Saliva Samples as A Tool to Study the Effect of Meal Timing on Metabolic And Inflammatory Biomarkers

 ,
,  , and
, and .png)
Abstract
:1. Introduction
2. Materials and Methods
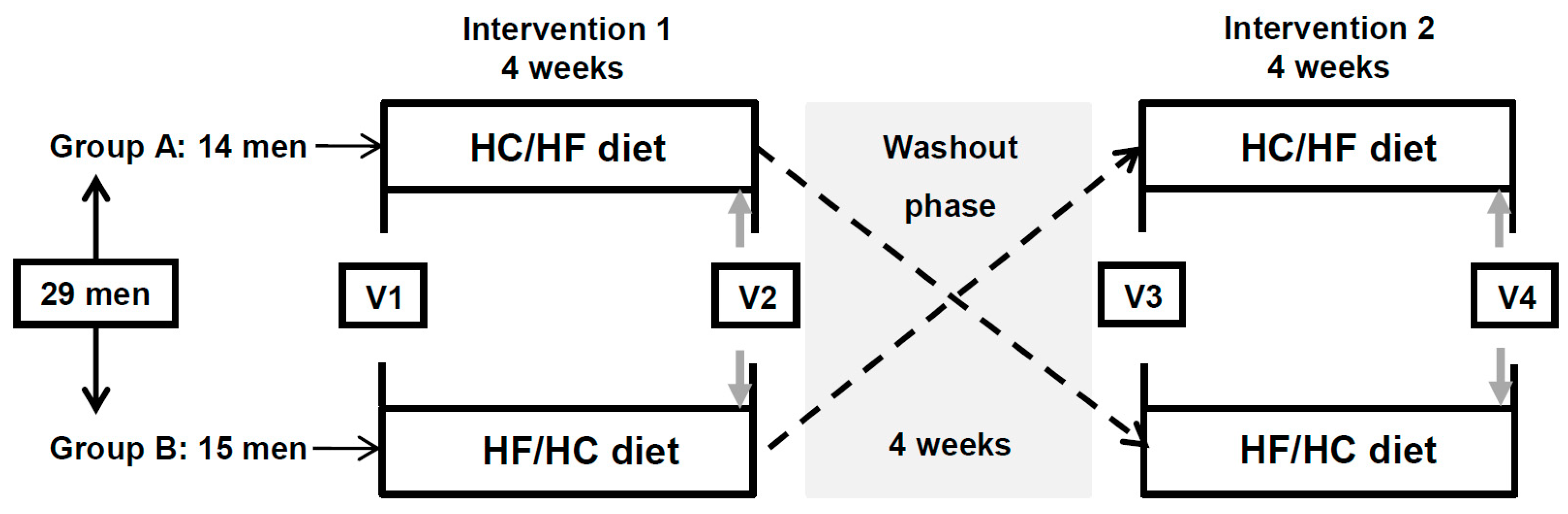
2.1. Study Design
2.2. Saliva Sample Collection
2.3. Sample Analysis
2.4. Statistical Analysis
3. Results
3.1. Study Population and Adherence to Dietary Interventions
3.2. Detectable Salivary Markers
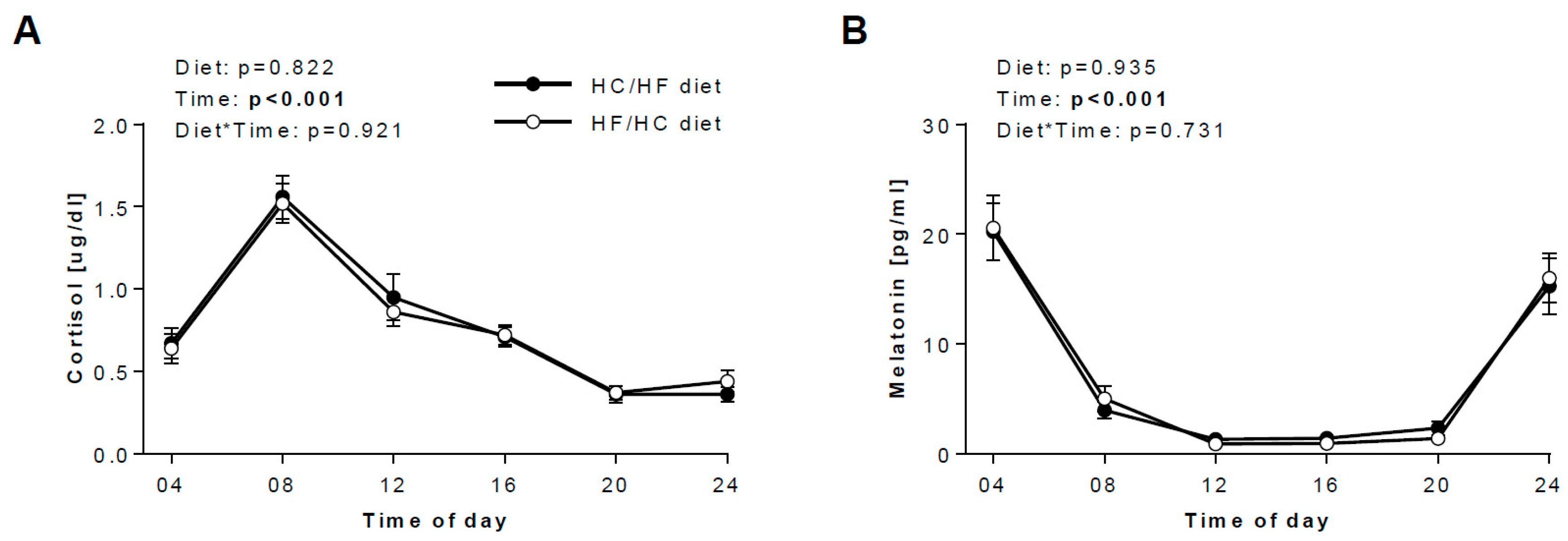
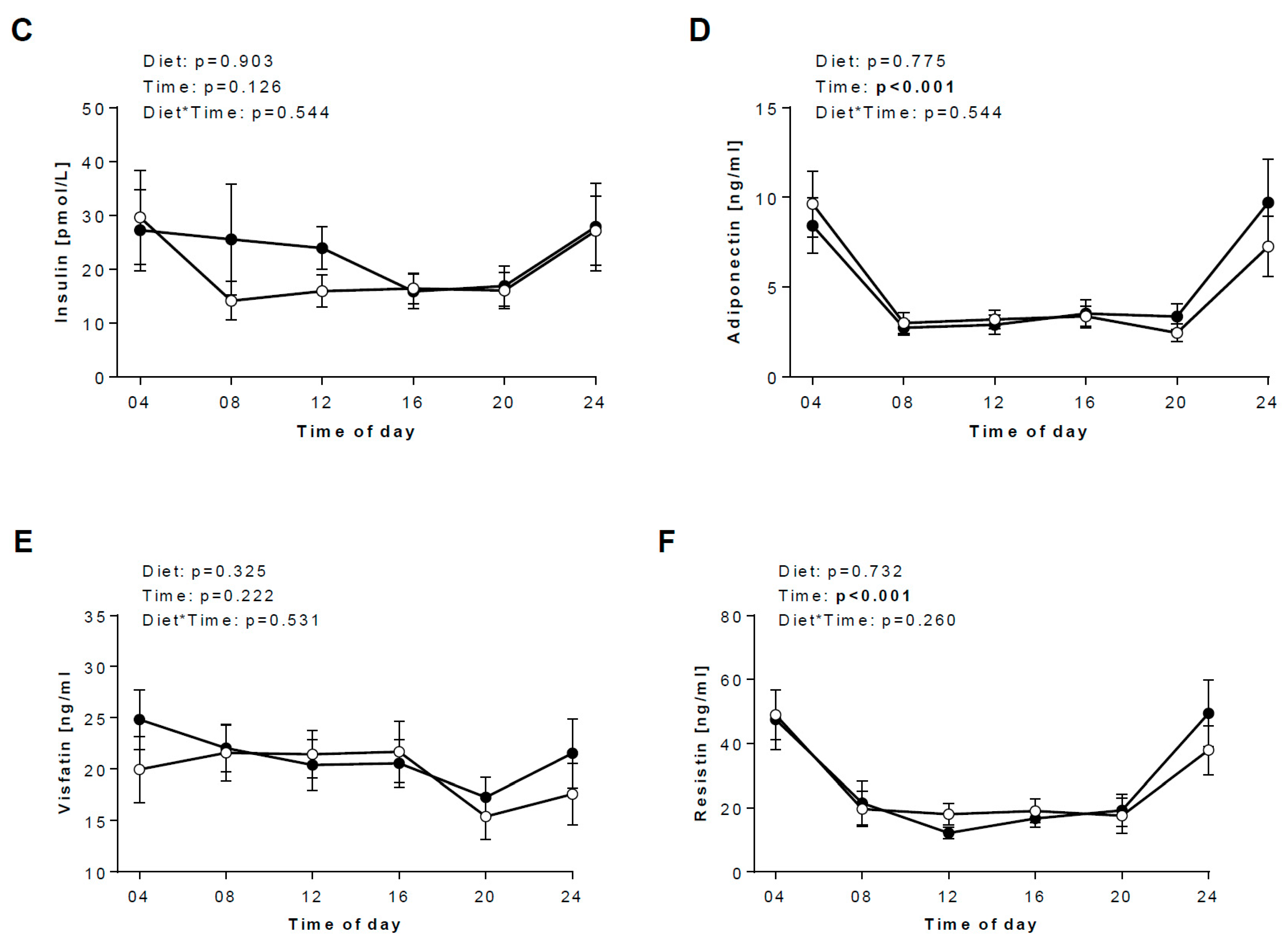
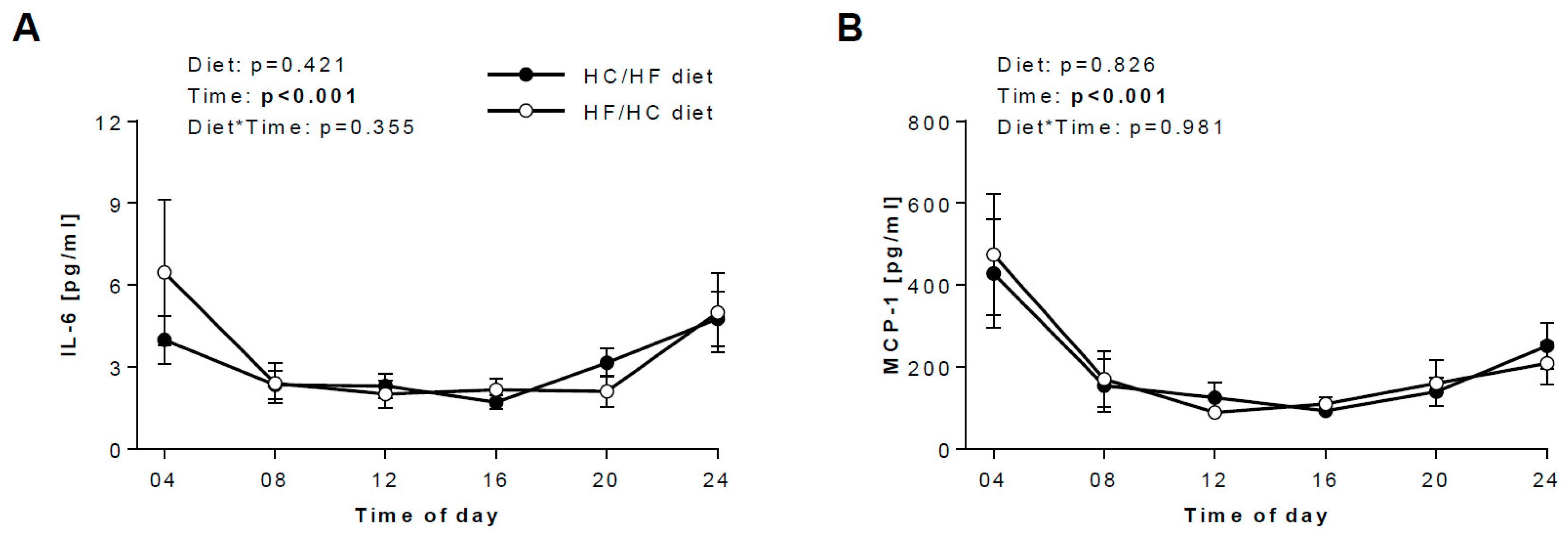
3.3. Diurnal Rhythms of Salivary Markers
3.4. Daily hormonal profiles in response to the diets
4. Discussion
Supplementary Materials
Author Contributions
Financial Support
Acknowledgments
Conflicts of Interest
Abbreviations
| BMI | Body mass index |
| CHO | Carbohydrates |
| CID | Clinical investigation day |
| EN% | Energy percent |
| FA | Fatty acid |
| FFA | Free fatty acid |
| HC/HF | Isocaloric carbohydrate-rich diet until 13:30 and fat-rich diet between 16:30 and 22:00 |
| HF/HC | Isocaloric fat-rich diet until 13:30 and carbohydrate-rich diet between 16:30 and 22:00 |
| IL-6 | Interleukin-6 |
| Kcal | Kilo calories |
| MCP-1 | Monocyte chemoattractant protein-1; |
| MTT-HC | High-carb meal tolerance test |
| MTT-HF | High-fat meal tolerance test |
| SFA | Saturated fatty acids |
References
- Kessler, K.; Pivovarova-Ramich, O. Meal Timing, Aging, and Metabolic Health. Int. J. Mol. Sci. 2019, 20, 1911. [Google Scholar] [CrossRef] [Green Version]
- Arble, D.M.; Bass, J.; Laposky, A.D.; Vitaterna, M.H.; Turek, F.W. Circadian timing of food intake contributes to weight gain. Obesity (Silver Spring) 2009, 17, 2100–2102. [Google Scholar] [CrossRef]
- Chaix, A.; Zarrinpar, A.; Miu, P.; Panda, S. Time-restricted feeding is a preventative and therapeutic intervention against diverse nutritional challenges. Cell Metab. 2014, 20, 991–1005. [Google Scholar] [CrossRef] [Green Version]
- Hatori, M.; Vollmers, C.; Zarrinpar, A.; DiTacchio, L.; Bushong, E.A.; Gill, S.; Leblanc, M.; Chaix, A.; Joens, M.; Fitzpatrick, J.A.; et al. Time-restricted feeding without reducing caloric intake prevents metabolic diseases in mice fed a high-fat diet. Cell Metab. 2012, 15, 848–860. [Google Scholar] [CrossRef] [Green Version]
- Allison, K.C.; Goel, N. Timing of eating in adults across the weight spectrum: Metabolic factors and potential circadian mechanisms. Physiol. Behave.r 2018, 192, 158–166. [Google Scholar] [CrossRef]
- Garaulet, M.; Gomez-Abellan, P.; Alburquerque-Bejar, J.J.; Lee, Y.C.; Ordovas, J.M.; Scheer, F.A. Timing of food intake predicts weight loss effectiveness. Int. J. Obes. (Lond) 2013, 37, 604–611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jakubowicz, D.; Barnea, M.; Wainstein, J.; Froy, O. High caloric intake at breakfast vs. dinner differentially influences weight loss of overweight and obese women. Obesity (Silver Spring) 2013, 21, 2504–2512. [Google Scholar] [CrossRef] [PubMed]
- Bray, M.S. Time-of-day-dependent dietary fat consumption influences multiple cardiometabolic syndrome parameters in mice. Int. J. Obes. (Lond.). 2010, 34, 1589–1598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almoosawi, S.; Prynne, C.J.; Hardy, R.; Stephen, A.M. Time-of-day of energy intake: Association with hypertension and blood pressure 10 years later in the 1946 British Birth Cohort. J. Hypertens. 2013, 31, 882–892. [Google Scholar] [CrossRef] [PubMed]
- Almoosawi, S.; Prynne, C.J.; Hardy, R.; Stephen, A.M. Diurnal eating rhythms: Association with long-term development of diabetes in the 1946 British birth cohort. Nutr. Metab. Cardiovasc. Dis. 2013, 23, 1025–1030. [Google Scholar] [CrossRef]
- Kessler, K.; Hornemann, S.; Petzke, K.J.; Kemper, M.; Kramer, A.; Pfeiffer, A.F.; Pivovarova, O.; Rudovich, N. The effect of diurnal distribution of carbohydrates and fat on glycaemic control in humans: A randomized controlled trial. Sci. Rep. 2017, 7, 44170. [Google Scholar] [CrossRef] [Green Version]
- Kessler, K.; Hornemann, S.; Petzke, K.J.; Kemper, M.; Markova, M.; Rudovich, N.; Grune, T.; Kramer, A.; Pfeiffer, A.F.H.; Pivovarova-Ramich, O. Diurnal distribution of carbohydrates and fat affects substrate oxidation and adipokine secretion in humans. Am. J. Clin. Nutr. 2018, accepted. [Google Scholar] [CrossRef] [PubMed]
- Roblegg, E.; Coughran, A.; Sirjani, D. Saliva: An all-rounder of our body. Eur. J. Pharm. Biopharm. 2019, 142, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Bellagambi, F.G.; Lomonaco, T.; Salvo, P.; Vivaldi, F.; Hangouët, M.; Ghimenti, S.; Biagini, D.; Di Francesco, F.; Fuoco, R.; Errachid, A. Saliva sampling: Methods and devices. An overview. Trends Analyt. Chem. 2020, in press. [Google Scholar] [CrossRef]
- Chiappin, S.; Antonelli, G.; Gatti, R.; De Palo, E.F. Saliva specimen: A new laboratory tool for diagnostic and basic investigation. Clin. Chim. Acta. 2007, 383, 30–40. [Google Scholar] [CrossRef]
- Lomonaco, T.; Ghimenti, S.; Piga, I.; Biagini, D.; Onor, M.; Fuoco, R.; Paolicchi, A.; Ruocco, L.; Pellegrini, G.; Trivella, M.G.; et al. Monitoring of warfarin therapy: Preliminary results from a longitudinal pilot study. Microchem. J. 2018, 136, 170–176. [Google Scholar] [CrossRef]
- Ghimenti, S.; Lomonaco, T.; Onor, M.; Murgia, L.; Paolicchi, A.; Fuoco, R.; Ruocco, L.; Pellegrini, G.; Trivella, M.G.; Di Francesco, F. Measurement of warfarin in the oral fluid of patients undergoing anticoagulant oral therapy. PLoS ONE 2011, 6, e28182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desai, G.S.; Mathews, S.T. Saliva as a non-invasive diagnostic tool for inflammation and insulin-resistance. World J. Diabetes 2014, 5, 730–738. [Google Scholar] [CrossRef] [PubMed]
- Bonne, N.J.; Wong, D.T. Salivary biomarker development using genomic, proteomic and metabolomic approaches. Genome. Med. 2012, 4, 82. [Google Scholar] [CrossRef] [PubMed]
- Dallmann, R.; Viola, A.U.; Tarokh, L.; Cajochen, C.; Brown, S.A. The human circadian metabolome. Proc. Natl. Acad. Sci. USA 2012, 109, 2625–2629. [Google Scholar] [CrossRef] [Green Version]
- Collado, M.C.; Engen, P.A.; Bandin, C.; Cabrera-Rubio, R.; Voigt, R.M.; Green, S.J.; Naqib, A.; Keshavarzian, A.; Scheer, F.; Garaulet, M. Timing of food intake impacts daily rhythms of human salivary microbiota: A randomized, crossover study. FASEB J. 2018, 32, 2060–2072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pirsean, C.; Negut, C.; Stefan-van Staden, R.I.; Dinu-Pirvu, C.E.; Armean, P.; Udeanu, D.I. The salivary levels of leptin and interleukin-6 as potential inflammatory markers in children obesity. PLoS ONE 2019, 14, e0210288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodson, J.M.; Kantarci, A.; Hartman, M.L.; Denis, G.V.; Stephens, D.; Hasturk, H.; Yaskell, T.; Vargas, J.; Wang, X.; Cugini, M.; et al. Metabolic disease risk in children by salivary biomarker analysis. PLoS ONE 2014, 9, e98799. [Google Scholar] [CrossRef] [PubMed]
- Pasic, J.; Pickup, J.C. Salivary insulin in normal and type I diabetic subjects. Diabetes Care 1988, 11, 489–494. [Google Scholar] [CrossRef]
- Browne, R.W.; Kantarci, A.; LaMonte, M.J.; Andrews, C.A.; Hovey, K.M.; Falkner, K.L.; Cekici, A.; Stephens, D.; Genco, R.J.; Scannapieco, F.A.; et al. Performance of multiplex cytokine assays in serum and saliva among community-dwelling postmenopausal women. PLoS ONE 2013, 8, e59498. [Google Scholar] [CrossRef] [Green Version]
- Aydin, S.; Halifeoglu, I.; Ozercan, I.H.; Erman, F.; Kilic, N.; Aydin, S.; Ilhan, N.; Ilhan, N.; Ozkan, Y.; Akpolat, N.; et al. A comparison of leptin and ghrelin levels in plasma and saliva of young healthy subjects. Peptides 2005, 26, 647–652. [Google Scholar] [CrossRef]
- Mamali, I.; Roupas, N.D.; Armeni, A.K.; Theodoropoulou, A.; Markou, K.B.; Georgopoulos, N.A. Measurement of salivary resistin, visfatin and adiponectin levels. Peptides 2012, 33, 120–124. [Google Scholar] [CrossRef]
- Groschl, M.; Rauh, M.; Wagner, R.; Neuhuber, W.; Metzler, M.; Tamguney, G.; Zenk, J.; Schoof, E.; Dorr, H.G.; Blum, W.F.; et al. Identification of leptin in human saliva. J. Clin. Endocrinol. Metab. 2001, 86, 5234–5239. [Google Scholar] [CrossRef] [Green Version]
- Tvarijonaviciute, A.; Martinez-Lozano, N.; Rios, R.; Marcilla de Teruel, M.C.; Garaulet, M.; Ceron, J.J. Saliva as a non-invasive tool for assessment of metabolic and inflammatory biomarkers in children. Clin. Nutr. 2019. [Google Scholar] [CrossRef]
- El-Farhan, N.; Rees, D.A.; Evans, C. Measuring cortisol in serum, urine and saliva - are our assays good enough? Ann. Clin. Biochem. 2017, 54, 308–322. [Google Scholar] [CrossRef]
- Shirakawa, S.; Tsuchiya, S.; Tsutsumi, Y.; Kotorii, T.; Uchimura, N.; Sakamoto, T.; Yamada, S. Time course of saliva and serum melatonin levels after ingestion of melatonin. Psychiatry Clin. Neurosci. 1998, 52, 266–267. [Google Scholar] [CrossRef] [PubMed]
- Reinhardt, E.L.; Fernandes, P.; Markus, R.P.; Fischer, F.M. Night work effects on salivary cytokines TNF, IL-1beta and IL-6. Chronobiol. Int. 2019, 36, 11–26. [Google Scholar] [CrossRef] [PubMed]
- Pivovarova, O.; Jurchott, K.; Rudovich, N.; Hornemann, S.; Lu, Y.; Mockel, S.; Murahovschi, V.; Kessler, K.; Seltmann, A.C.; Maser-Gluth, C.; et al. Changes of dietary fat and carbohydrate content alter central and peripheral clock in humans. J. Clin. Endocrinol. Metab. 2015, jc20143868. [Google Scholar] [CrossRef] [Green Version]
- Asher, G.; Schibler, U. Crosstalk between components of circadian and metabolic cycles in mammals. Cell Metab. 2011, 13, 125–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shostak, A.; Husse, J.; Oster, H. Circadian regulation of adipose function. Adipocyte 2013, 2, 201–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheiermann, C.; Kunisaki, Y.; Frenette, P.S. Circadian control of the immune system. Nat. Rev. Immunol. 2013, 13, 190–198. [Google Scholar] [CrossRef] [Green Version]
- Gavrila, A. Diurnal and ultradian dynamics of serum adiponectin in healthy men: Comparison with leptin, circulating soluble leptin receptor, and cortisol patterns. J. Clin. Endocrinol. Metab. 2003, 88, 2838–2843. [Google Scholar] [CrossRef] [Green Version]
- Oliver, P.; Ribot, J.; Rodriguez, A.M.; Sanchez, J.; Pico, C.; Palou, A. Resistin as a putative modulator of insulin action in the daily feeding/fasting rhythm. Pflugers. Arch. 2006, 452, 260–267. [Google Scholar] [CrossRef]
- Aziz, N.A.; Pijl, H.; Frolich, M.; Roelfsema, F.; Roos, R.A. Leptin, adiponectin, and resistin secretion and diurnal rhythmicity are unaltered in Parkinson’s disease. Mov. Disord. 2011, 26, 760–761. [Google Scholar] [CrossRef]
- Wehrens, S.M.T.; Christou, S.; Isherwood, C.; Middleton, B.; Gibbs, M.A.; Archer, S.N.; Skene, D.J.; Johnston, J.D. Meal Timing Regulates the Human Circadian System. Curr. Biol. CB 2017, 27, 1768–1775. [Google Scholar] [CrossRef] [Green Version]
- Poggiogalle, E.; Jamshed, H.; Peterson, C.M. Circadian regulation of glucose, lipid, and energy metabolism in humans. Metabolism 2018, 84, 11–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lomonaco, T.; Ghimenti, S.; Biagini, D.; Bramanti, E.; Onor, M.; Bellagambi, F.G.; Fuoco, R.; Di Francesco, F. The effect of sampling procedures on the urate in oral fluid and lactate concentration in oral fluid. Microchem. J. 2018, 136, 255–262. [Google Scholar] [CrossRef]
- Lomonaco, T.; Ghimenti, S.; Piga, I.; Biagini, D.; Onor, M.; Fuoco, R.; Di Francesco, F. Influence of sampling on the determination of warfarin and warfarin alcohols in oral fluid. PLoS ONE 2014, 9, e114430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dawes, C.; Ong, B.Y. Circadian rhythms in the concentrations of protein and the main electrolytes in human unstimulated parotid saliva. Arch. Oral. Biol. 1973, 18, 1233–1242. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HC/HF diet | HF/HC diet | P1 | Pcorr2 | |||||
|---|---|---|---|---|---|---|---|---|
| Pre | Post | Δ% | Pre | Post | Δ% | |||
| Weight [kg] | 86.8 ± 2.8 | 86.4 ± 2.9 | −0.5 | 87.1 ± 2.9 | 86.6 ± 2.9 | −0.6 | 0.936 | |
| BMI [kg/m²] | 27.0 ± 0.7 | 26.8 ± 0.8 | −0.7 | 27.1 ± 0.8 | 26.9 ± 0.8 | −0.7 | 0.917 | |
| Total body fat [%] | 24.7 ± 1.5 | 24.2 ± 1.7 | −2.0 | 24.5 ± 1.7 | 23.3 ± 1.7 | −4.9 | 0.206 | |
| Triglycerides [mmol/L] | 1.15 ± 0.13 | 1.07 ± 0.11 | −7.0 | 1.25 ± 0.16 | 1.17 ± 0.11 | −6.4 | 0.666 | 0.666 |
| Total cholesterol [mmol/L] | 5.22 ± 0.17 | 4.78 ± 0.16 | −8.4 | 5.24 ± 0.18 | 4.88 ± 0.18 | −6.9 | 0.347 | 0.273 |
| HDL cholesterol [mmol/L] | 1.24 ± 0.04 | 1.08 ± 0.03 | −12.9 | 1.21 ± 0.04 | 1.08 ± 0.03 | −10.7 | 0.737 | 0.704 |
| LDL cholesterol [mmol/L] | 3.46 ± 0.16 | 3.21 ± 0.15 | −7.2 | 3.46 ± 0.17 | 3.27 ± 0.16 | −5.5 | 0.571 | 0.527 |
| NEFA [mmol/L] | 0.50 ± 0.03 | 0.45 ± 0.02 | −10.0 | 0.49 ± 0.03 | 0.45 ± 0.03 | −8.2 | 0.238 | 0.233 |
| Glucose [mmol/L] | 5.91 ± 0.12 | 5.35 ± 0.07 | −9.5** | 5.84 ± 0.1 | 5.33 ± 0.08 | −8.7** | 0.407 | 0.415 |
| Insulin [pmol/L] | 38.46 ± 4.92 | 33.06 ± 2.58 | −14.0 | 41.22 ± 5.52 | 33.72 ± 4.02 | −18.2 | 0.524 | 0.525 |
| HOMA−IR [mmol· mU· l−2] | 1.67 ± 0.29 | 1.29 ± 0.12 | −22.8 | 1.70 ± 0.29 | 1.24 ± 0.16 | −27.1 | 0.537 | 0.539 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kessler, K.; Hornemann, S.; Rudovich, N.; Weber, D.; Grune, T.; Kramer, A.; Pfeiffer, A.F.H.; Pivovarova-Ramich, O. Saliva Samples as A Tool to Study the Effect of Meal Timing on Metabolic And Inflammatory Biomarkers. Nutrients 2020, 12, 340. https://doi.org/10.3390/nu12020340
Kessler K, Hornemann S, Rudovich N, Weber D, Grune T, Kramer A, Pfeiffer AFH, Pivovarova-Ramich O. Saliva Samples as A Tool to Study the Effect of Meal Timing on Metabolic And Inflammatory Biomarkers. Nutrients. 2020; 12(2):340. https://doi.org/10.3390/nu12020340
Chicago/Turabian StyleKessler, Katharina, Silke Hornemann, Natalia Rudovich, Daniela Weber, Tilman Grune, Achim Kramer, Andreas F. H. Pfeiffer, and Olga Pivovarova-Ramich. 2020. "Saliva Samples as A Tool to Study the Effect of Meal Timing on Metabolic And Inflammatory Biomarkers" Nutrients 12, no. 2: 340. https://doi.org/10.3390/nu12020340
APA StyleKessler, K., Hornemann, S., Rudovich, N., Weber, D., Grune, T., Kramer, A., Pfeiffer, A. F. H., & Pivovarova-Ramich, O. (2020). Saliva Samples as A Tool to Study the Effect of Meal Timing on Metabolic And Inflammatory Biomarkers. Nutrients, 12(2), 340. https://doi.org/10.3390/nu12020340