Omega-3 Polyunsaturated Fatty Acids Inhibit the Function of Human URAT1, a Renal Urate Re-Absorber

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Culture
2.3. Preparation of Protein Lysates and Immunoblotting
2.4. Confocal Microscopy
2.5. Urate Uptake Assay Using URAT1-Expressing 293A Cells
2.6. Quantification and Statistical Analysis
3. Results
3.1. URAT1-Mediated Urate Uptake in 293A Cells
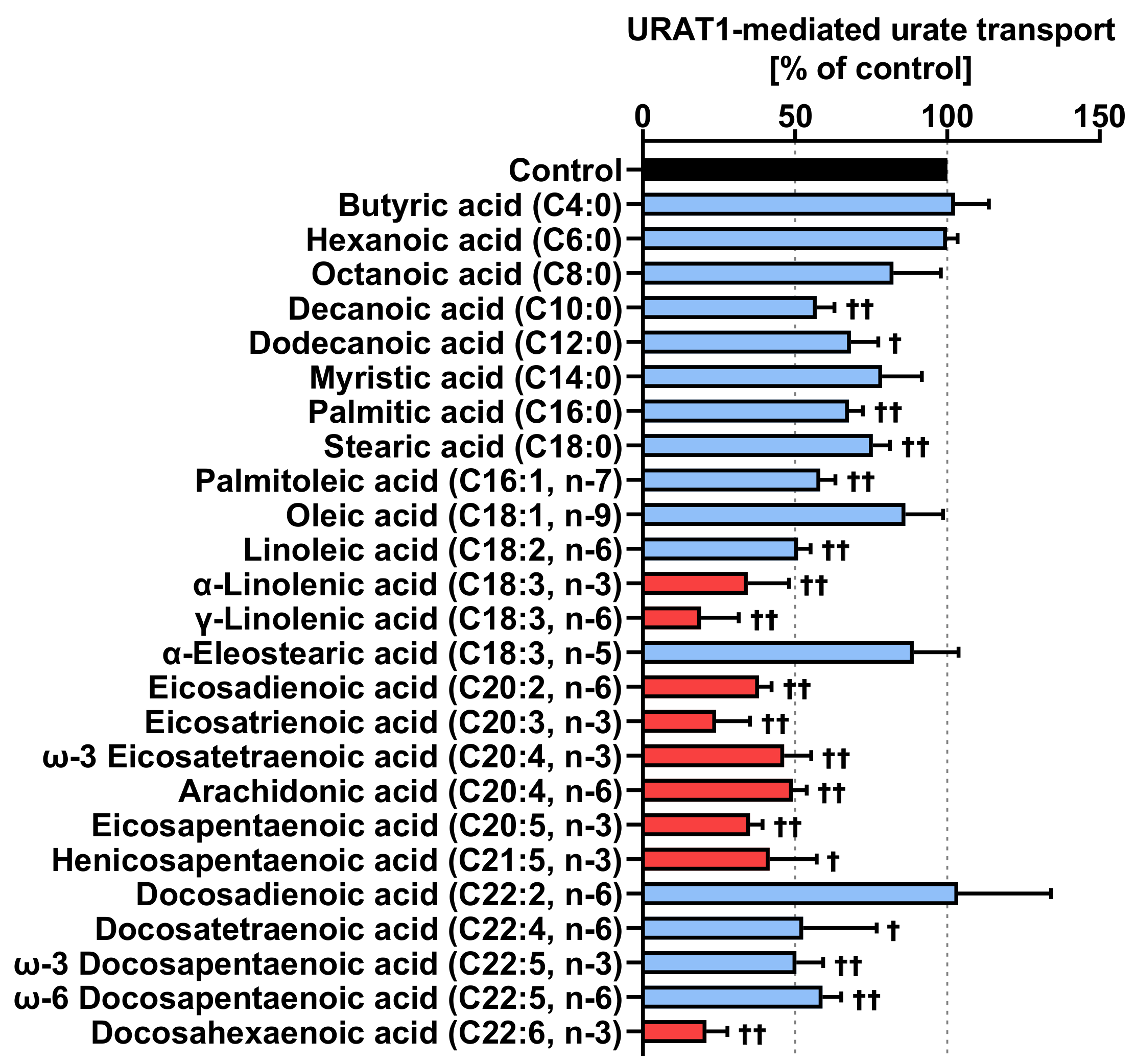
3.2. Unsaturated Fatty Acids Are Stronger Inhibitors of URAT1 Activity Than Saturated Fatty Acids
3.3. ω-3 Fatty Acids Are the Most Effective URAT1 Inhibitors
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A




References
- Calder, P.C. Functional Roles of Fatty Acids and Their Effects on Human Health. JPEN J. Parenter. Enteral. Nutr. 2015, 39, 18S–32S. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, Q.V.; Malau-Aduli, B.S.; Cavalieri, J.; Malau-Aduli, A.E.O.; Nichols, P.D. Enhancing Omega-3 Long-Chain Polyunsaturated Fatty Acid Content of Dairy-Derived Foods for Human Consumption. Nutrients 2019, 11, 743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahidi, F.; Ambigaipalan, P. Omega-3 Polyunsaturated Fatty Acids and Their Health Benefits. Annu. Rev. Food Sci. Technol. 2018, 9, 345–381. [Google Scholar] [PubMed]
- Zarate, R.; El Jaber-Vazdekis, N.; Tejera, N.; Perez, J.A.; Rodriguez, C. Significance of long chain polyunsaturated fatty acids in human health. Clin. Transl. Med. 2017, 6, 25. [Google Scholar]
- Bowen, K.J.; Harris, W.S.; Kris-Etherton, P.M. Omega-3 Fatty Acids and Cardiovascular Disease: Are There Benefits? Curr. Treat. Options Cardiovasc. Med. 2016, 18, 69. [Google Scholar]
- Dalbeth, N.; Merriman, T.R.; Stamp, L.K. Gout. Lancet 2016, 388, 2039–2052. [Google Scholar] [CrossRef]
- Dalbeth, N.; Choi, H.K.; Joosten, L.A.B.; Khanna, P.P.; Matsuo, H.; Perez-Ruiz, F.; Stamp, L.K. Gout. Nat. Rev. Dis. Primers. 2019, 5, 69. [Google Scholar] [CrossRef]
- Major, T.J.; Dalbeth, N.; Stahl, E.A.; Merriman, T.R. An update on the genetics of hyperuricaemia and gout. Nat. Rev. Rheumatol. 2018, 14, 341–353. [Google Scholar]
- Wu, X.W.; Muzny, D.M.; Lee, C.C.; Caskey, C.T. Two independent mutational events in the loss of urate oxidase during hominoid evolution. J. Mol. Evol. 1992, 34, 78–84. [Google Scholar] [CrossRef]
- Bobulescu, I.A.; Moe, O.W. Renal transport of uric acid: evolving concepts and uncertainties. Adv. Chronic Kidney Dis. 2012, 19, 358–371. [Google Scholar] [CrossRef] [Green Version]
- Hyndman, D.; Liu, S.; Miner, J.N. Urate Handling in the Human Body. Curr. Rheumatol. Rep. 2016, 18, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enomoto, A.; Kimura, H.; Chairoungdua, A.; Shigeta, Y.; Jutabha, P.; Cha, S.H.; Hosoyamada, M.; Takeda, M.; Sekine, T.; Igarashi, T.; et al. Molecular identification of a renal urate anion exchanger that regulates blood urate levels. Nature 2002, 417, 447–452. [Google Scholar] [CrossRef] [PubMed]
- Miner, J.N.; Tan, P.K.; Hyndman, D.; Liu, S.; Iverson, C.; Nanavati, P.; Hagerty, D.T.; Manhard, K.; Shen, Z.; Girardet, J.-L.; et al. Lesinurad, a novel, oral compound for gout, acts to decrease serum uric acid through inhibition of urate transporters in the kidney. Arthritis. Res. Ther. 2016, 18, 214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taniguchi, T.; Ashizawa, N.; Matsumoto, K.; Saito, R.; Motoki, K.; Sakai, M.; Chikamatsu, N.; Hagihara, C.; Hashiba, M.; Iwanaga, T. Pharmacological Evaluation of Dotinurad, a Selective Urate Reabsorption Inhibitor. J. Pharmacol. Exp. Ther. 2019, 371, 162–170. [Google Scholar] [CrossRef]
- Toyoda, Y.; Takada, T.; Saito, H.; Hirata, H.; Ota-Kontani, A.; Kobayashi, N.; Tsuchiya, Y.; Suzuki, H. Inhibitory effect of Citrus flavonoids on the in vitro transport activity of human urate transporter 1 (URAT1/SLC22A12), a renal re-absorber of urate. N.P.J. Sci. Food. 2020, 4, 3. [Google Scholar] [CrossRef] [Green Version]
- Tashiro, Y.; Sakai, R.; Hirose-Sugiura, T.; Kato, Y.; Matsuo, H.; Takada, T.; Suzuki, H.; Makino, T. Effects of Osthol Isolated from Cnidium monnieri Fruit on Urate Transporter 1. Molecules 2018, 23, 2837. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.; Fong, W.P.; Cheng, C.H. Morin (3,5,7,2’,4’-pentahydroxyflavone) exhibits potent inhibitory actions on urate transport by the human urate anion transporter (hURAT1) expressed in human embryonic kidney cells. Drug. Metab. Dispos. 2007, 35, 981–986. [Google Scholar] [CrossRef]
- Miyata, H.; Takada, T.; Toyoda, Y.; Matsuo, H.; Ichida, K.; Suzuki, H. Identification of Febuxostat as a New Strong ABCG2 Inhibitor: Potential Applications and Risks in Clinical Situations. Front. Pharmacol. 2016, 7, 518. [Google Scholar] [CrossRef] [Green Version]
- Toyoda, Y.; Sakurai, A.; Mitani, Y.; Nakashima, M.; Yoshiura, K.-I.; Nakagawa, H.; Sakai, Y.; Ota, I.; Lezhava, A.; Hayashizaki, Y.; et al. Earwax, osmidrosis, and breast cancer: why does one SNP (538G>A) in the human ABC transporter ABCC11 gene determine earwax type? FASEB J. 2009, 23, 2001–2013. [Google Scholar] [CrossRef]
- Toyoda, Y.; Mancikova, A.; Krylov, V.; Morimoto, K.; Pavelcová, K.; Bohatá, J.; Pavelka, K.; Suzuki, H.; Matsuo, H.; Takada , T.; et al. Functional Characterization of Clinically-Relevant Rare Variants in ABCG2 Identified in a Gout and Hyperuricemia Cohort. Cells 2019, 8, 363. [Google Scholar] [CrossRef] [Green Version]
- Ibarguren, M.; Lopez, D.J.; Escriba, P.V. The effect of natural and synthetic fatty acids on membrane structure, microdomain organization, cellular functions and human health. Biochim. Biophys. Acta. 2014, 1838, 1518–1528. [Google Scholar] [CrossRef] [Green Version]
- Calder, P.C. Omega-3 polyunsaturated fatty acids and inflammatory processes: nutrition or pharmacology? Br. J. Clin. Pharmacol. 2013, 75, 645–662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abhishek, A.; Valdes, A.M.; Doherty, M. Low omega-3 fatty acid levels associate with frequent gout attacks: a case control study. Ann. Rheum. Dis. 2016, 75, 784–785. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhang, Y.; Terkeltaub, R.; Chen, C.; Neogi, T. Effect of Dietary and Supplemental Omega-3 Polyunsaturated Fatty Acids on Risk of Recurrent Gout Flares. Arthritis Rheumatol. 2019, 71, 1580–1586. [Google Scholar] [CrossRef] [PubMed]
- Iverson, C.; Bacong, A.; Liu, S.; Baumgartner, S.; Lundstrom, T.; Oscarsson, J.; Miner, J.N. Omega-3-carboxylic acids provide efficacious anti-inflammatory activity in models of crystal-mediated inflammation. Sci. Rep. 2018, 8, 1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tate, G.A.; Mandell, B.F.; Karmali, R.A.; Laposata, M.; Baker, D.G.; Schumacher, H.R., Jr.; Zurier, R.B. Suppression of monosodium urate crystal-induced acute inflammation by diets enriched with gamma-linolenic acid and eicosapentaenoic acid. Arthritis Rheum. 1988, 31, 1543–1551. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Li, K.; Asimi, S.; Chen, Q.; Li, D. Effect of vitamin B-12 and n-3 polyunsaturated fatty acids on plasma homocysteine, ferritin, C-reaction protein, and other cardiovascular risk factors: a randomized controlled trial. Asia. Pac. J. Clin. Nutr. 2015, 24, 403–411. [Google Scholar]
- Stea, T.H.; Stolevik, S.B.; Berntsen, S.; Ezzathkah Bastani, N.; Paulsen, G.; Lohne Seiler, H.; Hetlelid, K.J.; Blomhoff, R.; Mansoor, M.A. Effect of Omega-3 and Vitamins E + C Supplements on the Concentration of Serum B-Vitamins and Plasma Redox Aminothiol Antioxidant Status in Elderly Men after Strength Training for Three Months. Ann. Nutr. Metab. 2016, 68, 145–155. [Google Scholar] [CrossRef]
- Vitart, V.; Rudan, I.; Hayward, C.; Gray, N.K.; Floyd, J.; Knott, S.A.; Kolcic, I.; Polasek, O.; Graessler, J.; Wilson, J.F.; et al. SLC2A9 is a newly identified urate transporter influencing serum urate concentration, urate excretion and gout. Nat. Genet. 2008, 40, 437–442. [Google Scholar] [CrossRef]
- Matsuo, H.; Chiba, T.; Nagamori, S.; Nakayama, A.; Domoto, H.; Phetdee, K.; Wiriyasermkul, P.; Kikuchi, Y.; Oda, T.; Nishiyama, J.; et al. Mutations in glucose transporter 9 gene SLC2A9 cause renal hypouricemia. Am. J. Hum. Genet. 2008, 83, 744–751. [Google Scholar] [CrossRef] [Green Version]
- Higashino, T.; Morimoto, K.; Nakaoka, H.; Toyoda, Y.; Kawamura, Y.; Shimizu, S.; Nakamura, T.; Hosomichi, K.; Nakayama, A.; Ooyama, K.; et al. Dysfunctional missense variant of OAT10/SLC22A13 decreases gout risk and serum uric acid levels. Ann Rheum Dis 2020, 79, 164–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ichida, K.; Matsuo, H.; Takada, T.; Nakayama, A.; Murakami, K.; Shimizu, T.; Yamanashi, Y.; Kasuga, H.; Nakashima, H.; Nakamura, T.; et al. Decreased extra-renal urate excretion is a common cause of hyperuricemia. Nat. Commun. 2012, 3, 764. [Google Scholar] [PubMed] [Green Version]
- Matsuo, H.; Takada, T.; Ichida, K.; Nakamura, T.; Nakayama, A.; Ikebuchi, Y.; Ito, K.; Kusanagi, Y.; Chiba, T.; Tadokoro, S.; et al. Common defects of ABCG2, a high-capacity urate exporter, cause gout: A function-based genetic analysis in a Japanese population. Sci. Transl. Med. 2009, 1, 5ra11. [Google Scholar] [CrossRef] [PubMed]
- Woodward, O.M.; Kottgen, A.; Coresh, J.; Boerwinkle, E.; Guggino, W.B.; Kottgen, M. Identification of a urate transporter, ABCG2, with a common functional polymorphism causing gout. Proc. Natl. Acad. Sci. USA 2009, 106, 10338–10342. [Google Scholar] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reagent or Resource | Source | Identifier |
|---|---|---|
| Antibodies | ||
| Rabbit polyclonal anti-EGFP | Life Technologies | Cat# A11122; RRID: AB_221569; 1:1,000 dilution 1 |
| Rabbit polyclonal anti-α-tubulin | Abcam | Cat# ab15246; RRID: AB_301787; 1:1,000 dilution 1 |
| Donkey anti-rabbit IgG-horseradish peroxidase (HRP)-conjugate | GE Healthcare | Cat# NA934V; RRID: AB_772206; 1:3,000 dilution 1 |
| Chemicals | ||
| [8-14C]-Uric acid (53 mCi/mmol) | American Radiolabeled Chemicals | Cat# ARC0513 |
| Arachidonic acid | Cayman Chemical | Cat# 90010; CAS: 506-32-1; Purity: ≥98% |
| Benzbromarone | FUJIFILM Wako Pure Chemical | Cat# 028-1585; CAS: 3562-84-3; Purity: >98% |
| Butyric acid | SIGMA-ALDRICH | Cat# B103500-5ML; CAS: 107-92-6; Purity: ≥99% |
| Decanoic acid | FUJIFILM Wako Pure Chemical | Cat# 033-01073; CAS: 334-48-5; Purity: ≥98% |
| Dimethyl sulfoxide | Nacalai Tesque | Cat# 13445-74; CAS: 67-68-5 |
| Docosadienoic acid | Cayman Chemical | Cat# 20749; CAS: 17735-98-7; Purity: ≥98% |
| Docosahexaenoic acid | Cayman Chemical | Cat# 90310; CAS: 6217-54-5; Purity: ≥98% |
| Docosatetraenoic acid | Cayman Chemical | Cat# 90300; CAS: 28874-58-0; Purity: ≥98% |
| Dodecanoic acid | SIGMA-ALDRICH | Cat# L556-25G; CAS: 143-07-7; Purity: ≥98% |
| Eicosadienoic acid | Cayman Chemical | Cat# 90330; CAS: 2091-39-6; Purity: ≥98% |
| Eicosapentaenoic acid | Cayman Chemical | Cat# 90110; CAS: 10417-94-4; Purity: ≥98% |
| Eicosatrienoic acid | Cayman Chemical | Cat# 90192; CAS: 20590-32-3; Purity: ≥98% |
| Henicosapentaenoic acid | Cayman Chemical | Cat# 10670; CAS: 24257-10-1; Purity: ≥95% |
| Hexanoic acid | FUJIFILM Wako Pure Chemical | Cat# 081-06292; CAS: 142-62-1; Purity: ≥99% |
| Linoleic acid | Cayman Chemical | Cat# 90150; CAS: 60-33-3; Purity: ≥98% |
| Myristic acid | FUJIFILM Wako Pure Chemical | Cat# 130-03432; CAS: 544-63-8; Purity: ≥98% |
| Octanoic acid | SIGMA-ALDRICH | Cat# C2875-10ML; CAS: 124-07-2; Purity: ≥99% |
| Oleic acid | Cayman Chemical | Cat# 90260; CAS: 112-80-1; Purity: ≥98% |
| Palmitic acid | Cayman Chemical | Cat# 10006627; CAS: 57-10-3; Purity: ≥98% |
| Palmitoleic acid | Cayman Chemical | Cat# 10009871; CAS: 373-49-9; Purity: ≥99% |
| Polyethelenimine “MAX” | Polysciences | Cat# 24765; CAS: 49553-93-7 |
| Stearic acid | SIGMA-ALDRICH | Cat# S4751-1G; CAS: 57-11-4; Purity: ≥98.5% |
| α-Eleostearic acid | Cayman Chemical | Cat# 10008349; CAS: 506-23-0; Purity: ≥95% |
| α-Linolenic acid | Cayman Chemical | Cat# 90210; CAS: 463-40-1; Purity: ≥98% |
| γ-Linolenic acid | Cayman Chemical | Cat# 90220; CAS: 506-26-3; Purity: ≥98% |
| ω-3 Eicosatetraenoic acid | Larodan Fine Chemicals | Cat# 10-2024; CAS: 24880-40-8; Purity: ≥98% |
| ω-3 Docosapentaenoic acid | Cayman Chemical | Cat# 90165; CAS: 24880-45-3; Purity: ≥98% |
| Critical Commercial Assays | ||
| PierceTM BCA Protein Assay Reagent A & B | Thermo Fisher Scientific | Cat# 23223, Cat# 23224 |
| PureLinkTM HiPure Plasmid Filter Midiprep Kit | Thermo Fisher Scientific | Cat# K210015 |
| Recombinant DNA | ||
| The complete URAT1 cDNA | Miyata et al., 2016 [18] | NCBI Reference Sequence: NM_144585.3 |
| Experimental Models: Cell Lines | ||
| 293A | Invitrogen | R70507 |
| Software and Algorithms | ||
| Excel 2019 | Microsoft | https://products.office.com/ja-jp/home |
| Statcel4 add-in software | OMS Publishing | http://www.oms-publ.co.jp/ |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saito, H.; Toyoda, Y.; Takada, T.; Hirata, H.; Ota-Kontani, A.; Miyata, H.; Kobayashi, N.; Tsuchiya, Y.; Suzuki, H. Omega-3 Polyunsaturated Fatty Acids Inhibit the Function of Human URAT1, a Renal Urate Re-Absorber. Nutrients 2020, 12, 1601. https://doi.org/10.3390/nu12061601
Saito H, Toyoda Y, Takada T, Hirata H, Ota-Kontani A, Miyata H, Kobayashi N, Tsuchiya Y, Suzuki H. Omega-3 Polyunsaturated Fatty Acids Inhibit the Function of Human URAT1, a Renal Urate Re-Absorber. Nutrients. 2020; 12(6):1601. https://doi.org/10.3390/nu12061601
Chicago/Turabian StyleSaito, Hiroki, Yu Toyoda, Tappei Takada, Hiroshi Hirata, Ami Ota-Kontani, Hiroshi Miyata, Naoyuki Kobayashi, Youichi Tsuchiya, and Hiroshi Suzuki. 2020. "Omega-3 Polyunsaturated Fatty Acids Inhibit the Function of Human URAT1, a Renal Urate Re-Absorber" Nutrients 12, no. 6: 1601. https://doi.org/10.3390/nu12061601
APA StyleSaito, H., Toyoda, Y., Takada, T., Hirata, H., Ota-Kontani, A., Miyata, H., Kobayashi, N., Tsuchiya, Y., & Suzuki, H. (2020). Omega-3 Polyunsaturated Fatty Acids Inhibit the Function of Human URAT1, a Renal Urate Re-Absorber. Nutrients, 12(6), 1601. https://doi.org/10.3390/nu12061601