Energy Restriction Enhances Adult Hippocampal Neurogenesis-Associated Memory after Four Weeks in an Adult Human Population with Central Obesity; a Randomized Controlled Trial

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Standards
2.2. Participant Selection
2.3. Study Protocol
2.4. Mnemonic Similarity Task
2.5. Anthropometry
2.6. Statistics
3. Results
3.1. Cohort Characteristics
3.2. Compliance and Data Exclusion
3.3. Energy Restriction, Regardless of Method, Causes Weight Loss and Changes in Body Composition
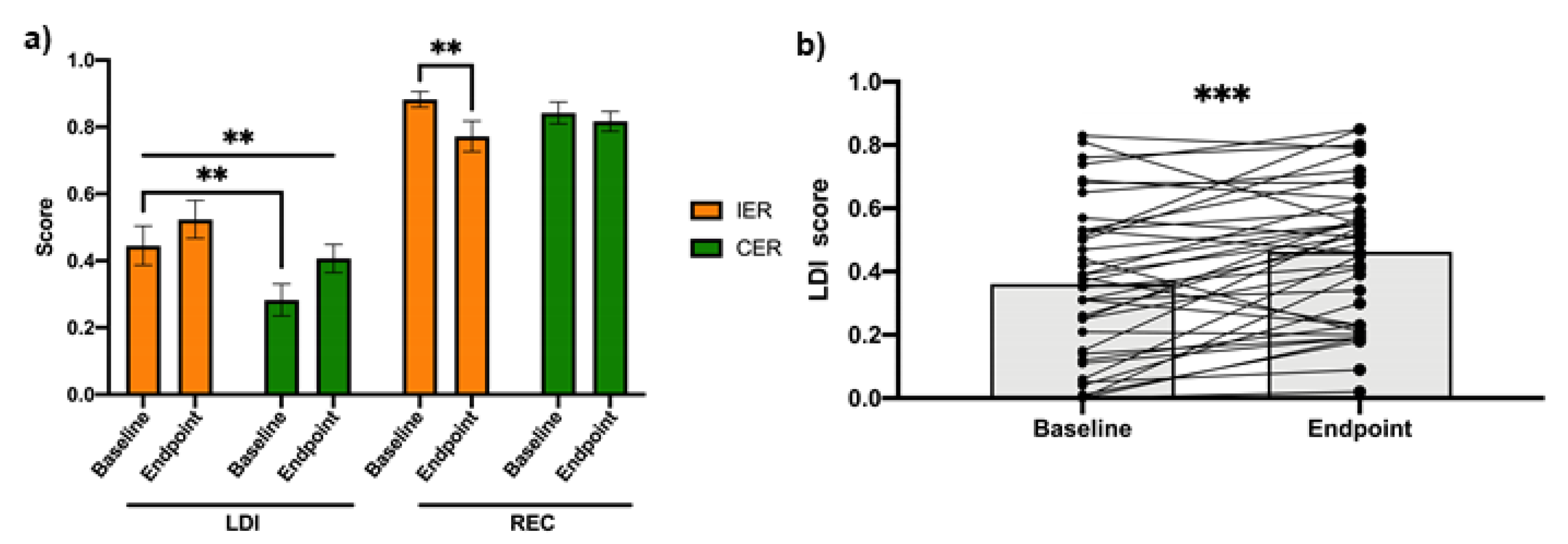
3.4. Energy Restriction, Regardless of Method, Improves Adult-Hippocampal Neurogenesis Dependent Cognition
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zamroziewicz, M.K.; Barbey, A.K. Nutritional Cognitive Neuroscience: Innovations for Healthy Brain Aging. Front Neurosci. 2016, 10, 240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kesby, J.P.; Kim, J.J.; Scadeng, M.; Woods, G.; Kado, D.M.; Olefsky, J.M.; Jeste, D.V.; Achim, C.L.; Semenova, S. Spatial cognition in adult and aged mice exposed to high-fat diet. PLoS ONE 2015, 10, e0140034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Underwood, E.L.; Thompson, L.T. High-fat diet impairs spatial memory and hippocampal intrinsic excitability and sex-dependently alters circulating insulin and hippocampal insulin sensitivity. Biol. Sex Differ. 2016, 7, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haapala, E.A.; Eloranta, A.M.; Venäläinen, T.; Schwab, U.; Lindi, V.; Lakka, T.A. Associations of diet quality with cognition in children-the Physical Activity and Nutrition in Children Study. Br. J. Nutr. 2015, 114, 1080–1087. [Google Scholar] [CrossRef] [Green Version]
- Kamo, T.; Nishida, Y. Direct and indirect effects of nutritional status, physical function and cognitive function on activities of daily living in Japanese older adults requiring long-term care. Geriatr. Gerontol. Int. 2014, 14, 799–805. [Google Scholar] [CrossRef]
- Kempermann, G.; Gage, F.H.; Aigner, L.; Song, H.; Curtis, M.A.; Thuret, S.; Kuhn, H.G.; Jessberger, S.; Frankland, P.W.; Cameron, H.A.; et al. Human Adult Neurogenesis: Evidence and Remaining Questions. Cell Stem Cell 2018, 23, 25–30. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.; Thuret, S. Adult Human Hippocampal Neurogenesis: Controversy and Evidence. Trends Mol. Med. 2018, 24, 521–522. [Google Scholar] [CrossRef] [Green Version]
- Eriksson, P.S.; Perfilieva, E.; Björk-Eriksson, T.; Alborn, A.M.; Nordborg, C.; Peterson, D.A.; Gage, F.H. Neurogenesis in the adult human hippocampus. Nat. Med. 1998, 4, 1313–1317. [Google Scholar] [CrossRef]
- Spalding, K.L.; Bergmann, O.; Alkass, K.; Bernard, S.; Salehpour, M.; Huttner, H.B.; Boström, E.; Westerlund, I.; Vial, C.; Buchholz, B.A.; et al. Dynamics of hippocampal neurogenesis in adult humans. Cell 2013, 153, 1219. [Google Scholar] [CrossRef] [Green Version]
- Sorrells, S.F.; Paredes, M.F.; Cebrian-Silla, A.; Sandoval, K.; Qi, D.; Kelley, K.W.; James, D.; Mayer, S.; Chang, J.; Auguste, K.I.; et al. Human hippocampal neurogenesis drops sharply in children to undetectable levels in adults. Nature 2018, 555, 377–381. [Google Scholar] [CrossRef]
- Boldrini, M.; Fulmore, C.A.; Tartt, A.N.; Simeon, L.R.; Pavlova, I.; Poposka, V.; Rosoklija, G.B.; Stankov, A.; Arango, V.; Dwork, A.J.; et al. Human Hippocampal Neurogenesis Persists throughout Aging. Cell Stem Cell 2018, 22, 589–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno-Jiménez, E.P.; Flor-García, M.; Terreros-Roncal, J.; Rábano, A.; Cafini, F.; Pallas-Bazarra, N.; Ávila, J.; Llorens-Martín, M. Adult hippocampal neurogenesis is abundant in neurologically healthy subjects and drops sharply in patients with Alzheimer’s disease. Nature Medicine. Nat. Publ. Group 2019, 25, 554–560. [Google Scholar]
- Gonçalves, J.T.; Schafer, S.T.; Gage, F.H. Adult Neurogenesis in the Hippocampus: From Stem Cells to Behavior. Cell Cell Press 2016, 167, 897–914. [Google Scholar] [CrossRef] [Green Version]
- Toda, T.; Gage, F.H. Review: Adult neurogenesis contributes to hippocampal plasticity. Cell Tissue Res. 2018, 373, 693–709. [Google Scholar] [CrossRef] [PubMed]
- Anacker, C.; Luna, V.M.; Stevens, G.S.; Millette, A.; Shores, R.; Jimenez, J.C.; Chen, B.; Hen, R. Hippocampal neurogenesis confers stress resilience by inhibiting the ventral dentate gyrus. Nature 2018, 559, 98–102. [Google Scholar] [CrossRef] [PubMed]
- Cameron, H.A.; Glover, L.R. Adult Neurogenesis: Beyond Learning and Memory. Annu. Rev. Psychol. 2015, 66, 53–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fanselow, M.S.; Dong, H.W. Are the Dorsal and Ventral Hippocampus Functionally Distinct Structures? Neuron 2010, 65, 7–19. [Google Scholar] [CrossRef] [Green Version]
- Stark, S.M.; Yassa, M.A.; Lacy, J.W.; Stark, C.E.L. A task to assess behavioral pattern separation (BPS) in humans: Data from healthy aging and mild cognitive impairment. Neuropsychologia 2013, 51, 2442–2449. [Google Scholar] [CrossRef] [Green Version]
- Bakker, A.; Kirwan, C.B.; Miller, M.; Stark, C.E.L. Pattern separation in the human hippocampal CA3 and dentate gyrus. Science 2008, 319, 1640–1642. [Google Scholar] [CrossRef] [Green Version]
- Lacy, J.W.; Yassa, M.A.; Stark, S.M.; Muftuler, L.T.; Stark, C.E.L. Distinct pattern separation related transfer functions in human CA3/dentate and CA1 revealed using highresolution fMRI and variable mnemonic similarity. Learn. Mem. 2011, 18, 15–18. [Google Scholar] [CrossRef] [Green Version]
- Clelland, C.D.; Choi, M.; Romberg, C.; Clemenson, G.D.; Fragniere, A.; Tyers, P.; Jessberger, S.; Saksida, L.M.; Barker, R.A.; Gage, F.H.; et al. A functional role for adult hippocampal neurogenesis in spatial pattern separation. Science 2009, 325, 210–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahay, A.; Scobie, K.N.; Hill, A.S.; O’Carroll, C.M.; Kheirbek, M.A.; Burghardt, N.S.; Fenton, A.A.; Dranovsky, A.; Hen, R. Increasing adult hippocampal neurogenesis is sufficient to improve pattern separation. Nature 2011, 472, 466–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gould, E.; Reeves, A.J.; Fallah, M.; Tanapat, P.; Gross, C.G.; Fuchs, E. Hippocampal neurogenesis in adult Old World primates. Proc. Natl. Acad. Sci. USA 1999, 96, 5263–5267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, R.E. Recognition memory: An old idea given new life. Curr. Biol. 2013, 23, 725–727. [Google Scholar] [CrossRef] [Green Version]
- Bird, C.M. The role of the hippocampus in recognition memory. Cortex 2017, 93, 155–165. [Google Scholar] [CrossRef] [Green Version]
- Bachevalier, J.; Nemanic, S.; Alvarado, M.C. The influence of context on recognition memory in monkeys: Effects of hippocampal, parahippocampal and perirhinal lesions. Behav. Brain Res. 2015, 285, 89–98. [Google Scholar] [CrossRef] [Green Version]
- Cohen, S.J.; Stackman, R.W. Assessing rodent hippocampal involvement in the novel object recognition task. A review. Behav. Brain Res. 2015, 285, 105–117. [Google Scholar] [CrossRef]
- Lafenêtre, P.; Leske, O.; Ma-Högemeie, Z.; Haghikia, A.; Bichler, Z.; Wahle, P.; Heumann, R. Exercise can rescue recognition memory impairment in a model with reduced adult hippocampal neurogenesis. Front Behav. Neurosci. 2010, 3, 34. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.I.; Lee, J.W.; Lee, Y.A.; Lee, D.H.; Han, N.S.; Choi, Y.K.; Hwang, B.R.; Kim, H.J.; Han, J.S. Sexual activity counteracts the suppressive effects of chronic stress on adult hippocampal neurogenesis and recognition memory. Brain Res. 2013, 1538, 26–40. [Google Scholar] [CrossRef]
- Zainuddin, M.S.A.; Thuret, S. Nutrition, adult hippocampal neurogenesis and mental health. Br. Med. Bull. 2012, 103, 89–114. [Google Scholar] [CrossRef] [Green Version]
- Mattson, M.P. Neuroprotective signaling and the aging brain: Take away my food and let me run. Brain Res. 2000, 886, 47–53. [Google Scholar] [CrossRef]
- Horne, B.D.; Muhlestein, J.B.; Anderson, J.L. Health effects of intermittent fasting: Hormesis or harm? A systematic review. Am. J. Clin. Nutr. 2015, 102, 464–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, T.; Dias, G.P.; Thuret, S. Effects of diet on brain plasticity in animal and human studies: Mind the gap. Neural Plast. 2014, 2014, 563160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mattson, M.P.; Longo, V.D.; Harvie, M. Impact of intermittent fasting on health and disease processes. Ageing Res. Rev. 2017, 39, 46–58. [Google Scholar] [CrossRef] [PubMed]
- Pinto, A.M.; Bordoli, C.; Buckner, L.P.; Kim, C.; Kaplan, P.C.; Del Arenal, I.M.; Jeffcock, E.J.; Hall, W.L. Intermittent energy restriction is comparable to continuous energy restriction for cardiometabolic health in adults with central obesity: A randomized controlled trial; the Met-IER study. Clin. Nutr. 2019. [Google Scholar] [CrossRef]
- Stark, S.M.; Stevenson, R.; Wu, C.; Rutledge, S.; Stark, C.E.L. Stability of age-related deficits in the mnemonic similarity task across task variations. Behav. Neurosci. 2015, 129, 257–268. [Google Scholar] [CrossRef]
- Brownlow, M.L.; Joly-Amado, A.; Azam, S.; Elza, M.; Selenica, M.L.; Pappas, C.; Small, B.; Engelman, R.; Gordon, M.N.; Morgan, D. Partial rescue of memory deficits induced by calorie restriction in a mouse model of tau deposition. Behav. Brain Res. 2014, 271, 79–88. [Google Scholar] [CrossRef]
- Carter, C.S.; Leeuwenburgh, C.; Daniels, M.; Foster, T.C. Influence of calorie restriction on measures of age-related cognitive decline: Role of increased physical activity. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2009, 64, 850–859. [Google Scholar] [CrossRef] [Green Version]
- Wu, P.; Jiang, C.; Shen, Q.; Hu, Y. Systematic gene expression profile of hypothalamus in calorie-restricted mice implicates the involvement of mTOR signaling in neuroprotective activity. Mech. Ageing Dev. 2009, 130, 602–610. [Google Scholar] [CrossRef]
- Kaptan, Z.; Akgün-Dar, K.; Kapucu, A.; Dedeakayoʇullari, H.; Batu, Ş.; Üzüm, G. Long term consequences on spatial learning-memory of low-calorie diet during adolescence in female rats; Hippocampal and prefrontal cortex BDNF level, expression of NeuN and cell proliferation in dentate gyrus. Brain Res. 2015, 1618, 194–204. [Google Scholar] [CrossRef]
- Li, L.; Wang, Z.; Zuo, Z. Chronic intermittent fasting improves cognitive functions and brain structures in mice. PLoS ONE 2013, 8, e66069. [Google Scholar] [CrossRef] [Green Version]
- Ahn, J.H.; Shin, B.N.; Song, M.; Kim, H.; Park, J.H.; Lee, T.K.; Park, C.W.; Park, Y.E.; Lee, J.C.; Yong, J.H. Intermittent fasting increases the expressions of SODs and catalase in granule and polymorphic cells and enhances neuroblast dendrite complexity and maturation in the adult gerbil dentate gyrus. Mol. Med. Rep. 2019, 19, 1721–1727. [Google Scholar] [CrossRef] [PubMed]
- Geng, Y.Q.; Guan, J.T.; Xu, M.Y.; Xu, X.H.; Fu, Y.C. Behavioral Study of Calorie-restricted Rats from Early Old Age. In Proceedings of the 2007 29th Annual International Conference of the IEEE Engineering in Medicine and Biology Society, Lyon, France, 22–26 August 2007; pp. 2393–2395. [Google Scholar]
- Wu, P.; Shen, Q.; Dong, S.; Xu, Z.; Tsien, J.Z.; Hu, Y. Calorie restriction ameliorates neurodegenerative phenotypes in forebrain-specific presenilin-1 and presenilin-2 double knockout mice. Neurobiol. Aging 2008, 29, 1502–1511. [Google Scholar] [CrossRef] [PubMed]
- Longo, V.D.; Mattson, M.P. Fasting: Molecular Mechanisms and Clinical Applications. Cell Metab. 2014, 19, 181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.; Seroogy, K.B.; Mattson, M.P. Dietary restriction enhances neurotrophin expression and neurogenesis in the hippocampus of adult mice. J. Neurochem. 2002, 80, 539–547. [Google Scholar] [CrossRef] [PubMed]
- Michael Anson, R.; Guo, Z.; de Cabo, R.; Iyun, T.; Rios, M.; Hagepanos, A.; Ingram, D.K.; Lane, M.A.; Mattson, M.P. Intermittent fasting dissociates beneficial effects of dietary restriction on glucose metabolism and neuronal resistance to injury from calorie intake. Proc. Natl. Acad. Sci. USA 2003, 100, 6216–6220. [Google Scholar] [CrossRef] [Green Version]
- Maruszak, A.; Pilarski, A.; Murphy, T.; Branch, N.; Thuret, S. Hippocampal neurogenesis in Alzheimer’s disease: Is there a role for dietary modulation? J. Alzheimer’s Dis. 2014, 38, 11–38. [Google Scholar] [CrossRef]
- Manzanero, S.; Erion, J.R.; Santro, T.; Steyn, F.J.; Chen, C.; Arumugam, T.V.; Stranahan, A.M. Intermittent fasting attenuates increases in neurogenesis after ischemia and reperfusion and improves recovery. J. Cereb. Blood Flow Metab. 2014, 34, 897–905. [Google Scholar] [CrossRef] [Green Version]
- Komatsu, T.; Chiba, T.; Yamaza, H.; Yamashita, K.; Shimada, A.; Hoshiyama, Y.; Henmi, T.; Ohtani, H.; Higami, Y.; de Cabo, R.; et al. Manipulation of caloric content but not diet composition, attenuates the deficit in learning and memory of senescence-accelerated mouse strain P8. Exp. Gerontol. 2008, 43, 339–346. [Google Scholar] [CrossRef]
- Wei, M.; Brandhorst, S.; Shelehchi, M.; Mirzaei, H.; Cheng, C.W.; Budniak, J.; Groshen, S.; Mack, W.J.; Guen, E.; Di Biase, S.; et al. Fasting-mimicking diet and markers/risk factors for aging, diabetes, cancer, and cardiovascular disease. Sci. Transl. Med. 2017, 9, 8700. [Google Scholar] [CrossRef]
- Madeo, F.; Carmona-Gutierrez, D.; Hofer, S.J.; Kroemer, G. Caloric Restriction Mimetics against Age-Associated Disease: Targets, Mechanisms, and Therapeutic Potential. Cell Metab. 2019, 29, 592–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vallejo, E. Hunger diet on alternate days in the nutrition of the aged. Prensa Med. Argent. 1957, 44, 119–120. [Google Scholar] [PubMed]
- Redman, L.M.; Ravussin, E. Caloric restriction in humans: Impact on physiological, psychological, and behavioral outcomes. Antioxid. Redox Signal. 2011, 14, 275–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stekovic, S.; Hofer, S.J.; Tripolt, N.; Aon, M.A.; Royer, P.; Pein, L.; Stadler, J.T.; Pendl, T.; Prietl, B.; Url, J.; et al. Alternate Day Fasting Improves Physiological and Molecular Markers of Aging in Healthy, Non-obese Humans. Cell Metab. 2019, 30, 462–476. [Google Scholar] [CrossRef] [PubMed]
- Anton, S.D.; Lee, S.A.; Donahoo, W.T.; McLaren, C.; Manini, T.; Leeuwenburgh, C.; Pahor, M. The effects of time restricted feeding on overweight, older adults: A pilot study. Nutrients 2019, 11, 1500. [Google Scholar] [CrossRef] [Green Version]
- Tinsley, G.M.; La Bounty, P.M. Effects of intermittent fasting on body composition and clinical health markers in humans. Nutr. Rev. 2015, 73, 661–674. [Google Scholar] [CrossRef]
- Sutton, E.F.; Beyl, R.; Early, K.S.; Cefalu, W.T.; Ravussin, E.; Peterson, C.M. Early Time-Restricted Feeding Improves Insulin Sensitivity, Blood Pressure, and Oxidative Stress Even without Weight Loss in Men with Prediabetes. Cell Metab. 2018, 27, 1212–1221. [Google Scholar] [CrossRef] [Green Version]
- Prehn, K.; Jumpertz von Schwartzenberg, R.; Mai, K.; Zeitz, U.; Witte, A.V.; Hampel, D.; Szela, A.M.; Fabian, S.; Grittner, U.; Spranger, J.; et al. Caloric Restriction in Older Adults—Differential Effects of Weight Loss and Reduced Weight on Brain Structure and Function. Cereb. Cortex 2016, 27, 1765–1778. [Google Scholar] [CrossRef]
- Hussin, N.M.; Shahar, S.; Teng, N.I.M.F.; Ngah, W.Z.W.; Das, S.K. Efficacy of Fasting and Calorie Restriction (FCR) on mood and depression among ageing men. J. Nutr. Health Aging 2013, 17, 674–680. [Google Scholar] [CrossRef]
- Fitzgerald, K.C.; Vizthum, D.; Henry-Barron, B.; Schweitzer, A.; Cassard, S.D.; Kossoff, E.; Hartman, A.L.; Kapogiannis, D.; Sullivan, P.; Baer, D.J.; et al. Effect of intermittent vs. daily calorie restriction on changes in weight and patient-reported outcomes in people with multiple sclerosis. Mult. Scler. Relat. Disord. 2018, 23, 33–39. [Google Scholar] [CrossRef]
- Jamshed, H.; Beyl, R.A.; Manna, D.L.D.; Yang, E.S.; Ravussin, E.; Peterson, C.M. Early time-restricted feeding improves 24-hour glucose levels and affects markers of the circadian clock, aging, and autophagy in humans. Nutrients 2019, 11, 1234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hullinger, R.; Puglielli, L. Molecular and cellular aspects of age-related cognitive decline and Alzheimer’s disease. Behav. Brain Res. 2017, 322, 191–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakashiba, T.; Cushman, J.D.; Pelkey, K.A.; Renaudineau, S.; Buhl, D.L.; McHugh, T.J.; Rodriguez Barrera, V.; Chittajallu, R.; Iwamoto, K.S.; McBain, C.J.; et al. Young dentate granule cells mediate pattern separation, whereas old granule cells facilitate pattern completion. Cell 2012, 149, 188–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oreopoulos, A.; Kalantar-Zadeh, K.; Sharma, A.M.; Fonarow, G.C. The obesity paradox in the elderly: Potential mechanisms and clinical implications. Clin. Geriatr. Med. 2009, 25, 643–659. [Google Scholar] [CrossRef] [PubMed]
- Sellbom, K.S.; Gunstad, J. Cognitive function and decline in obesity. J. Alzheimers Dis. 2012, 30, 89–95. [Google Scholar] [CrossRef]
- Kuo, H.-K.; Jones, R.N.; Milberg, W.P.; Tennstedt, S.; Talbot, L.; Morris, J.N.; Lipsitz, L.A. Cognitive function in normal-weight, overweight, and obese older adults: An analysis of the Advanced Cognitive Training for Independent and Vital Elderly cohort. J. Am. Geriatr. Soc. 2006, 54, 97–103. [Google Scholar] [CrossRef] [Green Version]
- Memel, M.; Bourassa, K.; Woolverton, C.; Sbarra, D.A. Body Mass and Physical Activity Uniquely Predict Change in Cognition for Aging Adults. Ann. Behav. Med. 2016, 50, 397–408. [Google Scholar] [CrossRef]
- Skinner, J.S.; Abel, W.M.; McCoy, K.; Wilkins, C.H. Exploring the “obesity paradox” as a correlate of cognitive and physical function in community-dwelling black and white older adults. Ethn. Dis. 2017, 27, 387–394. [Google Scholar] [CrossRef] [Green Version]
- Hariton, E.; Locascio, J.J. Randomised controlled trials—The gold standard for effectiveness research: Study design: Randomised controlled trials. Int. J. Obstet. Gynaecol. 2018, 125, 1716. [Google Scholar] [CrossRef] [Green Version]
- Stark, S.M.; Kirwan, C.B.; Stark, C.E.L. Mnemonic Similarity Task: A Tool for Assessing Hippocampal Integrity. Trends Cogn. Sci. 2019, 23, 938–951. [Google Scholar] [CrossRef]
- Ho, N.F.; Hooker, J.M.; Sahay, A.; Holt, D.J.; Roffman, J.L. In vivo imaging of adult human hippocampal neurogenesis: Progress, pitfalls and promise. Mol. Psychiatry 2013, 18, 404–416. [Google Scholar] [CrossRef] [PubMed]
- Rounds, T.; Harvey, J. Enrollment Challenges: Recruiting Men to Weight Loss Interventions. Am. J. Mens Health 2019, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snyder, J.S.; Soumier, A.; Brewer, M.; Pickel, J.; Cameron, H.A. Adult hippocampal neurogenesis buffers stress responses and depressive behaviour. Nature 2011, 476, 458–462. [Google Scholar] [CrossRef] [PubMed]
- Kempermann, G. Environmental enrichment, new neurons and the neurobiology of individuality. Nat. Rev. Neuroscience 2019, 20, 235–245. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Characteristic | IER (n = 20) | CER (n = 23) | P Value |
|---|---|---|---|
| Age (years) | 50.0 ± 12.7 | 55.7 ± 8.2 | 0.066 |
| Weight (kg) | 87.6 ± 16.8 | 89.1 ± 20.0 | 0.820 |
| BMI (kg/m2) | 32.0 ± 4.7 | 30.9 ± 5.6 | 0.529 |
| Waist circumference (cm) | 107.7 ± 9.3 | 110.5 ± 15.8 | 0.971 |
| Hip circumference (cm) | 114.4 ± 8.8 | 115.7 ± 12.6 | 0.686 |
| Waist to hip ratio | 0.9 ± 0.1 | 1.0 ± 0.1 | 0.557 |
| Body fat (%) | 39.9 ± 6.5 | 37.5 ± 7.1 | 0.258 |
| Resting metabolic rate (kcal/day) | 1404.2 ± 276.6 | 1378.2 ± 304.1 | 0.772 |
| Physical activity levels | 1.44 ± 0.2 | 1.45 ± 0.1 | 0.925 |
| Total energy expenditure (kcal/day) | 1999.1 ± 357.1 | 2036.2 ± 476.9 | 0.818 |
| Characteristic | IER (n = 18) | CER (n = 21) | P Value |
|---|---|---|---|
| % Change in weight (kg) | −3.1 ± 1.6 | −2.8 ± 1.6 | 0.578 |
| % Change in BMI (kg/m2) | −3.1 ± 1.5 | −2.8 ± 1.6 | 0.631 |
| % Change in waist circumference (cm) | −3.8 ± 2.4 | −4.3 ± 2.9 | 0.589 |
| % Change in hip circumference (cm) | −1.9 ± 1.6 | −2.0 ± 2.7 | 0.932 |
| % Change in waist to hip ratio | −1.9 ± 2.5 | −2.3 ± 3.1 | 0.675 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, C.; Pinto, A.M.; Bordoli, C.; Buckner, L.P.; Kaplan, P.C.; del Arenal, I.M.; Jeffcock, E.J.; Hall, W.L.; Thuret, S. Energy Restriction Enhances Adult Hippocampal Neurogenesis-Associated Memory after Four Weeks in an Adult Human Population with Central Obesity; a Randomized Controlled Trial. Nutrients 2020, 12, 638. https://doi.org/10.3390/nu12030638
Kim C, Pinto AM, Bordoli C, Buckner LP, Kaplan PC, del Arenal IM, Jeffcock EJ, Hall WL, Thuret S. Energy Restriction Enhances Adult Hippocampal Neurogenesis-Associated Memory after Four Weeks in an Adult Human Population with Central Obesity; a Randomized Controlled Trial. Nutrients. 2020; 12(3):638. https://doi.org/10.3390/nu12030638
Chicago/Turabian StyleKim, Curie, Ana Margarida Pinto, Claire Bordoli, Luke Patrick Buckner, Polly Charlotte Kaplan, Ines Maria del Arenal, Emma Jane Jeffcock, Wendy L Hall, and Sandrine Thuret. 2020. "Energy Restriction Enhances Adult Hippocampal Neurogenesis-Associated Memory after Four Weeks in an Adult Human Population with Central Obesity; a Randomized Controlled Trial" Nutrients 12, no. 3: 638. https://doi.org/10.3390/nu12030638
APA StyleKim, C., Pinto, A. M., Bordoli, C., Buckner, L. P., Kaplan, P. C., del Arenal, I. M., Jeffcock, E. J., Hall, W. L., & Thuret, S. (2020). Energy Restriction Enhances Adult Hippocampal Neurogenesis-Associated Memory after Four Weeks in an Adult Human Population with Central Obesity; a Randomized Controlled Trial. Nutrients, 12(3), 638. https://doi.org/10.3390/nu12030638