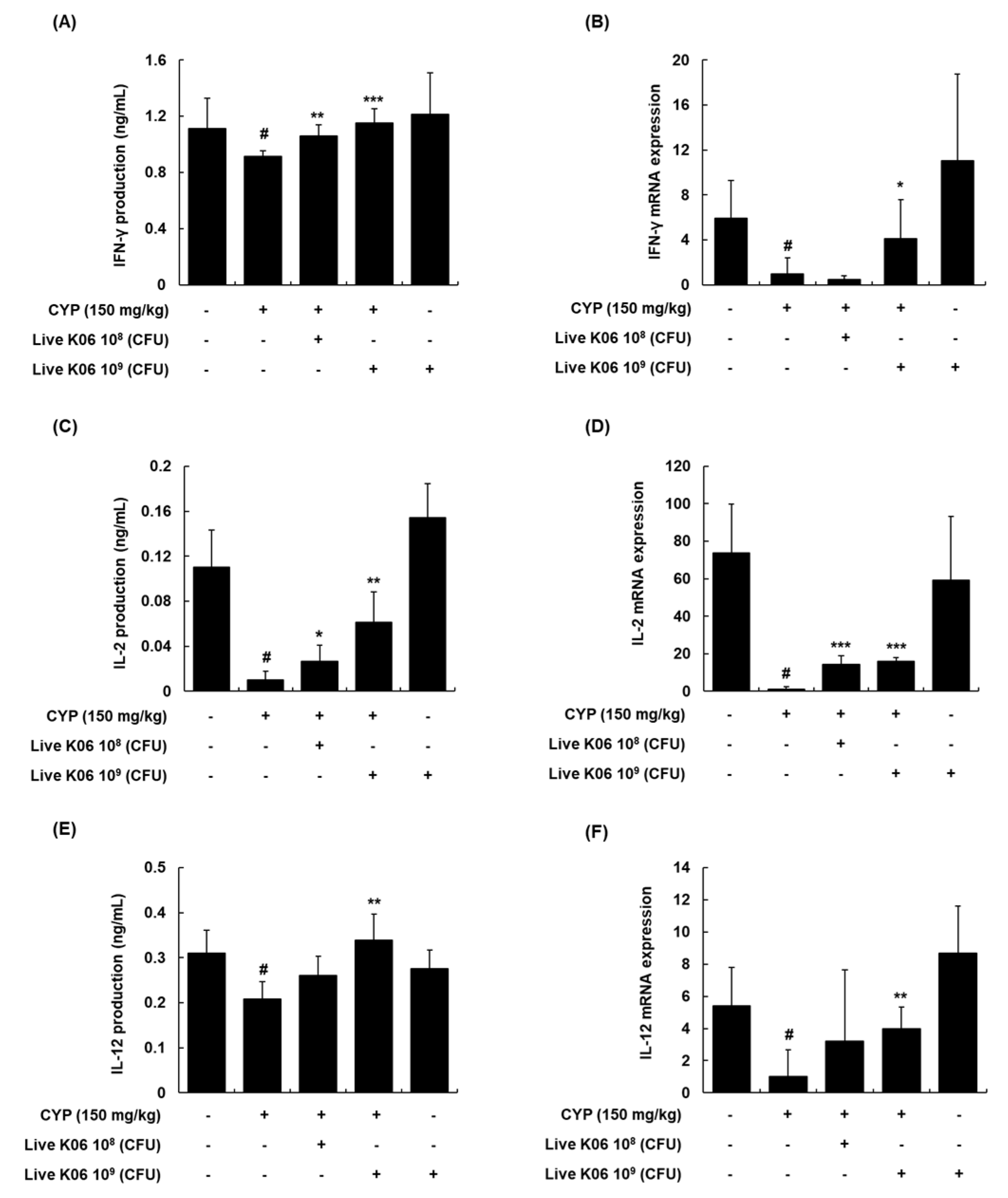
3.4. Regulation of Th1 Cytokines and Their mRNA Expression in Splenocytes of CYP-Treated Mice by Live K040706
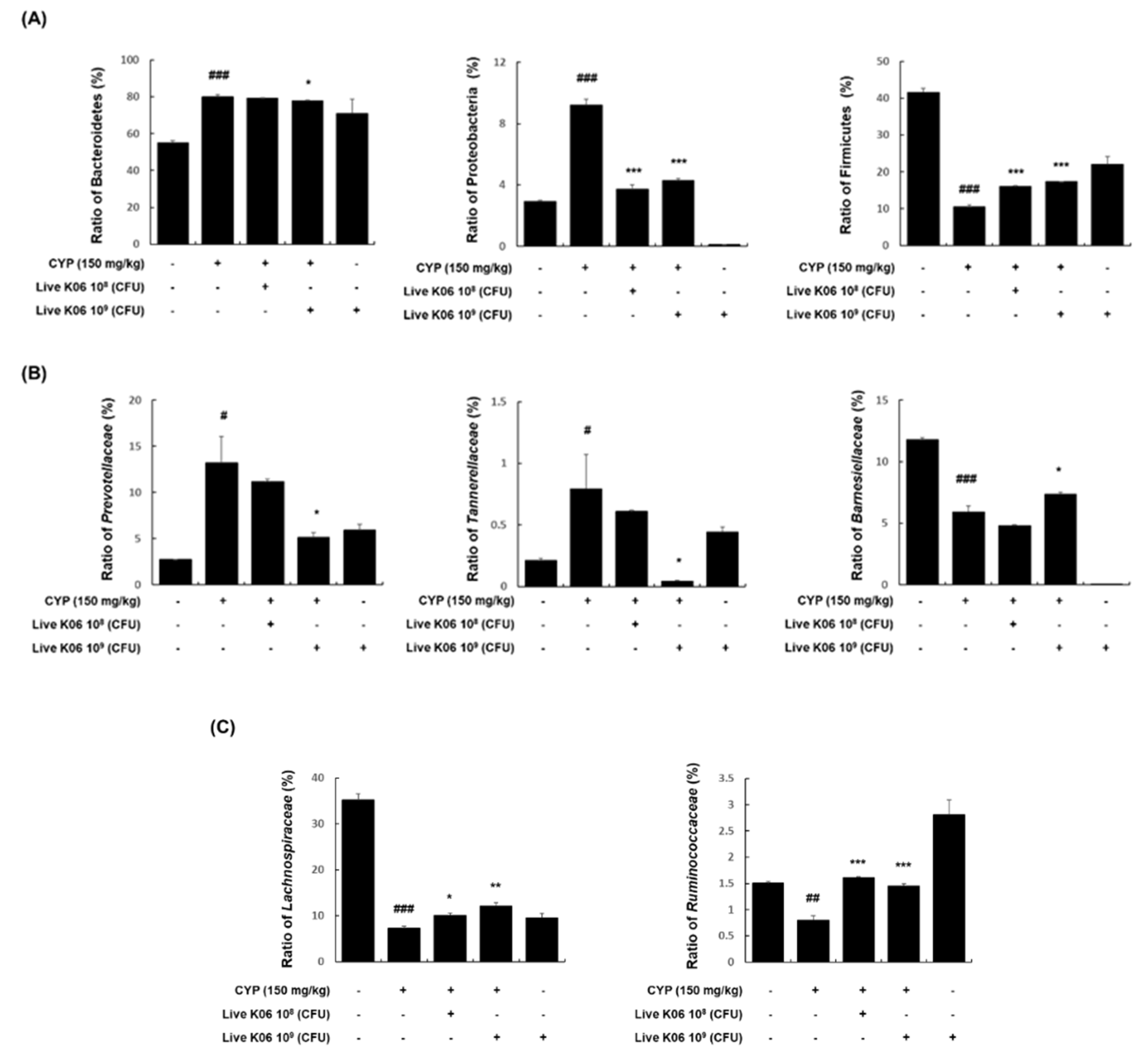
3.5. Effects of K040706 on Microbiota Composition in CYP-Treated Mice
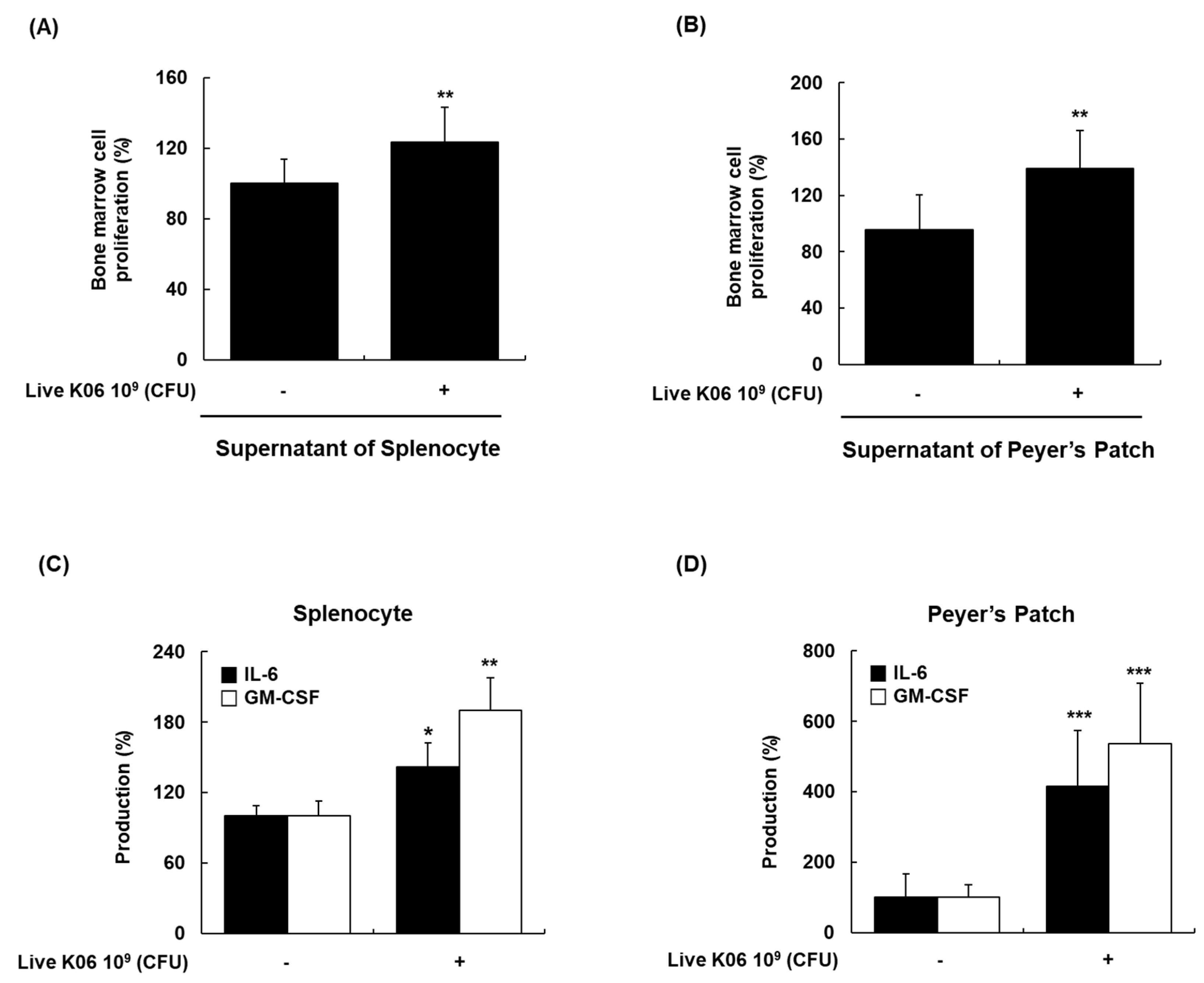
3.6. Promotion of BM Cell Hematopoiesis in Mice by Live K040706
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Hachimura, S.; Totsuka, M.; Hosono, A. Immunomodulation by food: Impact on gut immunity and immune cell function. Biosci. Biotechnol. Biochem. 2018, 82, 584–599. [Google Scholar] [CrossRef] [PubMed]
- Dhama, K.; Saminathan, M.; Jacob, S.S.; Singh, M.; Karthik, A.R.T.; Malik, L.T.S.Y.S.; Singh, R.K. Effect of Immunomodulation and Immunomodulatory Agents on Health with some Bioactive Principles, Modes of Action and Potent Biomedical Applications. Int. J. Pharmacol. 2015, 11, 253–290. [Google Scholar] [CrossRef] [Green Version]
- Kidd, P. Th1/Th2 balance: The hypothesis, its limitations, and implications for health and disease. Altern. Med. Rev. 2003, 8, 223–246. [Google Scholar] [PubMed]
- Romagnani, S. Th1/Th2 cells. Inflamm. Bowel Dis. 1999, 5, 285–294. [Google Scholar] [CrossRef] [PubMed]
- Walker, J.A.; McKenzie, A.N.J. TH2 cell development and function. Nat. Rev. Immunol. 2018, 18, 121–133. [Google Scholar] [CrossRef] [PubMed]
- De Vrese, M.; Schrezenmeir, J. Probiotics, prebiotics, and synbiotics. Adv. Biochem. Eng. Biotechnol. 2008, 111, 1–66. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.J.; Im, S.H. Probiotics as an Immune Modulator. J. Nutr. Sci. Vitaminol. 2015, 61 (Suppl. S103–S105). [Google Scholar] [CrossRef] [Green Version]
- Behnsen, J.; Deriu, E.; Sassone-Corsi, M.; Raffatellu, M. Probiotics: Properties, examples, and specific applications. Cold Spring Harb. Perspect Med. 2013, 3, a010074. [Google Scholar] [CrossRef] [Green Version]
- Lim, S.M.; Jeong, J.J.; Woo, K.H.; Han, M.J.; Kim, D.H. Lactobacillus sakei OK67 ameliorates high-fat diet-induced blood glucose intolerance and obesity in mice by inhibiting gut microbiota lipopolysaccharide production and inducing colon tight junction protein expression. Nutr. Res. 2016, 36, 337–348. [Google Scholar] [CrossRef]
- Jung, J.Y.; Shin, J.S.; Lee, S.G.; Rhee, Y.K.; Cho, C.W.; Hong, H.D.; Lee, K.T. Lactobacillus sakei K040706 evokes immunostimulatory effects on macrophages through TLR 2-mediated activation. Int. Immunopharmacol. 2015, 28, 88–96. [Google Scholar] [CrossRef]
- Seo, S.H.; Shin, J.S.; Lee, W.S.; Rhee, Y.K.; Cho, C.W.; Hong, H.D.; Lee, K.T. Anti-colitis effect of Lactobacillus sakei K040706 via suppression of inflammatory responses in the dextran sulfate sodium-induced colitis mice model. J. Funct. Foods 2017, 29, 256–268. [Google Scholar] [CrossRef]
- Shin, J.S.; Chung, S.H.; Lee, W.S.; Lee, J.Y.; Kim, J.L.; Lee, K.T. Immunostimulatory effects of cordycepin-enriched WIB-801CE from Cordyceps militaris in splenocytes and cyclophosphamide-induced immunosuppressed mice. Phytother. Res. 2018, 32, 132–139. [Google Scholar] [CrossRef] [PubMed]
- Han, H.S.; Shin, J.S.; Song, Y.R.; Rhee, Y.K.; Cho, C.W.; Ryu, J.H.; Inn, K.S.; Hong, H.D.; Lee, K.T. Immunostimulatory effects of polysaccharides isolated from young barley leaves (Hordeum vulgare L.) with dual activation of Th1 and Th2 in splenic T cells and cyclophosphamide-induced immunosuppressed mice. Int. J. Biol. Macromol. 2020, 147, 954–964. [Google Scholar] [CrossRef] [PubMed]
- Hong, T.; Matsumoto, T.; Kiyohara, H.; Yamada, H. Enhanced production of hematopoietic growth factors through T cell activation in Peyer’s patches by oral administration of Kampo (Japanese herbal) medicine, “Juzen-Taiho-To”. Phytomedicine 1998, 5, 353–360. [Google Scholar] [CrossRef]
- Uharek, L.; Zeis, M.; Glass, B.; Steinmann, J.; Dreger, P.; Gassmann, W.; Schmitz, N.; Muller-Ruchholtz, W. High lytic activity against human leukemia cells after activation of allogeneic NK cells by IL-12 and IL-2. Leukemia 1996, 10, 1758–1764. [Google Scholar]
- Mahrus, S.; Craik, C.S. Selective chemical functional probes of granzymes A and B reveal granzyme B is a major effector of natural killer cell-mediated lysis of target cells. Chem. Biol. 2005, 12, 567–577. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Sun, R. Tumor immunotherapy: New aspects of natural killer cells. Chin. J. Cancer Res. 2018, 30, 173–196. [Google Scholar] [CrossRef]
- Chen, X.; Nie, W.; Yu, G.; Li, Y.; Hu, Y.; Lu, J.; Jin, L. Antitumor and immunomodulatory activity of polysaccharides from Sargassum fusiforme. Food Chem. Toxicol. 2012, 50, 695–700. [Google Scholar] [CrossRef]
- Cerqueira, F.; Cordeiro-Da-Silva, A.; Gaspar-Marques, C.; Simoes, F.; Pinto, M.M.; Nascimento, M.S. Effect of abietane diterpenes from Plectranthus grandidentatus on T- and B-lymphocyte proliferation. Bioorg. Med. Chem. 2004, 12, 217–223. [Google Scholar] [CrossRef]
- Mosmann, T.R.; Coffman, R.L. TH1 and TH2 cells: Different patterns of lymphokine secretion lead to different functional properties. Annu. Rev. Immunol. 1989, 7, 145–173. [Google Scholar] [CrossRef]
- Scharton, T.M.; Scott, P. Natural killer cells are a source of interferon gamma that drives differentiation of CD4+ T cell subsets and induces early resistance to Leishmania major in mice. J. Exp. Med. 1993, 178, 567–577. [Google Scholar] [CrossRef] [PubMed]
- Trinchieri, G.; Matsumoto-Kobayashi, M.; Clark, S.C.; Seehra, J.; London, L.; Perussia, B. Response of resting human peripheral blood natural killer cells to interleukin 2. J. Exp. Med. 1984, 160, 1147–1169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trinchieri, G. Immunobiology of interleukin-12. Immunol. Res. 1998, 17, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Viaud, S.; Saccheri, F.; Mignot, G.; Yamazaki, T.; Daillere, R.; Hannani, D.; Enot, D.P.; Pfirschke, C.; Engblom, C.; Pittet, M.J.; et al. The intestinal microbiota modulates the anticancer immune effects of cyclophosphamide. Science 2013, 342, 971–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barreda, D.R.; Belosevic, M. Transcriptional regulation of hemopoiesis. Dev. Comp. Immunol. 2001, 25, 763–789. [Google Scholar] [CrossRef]
- Gill, H.S.; Guarner, F. Probiotics and human health: A clinical perspective. Postgrad. Med. J. 2004, 80, 516–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molina, M.A.; Diaz, A.M.; Hesse, C.; Ginter, W.; Gentilini, M.V.; Nunez, G.G.; Canellada, A.M.; Sparwasser, T.; Berod, L.; Castro, M.S.; et al. Immunostimulatory Effects Triggered by Enterococcus faecalis CECT7121 Probiotic Strain Involve Activation of Dendritic Cells and Interferon-Gamma Production. PLoS ONE 2015, 10, e0127262. [Google Scholar] [CrossRef] [Green Version]
- Meng, Y.; Li, B.; Jin, D.; Zhan, M.; Lu, J.; Huo, G. Immunomodulatory activity of Lactobacillus plantarum KLDS1.0318 in cyclophosphamide-treated mice. Food Nutr. Res. 2018, 62. [Google Scholar] [CrossRef]
- Thomson, A.W.; Mathie, I.H.; Sewell, H.F. Cyclophosphamide-induced eosinophilia in the rat: Concomitant changes in T-cell subsets, B cells and large granular lymphocytes within lymphoid tissues. Immunology 1987, 60, 383–388. [Google Scholar]
- Misra, R.R.; Bloom, S.E. Roles of dosage, pharmacokinetics, and cellular sensitivity to damage in the selective toxicity of cyclophosphamide towards B and T cells in development. Toxicology 1991, 66, 239–256. [Google Scholar] [CrossRef]
- Thapa, P.; Farber, D.L. The Role of the Thymus in the Immune Response. Thorac. Surg. Clin. 2019, 29, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Commins, S.P.; Borish, L.; Steinke, J.W. Immunologic messenger molecules: Cytokines, interferons, and chemokines. J. Allergy Clin. Immunol. 2010, 125, S53–S72. [Google Scholar] [CrossRef] [PubMed]
- Ren, D.; Li, C.; Qin, Y.; Yin, R.; Du, S.; Liu, H.; Zhang, Y.; Wang, C.; Rong, F.; Jin, N. Evaluation of immunomodulatory activity of two potential probiotic Lactobacillus strains by in vivo tests. Anaerobe 2015, 35, 22–27. [Google Scholar] [CrossRef]
- Becker, P.S.; Suck, G.; Nowakowska, P.; Ullrich, E.; Seifried, E.; Bader, P.; Tonn, T.; Seidl, C. Selection and expansion of natural killer cells for NK cell-based immunotherapy. Cancer Immunol. Immunother. 2016, 65, 477–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, Y.; Lu, Y.; Wang, D.; Liu, J.; Song, X.; Zhang, W.; Zhao, X.; Nguyen, T.L.; Hu, Y. Effect of epimedium polysaccharide-propolis flavone immunopotentiator on immunosuppression induced by cyclophosphamide in chickens. Cell Immunol. 2013, 281, 37–43. [Google Scholar] [CrossRef]
- Yoo, H.J.; You, D.J.; Lee, K.W. Characterization and Immunomodulatory Effects of High Molecular Weight Fucoidan Fraction from the Sporophyll of Undaria pinnatifida in Cyclophosphamide-Induced Immunosuppressed Mice. Mar. Drugs 2019, 17, 447. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Cong, L.; Wang, C.; Li, H.; Zhang, C.; Guan, X.; Liu, P.; Xie, Y.; Chen, J.; Sun, J. Immunomodulatory effect of Schisandra polysaccharides in cyclophosphamide-induced immunocompromised mice. Exp. Ther. Med. 2018, 15, 4755–4762. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.G.; Sahoo, A.; Im, S.H. Epigenetic regulation of cytokine gene expression in T lymphocytes. Yonsei Med. J. 2009, 50, 322–330. [Google Scholar] [CrossRef]
- Mukhopadhya, I.; Hansen, R.; El-Omar, E.M.; Hold, G.L. IBD-what role do Proteobacteria play? Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 219–230. [Google Scholar] [CrossRef]
- Woting, A.; Blaut, M. The Intestinal Microbiota in Metabolic Disease. Nutrients 2016, 8, 202. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, C.; Xia, Q.; Liu, D.; Tan, X.; Li, Y.; Cao, Y. Treatment with Subcritical Water-Hydrolyzed Citrus Pectin Ameliorated Cyclophosphamide-Induced Immunosuppression and Modulated Gut Microbiota Composition in ICR Mice. Molecules 2020, 25, 1302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, Y.; Yan, Y.; Chen, D.; Ran, L.; Mi, J.; Lu, L.; Jing, B.; Li, X.; Zeng, X.; Cao, Y. Modulating effects of polysaccharides from the fruits of Lycium barbarum on the immune response and gut microbiota in cyclophosphamide-treated mice. Food Funct. 2019, 10, 3671–3683. [Google Scholar] [CrossRef] [PubMed]
- Montassier, E.; Al-Ghalith, G.A.; Ward, T.; Corvec, S.; Gastinne, T.; Potel, G.; Moreau, P.; de la Cochetiere, M.F.; Batard, E.; Knights, D. Pretreatment gut microbiome predicts chemotherapy-related bloodstream infection. Genome Med. 2016, 8, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopetuso, L.R.; Petito, V.; Graziani, C.; Schiavoni, E.; Paroni Sterbini, F.; Poscia, A.; Gaetani, E.; Franceschi, F.; Cammarota, G.; Sanguinetti, M.; et al. Gut Microbiota in Health, Diverticular Disease, Irritable Bowel Syndrome, and Inflammatory Bowel Diseases: Time for Microbial Marker of Gastrointestinal Disorders. Dig. Dis. 2018, 36, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Naftali, T.; Reshef, L.; Kovacs, A.; Porat, R.; Amir, I.; Konikoff, F.M.; Gophna, U. Distinct Microbiotas are Associated with Ileum-Restricted and Colon-Involving Crohn’s Disease. Inflamm. Bowel Dis. 2016, 22, 293–302. [Google Scholar] [CrossRef]
- Orkin, S.H. Development of the hematopoietic system. Curr. Opin. Genet. Dev. 1996, 6, 597–602. [Google Scholar] [CrossRef]
- Lee, H.S.; Lee, I.H.; Park, S.I.; Lee, D.Y. Network Pharmacology-Based Investigation of the System-Level Molecular Mechanisms of the Hematopoietic Activity of Samul-Tang, a Traditional Korean Herbal Formula. Evid. Based Complement Alternat. Med. 2020, 2020, 9048089. [Google Scholar] [CrossRef]
- Zhao, J.L.; Ma, C.; O’Connell, R.M.; Mehta, A.; DiLoreto, R.; Heath, J.R.; Baltimore, D. Conversion of danger signals into cytokine signals by hematopoietic stem and progenitor cells for regulation of stress-induced hematopoiesis. Cell Stem Cell 2014, 14, 445–459. [Google Scholar] [CrossRef] [Green Version]
- Wilhelmsen, K.; Mesa, K.R.; Prakash, A.; Xu, F.; Hellman, J. Activation of endothelial TLR2 by bacterial lipoprotein upregulates proteins specific for the neutrophil response. Innate Immun. 2012, 18, 602–616. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward | Reverse | Tm (°C) |
|---|---|---|---|
| IFN-γ | GCTTCCTGAGGCTGGATTC | TACCTTCTTCAGCAACAGCAAG | 55 |
| IL-2 | TCCTGGGGAGTTTCAGGTTC | CTCTACAGCGGAAGCACAGC | 55 |
| IL-12 | TCTGCAGAGAAGGTCACACT | ATGAAGAAGCTGGTGCTGTA | 55 |
| β-actin | ATCACTATTGGCAACGAGCG | TCAGCAATGCCTGGGTACAT | 55 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, S.-Y.; Shin, J.-S.; Chung, K.-S.; Han, H.-S.; Lee, H.-H.; Lee, J.-H.; Kim, S.-Y.; Ji, Y.W.; Ha, Y.; Kang, J.; et al. Immunostimulatory Effects of Live Lactobacillus sakei K040706 on the CYP-Induced Immunosuppression Mouse Model. Nutrients 2020, 12, 3573. https://doi.org/10.3390/nu12113573
Kim S-Y, Shin J-S, Chung K-S, Han H-S, Lee H-H, Lee J-H, Kim S-Y, Ji YW, Ha Y, Kang J, et al. Immunostimulatory Effects of Live Lactobacillus sakei K040706 on the CYP-Induced Immunosuppression Mouse Model. Nutrients. 2020; 12(11):3573. https://doi.org/10.3390/nu12113573
Chicago/Turabian StyleKim, Seo-Yeon, Ji-Sun Shin, Kyung-Sook Chung, Hee-Soo Han, Hwi-Ho Lee, Jeong-Hun Lee, Su-Yeon Kim, Yong Woo Ji, Yejin Ha, Jooyeon Kang, and et al. 2020. "Immunostimulatory Effects of Live Lactobacillus sakei K040706 on the CYP-Induced Immunosuppression Mouse Model" Nutrients 12, no. 11: 3573. https://doi.org/10.3390/nu12113573
APA StyleKim, S. -Y., Shin, J. -S., Chung, K. -S., Han, H. -S., Lee, H. -H., Lee, J. -H., Kim, S. -Y., Ji, Y. W., Ha, Y., Kang, J., Rhee, Y. K., & Lee, K. -T. (2020). Immunostimulatory Effects of Live Lactobacillus sakei K040706 on the CYP-Induced Immunosuppression Mouse Model. Nutrients, 12(11), 3573. https://doi.org/10.3390/nu12113573