Microbiota and Lifestyle: A Special Focus on Diet

 , , , and
, , , and 
Abstract
:1. Introduction
2. Diet
2.1. Methodology
2.1.1. Search Strategy
2.1.2. Selection Criteria
2.1.3. Data Collection Process
2.2. Results and Discussion
2.2.1. Probiotics
2.2.2. Yogurt
2.2.3. Prebiotics
2.2.4. Alcoholic Beverages
2.2.5. Refined Sugars and Sweeteners
2.2.6. Fats
3. Stress
4. Other Lifestyle Factors
4.1. Physical Activity
4.2. Drug and Air Pollutants
4.3. Tobacco Consumption
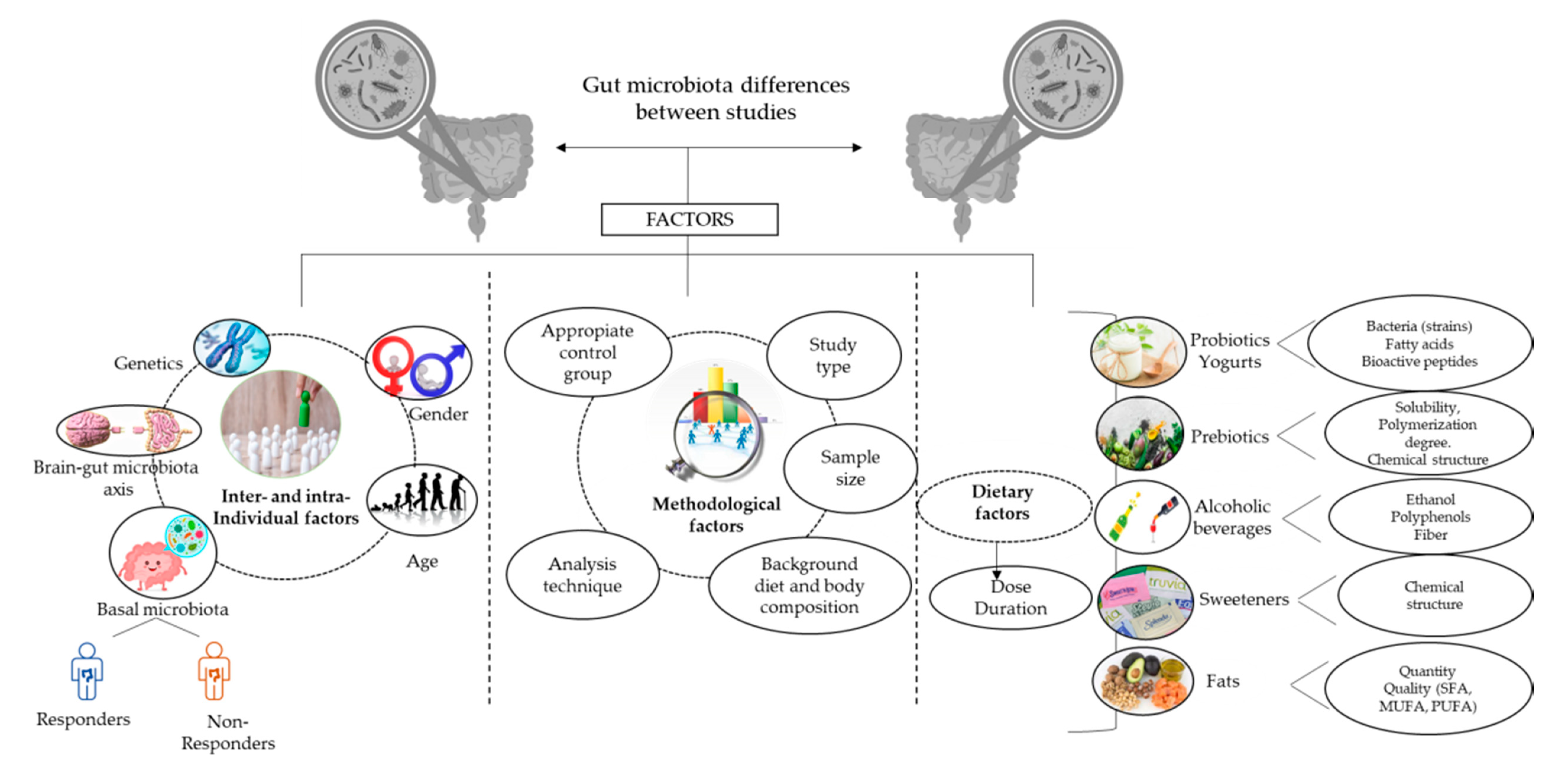
5. Key Factors Involved in the Diet–Gut Microbiota Interaction
5.1. Intra-Individual Factors
5.2. Inter-Individual Factors
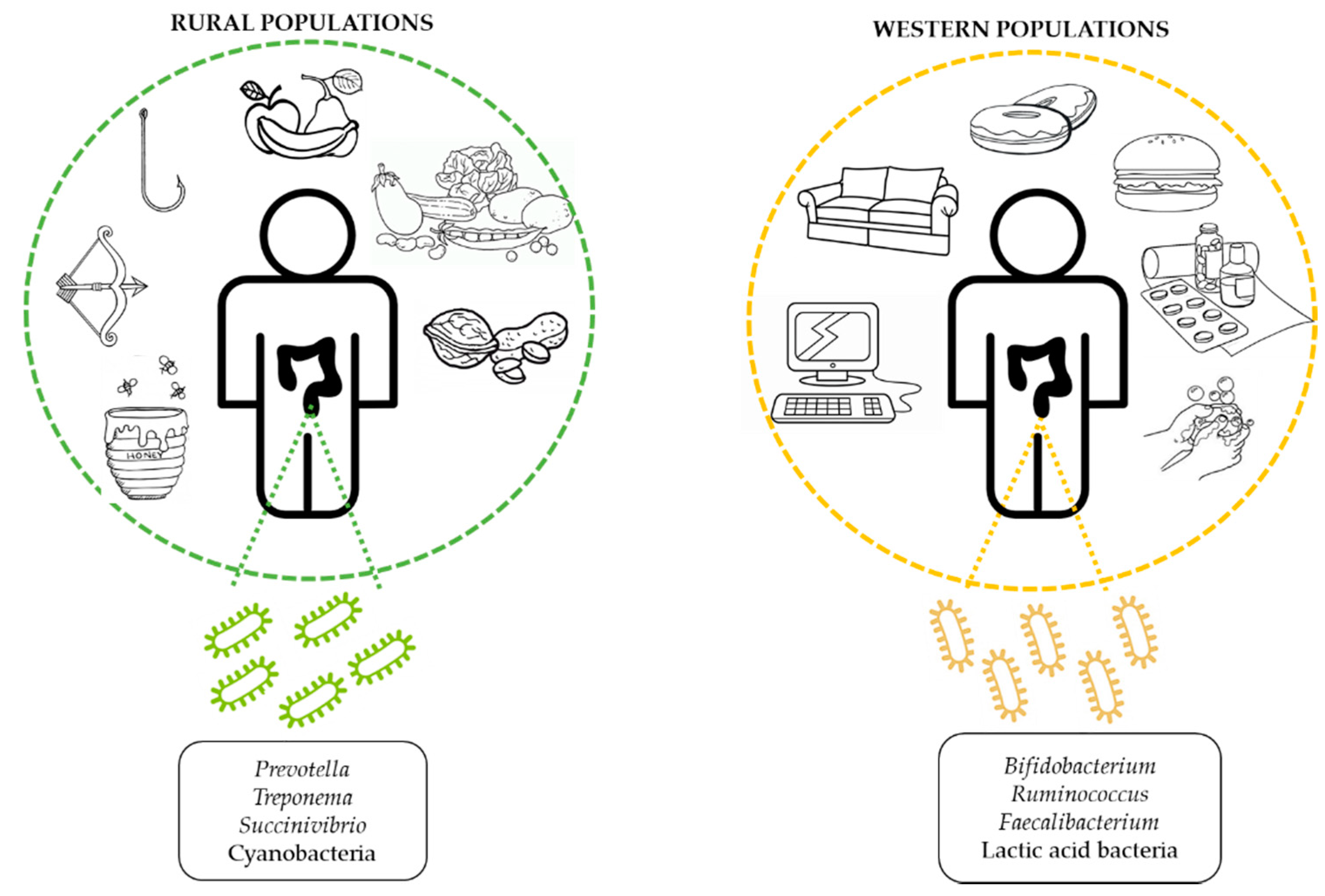
5.2.1. Geography
5.2.2. Gender
5.2.3. Age
5.3. Methodological Factors
5.4. Dietary Factors
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- WHO|Resolutions. Prevention and Control of Non-Communicable Diseases. Available online: https://www.who.int/nmh/events/un_ncd_summit2011/resolutions/en/ (accessed on 8 December 2019).
- Gómez-Gallego, C.; García-Mantrana, I.; Martínez-Costa, C.; Salminen, S.; Isolauri, E.; Collado, M.C. The Microbiota and Malnutrition: Impact of Nutritional Status During Early Life. Annu. Rev. Nutr. 2019, 39, 267–290. [Google Scholar] [CrossRef] [PubMed]
- West, C.E.; Renz, H.; Jenmalm, M.C.; Kozyrskyj, A.L.; Allen, K.J.; Vuillermin, P.; Prescott, S.L. The Gut Microbiota and Inflammatory Noncommunicable Diseases: Associations and Potentials for Gut Microbiota Therapies. J. Allergy Clin. Immunol. 2015, 135, 3–13. [Google Scholar] [CrossRef] [Green Version]
- Strachan, D.P. Hay Fever, Hygiene, and Household Size. Br. Med. J. 1989, 299, 1259–1260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lloyd-Price, J.; Abu-Ali, G.; Huttenhower, C. The Healthy Human Microbiome. Genome Med. 2016, 8, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rothschild, D.; Weissbrod, O.; Barkan, E.; Kurilshikov, A.; Korem, T.; Zeevi, D.; Costea, P.I.; Godneva, A.; Kalka, I.N.; Bar, N.; et al. Environment Dominates over Host Genetics in Shaping Human Gut Microbiota. Nature 2018, 555, 210–215. [Google Scholar] [CrossRef]
- Bäckhed, F.; Fraser, C.M.; Ringel, Y.; Sanders, M.E.; Sartor, R.B.; Sherman, P.M.; Versalovic, J.; Young, V.; Finlay, B.B. Defining a Healthy Human Gut Microbiome: Current Concepts, Future Directions, and Clinical Applications. Cell Host Microbe 2012, 12, 611–622. [Google Scholar] [CrossRef] [Green Version]
- Candela, M.; Biagi, E.; Maccaferri, S.; Turroni, S.; Brigidi, P. Intestinal Microbiota Is a Plastic Factor Responding to Environmental Changes. Trends Microbiol. 2012, 20, 385–391. [Google Scholar] [CrossRef] [PubMed]
- Levy, M.; Kolodziejczyk, A.A.; Thaiss, C.A.; Elinav, E. Dysbiosis and the Immune System. Nat. Rev. Immunol. 2017, 17, 219–232. [Google Scholar] [CrossRef]
- Tebani, A.; Bekri, S. Paving the Way to Precision Nutrition through Metabolomics. Front. Nutr. 2019, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loke, P.; Lim, Y.A.L. Helminths and the Microbiota: Parts of the Hygiene Hypothesis. Parasite Immunol. 2015, 37, 314–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sender, R.; Fuchs, S.; Milo, R. Revised Estimates for the Number of Human and Bacteria Cells in the Body. PLoS Biol. 2016, 14, e1002533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turnbaugh, P.J.; Ley, R.E.; Hamady, M.; Fraser-Liggett, C.M.; Knight, R.; Gordon, J.I. The Human Microbiome Project. Nature 2007, 449, 804–810. [Google Scholar] [CrossRef] [PubMed]
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.; Gasbarrini, A.; Mele, M. What Is the Healthy Gut Microbiota Composition? A Changing Ecosystem across Age, Environment, Diet, and Diseases. Microorganisms 2019, 7, 14. [Google Scholar] [CrossRef] [Green Version]
- Belkaid, Y.; Harrison, O.J. Homeostatic Immunity and the Microbiota. Immunity 2017, 46, 562–576. [Google Scholar] [CrossRef]
- Doré, J.; Multon, M.C.; Béhier, J.M.; Affagard, H.; Andremont, A.; Barthélémy, P.; Batitsa, R.; Bonneville, M.; Bonny, C.; Boyaval, G.; et al. The Human Gut Microbiome as Source of Innovation for Health: Which Physiological and Therapeutic Outcomes Could We Expect? Therapie 2017, 72, 21–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antonini, M.; Conte, M.L.; Sorini, C.; Falcone, M. How the Interplay between the Commensal Microbiota, Gut Barrier Integrity, and Mucosal Immunity Regulates Brain Autoimmunity. Front. Immunol. 2019, 10. [Google Scholar] [CrossRef]
- Dahl, W.J.; Stewart, M.L. Position of the Academy of Nutrition and Dietetics: Health Implications of Dietary Fiber. J. Acad. Nutr. Diet. 2015, 115, 1861–1870. [Google Scholar] [CrossRef]
- Bo, S.; Ciccone, G.; Pearce, N.; Merletti, F.; Gentile, L.; Cassader, M.; Pagano, G. Prevalence of Undiagnosed Metabolic Syndrome in a Population of Adult Asymptomatic Subjects. Diabetes Res. Clin. Pract. 2007, 75, 362–365. [Google Scholar] [CrossRef]
- Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K.S.; Manichanh, C.; Nielsen, T.; Pons, N.; Levenez, F.; Yamada, T.; et al. A Human Gut Microbial Gene Catalogue Established by Metagenomic Sequencing. Nature 2010, 464, 59–65. [Google Scholar] [CrossRef] [Green Version]
- Tap, J.; Furet, J.-P.; Bensaada, M.; Philippe, C.; Roth, H.; Rabot, S.; Lakhdari, O.; Lombard, V.; Henrissat, B.; Corthier, G.; et al. Gut Microbiota Richness Promotes Its Stability upon Increased Dietary Fibre Intake in Healthy Adults. Environ. Microbiol. 2015, 17, 4954–4964. [Google Scholar] [CrossRef]
- Lozupone, C.A.; Stombaugh, J.I.; Gordon, J.I.; Jansson, J.K.; Knight, R. Diversity, Stability and Resilience of the Human Gut Microbiota. Nature 2012, 489, 220–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reuter, G. The Lactobacillus and Bifidobacterium Microflora of the Human Intestine: Composition and Succession. Curr. Issues Intest. Microbiol. 2001, 2, 43–53. [Google Scholar] [PubMed]
- Shafquat, A.; Joice, R.; Simmons, S.L.; Huttenhower, C. Functional and Phylogenetic Assembly of Microbial Communities in the Human Microbiome. Trends Microbiol. 2014, 22, 261–266. [Google Scholar] [CrossRef] [Green Version]
- Derrien, M.; van Hylckama Vlieg, J.E.T. Fate, Activity, and Impact of Ingested Bacteria within the Human Gut Microbiota. Trends Microbiol. 2015, 23, 354–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McNulty, N.P.; Yatsunenko, T.; Hsiao, A.; Faith, J.J.; Muegge, B.D.; Goodman, A.L.; Henrissat, B.; Oozeer, R.; Cools-Portier, S.; Gobert, G.; et al. The Impact of a Consortium of Fermented Milk Strains on the Gut Microbiome of Gnotobiotic Mice and Monozygotic Twins. Sci. Transl. Med. 2011, 3, 106ra106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huttenhower, C.; Gevers, D.; Knight, R.; Abubucker, S.; Badger, J.H.; Chinwalla, A.T.; Creasy, H.H.; Earl, A.M.; Fitzgerald, M.G.; Fulton, R.S.; et al. Structure, Function and Diversity of the Healthy Human Microbiome. Nature 2012, 486, 207–214. [Google Scholar] [CrossRef] [Green Version]
- Neish, A.S. Microbes in Gastrointestinal Health and Disease. Gastroenterology 2009, 136, 65–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yatsunenko, T.; Rey, F.E.; Manary, M.J.; Trehan, I.; Dominguez-Bello, M.G.; Contreras, M.; Magris, M.; Hidalgo, G.; Baldassano, R.N.; Anokhin, A.P.; et al. Human Gut Microbiome Viewed across Age and Geography. Nature 2012, 486, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Björkman, A.; Cai, K.; Liu, G.; Wang, C.; Li, Y.; Xia, H.; Sun, L.; Kristiansen, K.; Wang, J.; et al. Impact of a 3-Months Vegetarian Diet on the Gut Microbiota and Immune Repertoire. Front. Immunol. 2018, 9, 908. [Google Scholar] [CrossRef] [PubMed]
- Mitsou, E.K.; Kakali, A.; Antonopoulou, S.; Mountzouris, K.C.; Yannakoulia, M.; Panagiotakos, D.B.; Kyriacou, A. Adherence to the Mediterranean Diet Is Associated with the Gut Microbiota Pattern and Gastrointestinal Characteristics in an Adult Population. Br. J. Nutr. 2017, 117, 1645–1655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Filippis, F.; Pellegrini, N.; Vannini, L.; Jeffery, I.B.; La Storia, A.; Laghi, L.; I Serrazanetti, D.; Di Cagno, R.; Ferrocino, I.; Lazzi, C.; et al. High-Level Adherence to a Mediterranean Diet Beneficially Impacts the Gut Microbiota and Associated Metabolome. Gut 2016, 65, 1812–1821. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Navarro, T.; Díaz, I.; Gutiérrez-Díaz, I.; Rodríguez-Carrio, J.; Suárez, A.; de los Reyes-Gavilán, C.G.; Gueimonde, M.; Salazar, N.; González, S. Exploring the Interactions between Serum Free Fatty Acids and Fecal Microbiota in Obesity through a Machine Learning Algorithm. Food Res. Int. 2019, 121, 533–541. [Google Scholar] [CrossRef]
- Myles, I.A. Fast Food Fever: Reviewing the Impacts of the Western Diet on Immunity. Nutr. J. 2014, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sofi, F.; Macchi, C.; Abbate, R.; Gensini, G.F.; Casini, A. Mediterranean Diet and Health. BioFactors 2013, 39, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Rinninella, E.; Cintoni, M.; Raoul, P.; Lopetuso, L.R.; Scaldaferri, F.; Pulcini, G.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. Food Components and Dietary Habits: Keys for a Healthy Gut Microbiota Composition. Nutrients 2019, 11, 2393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uyeno, Y.; Sekiguchi, Y.; Kamagata, Y. Impact of Consumption of Probiotic Lactobacilli-Containing Yogurt on Microbial Composition in Human Feces. Int. J. Food Microbiol. 2008, 122, 16–22. [Google Scholar] [CrossRef]
- García-Albiach, R.; Pozuelo de Felipe, M.J.; José, M.; de Felipe, P.; Angulo, S.; Morosini, M.-I.; Bravo, D.; Baquero, F.; del Campo, R. Molecular Analysis of Yogurt Containing Lactobacillus Delbrueckii Subsp. Bulgaricus and Streptococcus Thermophilus in Human Intestinal Microbiota. Am. J. Clin. Nutr. 2008, 87, 91–96. [Google Scholar] [CrossRef]
- Sierra, S.; Lara-Villoslada, F.; Sempere, L.; Olivares, M.; Boza, J.; Xaus, J. Intestinal and Immunological Effects of Daily Oral Administration of Lactobacillus Salivarius CECT5713 to Healthy Adults. Anaerobe 2010, 16, 195–200. [Google Scholar] [CrossRef]
- Larsen, C.N.; Nielsen, S.; Kaestel, P.; Brockmann, E.; Bennedsen, M.; Christensen, H.R.; Eskesen, D.C.; Jacobsen, B.L.; Michaelsen, K.F. Dose-Response Study of Probiotic Bacteria Bifidobacterium Animalis Subsp Lactis BB-12 and Lactobacillus Paracasei Subsp Paracasei CRL-341 in Healthy Young Adults. Eur. J. Clin. Nutr. 2006, 60, 1284–1293. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Wang, L.; Guo, Z.; Sun, Z.; Gesudu, Q.; Kwok, L.; Menghebilige; Zhang, H. 454 Pyrosequencing Reveals Changes in the Faecal Microbiota of Adults Consuming Lactobacillus Casei Zhang. FEMS Microbiol. Ecol. 2014, 88, 612–622. [Google Scholar] [CrossRef] [Green Version]
- Ferrario, C.; Taverniti, V.; Milani, C.; Fiore, W.; Laureati, M.; De Noni, I.; Stuknyte, M.; Chouaia, B.; Riso, P.; Guglielmetti, S. Modulation of Fecal Clostridiales Bacteria and Butyrate by Probiotic Intervention with Lactobacillus Paracasei DG Varies among Healthy Adults. J. Nutr. 2014, 144, 1787–1796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, B.R.; Shin, J.; Guevarra, R.B.; Lee, J.H.; Kim, D.W.; Seol, K.H.; Lee, J.H.; Kim, H.B.; Isaacson, R.E. Deciphering Diversity Indices for a Better Understanding of Microbial Communities. J. Microbiol. Biotechnol. 2017, 27, 2089–2093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toscano, M.; De Grandi, R.; Stronati, L.; De Vecchi, E.; Drago, L. Effect of Lactobacillus Rhamnosus HN001 and Bifidobacterium Longum BB536 on the Healthy Gut Microbiota Composition at Phyla and Species Level: A Preliminary Study. World J. Gastroenterol. 2017, 23, 2696–2704. [Google Scholar] [CrossRef] [PubMed]
- Toscano, M.; De Grandi, R.; Miniello, V.L.; Mattina, R.; Drago, L. Ability of Lactobacillus Kefiri LKF01 (DSM32079) to Colonize the Intestinal Environment and Modify the Gut Microbiota Composition of Healthy Individuals. Dig. Liver Dis. 2017, 49, 261–267. [Google Scholar] [CrossRef] [Green Version]
- Gargari, G.; Taverniti, V.; Balzaretti, S.; Ferrario, C.; Gardana, C.; Simonetti, P.; Guglielmetti, S. Consumption of a Bifidobacterium Bifidum Strain for 4 Weeks Modulates Dominant Intestinal Bacterial Taxa and Fecal Butyrate in Healthy Adults. Appl. Environ. Microbiol. 2016, 82, 5850–5859. [Google Scholar] [CrossRef] [Green Version]
- Verdenelli, M.C.; Silvi, S.; Cecchini, C.; Orpianesi, C.; Cresci, A. Influence of a Combination of Two Potential Probiotic Strains, Lactobacillus Rhamnosus IMC 501® and Lactobacillus Paracasei IMC 502® on Bowel Habits of Healthy Adults. Lett. Appl. Microbiol. 2011, 52, 596–602. [Google Scholar] [CrossRef]
- Olivares, M.; Díaz-Ropero, M.P.; Gómez, N.; Sierra, S.; Lara-Villoslada, F.; Martín, R.; Rodríguez, J.M.; Xaus, J. Dietary Deprivation of Fermented Foods Causes a Fall in Innate Immune Response. Lactic Acid Bacteria Can Counteract the Immunological Effect of This Deprivation. J. Dairy Res. 2006, 73, 492–498. [Google Scholar] [CrossRef]
- Zhang, H.; Sun, J.; Liu, X.; Hong, C.; Zhu, Y.; Liu, A.; Li, S.; Guo, H.; Ren, F. Lactobacillus Paracasei Subsp. Paracasei LC01 Positively Modulates Intestinal Microflora in Healthy Young Adults. J. Microbiol. 2013, 51, 777–782. [Google Scholar] [CrossRef]
- Savard, P.; Lamarche, B.; Paradis, M.E.; Thiboutot, H.; Laurin, É.; Roy, D. Impact of Bifidobacterium Animalis Subsp. Lactis BB-12 and, Lactobacillus Acidophilus LA-5-Containing Yoghurt, on Fecal Bacterial Counts of Healthy Adults. Int. J. Food Microbiol. 2011, 149, 50–57. [Google Scholar] [CrossRef]
- Filteau, M.; Matamoros, S.; Savard, P.; Roy, D. Molecular Monitoring of Fecal Microbiota in Healthy Adults Following Probiotic Yogurt Intake. Pharma Nutr. 2013, 1, 123–129. [Google Scholar] [CrossRef]
- Burton, K.J.; Rosikiewicz, M.; Pimentel, G.; Bütikofer, U.; von Ah, U.; Voirol, M.-J.; Croxatto, A.; Aeby, S.; Drai, J.; McTernan, P.G.; et al. Probiotic Yogurt and Acidified Milk Similarly Reduce Postprandial Inflammation and Both Alter the Gut Microbiota of Healthy, Young Men. Br. J. Nutr. 2017, 117, 1312–1322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volokh, O.; Klimenko, N.; Berezhnaya, Y.; Tyakht, A.; Nesterova, P.; Popenko, A.; Alexeev, D. Human Gut Microbiome Response Induced by Fermented Dairy Product Intake in Healthy Volunteers. Nutrients 2019, 11, 547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rochet, V.; Rigottier-Gois, L.; Sutren, M.; Krementscki, M.-N.; Andrieux, C.; Furet, J.-P.; Tailliez, P.; Levenez, F.; Mogenet, A.; Bresson, J.-L.; et al. Effects of Orally Administered Lactobacillus Casei DN-114 001 on the Composition or Activities of the Dominant Faecal Microbiota in Healthy Humans. Br. J. Nutr. 2006, 95, 421–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramnani, P.; Costabile, A.; Bustillo, A.G.R.; Gibson, G.R. A Randomised, Double- Blind, Cross-over Study Investigating the Prebiotic Effect of Agave Fructans in Healthy Human Subjects. J. Nutr. Sci. 2015, 4. [Google Scholar] [CrossRef] [Green Version]
- Holscher, H.D.; Bauer, L.L.; Gourineni, V.; Pelkman, C.L.; Fahey, G.C.; Swanson, K.S. Agave Inulin Supplementation Affects the Fecal Microbiota of Healthy Adults Participating in a Randomized, Double-Blind, Placebo-Controlled, Crossover Trial. J. Nutr. 2015, 145, 2025–2032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarke, S.T.; Green-Johnson, J.M.; Brooks, S.P.J.; Ramdath, D.D.; Bercik, P.; Avila, C.; Inglis, G.D.; Green, J.; Yanke, L.J.; Selinger, L.B.; et al. Β2-1 Fructan Supplementation Alters Host Immune Responses in a Manner Consistent with Increased Exposure to Microbial Components: Results from a Double-Blinded, Randomised, Cross-over Study in Healthy Adults. Br. J. Nutr. 2016, 115, 1748–1759. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; Li, P.; Chen, M.; Luo, Y.; Prabhakar, M.; Zheng, H.; He, Y.; Qi, Q.; Long, H.; Zhang, Y.; et al. Fructooligosaccharide (FOS) and Galactooligosaccharide (GOS) Increase Bifidobacterium but Reduce Butyrate Producing Bacteria with Adverse Glycemic Metabolism in Healthy Young Population. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef]
- Elison, E.; Vigsnaes, L.K.; Rindom Krogsgaard, L.; Rasmussen, J.; Sorensen, N.; McConnell, B.; Hennet, T.; Sommer, M.O.A.; Bytzer, P. Oral Supplementation of Healthy Adults with 2′-O-Fucosyllactose and Lacto-N-Neotetraose Is Well Tolerated and Shifts the Intestinal Microbiota. Br. J. Nutr. 2016, 116, 1356–1368. [Google Scholar] [CrossRef] [Green Version]
- Healey, G.; Murphy, R.; Butts, C.; Brough, L.; Whelan, K.; Coad, J. Habitual Dietary Fibre Intake Influences Gut Microbiota Response to an Inulin-Type Fructan Prebiotic: A Randomised, Double-Blind, Placebo-Controlled, Cross-over, Human Intervention Study. Br. J. Nutr. 2018, 119, 176–189. [Google Scholar] [CrossRef] [PubMed]
- Costabile, A.; Kolida, S.; Klinder, A.; Gietl, E.; Bäuerlein, M.; Frohberg, C.; Landschütze, V.; Gibson, G.R. A Double-Blind, Placebo-Controlled, Cross-over Study to Establish the Bifidogenic Effect of a Very-Long-Chain Inulin Extracted from Globe Artichoke (Cynara Scolymus) in Healthy Human Subjects. Br. J. Nutr. 2010, 104, 1007–1017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burns, A.M.; Solch, R.J.; Dennis-Wall, J.C.; Ukhanova, M.; Nieves, C.; Mai, V.; Christman, M.C.; Gordon, D.T.; Langkamp-Henken, B. In Healthy Adults, Resistant Maltodextrin Produces a Greater Change in Fecal Bifidobacteria Counts and Increases Stool Wet Weight: A Double-Blind, Randomized, Controlled Crossover Study. Nutr. Res. 2018, 60, 33–42. [Google Scholar] [CrossRef]
- Alfa, M.J.; Strang, D.; Tappia, P.S.; Graham, M.; Van Domselaar, G.; Forbes, J.D.; Laminman, V.; Olson, N.; DeGagne, P.; Bray, D.; et al. A Randomized Trial to Determine the Impact of a Digestion Resistant Starch Composition on the Gut Microbiome in Older and Mid-Age Adults. Clin. Nutr. 2018, 37, 797–807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Childs, C.E.; Röytiö, H.; Alhoniemi, E.; Fekete, A.A.; Forssten, S.D.; Hudjec, N.; Lim, Y.N.; Steger, C.J.; Yaqoob, P.; Tuohy, K.M.; et al. Xylo-Oligosaccharides Alone or in Synbiotic Combination with Bifidobacterium Animalis Subsp. Lactis Induce Bifidogenesis and Modulate Markers of Immune Function in Healthy Adults: A Double-Blind, Placebo-Controlled, Randomised, Factorial Cross-over Study. Br. J. Nutr. 2014, 111, 1945–1956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walton, G.E.; Lu, C.; Trogh, I.; Arnaut, F.; Gibson, G.R. A Randomised, Double-Blind, Placebo Controlled Cross-over Study to Determine the Gastrointestinal Effects of Consumption of Arabinoxylan- Oligosaccharides Enriched Bread in Healthy Volunteers. Nutr. J. 2012, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hooda, S.; Boler, B.M.V.; Serao, M.C.R.; Brulc, J.M.; Staeger, M.A.; Boileau, T.W.; Dowd, S.E.; Fahey, G.C.; Swanson, K.S. 454 Pyrosequencing Reveals a Shift in Fecal Microbiota of Healthy Adult Men Consuming Polydextrose or Soluble Corn Fiber. J. Nutr. 2012, 142, 1259–1265. [Google Scholar] [CrossRef]
- Baxter, N.T.; Schmidt, A.W.; Venkataraman, A.; Kim, K.S.; Waldron, C.; Schmidt, T.M. Dynamics of Human Gut Microbiota and Short-Chain Fatty Acids in Response to Dietary Interventions with Three Fermentable Fibers. MBio 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Lecerf, J.M.; Dépeint, F.; Clerc, E.; Dugenet, Y.; Niamba, C.N.; Rhazi, L.; Cayzeele, A.; Abdelnour, G.; Jaruga, A.; Younes, H.; et al. Xylo-Oligosaccharide (XOS) in Combination with Inulin Modulates Both the Intestinal Environment and Immune Status in Healthy Subjects, While XOS Alone Only Shows Prebiotic Properties. Br. J. Nutr. 2012, 108, 1847–1858. [Google Scholar] [CrossRef]
- Ampatzoglou, A.; Atwal, K.K.; Maidens, C.M.; Williams, C.L.; Ross, A.B.; Thielecke, F.; Jonnalagadda, S.S.; Kennedy, O.B.; Yaqoob, P. Increased Whole Grain Consumption Does Not Affect Blood Biochemistry, Body Composition, or Gut Microbiology in Healthy, Low-Habitual Whole Grain Consumers. J. Nutr. 2015, 145, 215–221. [Google Scholar] [CrossRef] [Green Version]
- Vanegas, S.M.; Meydani, M.; Barnett, J.B.; Goldin, B.; Kane, A.; Rasmussen, H.; Brown, C.; Vangay, P.; Knights, D.; Jonnalagadda, S.; et al. Substituting Whole Grains for Refined Grains in a 6-Wk Randomized Trial Has a Modest Effect on Gut Microbiota and Immune and Inflammatory Markers of Healthy Adults. Am. J. Clin. Nutr. 2017, 105, 635–650. [Google Scholar] [CrossRef] [Green Version]
- Vendrame, S.; Guglielmetti, S.; Riso, P.; Arioli, S.; Klimis-Zacas, D.; Porrini, M. Six-Week Consumption of a Wild Blueberry Powder Drink Increases Bifidobacteria in the Human Gut. J. Agric. Food Chem. 2011, 59, 12815–12820. [Google Scholar] [CrossRef]
- Guglielmetti, S.; Fracassetti, D.; Taverniti, V.; Del Bo’, C.; Vendrame, S.; Klimis-Zacas, D.; Arioli, S.; Riso, P.; Porrini, M. Differential Modulation of Human Intestinal Bifidobacterium Populations after Consumption of a Wild Blueberry (Vaccinium Angustifolium) Drink. J. Agric. Food Chem. 2013, 61, 8134–8140. [Google Scholar] [CrossRef] [PubMed]
- Wallace, A.J.; Eady, S.L.; Hunter, D.C.; Skinner, M.A.; Huffman, L.; Ansell, J.; Blatchford, P.; Wohlers, M.; Herath, T.D.; Hedderley, D.; et al. No Difference in Fecal Levels of Bacteria or Short Chain Fatty Acids in Humans, When Consuming Fruit Juice Beverages Containing Fruit Fiber, Fruit Polyphenols, and Their Combination. Nutr. Res. 2015, 35, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Klinder, A.; Shen, Q.; Heppel, S.; Lovegrove, J.A.; Rowland, I.; Tuohy, K.M. Impact of Increasing Fruit and Vegetables and Flavonoid Intake on the Human Gut Microbiota. Food Funct. 2016, 7, 1788–1796. [Google Scholar] [CrossRef] [PubMed]
- Tzounis, X.; Rodriguez-Mateos, A.; Vulevic, J.; Gibson, G.R.; Kwik-Uribe, C.; Spencer, J.P.E. Prebiotic Evaluation of Cocoa-Derived Flavanols in Healthy Humans by Using a Randomized, Controlled, Double-Blind, Crossover Intervention Study. Am. J. Clin. Nutr. 2011, 93, 62–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, X.; Long, Y.; Ji, Z.; Gao, J.; Fu, T.; Yan, M.; Zhang, L.; Su, H.; Zhang, W.; Wen, X.; et al. Green Tea Liquid Consumption Alters the Human Intestinal and Oral Microbiome. Mol. Nutr. Food Res. 2018, 62. [Google Scholar] [CrossRef] [Green Version]
- Janssens, P.L.H.R.; Penders, J.; Hursel, R.; Budding, A.E.; Savelkoul, P.H.M.; Westerterp-Plantenga, M.S. Long-Term Green Tea Supplementation Does Not Change the Human Gut Microbiota. PLoS ONE 2016, 11. [Google Scholar] [CrossRef]
- Cuervo, A.; Reyes-Gavilán, C.G.; Ruas-Madiedo, P.; Lopez, P.; Suarez, A.; Gueimonde, M.; González, S. Red Wine Consumption Is Associated with Fecal Microbiota and Malondialdehyde in a Human Population. J. Am. Coll. Nutr. 2015, 34, 135–141. [Google Scholar] [CrossRef] [Green Version]
- Stadlbauer, V.; Horvath, A.; Komarova, I.; Schmerboeck, B.; Feldbacher, N.; Wurm, S.; Klymiuk, I.; Durdevic, M.; Rainer, F.; Blesl, A.; et al. A Single Alcohol Binge Impacts on Neutrophil Function without Changes in Gut Barrier Function and Gut Microbiome Composition in Healthy Volunteers. PLoS ONE 2019, 14. [Google Scholar] [CrossRef] [Green Version]
- Queipo-Ortuño, M.I.; Boto-Ordóñez, M.; Murri, M.; Gomez-Zumaquero, J.M.; Clemente-Postigo, M.; Estruch, R.; Cardona Diaz, F.; Andrés-Lacueva, C.; Tinahones, F.J. Influence of Red Wine Polyphenols and Ethanol on the Gut Microbiota Ecology and Biochemical Biomarkers. Am. J. Clin. Nutr. 2012, 95, 1323–1334. [Google Scholar] [CrossRef] [PubMed]
- Barroso, E.; Muñoz-González, I.; Jiménez, E.; Bartolomé, B.; Moreno-Arribas, M.V.; Peláez, C.; del Carmen Martínez-Cuesta, M.; Requena, T. Phylogenetic Profile of Gut Microbiota in Healthy Adults after Moderate Intake of Red Wine. Mol. Nutr. Food Res. 2017, 61. [Google Scholar] [CrossRef]
- Hernández-Quiroz, F.; Nirmalkar, K.; Villalobos-Flores, L.E.; Murugesan, S.; Cruz-Narváez, Y.; Rico-Arzate, E.; Hoyo-Vadillo, C.; Chavez-Carbajal, A.; Pizano-Zárate, M.L.; García-Mena, J. Influence of Moderate Beer Consumption on Human Gut Microbiota and Its Impact on Fasting Glucose and β-Cell Function. Alcohol 2020, 85, 77–94. [Google Scholar] [CrossRef] [PubMed]
- Gostner, A.; Blaut, M.; Schäffer, V.; Kozianowski, G.; Theis, S.; Klingeberg, M.; Dombrowski, Y.; Martin, D.; Ehrhardt, S.; Taras, D.; et al. Effect of Isomalt Consumption on Faecal Microflora and Colonic Metabolism in Healthy Volunteers. Br. J. Nutr. 2006, 95, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Finney, M.; Smullen, J.; Foster, H.A.; Brokx, S.; Storey, D.M. Effects of Low Doses of Lactitol on Faecal Microflora, PH, Short Chain Fatty Acids and Gastrointestinal Symptomology. Eur. J. Nutr. 2007, 46, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Frankenfeld, C.L.; Sikaroodi, M.; Lamb, E.; Shoemaker, S.; Gillevet, P.M. High-Intensity Sweetener Consumption and Gut Microbiome Content and Predicted Gene Function in a Cross-Sectional Study of Adults in the United States. Ann. Epidemiol. 2015, 25, 736–742.e4. [Google Scholar] [CrossRef]
- Suez, J.; Korem, T.; Zeevi, D.; Zilberman-Schapira, G.; Thaiss, C.A.; Maza, O.; Israeli, D.; Zmora, N.; Gilad, S.; Weinberger, A.; et al. Artificial Sweeteners Induce Glucose Intolerance by Altering the Gut Microbiota. Nature 2014, 514, 181–186. [Google Scholar] [CrossRef]
- Wan, Y.; Wang, F.; Yuan, J.; Li, J.; Jiang, D.; Zhang, J.; Li, H.; Wang, R.; Tang, J.; Huang, T.; et al. Effects of Dietary Fat on Gut Microbiota and Faecal Metabolites, and Their Relationship with Cardiometabolic Risk Factors: A 6-Month Randomised Controlled-Feeding Trial. Gut 2019, 68, 1417–1429. [Google Scholar] [CrossRef] [Green Version]
- Lang, J.M.; Pan, C.; Cantor, R.M.; Tang, W.H.W.; Garcia-Garcia, J.C.; Kurtz, I.; Hazen, S.L.; Bergeron, N.; Krauss, R.M.; Lusis, A.J. Impact of Individual Traits, Saturated Fat, and Protein Source on the Gut Microbiome. MBio 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Watson, H.; Mitra, S.; Croden, F.C.; Taylor, M.; Wood, H.M.; Perry, S.L.; Spencer, J.A.; Quirke, P.; Toogood, G.J.; Lawton, C.L.; et al. A Randomised Trial of the Effect of Omega-3 Polyunsaturated Fatty Acid Supplements on the Human Intestinal Microbiota. Gut 2018, 67, 1974–1983. [Google Scholar] [CrossRef]
- Ott, B.; Skurk, T.; Lagkouvardos, L.; Fischer, S.; Büttner, J.; Lichtenegger, M.; Clavel, T.; Lechner, A.; Rychlik, M.; Haller, D.; et al. Short-Term Overfeeding with Dairy Cream Does Not Modify Gut Permeability, the Fecal Microbiota, or Glucose Metabolism in Young Healthy Men. J. Nutr. 2018, 148, 77–85. [Google Scholar] [CrossRef] [Green Version]
- Redondo, N.; García-González, N.; Diaz-Prieto, L.E.; Olmedilla-Alonso, B.; Martín-Diana, A.B.; Asensio-Vegas, C.; Nova, E. Effects of Ewe’s Milk Yogurt (Whole and Semi-Skimmed) and Cow’s Milk Yogurt on Inflammation Markers and Gut Microbiota of Subjects with Borderline-High Plasma Cholesterol Levels: A Crossover Study. Eur. J. Nutr. 2019, 58, 1113–1124. [Google Scholar] [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert Consensus Document: The International Scientific Association for Probiotics and Prebiotics Consensus Statement on the Scope and Appropriate Use of the Term Probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mercasa. Tendencias en los Mercados Mundiales de Leche y Productos Lácteos. Distribución y Consumo. Available online: https://www.mercasa.es/media/publicaciones/224/1450218713_Tendencia_mercados_mundiales_leche.pdf (accessed on 19 November 2019).
- Tannock, G.W.; Munro, K.; Harmsen, H.J.; Welling, G.W.; Smart, J.; Gopal, P.K. Analysis of the Fecal Microflora of Human Subjects Consuming a Probiotic Product Containing Lactobacillus Rhamnosus DR20. Appl. Environ. Microbiol. 2000, 66, 2578–2588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plaza-Díaz, J.; Ruiz-Ojeda, F.; Gil-Campos, M.; Gil, A. Immune-Mediated Mechanisms of Action of Probiotics and Synbiotics in Treating Pediatric Intestinal Diseases. Nutrients 2018, 10, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacDonald, T.T.; Bell, I. Probiotics and the Immune Response to Vaccines. Proc. Nutr. Soc. 2010, 69, 442–446. [Google Scholar] [CrossRef] [Green Version]
- Moens, F.; Verce, M.; De Vuyst, L. Lactate- and Acetate-Based Cross-Feeding Interactions between Selected Strains of Lactobacilli, Bifidobacteria and Colon Bacteria in the Presence of Inulin-Type Fructans. Int. J. Food Microbiol. 2017, 241, 225–236. [Google Scholar] [CrossRef]
- Turroni, F.; Bottacini, F.; Foroni, E.; Mulder, I.; Kim, J.-H.; Zomer, A.; Sánchez, B.; Bidossi, A.; Ferrarini, A.; Giubellini, V.; et al. Genome Analysis of Bifidobacterium Bifidum PRL2010 Reveals Metabolic Pathways for Host-Derived Glycan Foraging. Proc. Natl. Acad. Sci. USA 2010, 107, 19514–19519. [Google Scholar] [CrossRef] [Green Version]
- Le Chatelier, E.; Nielsen, T.; Qin, J.; Prifti, E.; Hildebrand, F.; Falony, G.; Almeida, M.; Arumugam, M.; Batto, J.-M.; Kennedy, S.; et al. Richness of Human Gut Microbiome Correlates with Metabolic Markers. Nature 2013, 500, 541–546. [Google Scholar] [CrossRef]
- Kristensen, N.B.; Bryrup, T.; Allin, K.H.; Nielsen, T.; Hansen, T.H.; Pedersen, O. Alterations in Fecal Microbiota Composition by Probiotic Supplementation in Healthy Adults: A Systematic Review of Randomized Controlled Trials. Genome Med. 2016, 8. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.W.; Suda, W.; Kim, S.; Oshima, K.; Fukuda, S.; Ohno, H.; Morita, H.; Hattori, M. Robustness of Gut Microbiota of Healthy Adults in Response to Probiotic Intervention Revealed by High-Throughput Pyrosequencing. DNA Res. 2013, 20, 241–253. [Google Scholar] [CrossRef]
- Wang, H.; Livingston, K.A.; Fox, C.S.; Meigs, J.B.; Jacques, P.F. Yogurt Consumption Is Associated with Better Diet Quality and Metabolic Profile in American Men and Women. Nutr. Res. 2013, 33, 18–26. [Google Scholar] [CrossRef] [Green Version]
- Adolfsson, O.; Meydani, S.N.; Russell, R.M. Yogurt and Gut Function. Am. J. Clin. Nutr. 2004, 80, 245–256. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, M.A.; Panahi, S.; Daniel, N.; Tremblay, A.; Marette, A. Yogurt and Cardiometabolic Diseases: A Critical Review of Potential Mechanisms. Adv. Nutr. 2017, 8, 812–829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Meijl, L.E.C.; Vrolix, R.; Mensink, R.P. Dairy Product Consumption and the Metabolic Syndrome. Nutr. Res. Rev. 2008, 21, 148–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Augustin, O.; Rivero-Gutiérrez, B.; Mascaraque, C.; Sánchez de Medina, F. Food Derived Bioactive Peptides and Intestinal Barrier Function. Int. J. Mol. Sci. 2014, 15, 22857–22873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elli, M.; Callegari, M.L.; Ferrari, S.; Bessi, E.; Cattivelli, D.; Soldi, S.; Morelli, L.; Goupil Feuillerat, N.; Antoine, J.-M. Survival of Yogurt Bacteria in the Human Gut. Appl. Environ. Microbiol. 2006, 72, 5113–5117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mater, D.D.G.; Bretigny, L.; Firmesse, O.; Flores, M.-J.; Mogenet, A.; Bresson, J.-L.; Corthier, G. Streptococcus Thermophilus and Lactobacillus Delbrueckii Subsp. Bulgaricus Survive Gastrointestinal Transit of Healthy Volunteers Consuming Yogurt. FEMS Microbiol. Lett. 2005, 250, 185–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Del Campo, R.; Bravo, D.; Cantón, R.; Ruiz-Garbajosa, P.; García-Albiach, R.; Montesi-Libois, A.; Yuste, F.-J.; Abraira, V.; Baquero, F. Scarce Evidence of Yogurt Lactic Acid Bacteria in Human Feces after Daily Yogurt Consumption by Healthy Volunteers. Appl. Environ. Microbiol. 2005, 71, 547–549. [Google Scholar] [CrossRef] [Green Version]
- Morelli, L. Yogurt, Living Cultures, and Gut Health. Am. J. Clin. Nutr. 2014, 99 (Suppl. 5), 1248S–1250S. [Google Scholar] [CrossRef] [Green Version]
- Cândido, F.G.; Valente, F.X.; Grześkowiak, Ł.M.; Moreira, A.P.B.; Rocha, D.M.U.P.; Alfenas, R.C.G. Impact of Dietary Fat on Gut Microbiota and Low-Grade Systemic Inflammation: Mechanisms and Clinical Implications on Obesity. Int. J. Food Sci. Nutr. 2018, 69, 125–143. [Google Scholar] [CrossRef]
- Olmedilla-Alonso, B.; Nova-Rebato, E.; García-González, N.; Martín-Diana, A.-B.; Fontecha, J.; Delgado, D.; Gredilla, A.-E.; Bueno, F.; Asensio-Vegas, C. Effect of Ewe’s (Semi-Skimmed and Whole) and Cow’s Milk Yogurt Consumption on the Lipid Profile of Control Subjects: A Crossover Study. Food Nutr. Res. 2017, 61, 1391669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aoyama, T.; Nosaka, N.; Kasai, M. Research on the Nutritional Characteristics of Medium-Chain Fatty Acids. J. Med. Investig. 2007, 54, 385–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, S.; Wang, Y.; Jacoby, J.J.; Jiang, Y.; Zhang, Y.; Yu, L.L. Effects of Medium- and Long-Chain Triacylglycerols on Lipid Metabolism and Gut Microbiota Composition in C57BL/6J Mice. J. Agric. Food Chem. 2017, 65, 6599–6607. [Google Scholar] [CrossRef]
- Chaplin, A.; Parra, P.; Serra, F.; Palou, A. Conjugated Linoleic Acid Supplementation under a High-Fat Diet Modulates Stomach Protein Expression and Intestinal Microbiota in Adult Mice. PLoS ONE 2015, 10, e0125091. [Google Scholar] [CrossRef] [Green Version]
- Allaband, C.; McDonald, D.; Vázquez-Baeza, Y.; Minich, J.J.; Tripathi, A.; Brenner, D.A.; Loomba, R.; Smarr, L.; Sandborn, W.J.; Schnabl, B.; et al. Microbiome 101: Studying, Analyzing, and Interpreting Gut Microbiome Data for Clinicians. Clin. Gastroenterol. Hepatol. 2019, 17, 218–230. [Google Scholar] [CrossRef] [PubMed]
- Gibson, G.R.; Probert, H.M.; Van Loo, J.; Rastall, R.A.; Roberfroid, M.B. Dietary Modulation of the Human Colonic Microbiota: Updating the Concept of Prebiotics. Nutr. Res. Rev. 2004, 17, 259–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert Consensus Document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) Consensus Statement on the Definition and Scope of Prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberfroid, M.; Gibson, G.R.; Hoyles, L.; McCartney, A.L.; Rastall, R.; Rowland, I.; Wolvers, D.; Watzl, B.; Szajewska, H.; Stahl, B.; et al. Prebiotic Effects: Metabolic and Health Benefits. Br. J. Nutr. 2010, 104 (Suppl. 2). [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schell, M.A.; Karmirantzou, M.; Snel, B.; Vilanova, D.; Berger, B.; Pessi, G.; Zwahlen, M.C.; Desiere, F.; Bork, P.; Delley, M.; et al. The Genome Sequence of Bifidobacterium Longum Reflects Its Adaptation to the Human Gastrointestinal Tract. Proc. Natl. Acad. Sci. USA 2002, 99, 14422–14427. [Google Scholar] [CrossRef] [Green Version]
- So, D.; Whelan, K.; Rossi, M.; Morrison, M.; Holtmann, G.; Kelly, J.T.; Shanahan, E.R.; Staudacher, H.M.; Campbell, K.L. Dietary Fiber Intervention on Gut Microbiota Composition in Healthy Adults: A Systematic Review and Meta-Analysis. Am. J. Clin. Nutr. 2018, 107, 965–983. [Google Scholar] [CrossRef] [Green Version]
- Belenguer, A.; Duncan, S.H.; Calder, A.G.; Holtrop, G.; Louis, P.; Lobley, G.E.; Flint, H.J. Two Routes of Metabolic Cross-Feeding between Bifidobacterium Adolescentis and Butyrate-Producing Anaerobes from the Human Gut. Appl. Environ. Microbiol. 2006, 72, 3593–3599. [Google Scholar] [CrossRef] [Green Version]
- Garrido, D.; Dallas, D.C.; Mills, D.A. Consumption of Human Milk Glycoconjugates by Infant-Associated Bifidobacteria: Mechanisms and Implications. Microbiology 2013, 159, 649–664. [Google Scholar] [CrossRef]
- Timmis, K.; Jebok, F. Microbiome Yarns: Human Milk Oligosaccharides, Bifidobacterium and Immunopowergames1,2,3,4. Microb. Biotechnol. 2018, 11, 437–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mora-Cura, Y.N.; Meléndez-Rentería, N.P.; Delgado-García, M.; Contreras-Esquivel, J.C.; Morlett-Chávez, J.A.; Aguilar, C.N.; Rodríguez-Herrera, R. Fermentation of Dietetic Fiber from Green Bean and Prickly Pear Shell by Pure and Mixture Culture of Lactobacillus Acidophilus LA-5 and Bifidobacterium Bifidum 450B. Curr. Microbiol. 2017, 74, 691–701. [Google Scholar] [CrossRef] [PubMed]
- Rowland, I.; Gibson, G.; Heinken, A.; Scott, K.; Swann, J.; Thiele, I.; Tuohy, K. Gut Microbiota Functions: Metabolism of Nutrients and Other Food Components. Eur. J. Nutr. 2018, 57, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Scanlan, P.D.; Shanahan, F.; Marchesi, J.R. Culture-Independent Analysis of Desulfovibrios in the Human Distal Colon of Healthy, Colorectal Cancer and Polypectomized Individuals. FEMS Microbiol. Ecol. 2009, 69, 213–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardona, F.; Andrés-Lacueva, C.; Tulipani, S.; Tinahones, F.J.; Queipo-Ortuño, M.I. Benefits of Polyphenols on Gut Microbiota and Implications in Human Health. J. Nutr. Biochem. 2013, 24, 1415–1422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dueñas, M.; Muñoz-González, I.; Cueva, C.; Jiménez-Girón, A.; Sánchez-Patán, F.; Santos-Buelga, C.; Moreno-Arribas, M.V.; Bartolomé, B. A Survey of Modulation of Gut Microbiota by Dietary Polyphenols. Biomed. Res. Int. 2015, 2015. [Google Scholar] [CrossRef] [PubMed]
- Marín, L.; Miguélez, E.M.; Villar, C.J.; Lombó, F. Bioavailability of Dietary Polyphenols and Gut Microbiota Metabolism: Antimicrobial Properties. Biomed Res. Int. 2015, 2015, 905215. [Google Scholar] [CrossRef] [Green Version]
- Trošt, K.; Ulaszewska, M.M.; Stanstrup, J.; Albanese, D.; De Filippo, C.; Tuohy, K.M.; Natella, F.; Scaccini, C.; Mattivi, F. Host: Microbiome Co-Metabolic Processing of Dietary Polyphenols—An Acute, Single Blinded, Cross-over Study with Different Doses of Apple Polyphenols in Healthy Subjects. Food Res. Int. 2018, 112, 108–128. [Google Scholar] [CrossRef] [Green Version]
- Sæther, S.M.M.; Knapstad, M.; Askeland, K.G.; Skogen, J.C. Alcohol Consumption, Life Satisfaction and Mental Health among Norwegian College and University Students. Addict. Behav. Rep. 2019, 10. [Google Scholar] [CrossRef]
- Erol, A.; Karpyak, V.M. Sex and Gender-Related Differences in Alcohol Use and Its Consequences: Contemporary Knowledge and Future Research Considerations. Drug Alcohol Depend. 2015, 156, 1–13. [Google Scholar] [CrossRef]
- Mutlu, E.A.; Gillevet, P.M.; Rangwala, H.; Sikaroodi, M.; Naqvi, A.; Engen, P.A.; Kwasny, M.; Lau, C.K.; Keshavarzian, A. Colonic Microbiome Is Altered in Alcoholism. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 302. [Google Scholar] [CrossRef]
- Tsuruya, A.; Kuwahara, A.; Saito, Y.; Yamaguchi, H.; Tsubo, T.; Suga, S.; Inai, M.; Aoki, Y.; Takahashi, S.; Tsutsumi, E.; et al. Ecophysiological Consequences of Alcoholism on Human Gut Microbiota: Implications for Ethanol-Related Pathogenesis of Colon Cancer. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [Green Version]
- Leclercq, S.; Matamoros, S.; Cani, P.D.; Neyrinck, A.M.; Jamar, F.; Stärkel, P.; Windey, K.; Tremaroli, V.; Bäckhed, F.; Verbeke, K.; et al. Intestinal Permeability, Gut-Bacterial Dysbiosis, and Behavioral Markers of Alcohol-Dependence Severity. Proc. Natl. Acad. Sci. USA 2014, 111, E4485–E4493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiva-Blanch, G.; Badimon, L. Benefits and Risks of Moderate Alcohol Consumption on Cardiovascular Disease: Current Findings and Controversies. Nutrients 2019, 12, 108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Padilla, H.; Gaziano, J.M.; Djoussé, L. Alcohol Consumption and Risk of Heart Failure: A Meta-Analysis. Phys. Sportsmed. 2010, 38, 84–89. [Google Scholar] [CrossRef]
- Polsky, S.; Akturk, H.K. Alcohol Consumption, Diabetes Risk, and Cardiovascular Disease Within Diabetes. Curr. Diab. Rep. 2017, 17. [Google Scholar] [CrossRef] [PubMed]
- Goñi, I.; Díaz-Rubio, M.E.; Saura-Calixto, F. Dietary Fiber in Beer: Content, Composition, Colonic Fermentability, and Contribution to the Diet. In Beer in Health and Disease Prevention; Elsevier: Amsterdam, The Netherlands, 2008; pp. 299–307. [Google Scholar] [CrossRef]
- Quesada-Molina, M.; Muñoz-Garach, A.; Tinahones, F.J.; Moreno-Indias, I. A New Perspective on the Health Benefits of Moderate Beer Consumption: Involvement of the Gut Microbiota. Metabolites 2019, 9, 272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naumovski, N.; Panagiotakos, D.B.; D’Cunha, N.M. Untangling the Two-Way Relationship between Red Wine Polyphenols and Gut Microbiota. Gastroenterology 2020, 158, 48–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radovanović, A.N.; Jovančićević, B.S.; Radovanović, B.C.; Mihajilov-Krstev, T.; Zvezdanović, J.B. Antioxidant and Antimicrobial Potentials of Serbian Red Wines Produced from International Vitis Vinifera Grape Varieties. J. Sci. Food Agric. 2012, 92, 2154–2161. [Google Scholar] [CrossRef]
- Moreno Indias, I. Beneficios de Los Polifenoles Contenidos En La Cerveza Sobre La Microbiota Intestinal. Nutr. Hosp. 2017, 34, 41–44. [Google Scholar] [CrossRef] [PubMed]
- Elamin, E.E.; Masclee, A.A.; Dekker, J.; Jonkers, D.M. Ethanol Metabolism and Its Effects on the Intestinal Epithelial Barrier. Nutr. Rev. 2013, 71, 483–499. [Google Scholar] [CrossRef] [PubMed]
- Davis, S.C.; Yadav, J.S.; Barrow, S.D.; Robertson, B.K. Gut Microbiome Diversity Influenced More by the Westernized Dietary Regime than the Body Mass Index as Assessed Using Effect Size Statistic. Microbiologyopen 2017, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Musso, G.; Gambino, R.; Cassader, M. Interactions Between Gut Microbiota and Host Metabolism Predisposing to Obesity and Diabetes. Annu. Rev. Med. 2011, 62, 361–380. [Google Scholar] [CrossRef] [PubMed]
- Aron-Wisnewsky, J.; Prifti, E.; Belda, E.; Ichou, F.; Kayser, B.D.; Dao, M.C.; Verger, E.O.; Hedjazi, L.; Bouillot, J.L.; Chevallier, J.M.; et al. Major Microbiota Dysbiosis in Severe Obesity: Fate after Bariatric Surgery. Gut 2019, 68, 70–82. [Google Scholar] [CrossRef]
- Forslund, K.; Hildebrand, F.; Nielsen, T.; Falony, G.; Le Chatelier, E.; Sunagawa, S.; Prifti, E.; Vieira-Silva, S.; Gudmundsdottir, V.; Krogh Pedersen, H.; et al. Disentangling Type 2 Diabetes and Metformin Treatment Signatures in the Human Gut Microbiota. Nature 2015, 528, 262–266. [Google Scholar] [CrossRef]
- Payne, A.N.; Chassard, C.; Lacroix, C. Gut Microbial Adaptation to Dietary Consumption of Fructose, Artificial Sweeteners and Sugar Alcohols: Implications for Host-Microbe Interactions Contributing to Obesity. Obes. Rev. 2012, 13, 799–809. [Google Scholar] [CrossRef]
- Lambertz, J.; Weiskirchen, S.; Landert, S.; Weiskirchen, R. Fructose: A Dietary Sugar in Crosstalk with Microbiota Contributing to the Development and Progression of Non-Alcoholic Liver Disease. Front. Immunol. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Volynets, V.; Louis, S.; Pretz, D.; Lang, L.; Ostaff, M.J.; Wehkamp, J.; Bischoff, S.C. Intestinal Barrier Function and the Gut Microbiome Are Differentially Affected in Mice Fed a Western-Style Diet or Drinking Water Supplemented with Fructose. J. Nutr. 2017, 147, 770–780. [Google Scholar] [CrossRef] [Green Version]
- Ferrere, G.; Leroux, A.; Wrzosek, L.; Puchois, V.; Gaudin, F.; Ciocan, D.; Renoud, M.L.; Naveau, S.; Perlemuter, G.; Cassard, A.M. Activation of Kupffer Cells Is Associated with a Specific Dysbiosis Induced by Fructose or High Fat Diet in Mice. PLoS ONE 2016, 11. [Google Scholar] [CrossRef] [Green Version]
- Mary, F. Diet Soft Drinks. Available online: https://static.diabetesselfmanagement.com/pdfs/DSM0310_012.pdf (accessed on 19 November 2019).
- Mooradian, A.D.; Smith, M.; Tokuda, M. The Role of Artificial and Natural Sweeteners in Reducing the Consumption of Table Sugar: A Narrative Review. Clin. Nutr. ESPEN 2017, 18, 1–8. [Google Scholar] [CrossRef]
- Ruiz-Ojeda, F.J.; Plaza-Díaz, J.; Sáez-Lara, M.J.; Gil, A. Effects of Sweeteners on the Gut Microbiota: A Review of Experimental Studies and Clinical Trials. In Advances in Nutrition; Oxford University Press: Oxford, UK, 2019; Volume 10, pp. 31–48. [Google Scholar] [CrossRef] [Green Version]
- Turnbaugh, P.J.; Ridaura, V.K.; Faith, J.J.; Rey, F.E.; Knight, R.; Gordon, J.I. The Effect of Diet on the Human Gut Microbiome: A Metagenomic Analysis in Humanized Gnotobiotic Mice. Sci. Transl. Med. 2009, 1, ra14–ra16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, X.; Li, J.; Tang, R.; Zhang, G.; Zeng, H.; Wood, R.J.; Liu, Z. High Fat Diet Alters Gut Microbiota and the Expression of Paneth Cell-Antimicrobial Peptides Preceding Changes of Circulating Inflammatory Cytokines. Mediators Inflamm. 2017, 2017. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Kim, S.E.; Kim, A.R.; Kang, S.; Park, M.Y.; Sung, M.K. Dietary Fat Intake and Age Modulate the Composition of the Gut Microbiota and Colonic Inflammation in C57BL/6J Mice. BMC Microbiol. 2019, 19. [Google Scholar] [CrossRef] [PubMed]
- Araújo, J.R.; Tomas, J.; Brenner, C.; Sansonetti, P.J. Impact of High-Fat Diet on the Intestinal Microbiota and Small Intestinal Physiology before and after the Onset of Obesity. Biochimie 2017, 141, 97–106. [Google Scholar] [CrossRef]
- Szajewska, H.; Szajewski, T. Saturated Fat Controversy: Importance of Systematic Reviews and Meta-Analyses. Crit. Rev. Food Sci. Nutr. 2016, 56, 1947–1951. [Google Scholar] [CrossRef] [PubMed]
- Patterson, E.; Wall, R.; Fitzgerald, G.F.; Ross, R.P.; Stanton, C. Health Implications of High Dietary Omega-6 Polyunsaturated Fatty Acids. J. Nutr. Metab. 2012, 2012, 1–16. [Google Scholar] [CrossRef]
- Patterson, E.; O’ Doherty, R.M.; Murphy, E.F.; Wall, R.; O’ Sullivan, O.; Nilaweera, K.; Fitzgerald, G.F.; Cotter, P.D.; Ross, R.P.; Stanton, C. Impact of Dietary Fatty Acids on Metabolic Activity and Host Intestinal Microbiota Composition in C57BL/6J Mice. Br. J. Nutr. 2014, 111, 1905–1917. [Google Scholar] [CrossRef] [Green Version]
- Gabert, L.; Vors, C.; Louche-Pélissier, C.; Sauvinet, V.; Lambert-Porcheron, S.; Drai, J.; Laville, M.; Désage, M.; Michalski, M.-C. 13C Tracer Recovery in Human Stools after Digestion of a Fat-Rich Meal Labelled with [1,1,1-13C3]Tripalmitin and [1,1,1-13C3]Triolein. Rapid Commun. Mass Spectrom. 2011, 25, 2697–2703. [Google Scholar] [CrossRef]
- Yoon, B.K.; Jackman, J.A.; Valle-González, E.R.; Cho, N.J. Antibacterial Free Fatty Acids and Monoglycerides: Biological Activities, Experimental Testing, and Therapeutic Applications. Int. J. Mol. Sci. 2018, 19, 1114. [Google Scholar] [CrossRef] [Green Version]
- Ridlon, J.M.; Kang, D.-J.; Hylemon, P.B. Bile Salt Biotransformations by Human Intestinal Bacteria. J. Lipid Res. 2006, 47, 241–259. [Google Scholar] [CrossRef] [Green Version]
- Chávez-Talavera, O.; Tailleux, A.; Lefebvre, P.; Staels, B. Bile Acid Control of Metabolism and Inflammation in Obesity, Type 2 Diabetes, Dyslipidemia, and Nonalcoholic Fatty Liver Disease. Gastroenterology 2017, 152, 1679–1694.e3. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; Amar, J.; Iglesias, M.A.; Poggi, M.; Knauf, C.; Bastelica, D.; Neyrinck, A.M.; Fava, F.; Tuohy, K.M.; Chabo, C.; et al. Metabolic Endotoxemia Initiates Obesity and Insulin Resistance. Diabetes 2007, 56, 1761–1772. [Google Scholar] [CrossRef] [Green Version]
- Micha, R.; Khatibzadeh, S.; Shi, P.; Fahimi, S.; Lim, S.; Andrews, K.G.; Engell, R.E.; Powles, J.; Ezzati, M.; Mozaffarian, D. Global, Regional, and National Consumption Levels of Dietary Fats and Oils in 1990 and 2010: A Systematic Analysis Including 266 Country-Specific Nutrition Surveys. BMJ 2014, 348, g2272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McEwen, B.S. Neurobiological and Systemic Effects of Chronic Stress. Chronic Stress 2017, 1, 247054701769232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cryan, J.F.; Dinan, T.G. Mind-Altering Microorganisms: The Impact of the Gut Microbiota on Brain and Behaviour. Nat. Rev. Neurosci. 2012, 13, 701–712. [Google Scholar] [CrossRef]
- Heijtz, R.D.; Wang, S.; Anuar, F.; Qian, Y.; Björkholm, B.; Samuelsson, A.; Hibberd, M.L.; Forssberg, H.; Pettersson, S. Normal Gut Microbiota Modulates Brain Development and Behavior. Proc. Natl. Acad. Sci. USA 2011, 108, 3047–3052. [Google Scholar] [CrossRef] [Green Version]
- Desbonnet, L.; Clarke, G.; Shanahan, F.; Dinan, T.G.; Cryan, J.F. Microbiota Is Essential for Social Development in the Mouse. Mol. Psychiatry 2014, 19, 146–148. [Google Scholar] [CrossRef]
- Karl, J.P.; Margolis, L.M.; Madslien, E.H.; Murphy, N.E.; Castellani, J.W.; Gundersen, Y.; Hoke, A.V.; Levangie, M.W.; Kumar, R.; Chakraborty, N.; et al. Changes in Intestinal Microbiota Composition and Metabolism Coincide with Increased Intestinal Permeability in Young Adults under Prolonged Physiological Stress. Am. J. Physiol. Gastrointest. Liver Physiol. 2017, 312, G559–G571. [Google Scholar] [CrossRef] [Green Version]
- Li, H.L.; Lu, L.; Wang, X.S.; Qin, L.Y.; Wang, P.; Qiu, S.P.; Wu, H.; Huang, F.; Zhang, B.B.; Shi, H.L.; et al. Alteration of Gut Microbiota and Inflammatory Cytokine/Chemokine Profiles in 5-Fluorouracil Induced Intestinal Mucositis. Front. Cell. Infect. Microbiol. 2017, 7. [Google Scholar] [CrossRef]
- Garaulet, M.; Gómez-Abellán, P.; Alburquerque-Béjar, J.J.; Lee, Y.C.; Ordovás, J.M.; Scheer, F.A.J.L. Timing of Food Intake Predicts Weight Loss Effectiveness. Int. J. Obes. 2013, 37, 604–611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, X.; FitzGerald, G.A. Timing the Microbes: The Circadian Rhythm of the Gut Microbiome. J. Biol. Rhythms 2017, 32, 505–515. [Google Scholar] [CrossRef]
- Benedict, C.; Vogel, H.; Jonas, W.; Woting, A.; Blaut, M.; Schürmann, A.; Cedernaes, J. Gut Microbiota and Glucometabolic Alterations in Response to Recurrent Partial Sleep Deprivation in Normal-Weight Young Individuals. Mol. Metab. 2016, 5, 1175–1186. [Google Scholar] [CrossRef] [PubMed]
- Gottesmann, C. GABA Mechanisms and Sleep. Neuroscience 2002, 111, 231–239. [Google Scholar] [CrossRef]
- Ursin, R. Serotonin and Sleep. Sleep Med. Rev. 2002, 6, 55–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yunes, R.A.; Poluektova, E.U.; Dyachkova, M.S.; Klimina, K.M.; Kovtun, A.S.; Averina, O.V.; Orlova, V.S.; Danilenko, V.N. GABA Production and Structure of GadB/GadC Genes in Lactobacillus and Bifidobacterium Strains from Human Microbiota. Anaerobe 2016, 42, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Valles-Colomer, M.; Falony, G.; Darzi, Y.; Tigchelaar, E.F.; Wang, J.; Tito, R.Y.; Schiweck, C.; Kurilshikov, A.; Joossens, M.; Wijmenga, C.; et al. The Neuroactive Potential of the Human Gut Microbiota in Quality of Life and Depression. Nat. Microbiol. 2019, 4, 623–632. [Google Scholar] [CrossRef]
- Maukonen, M.; Kanerva, N.; Partonen, T.; Kronholm, E.; Tapanainen, H.; Kontto, J.; Männistö, S. Chronotype Differences in Timing of Energy and Macronutrient Intakes: A Population-Based Study in Adults. Obesity 2017, 25, 608–615. [Google Scholar] [CrossRef] [Green Version]
- Turek, F.W.; Joshu, C.; Kohsaka, A.; Lin, E.; Ivanova, G.; McDearmon, E.; Laposky, A.; Losee-Olson, S.; Easton, A.; Jensen, D.R.; et al. Obesity and Metabolic Syndrome in Circadian Clock Mutant Nice. Science 2005, 308, 1043–1045. [Google Scholar] [CrossRef] [Green Version]
- Vera, B.; Dashti, H.S.; Gómez-Abellán, P.; Hernández-Martínez, A.M.; Esteban, A.; Scheer, F.A.J.L.; Saxena, R.; Garaulet, M. Modifiable Lifestyle Behaviors, but Not a Genetic Risk Score, Associate with Metabolic Syndrome in Evening Chronotypes. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef] [Green Version]
- Bandín, C.; Scheer, F.A.J.L.; Luque, A.J.; Ávila-Gandiá, V.; Zamora, S.; Madrid, J.A.; Gómez-Abellán, P.; Garaulet, M. Meal Timing Affects Glucose Tolerance, Substrate Oxidation and Circadian-Related Variables: A Randomized, Crossover Trial. Int. J. Obes. 2015, 39, 828–833. [Google Scholar] [CrossRef] [PubMed]
- Skonieczna-Żydecka, K.; Marlicz, W.; Misera, A.; Koulaouzidis, A.; Łoniewski, I. Microbiome—The Missing Link in the Gut-Brain Axis: Focus on Its Role in Gastrointestinal and Mental Health. J. Clin. Med. 2018, 7, 521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, A.M.; Thompson, S.V.; Edwards, C.G.; Musaad, S.M.A.; Khan, N.A.; Holscher, H.D. Associations among Diet, the Gastrointestinal Microbiota, and Negative Emotional States in Adults. Nutr. Neurosci. 2019, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, A.; Lehto, S.M.; Harty, S.; Dinan, T.G.; Cryan, J.F.; Burnet, P.W.J. Psychobiotics and the Manipulation of Bacteria–Gut–Brain Signals. Trends Neurosci. 2016, 39, 763–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.N.; Yun, Y.; Ryu, S.; Chang, Y.; Kwon, M.J.; Cho, J.; Shin, H.; Kim, H.L. Correlation between Gut Microbiota and Personality in Adults: A Cross-Sectional Study. Brain. Behav. Immun. 2018, 69, 374–385. [Google Scholar] [CrossRef] [PubMed]
- Sutin, A.R.; Terracciano, A.; Deiana, B.; Naitza, S.; Ferrucci, L.; Uda, M.; Schlessinger, D.; Costa, P.T. High Neuroticism and Low Conscientiousness Are Associated with Interleukin-6. Psychol. Med. 2010, 40, 1485–1493. [Google Scholar] [CrossRef] [Green Version]
- Audet, M.C. Stress-Induced Disturbances along the Gut Microbiota-Immune-Brain Axis and Implications for Mental Health: Does Sex Matter? Front. Neuroendocrinol. 2019, 54. [Google Scholar] [CrossRef]
- Baxter, A.J.; Scott, K.M.; Vos, T.; Whiteford, H.A. Global Prevalence of Anxiety Disorders: A Systematic Review and Meta-Regression. Psychol. Med. 2013, 43, 897–910. [Google Scholar] [CrossRef]
- Clarke, S.F.; Murphy, E.F.; O’Sullivan, O.; Lucey, A.J.; Humphreys, M.; Hogan, A.; Hayes, P.; O’Reilly, M.; Jeffery, I.B.; Wood-Martin, R.; et al. Exercise and Associated Dietary Extremes Impact on Gut Microbial Diversity. Gut 2014, 63, 1913–1920. [Google Scholar] [CrossRef] [Green Version]
- Bressa, C.; Bailén-Andrino, M.; Pérez-Santiago, J.; González-Soltero, R.; Pérez, M.; Montalvo-Lominchar, M.G.; Maté-Muñoz, J.L.; Domínguez, R.; Moreno, D.; Larrosa, M. Differences in Gut Microbiota Profile between Women with Active Lifestyle and Sedentary Women. PLoS ONE 2017, 12. [Google Scholar] [CrossRef] [Green Version]
- Cronin, O.; Barton, W.; Skuse, P.; Penney, N.C.; Garcia-Perez, I.; Murphy, E.F.; Woods, T.; Nugent, H.; Fanning, A.; Melgar, S.; et al. A Prospective Metagenomic and Metabolomic Analysis of the Impact of Exercise and/or Whey Protein Supplementation on the Gut Microbiome of Sedentary Adults. mSystems 2018, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cronin, O.; O’Sullivan, O.; Barton, W.; Cotter, P.D.; Molloy, M.G.; Shanahan, F. Gut Microbiota: Implications for Sports and Exercise Medicine. Br. J. Sports Med. 2017, 51, 700–701. [Google Scholar] [CrossRef] [PubMed]
- Luo, B.; Xiang, D.; Nieman, D.C.; Chen, P. The Effects of Moderate Exercise on Chronic Stress-Induced Intestinal Barrier Dysfunction and Antimicrobial Defense. Brain. Behav. Immun. 2014, 39, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Cerdá, B.; Pérez, M.; Pérez-Santiago, J.D.; Tornero-Aguilera, J.F.; González-Soltero, R.; Larrosa, M. Gut Microbiota Modification: Another Piece in the Puzzle of the Benefits of Physical Exercise in Health? Front. Physiol. 2016, 7, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blaser, M.J. Our Missing Microbes: Short-Term Antibiotic Courses Have Long-Term Consequences. Cleve. Clin. J. Med. 2018, 85, 928–930. [Google Scholar] [CrossRef] [Green Version]
- Falony, G.; Joossens, M.; Vieira-Silva, S.; Wang, J.; Darzi, Y.; Faust, K.; Kurilshikov, A.; Bonder, M.J.; Valles-Colomer, M.; Vandeputte, D.; et al. Population-Level Analysis of Gut Microbiome Variation. Science 2016, 352, 560–564. [Google Scholar] [CrossRef]
- Kennedy, E.A.; King, K.Y.; Baldridge, M.T. Mouse Microbiota Models: Comparing Germ-Free Mice and Antibiotics Treatment as Tools for Modifying Gut Bacteria. Front. Physiol. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Covington, A.; Pamer, E.G. The Intestinal Microbiota: Antibiotics, Colonization Resistance, and Enteric Pathogens. Immunol. Rev. 2017, 279, 90–105. [Google Scholar] [CrossRef]
- Ticinesi, A.; Milani, C.; Lauretani, F.; Nouvenne, A.; Mancabelli, L.; Lugli, G.A.; Turroni, F.; Duranti, S.; Mangifesta, M.; Viappiani, A.; et al. Gut Microbiota Composition Is Associated with Polypharmacy in Elderly Hospitalized Patients. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [Green Version]
- Gloux, K.; Berteau, O.; El Oumami, H.; Béguet, F.; Leclerc, M.; Doré, J. A Metagenomic β-Glucuronidase Uncovers a Core Adaptive Function of the Human Intestinal Microbiome. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. 1), 4539–4546. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, M.; Kurita, A.; Asahara, T.; Takakura, A.; Katono, K.; Iwasaki, M.; Ryuge, S.; Wada, M.; Onoda, S.; Yanaihara, T.; et al. Metabolism of Irinotecan and Its Active Metabolite SN-38 by Intestinal Microflora in Rats. Oncol. Rep. 2008, 20, 727–730. [Google Scholar] [PubMed]
- Clayton, T.A.; Baker, D.; Lindon, J.C.; Everett, J.R.; Nicholson, J.K. Pharmacometabonomic Identification of a Significant Host-Microbiome Metabolic Interaction Affecting Human Drug Metabolism. Proc. Natl. Acad. Sci. USA 2009, 106, 14728–14733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dujardin, C.E.; Mars, R.A.T.; Manemann, S.M.; Kashyap, P.C.; Clements, N.S.; Hassett, L.C.; Roger, V.L. Impact of Air Quality on the Gastrointestinal Microbiome: A Review. Environ. Res. 2020, 186. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, G.G.; Hubbard, J.; Korzenik, J.; Sands, B.E.; Panaccione, R.; Ghosh, S.; Wheeler, A.J.; Villeneuve, P.J. The Inflammatory Bowel Diseases and Ambient Air Pollution: A Novel Association. Am. J. Gastroenterol. 2010, 105, 2412–2419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Opstelten, J.L.; Plassais, J.; Van Mil, S.W.C.; Achouri, E.; Pichaud, M.; Siersema, P.D.; Oldenburg, B.; Cervino, A.C.L. Gut Microbial Diversity Is Reduced in Smokers with Crohn’s Disease. Inflamm. Bowel Dis. 2016, 22, 2070–2077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biedermann, L.; Zeitz, J.; Mwinyi, J.; Sutter-Minder, E.; Rehman, A.; Ott, S.J.; Steurer-Stey, C.; Frei, A.; Frei, P.; Scharl, M.; et al. Smoking Cessation Induces Profound Changes in the Composition of the Intestinal Microbiota in Humans. PLoS ONE 2013, 8. [Google Scholar] [CrossRef]
- Berkowitz, L.; Schultz, B.M.; Salazar, G.A.; Pardo-Roa, C.; Sebastián, V.P.; Álvarez-Lobos, M.M.; Bueno, S.M. Impact of Cigarette Smoking on the Gastrointestinal Tract Inflammation: Opposing Effects in Crohn’s Disease and Ulcerative Colitis. Front. Immunol. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- McBurney, M.I.; Davis, C.; Fraser, C.M.; Schneeman, B.O.; Huttenhower, C.; Verbeke, K.; Walter, J.; Latulippe, M.E. Establishing What Constitutes a Healthy Human Gut Microbiome: State of the Science, Regulatory Considerations, and Future Directions. J. Nutr. 2019, 1882–1895. [Google Scholar] [CrossRef]
- Gomez, A.; Petrzelkova, K.J.; Burns, M.B.; Yeoman, C.J.; Amato, K.R.; Vlckova, K.; Modry, D.; Todd, A.; Jost Robinson, C.A.; Remis, M.J.; et al. Gut Microbiome of Coexisting BaAka Pygmies and Bantu Reflects Gradients of Traditional Subsistence Patterns. Cell Rep. 2016, 14, 2142–2153. [Google Scholar] [CrossRef] [Green Version]
- Warnecke, F.; Luginbühl, P.; Ivanova, N.; Ghassemian, M.; Richardson, T.H.; Stege, J.T.; Cayouette, M.; McHardy, A.C.; Djordjevic, G.; Aboushadi, N.; et al. Metagenomic and Functional Analysis of Hindgut Microbiota of a Wood-Feeding Higher Termite. Nature 2007, 450, 560–565. [Google Scholar] [CrossRef]
- Zimmer, J.; Lange, B.; Frick, J.S.; Sauer, H.; Zimmermann, K.; Schwiertz, A.; Rusch, K.; Klosterhalfen, S.; Enck, P. A Vegan or Vegetarian Diet Substantially Alters the Human Colonic Faecal Microbiota. Eur. J. Clin. Nutr. 2012, 66, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Nam, Y.D.; Jung, M.J.; Roh, S.W.; Kim, M.S.; Bae, J.W. Comparative Analysis of Korean Human Gut Microbiota by Barcoded Pyrosequencing. PLoS ONE 2011, 6. [Google Scholar] [CrossRef]
- Schnorr, S.L.; Candela, M.; Rampelli, S.; Centanni, M.; Consolandi, C.; Basaglia, G.; Turroni, S.; Biagi, E.; Peano, C.; Severgnini, M.; et al. Gut Microbiome of the Hadza Hunter-Gatherers. Nat. Commun. 2014, 5. [Google Scholar] [CrossRef]
- Bédard, A.; Riverin, M.; Dodin, S.; Corneau, L.; Lemieux, S. Sex Differences in the Impact of the Mediterranean Diet on Cardiovascular Risk Profile. Br. J. Nutr. 2012, 108, 1428–1434. [Google Scholar] [CrossRef] [PubMed]
- Vom Steeg, L.G.; Klein, S.L. SeXX Matters in Infectious Disease Pathogenesis. PLoS Pathog. 2016, 12, e1005374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fish, E.N. The X-Files in Immunity: Sex-Based Differences Predispose Immune Responses. Nat. Rev. Immunol. 2008, 8, 737–744. [Google Scholar] [CrossRef] [PubMed]
- Markle, J.G.M.; Frank, D.N.; Mortin-Toth, S.; Robertson, C.E.; Feazel, L.M.; Rolle-Kampczyk, U.; Von Bergen, M.; McCoy, K.D.; Macpherson, A.J.; Danska, J.S. Sex Differences in the Gut Microbiome Drive Hormone-Dependent Regulation of Autoimmunity. Science 2013, 339, 1084–1088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliver, J.E.; Silman, A.J. Why Are Women Predisposed to Autoimmune Rheumatic Diseases? Arthritis Res. Ther. 2009, 11. [Google Scholar] [CrossRef] [Green Version]
- Bianchi, I.; Lleo, A.; Gershwin, M.E.; Invernizzi, P. The X Chromosome and Immune Associated Genes. J. Autoimmun. 2012, 38, J187–J192. [Google Scholar] [CrossRef]
- Yurkovetskiy, L.; Burrows, M.; Khan, A.A.; Graham, L.; Volchkov, P.; Becker, L.; Antonopoulos, D.; Umesaki, Y.; Chervonsky, A.V. Gender Bias in Autoimmunity Is Influenced by Microbiota. Immunity 2013, 39, 400–412. [Google Scholar] [CrossRef] [Green Version]
- Brennan, L.; Gibbons, H. Sex Matters: A Focus on the Impact of Biological Sex on Metabolomic Profiles and Dietary Interventions. Proc. Nutr. Soc. 2020, 79, 205–209. [Google Scholar] [CrossRef] [PubMed]
- Kundu, P.; Blacher, E.; Elinav, E.; Pettersson, S. Our Gut Microbiome: The Evolving Inner Self. Cell 2017, 171, 1481–1493. [Google Scholar] [CrossRef] [PubMed]
- Vieira, A.T.; Castelo, P.M.; Ribeiro, D.A.; Ferreira, C.M. Influence of Oral and Gut Microbiota in the Health of Menopausal Women. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef]
- Parida, S.; Sharma, D. The Microbiome–Estrogen Connection and Breast Cancer Risk. Cells 2019, 8, 1642. [Google Scholar] [CrossRef] [Green Version]
- Dabek, M.; McCrae, S.I.; Stevens, V.J.; Duncan, S.H.; Louis, P. Distribution of β-Glucosidase and β-Glucuronidase Activity and of β-Glucuronidase Gene Gus in Human Colonic Bacteria. FEMS Microbiol. Ecol. 2008, 66, 487–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mestdagh, R.; Dumas, M.E.; Rezzi, S.; Kochhar, S.; Holmes, E.; Claus, S.P.; Nicholson, J.K. Gut Microbiota Modulate the Metabolism of Brown Adipose Tissue in Mice. J. Proteome Res. 2012, 11, 620–630. [Google Scholar] [CrossRef] [PubMed]
- Santos-Marcos, J.A.; Rangel-Zuñiga, O.A.; Jimenez-Lucena, R.; Quintana-Navarro, G.M.; Garcia-Carpintero, S.; Malagon, M.M.; Landa, B.B.; Tena-Sempere, M.; Perez-Martinez, P.; Lopez-Miranda, J.; et al. Influence of Gender and Menopausal Status on Gut Microbiota. Maturitas 2018, 116, 43–53. [Google Scholar] [CrossRef]
- Haro, C.; Rangel-Zúñiga, O.A.; Alcalá-Díaz, J.F.; Gómez-Delgado, F.; Pérez-Martínez, P.; Delgado-Lista, J.; Quintana-Navarro, G.M.; Landa, B.B.; Navas-Cortés, J.A.; Tena-Sempere, M.; et al. Intestinal Microbiota Is Influenced by Gender and Body Mass Index. PLoS ONE 2016, 11, e0154090. [Google Scholar] [CrossRef] [Green Version]
- Borgo, F.; Garbossa, S.; Riva, A.; Severgnini, M.; Luigiano, C.; Benetti, A.; Pontiroli, A.E.; Morace, G.; Borghi, E. Body Mass Index and Sex Affect Diverse Microbial Niches within the Gut. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Mueller, S.; Saunier, K.; Hanisch, C.; Norin, E.; Alm, L.; Midtvedt, T.; Cresci, A.; Silvi, S.; Orpianesi, C.; Verdenelli, M.C.; et al. Differences in Fecal Microbiota in Different European Study Populations in Relation to Age, Gender, and Country: A Cross-Sectional Study. Appl. Environ. Microbiol. 2006, 72, 1027–1033. [Google Scholar] [CrossRef] [Green Version]
- Kumbhare, S.V.; Patangia, D.V.V.; Patil, R.H.; Shouche, Y.S.; Patil, N.P. Factors Influencing the Gut Microbiome in Children: From Infancy to Childhood. J. Biosci. 2019, 44. [Google Scholar] [CrossRef]
- Cong, X.; Xu, W.; Janton, S.; Henderson, W.A.; Matson, A.; McGrath, J.M.; Maas, K.; Graf, J. Gut Microbiome Developmental Patterns in Early Life of Preterm Infants: Impacts of Feeding and Gender. PLoS ONE 2016, 11. [Google Scholar] [CrossRef] [PubMed]
- Koenig, J.E.; Spor, A.; Scalfone, N.; Fricker, A.D.; Stombaugh, J.; Knight, R.; Angenent, L.T.; Ley, R.E. Succession of Microbial Consortia in the Developing Infant Gut Microbiome. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. 1), 4578–4585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biagi, E.; Candela, M.; Turroni, S.; Garagnani, P.; Franceschi, C.; Brigidi, P. Ageing and Gut Microbes: Perspectives for Health Maintenance and Longevity. Pharmacol. Res. 2013, 69, 11–20. [Google Scholar] [CrossRef]
- Biagi, E.; Rampelli, S.; Turroni, S.; Quercia, S.; Candela, M.; Brigidi, P. The Gut Microbiota of Centenarians: Signatures of Longevity in the Gut Microbiota Profile. Mech. Ageing Dev. 2017, 165, 180–184. [Google Scholar] [CrossRef]
- Biau, D.J.; Kernéis, S.; Porcher, R. Statistics in Brief: The Importance of Sample Size in the Planning and Interpretation of Medical Research. Clin. Orthop. Relat. Res. 2008, 466, 2282–2288. [Google Scholar] [CrossRef] [Green Version]
- Claesson, M.J.; O’Sullivan, O.; Wang, Q.; Nikkilä, J.; Marchesi, J.R.; Smidt, H.; de Vos, W.M.; Ross, R.P.; O’Toole, P.W. Comparative Analysis of Pyrosequencing and a Phylogenetic Microarray for Exploring Microbial Community Structures in the Human Distal Intestine. PLoS ONE 2009, 4, e6669. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Yu, Z.; Ye, X.; Zou, S.; Li, H.; Yu, D.; Wu, H.; Chen, Y.; Dore, J.; Clement, K.; et al. A Marker of Endotoxemia Is Associated With Obesity and Related Metabolic Disorders in Apparently Healthy Chinese. Diabetes Care 2010, 33, 1925–1932. [Google Scholar] [CrossRef] [Green Version]





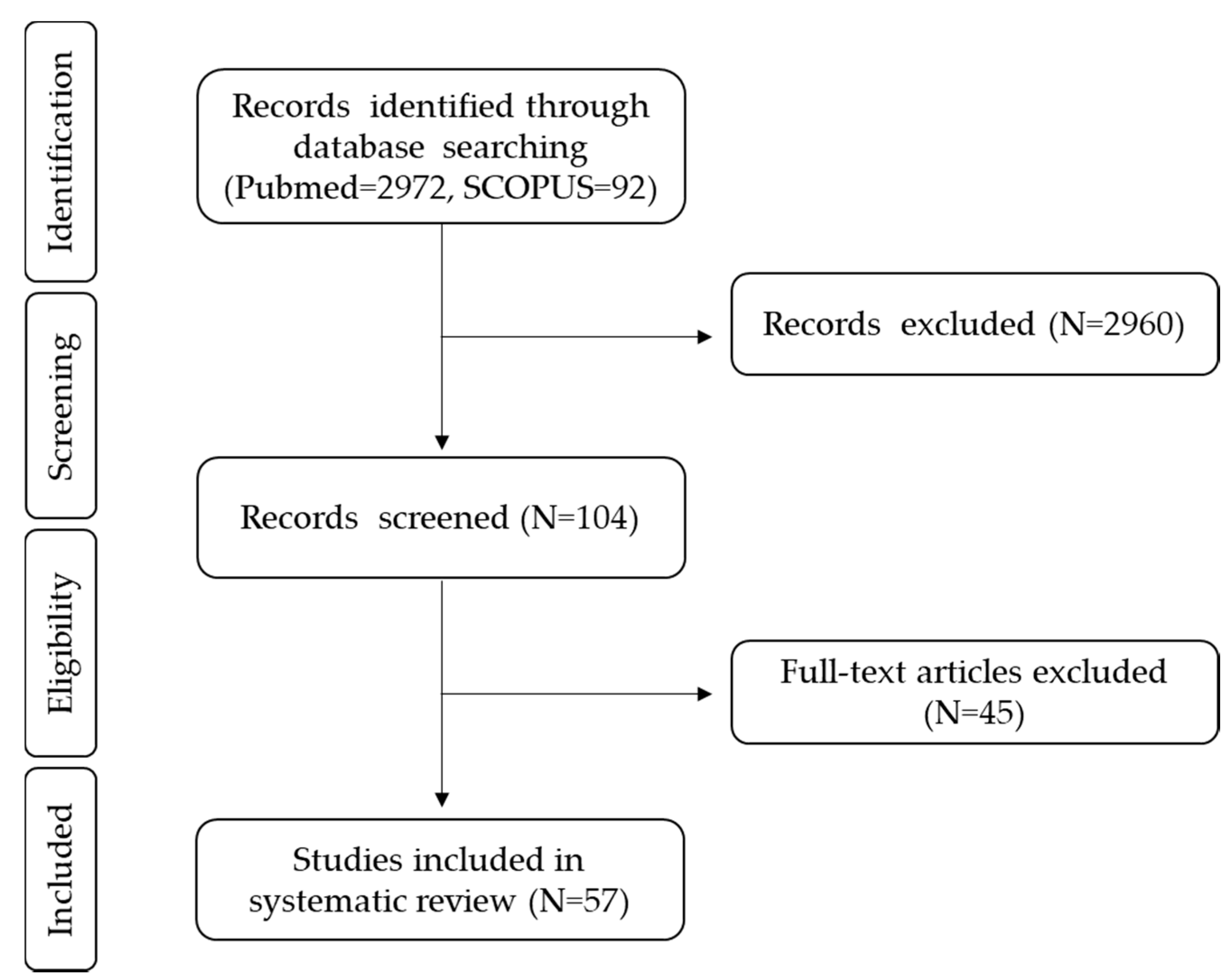
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ref. | Treatment | Study Type | Study Subjects | Analytic Technique | Results | Results |
|---|---|---|---|---|---|---|
| Yogurt/Probiotic vs. Control (C) | Differences vs. Basal Point | Differences vs. Control (C) | ||||
| Yogurt | ||||||
| [37] | Yogurt (108 cfu/g) | Randomized, parallel | 15 (6 W/9 M) | Culture + PCR | ↔Lactobacillus strains | -- |
| ↑Clostridium coccoides–Eubacterium rectale group | ||||||
| Duration: 20 d | Sequence-specific SSU rRNA cleavage with oligonucleotides | ↓Bacteroides–Prevotella group | ||||
| C: Comparison to basal point | Age: 24–46 y | ↔Bifidobacterium, C. leptum, Atopobium, Eggerthella, Collinsella, Lactobacillus, Enterococcus | ||||
| BMI: No data | ||||||
| [38] | Yogurt (107–108 cfu/g) | Randomized, DB, crossover | 79 (47 W/32 M) | DGGE | ↑Lactobacillus | ↔No significant changes |
| qPCR | ↓Bacteroides-Porphyromonas-Prevotella | |||||
| Duration: 4 wks | Age: 24 y | |||||
| C: Pasteurized yogurt | BMI: No data | |||||
| Capsules | ||||||
| [39] | Lactobacillus salivarius CECT5713 (2 × 108 cfu/d) | Randomized, DB, | 40 (20 W/20 M) | Culture | ↑Lactobacillus | ↑Lactobacillus |
| Duration: 6 wks | placebo-controlled, parallel | Age: 33 y | ||||
| C: Maltodextrin | BMI: No data | |||||
| [40] | (1) Lactobacillus paracasei ssp. paracasei CRL-431 (1011 cfu/d) | Randomized, DB, | 71 (46 W/25 M) | Culture | ↔Bacteroides, Bifidobacterium, Clostridia, Enterobacteriaceae, Enterococcus, Lactobacillus | ↔Bacteroides, Bifidobacterium, Clostridia, Enterobacteriaceae, Enterococcus, Lactobacillus |
| placebo-controlled, parallel | ||||||
| (2) Bifidobacterium animalis subsp. lactis BB-12 (1011 cfu/d) | Age: 26 y | |||||
| BMI: No data | ||||||
| Duration: 3 wks | ||||||
| C: Dextrose | ||||||
| [41] | Lactobacillus Zhang (1010 cfu/d) | Randomized, parallel | 24 (13 W/11 M) | 16s RNA gene Pyrosequencing | ↑β-diversity (Unifrac), Bifidobacterium, Fecalibacterium, Prevotella | -- |
| Duration: 4 wks | Age: 41 y | (V5–V6 regions, | ↓Blautia coccoides, Phascolarctobacterium, | |||
| BMI: 19.5–28.2 kg/m2 | ||||||
| C: Comparison to basal point | Roche) | Enterobacter | ||||
| qPCR | ||||||
| [42] | Lactobacillus paracasei DG (2.4 × 1013 cfu/d) | Randomized, DB, | 34 (19 W/15 M) | 16s RNA gene Sequencing | ↔α-diversity (Chao1, Shannon) | ↑Coprococcus |
| Duration: 4 wks | placebo-controlled, crossover | ↑β-diversity (Unifrac) | ↓Proteobacterias, B. coccoides | |||
| C: Maltodextrin + starch | Age: 35 y | (V3 region, Ion Torrent) | ||||
| BMI: 20–25 kg/m2 | ||||||
| [43] | Lactobacilluscasei (106–108 cfu/d); Lactobacillu brevis (106–108 cfu/d); Bifidobacterium longum + Lactobacillus lactis + Streptococcus thermophilus (106–108 cfu/d); Lactobacillus rhamnosus (106–108 cfu/d); Lactobacillus delbrueckii + St. thermophilus (106–108 cfu/d); B. animalis + Lactobacillus delbrueckii + St. thermophilus (106–108 cfu/d). Duration: 8 wks | Randomized, parallel | 18 (12 W/6 M) | 16s RNA gene Pyrosequencing | ↔β-diversity (Unifrac) | -- |
| ↑Firmicutes species after Lactobacillus and Bifidobacterium intake, Bacteroidetes species after Bifidobacterium intake | ||||||
| Age: 22 y | (V1-V2 regions, Roche) | |||||
| BMI: No data | ||||||
| C: Comparison to basal point | ||||||
| [44] | Bifidobacterium longum BB536 (4 × 108 cfu/d) + L. rhamnosus HN001 (109 cfu/d) | Randomized | 16 (4 W/12 M) | qPCR | ↔α-diversity (Chao1, Shannon, Simpson) | -- |
| 16s RNA gene Sequencing | ↓Firmicutes, Proteobacterias | |||||
| Age: 36 y | ↑Blautia producta, Blautia wexlerae, Haemophilus ducrey | |||||
| (V2-V4-V8, V3-V6, V7-V9 regions, Ion Torrent) | ↓Holdemania filiformis, Eubacterium. vulneris, Gemminer formicilis, Streptococcus sinensis | |||||
| Duration: 1 month | ||||||
| BMI: 20–26 kg/m2 | ||||||
| C: Comparison to basal point | ||||||
| [45] | Lactobacilluskefiri LKF01 (1010 cfu/d) | Randomized | 20 (16 W/4 M) | qPCR | ↔α-diversity (Chao1, Shannon, Simpson) | -- |
| Duration: 1 month | 16s RNA gene Sequencing | ↓Firmicutes, Bacteroides, Proteobacteria, Bilophila spp, Butyricicomonas spp, Flavonifractor spp, Oscillibacter spp, Prevotella spp | ||||
| C: Comparison to basal point | Age: 39 y | |||||
| (V2-V4-V8, V3-V6, V7-V9 regions, Ion Torrent) | ||||||
| BMI: 18.5–25 kg/m2 | ↑Lactobacillus | |||||
| [46] | Bifidobacterium bifidum strain Bb (3.8 × 109 cfu/d) | Randomized, DB, | 27 (13 W/14 M) | 16s RNA gene Sequencing | ↔α-diversity (Chao1, Shannon, Simpson), β-diversity (Unifrac) | -- |
| Age: 31 y | ||||||
| BMI: No data | ||||||
| (V3 region, Ion Torrent) | ||||||
| placebo-controlled, crossover | ↓Prevotellaceae | |||||
| Duration: 4 wks | ↑Rikenellaceae, Ruminococcaceae | |||||
| C: Maltodextrin | ||||||
| [47] | L. rhamnosus IMC501 + L. paracasei IMC502 (109 cfu/serving) | Randomized, DB, | 50 (27 W/23 M) | Cultures | ↔ Clostridium, Enterobacteriaceae, Bacteroides | ↑Lactobacillus, Bifidobacterium |
| placebo-controlled, parallel | qPCR | ↑Lactobacillus, ifidobacterium | ||||
| Durantion:12 wks | Age: 23–65 y | |||||
| C: food products without probiotics | BMI: No data | |||||
| Probiotics Fermented Milks | ||||||
| [48] | (1) Lactobacillus coryniformis CECT5711 (2 × 108 cfu/d) | Randomized, DB, | 30 (15 W/15 M) | Culture | ↑Lactobacillus | ↑Lactobacillus |
| (2) L. gasseri CECT5714 (2 × 108 cfu/d) | placebo-controlled, parallel | RADP | ||||
| Age: 23–43 y | ||||||
| Duration: 2 wks | BMI: No data | |||||
| C: Yogurt | ||||||
| [49] | L. paracasei ssp. paracasei LC01 (2 × 108 cfu/mL) | Randomized, DB, | 52 (31 W/21 M) | qPCR | ↑Lactobacillus, Roseburia intestinalis, Bifidobacterium, E. coli | ↑Lactobacillus, R. intestinalis |
| placebo-controlled, parallel | ↓Escherichia coli | |||||
| Duration: 4 wks | Age: 24 y | |||||
| C: Semi-skimmed milk | BMI: 19–29 kg/m2 | |||||
| [50] | L. acidophilus LA-5 (109 cfu/d) + | Randomized, DB, | 58 (38 W/20 M) | qPCR | ↑Bifidobacterium | ↑Bifidobacterium |
| B. animalis ssp.lactis BB-12 (109–1010 cfu/d) | placebo-controlled, parallel | ↑Lactobacillus | ||||
| Duration: 4 wks | Age: 32 y | Culture | ↑Lactobacillus | ↓Enterococcus | ||
| BMI: 20–28kg/m2 | ↓Enterococcus | |||||
| C: Pasterized Yogurt | ||||||
| [51] | L. acidophilus LA-5 (109 cfu/d) + | Randomized, | 58 (38 W/20 M) | T-RFLP | ↔β-diversity (Bray-Curtis) | ↔β-diversity (Bray-Curtis) |
| B. animalis ssp.lactis BB-12 (109–1010 cfu/d) | placebo-controlled, parallel | qPCR | ↔Bacteroides-Prevotella, B. coccoides, | ↔Bacteroides-Prevotella, | ||
| Duration: 4 wks | Age and BMI: No | C. leptum, Enterobacteriaceae, Enterococcus, Bifidobacterium | B. coccoides, C. leptum Enterobacteriaceae, | |||
| Enterococcus, Bifidobacterium | ||||||
| data | ||||||
| C: Yogurt | ||||||
| [52] | L. rhamnosus GG (2.83 × 106 cfu/g) | Randomized, crossover | 13 (0 W/13 M) | 16s RNA gene sequencing (V3-V4 regions, Illumina) | ↔ α-diversity (Shannon, Simpson) | ↑Intestinibacter bartlettii |
| Duration: 2 wks | ↓ Bilophila wadsworthia | ↓B. kashiwanohense, | ||||
| Age: 24 y | B. pseudocatenulatum, Megasphaera | |||||
| C: Milk acidified with 2% of D- (+)-glucono-δ-lactone (400 g) | BMI: 18.5–25 kg/m2 | |||||
| [53] | B. animalis ssp. lactis BB-12 (107 cfu/mL) | Parallel | 150 (No gender data) | 16s RNA gene Sequencing | ↔α-diversity (Chao1, Shannon) | -- |
| Duration: 1 month | ||||||
| C: Comparison to basal point | ↑β-diversity (Bray-Curtis), Bifidobacterium, Slackia, Streptococcus, Catenibacterium, Collinsella, Adlercreutzia | |||||
| Age: 18–40 y | (V4 region, Illumina) | |||||
| BMI: 18–28 kg/m2 | ||||||
| ↓Lachnoclostridium, Roseburia, Acidaminococcaceae | ||||||
| [26] | B. animalis ssp.lactis, L. delbrueckii ssp. bulgaricus, Lactococcus lactis ssp. cremoris, St. thermophilus [2 units/d (3.2 × 107 GE + 6.3 × 107 GE)] | Randomized, parallel | 14 (14 W/0 M) | 16s RNA gene Pyrosequencing | ↔No significant changes | -- |
| Age: 21–32 y | (V2 region, 454 FLX) | |||||
| BMI: No data | ||||||
| Duration: 7 wks | ||||||
| C: Comparison to basal point | ||||||
| [54] | L. casei DN-114001 (108 cfu/mL) | Randomized | 12 (7 W/5 M) | qPCR | ↑L. casei | -- |
| Duration: 10 d | FISH | ↔C.coccoides, F. prausnitzii, Bacteroides, Bifidobacterium, Atopobium, | ||||
| Age: 23–44 y | ||||||
| C: Comparison to basal point | BMI: No data | Lactobacilli–Enterococci, Enterobacteria | ||||
| a. Evidence from human studies relative to fiber effects on the gut microbiota composition of healthy adults | |||||||
|---|---|---|---|---|---|---|---|
| Ref. | Treatments | Study Type | Study Subjects | Analytic Technique | Results | Results | Results |
| Fiber vs. Control (C) | Differences vs. Basal Point | Differences between Treatment Groups | Differences vs. Control (C) | ||||
| Accepted Prebiotic Fibers | |||||||
| [55] | Agave fructan (5 g/d) | Randomized, DB, placebo-controlled crossover | 38 (19 W/19 M) | Colon culture model | ↑Bifidobacterium, Lactobacillus-Enterococcus group | -- | ↑Bifidobacterium, Lactobacillus-Enterococcus group |
| Age: 35 y | |||||||
| FISH | |||||||
| DP: 3–30 | BMI: 21.1–27.1 kg/m2 | PCR | |||||
| Duration: 3 wks | |||||||
| C: Maltodextrin | |||||||
| [56] | Agave inulin (5 or 7.5 g/d) | Randomized, DB, placebo-controlled crossover | 29 (No gender data) | 16s RNA gene Sequencing | -- | ↔No significant changes | (1) 5, 7.5 g: ↑Actinobacterias, Bifidobacteriaceae, Bifidobacterium, |
| DP: 25–34 | Age: 27 y | ||||||
| Duration: 3 wks | BMI: 18.5–29.5 kg/m2 | (V4 region, Illumina) | B. adolescentis, B. breve, B. longum, | ||||
| C: Agave inulin (0 g/d) | B. pseudolongum | ||||||
| ↓Desulfovibrio | |||||||
| (2) 7.5 g: ↓Lachnobacterium, Ruminococcus | |||||||
| [57] | β2–1 fructan | Randomized, DB, placebo-controlled crossover | 30 (17 W/13 M) | qPCR | ↑Bifidobacterium | -- | ↑Bifidobacterium |
| (inulin and short-chain oligosaccharides) (15 g/d) | Age: 28.1 y | ||||||
| BMI: 21.2–27.2 kg/m2 | |||||||
| Duration: 4 wks | |||||||
| C: Maltodextrin | |||||||
| [58] | FOS | Randomized, DB, crossover | (1)FOS: 34 | 16s RNA gene sequencing | (1) FOS: ↑Bifidobacterium | No statistical analysis performed | -- |
| GOS | (24 W/10 M) | ↓Phascolarctobacterium, Enterobacter, Turicibacter, Coprococcus, Salmonella | |||||
| (16 g/d) | Age: 21.9 y | ||||||
| Duration: 2 wks | BMI: 19.8–26.4 kg/m2 | (V2 region, Ion Torrent) | ↔α-diversity (Shannon) | ||||
| C: Comparison to basal point | (2)GOS: 35 | (2) GOS: ↑Bifidobacterium | |||||
| (25 W/10 M) | ↓α-diversity (Chao 1, Shannon, phylogenetic tree), Ruminococcus, Dehalobacterium, Synergistes, Holdemania | ||||||
| Age: 22.1 y | |||||||
| BMI: 19.8–26.4 kg/m2 | |||||||
| [59] | HMO (2-O-fucosyllactose (2′FL), lacto-N-neotetraose (LNnT), 2′FL + LNnT) | Randomized, DB, placebo-controlled parallel | 100 (49 W/51 M) | 16s RNA gene Sequencing (V3-V4 regions, Illumina) | (1) 2′FL (5, 10 g/d), LNnT and 2′FL + LNnT (5, 10, 20 g/d): ↑Actinobacterias | 20 g/d: ↑Actinobacterias | (1) 5, 10, 20 g/d: ↑Actinobacterias |
| Age: 30–40 y | (2) 2′FL (10 g/d), LNnT (5, 10, 20 g/d), 2′FL + LNnT (10, 20 g/d): | ||||||
| BMI: 20–28 kg/m2 | |||||||
| ↑Bifidobacterium, B. adolescentis | |||||||
| (5, 10 or 20 g/d) | (2) 2′FL (10 g/d): ↓Proteobacterias | (3) 2′FL + LNnT (20 g/d): ↑B. longum | |||||
| Duration: 2 wks | (3) LNnT and 2′FL + LNnT (20 g/d): ↓Firmicutes | ||||||
| C: Glucose (2 g) | |||||||
| [60] | Inulin-type fructan (16 g/d) | Randomized, DB, placebo-controlled crossover | 34 (13 W/21 M) | 16s RNA gene Sequencing (V3-V4 regions, Illumina) | HDF, LCF: ↑Actinobacterias, Bifidobacterium, Oscillospira | LDF vs. HDF: ↑Lactobacillus | ↑Actinobacterias, Bifidobacterium, Oscillospira |
| Duration: 3 wks | Age: 37 y | ||||||
| C: Maltodextrin (16 g/d) | BMI: 20–−27 kg/m2 | ↔α and β-diversity | HDF vs. LDF: ↑Ruminococcaceae, Fecalibacterium | ↓Firmicutes, Dorea, Coprococcus, Ruminococcus | |||
| High dietary fiber (HDF) or Low dietary fiber (LDF) | HDF: ↑Bifidobacterium, Fecalibacterium | ||||||
| ↔α-diversity (Shannon, Chao1), | |||||||
| ↓Firmicutes, Dorea, Coprococcus, Ruminococcus | β-diversity (Unifrac) | ||||||
| LCF: ↑Bifidobacterium | |||||||
| [61] | Very long chain inulin (10 g/d) | Randomized, DB, placebo-controlled crossover | 32 (18 W/14 M) | FISH | ↑Bifidobacterium | -- | ↑Bifidobacterium, Atopobium |
| Age: 25 y | Lactobacillus-Enterococcus | Lactobacillus-Enterococcus | |||||
| Duration: 3 wks | ↓Bacteroides-Prevotella | ||||||
| C: Maltodextrin (10 g/d) | BMI: 20–30 kg/m2 | ↔E.coli, E. rectale-C. coccoides, Ruminococcus | |||||
| Candidate Prebiotic Fibers | |||||||
| [62] | RMD (15 or 25 g/d) | Randomized, DB, placebo-controlled crossover | 49 (28 W/21 M) | qPCR | ↔ Bifidobacteria and total bacteria | No statistical analysis performed | (1) 15 g/d: ↔No significant changes |
| Duration: 3 wks | Age: 26 y | ||||||
| BMI: 21–28 kg/m2 | (2) 25 g/d: ↑Bifidobacterias | ||||||
| C: Maltodextrin | |||||||
| [63] | RPS (30 g/d) | Randomized, DB, placebo-controlled | 42 (24 W/18 M) | qPCR | ↑R. bromii, Bifidobacterium, | -- | ↑Bifidobacterium |
| Duration: 12 wks | Age: 42 y | 16s RNA gene Sequencing | ↓α-diversity | ||||
| B. ruminantium | (Shannon, Inverse Simpson) | ||||||
| C: Corn starch | BMI: No data | (V4 region, Illumina) | ↓R. obeum, R. torques, B. dentium | ||||
| [64] | XOS (8 g/d) | Randomized, DB, placebo-controlled crossover | 41 (20 W/21 M) | FISH | -- | -- | ↑Bifidobacterium |
| Duration: 3 wks | Age: 43 y | Flow cytometry | ↔Bacteroides/Prevotella, Clostridium I and II, Lactobacillus/Enterococcus Atopobium, B. lactis | ||||
| C: Maltodextrin | BMI: 20–30 kg/m2 | ||||||
| [65] | AXOS-enriched Bread (2.2 g/d) | Randomized, DB, placebo-controlled crossover | 40 (20 W/20 M) | FISH | ↑Bifidobacterium, Bacteroides | -- | -- |
| Duration: 21 d | Age: 31 y | ||||||
| Lactobacillus | |||||||
| C: Non-endoxylanase treated breads | BMI: 20.−26 kg/m2 | ||||||
| [66] | Polydextrose (PDX) | Randomized, DB, placebo-controlled crossover | 21 (0 W/21 M) | 16s RNA gene Pyrosequencing (V3-V4 regions, Roche) | -- | ↔α-diversity (Shannon, Chao1) | SCF, PDX: ↑Clostridiaceae, Veillonellaceae, Fecalibacterium, ↓Actinobacteria and Firmicutes |
| Soluble Corn Fiber (SCF) (21 g/d) | Age: 27 y | SCF vs. PDX: ↑Proteobacterias. Lactobacilli, Alcaligenaceae, Roseburia,↓Oscillospira, | |||||
| Duration: 21 d | BMI: 23–31 kg/m2 | ||||||
| ↔α-diversity (Shannon, Chao1) | |||||||
| C: No supplemental fiber (NFC) | PDX vs. SCF: ↑Verrucomicrobia, Clostridium, Akkermansia, | ||||||
| SCF: ↑Proteobacteria, Lactobacilli | |||||||
| PDX: ↑Clostridium, Akkermansia, | |||||||
| ↓Lachnospiraceae | |||||||
| C. leptum, ↓Hyphomicrobiaceae | |||||||
| Mixed Accepted and Candidates Prebiotic Fibers | |||||||
| [67] | (1) RPS (28–34 g/d) | Randomized, DB, placebo-controlled, parallel | 174 | qPCR | (1) RPS: ↑B. fecale/adolescentis/stercoris | No statistical analysis performed | -- |
| (2) RMS: ↑R. bromii | |||||||
| 16s RNA gene Sequencing | |||||||
| (3) Inulin: ↑Anaerostipes hadrus, | |||||||
| Age: 19 y | B. fecale/adolescentis, | ||||||
| B. longum/breve, | |||||||
| (2) RMS (20–24 g/d) | Gender and BMI: no data | (V4 region, Illumina) | B.catenulatum/pseudocatenulatum/ | ||||
| (3) Inulin (20 g/d) | kashiwanohense, B. bifidum, E. rectale | ||||||
| Duration: 2 wks | |||||||
| C: Amylase-accessible corn starch | |||||||
| [68] | (1) XOS (5 g/d) | Randomized, DB, placebo-controlled, parallel | 65 (33 W/26 M) | qPCR | XOS + inulin: ↑Lactobacillus | No statistical analysis performed | XOS + inulin/XOS: ↑Bifidobacterium (V2, V3), Peptostreptococcus (V2) |
| (2) XOS (1 g/d) + Inulin | ↔Firmicutes spp, Bacteroidetes spp, Clostridium, Staphylococcus, Eubacterium, Peptostreptococcus, Fusobacterium, Enterobacterium, F. prausnitzii, Roseburia spp. | ||||||
| Age: 18–24 y | |||||||
| (chicory) (3 g/d) | BMI: 18.5–27 kg/m2 | ||||||
| DP inulin: 10 | |||||||
| Duration: 4 wks | ↔Firmicutes spp, Bacteroidetes spp, Clostridium, Staphylococcus, Eubacterium, Fusobacterium, Enterobacterium, | ||||||
| C: Maltodextrin | |||||||
| F. prausnitzii, Roseburia spp. | |||||||
| Dietetic Fibers | |||||||
| [21] | Dietetic fiber (10 or 40 g/d) | Randomized, crossover | 19 (10 W/9 M) | qPCR | (1) 40 g/d: ↓E. coli | No statistical analysis performed | 40 g/d: ↑Microbial change (JSD metrics) in subjects with a low richness |
| 16s RNA gene Pyrosequencing (V3-V4 regions, Roche) | (2) 10, 40 g/d: ↔C. coccoides, C. leptum, Bacteroides-Prevotella, Bifidobacterium | ||||||
| Duration: 5 d | Age: 19–25 y | ||||||
| C: Comparison to basal point | BMI: 18.5–25 kg/m2 | ||||||
| [69] | (1) High Whole Grain (WG) Diet (>80 g/d WGs) | Randomized, crossover | 33 (21 W/12 M) | FISH | ↔No significant changes | ↔No significant changes | -- |
| Low consumers of WG diet | |||||||
| (2) Refined grain diet (<16 g/d WGs) | |||||||
| Age: 49 y | |||||||
| Duration: 6 wks | BMI: 20–35 kg/m2 | ||||||
| C: Comparison to basal point | |||||||
| [70] | Whole Grain Diet | Randomized, controlled, parallel | 81 (32 W/49 M) | 16s RNA gene Sequencing | ↔No significant changes | -- | ↑Lachnospira |
| (16 g fiber/1000 kcal) | Age: 54–55 y | (V4 region, Illumina) | ↓Enterobacteriaceae | ||||
| Duration: 6 wks | BMI: 20–35 kg/m2 | ↔α-diversity (phylogenetic tree), | |||||
| C: Refined grain diet | β-diversity (Unifrac) | ||||||
| (8 g fiber/1000 kcal) | |||||||
| b. Evidence from human studies relative to polyphenols effects on the gut microbiota composition of healthy adults | |||||||
| Ref. | Treatments | Study Type | Study Subjects | Analytic Technique | Results | Results | Results |
| Polyphenols vs. Control (C) | Differences vs. Basal Point | Differences between Treatment Groups | Differences vs. Control (C) | ||||
| [71,72] | Wild blueberry drink | Randomized, DB, placebo-controlled crossover | 15 (0 W/15 M) | qPCR | ↑Bifidobacterium | -- | -- |
| ↔Bacteroides, Prevotella, | |||||||
| Enterococcus, C. coccoides, | |||||||
| Bifidobacterium species | |||||||
| ↑B. longum subsp. infantis | |||||||
| (25 g/250 mL) | Age: 47 y | ||||||
| BMI: 22–28 kg/m2 | |||||||
| [Chlorogenic acid (127.5 mg) + anthocyanins (375 mg)] | |||||||
| Duration: 6 wks | |||||||
| C: Placebo drink | |||||||
| [73] | Boysenberry juice | Randomized, placebo-controlled crossover | 24 (5 M/20 W) | qPCR | -- | ↔Bacteroides-Prevotella- | ↔Bacteroides-Prevotella- |
| (anthocyanins, ellagitannins and ellagic acid derivatives; 750 mg) | Age: 50 y | Porphyromonas group, | |||||
| Porphyromonas group, | Bifidobacterium, | ||||||
| C. perfringens, Lactobacillus | |||||||
| BMI: 18–35 kg/m2 | Bifidobacterium, | ||||||
| C. perfringens, Lactobacillus | |||||||
| Duration: 4 wks | |||||||
| C: Placebo drink | |||||||
| [74] | Fruits and Vegetables (2 (6 wks), 4 (12 wk) and 6 portions (18 wks)) | Randomized, controlled, parallel | 122 (74 W/48 M) | FISH | (1) HF: ↑Bacteroides/Prevotella | No statistical analysis performed | LF: ↑Bifidobacterium |
| Age: 49–52 y | (2) LF: ↑Bifidobacterium, | ||||||
| BMI: 18–35 kg/m2 | Bacteroides/Prevotella, | ||||||
| Duration: 18 wks | High-flavonoid (HF)/Low-flavonoid (LF) | C. leptum-R. bromii/flavefaciens | |||||
| C: Habitual diet | |||||||
| [75] | Cocoa flavanols | Randomized, DB, crossover | 22 (12 M/10 W) | FISH | (1) HCF: ↑Bifidobacterium, Lactobacillus, Enterococcus | -- | HCF: ↑Bifidobacterium, Lactobacillus, Enterococcus |
| (catechin, epicatechin, theobromine) HCF: High–cocoa flavanol group; 494 mg/d | |||||||
| Age: 30 y | |||||||
| Duration: 4 wks | BMI: 20–25 kg/m2 | ↓C. histolyticum | |||||
| C: Low–cocoa flavanol (LCF) group (23 mg/d) | (2) LCF: ↑E. rectale-C. coccoides group, C. histolyticum | ||||||
| ↓C. histolyticum | |||||||
| [76] | Green tea [400 mL/d (100.2 μg gallic acid Eq/mL)] | Intervention | 12 (4 W/8 M) | 16s RNA gene Sequencing | ↑α-diversity (Simpson, Shannon and Chao1), ↑Actinobacteria, Firmicutes, | -- | -- |
| Age: 34 y | |||||||
| (V4-V5 regions, Illumina) | |||||||
| Butyrate-producing bacteria, | |||||||
| BMI: 18–24 kg/m2 | ↓Bacteroidetes members | ||||||
| Duration: 2 wks | |||||||
| C: Comparison to basal point | |||||||
| [77] | Green Tea | Randomized, single blind, placebo-controlled parallel | 58 (46 W/12 M) | 16S–23S rDNA Intergenic spacer region | ↔α-diversity (Shannon), Actinobacteria, Firmicutes, | -- | ↔α-diversity (Shannon), Actinobacteria, Firmicutes, |
| (>1.35 g Catechins; >0.56 g Epigallocatechin-3-gallate) | Age: 29 y | ||||||
| Bacteroidetes, Fusobacteria, Verrucomicrobia, Proteobacteria | |||||||
| Bacteroidetes, Fusobacteria, Verrucomicrobia, Proteobacteria | |||||||
| BMI: 18–25 kg/m2 | |||||||
| (9 capsules/d) | |||||||
| Duration: 12 wks | |||||||
| C: Microcrystalline cellulose | |||||||
| Ref. | Treatments | Study Type | Study Subjects | Analytic Technique | Results | Results | Results |
|---|---|---|---|---|---|---|---|
| Alcohol vs. Control (C) | Differences vs. Basal Point | Differences between Treatment Groups | Differences vs. Control (C) | ||||
| [78] | Red wine (100 mL/d) | Observational | 38 (27 W/11 M) | qPCR | -- | -- | ↓Bifidobacterium, |
| Age: 55–67 y | B. coccoides, C. leptum, Lactobacillus | ||||||
| C: Non-wine consumers | BMI: 22–30 kg/m2 | ||||||
| [79] | Vodka (2 mL in 300 mL orange or strawberry juice) | Observational | 15 (4 W/11 M) | 16s RNA gene Sequencing | ↔α-diversity (Chao1), β-diversity (Bray-Curtis) | ||
| ↔ Main phyla, families, genera and species analyzed | |||||||
| Age: 26 y | (V1-V2 region, Illumina) | ||||||
| BMI: 23–27 kg/m2 | |||||||
| C: Comparison to basal point | |||||||
| [80] | (1) RW (Red wine; 272 mL/d) | Randomized, controlled, crossover | 10 (0 W/10 M) | PCR | (1) RW: ↑Enterococcus, Prevotella, Bacteroides, B. uniformis, Bifidobacterium, | Gin vs. RW and DRW: | -- |
| Age: 48 y | DGGE + qPCR | ||||||
| E. lenta, B. cocoides-E. rectale group | ↑Clostridium, | ||||||
| (2) DRW: ↑Enterococcus, Bifidobacterium, E. lenta, | C. histolyticum | ||||||
| ↓Prevotella, Bifidobacterium, Enterococcus, E. lenta | |||||||
| (2) DRW (Dealcoholized red wine; 272 mL/d) | BMI: 24.6–30.8 kg/m2 | B. cocoides-E. rectale group | |||||
| (3) Gin (100 mL/d) | |||||||
| Duration: 20 d | |||||||
| C: Comparison to basal point | |||||||
| [81] | Red wine | Randomized, controlled, parallel | 20 | 16s RNA gene Sequencing | ↑α-diversity (Shannon-Weaver), Slackia, Gordonibacter, Oscillatoria, Veillonela | -- | -- |
| (272 mL/d) | Age: 20–48 y | ||||||
| Duration: 1 month | Gender and BMI: No data | (V1-V2 regions, Illumina) | |||||
| C: Non-wine consumers | |||||||
| [82] | (1) AB: Alcoholic beer | Interventional, 2 phases study, parallel | NAB: 35 (14 W/21 M) | 16s RNA gene Sequencing | (1) AB: ↑Bacteroidetes, Dysgonomonas, Pseudomonas, Succinivibrio | No statistical analysis performed | -- |
| (355 mL/d) | Age: 21–53 y | ↓Firmicutes | |||||
| (2) NAB: Non-alcoholic beer | (2) NBA: ↑α-diversity (Chao1, Shannon), β-diversity (Unifrac), | ||||||
| AB: 33 (15 W/18 M) | |||||||
| (V3 region, Roche) | ↑Bacteroidetes, Dialister, Actinomyces, Staphylococcus, Parabacteroides, Veillonella, Haemophilus, Lactococcus, Bacteroides, Weissella, Phascolarctobacterium, Streptococcus, Acinetobacter, Sutterella, Turicibacter, Lactobacillus | ||||||
| (355 mL/d) | Age: 21–55 y | ||||||
| Duration: 30 d | |||||||
| C: Comparison to basal point | BMI: No data | ||||||
| ↓Firmicutes |
| Ref. | Treatments Sweeteners vs Control (C) | Study type | Study subjects | Analytic technique | Results Differences vs. basal point | Results Differences vs. Control (C) |
|---|---|---|---|---|---|---|
| [83] | Isomalt Dose: 30 g/d Duration: 4 wk C: Sucrose | Randomized, DB, placebo-controlled, crossover | 19 (12 W/7 M) Age: 35 y BMI: 23-26 kg/m2 | Culture FISH | -- | ↑Bifidobacteria ↑Atopobium, Actinobacteria ↓Roseburia intestinalis, Bacteroides |
| [84] | Lactitol Dose: 0, 5 or 10 g/d Duration: 1 wk C: Sucrose | Randomized, DB, placebo-controlled, longitudinal | 75 (26 W/39 M) Age: 18-24 y BMI Women: 20-25 kg/m2 BMI Men: 20-26 kg/m2 | Culture | (1) 5 g: ↔No significant changes (2) 10 g: ↑Bifidobacterium ↔Bacterial counts of total anaerobes, aerobes, Enterobacteriaceae, Lactobacilli | -- |
| [85] | (1) Aspartame (2) Acesulfame-K Data collection: 24 h recalls C: Non-consumers | Observational | 31 (20 W/11 M) Age: 27 y BMI: 20-28 kg/m2 | LH-PCR | -- | ↑β-diversity (Unifrac) ↔No significant changes at class and order level |
| [86] | Non-caloric artificial sweeteners (NAS) Data collection: FFQ | Observational | 172 Age: 43 y Gender and BMI: No data | 16sRNA gene Sequencing (V2 region, Illumina) | Positive correlations with Actinobacteria, Enterobacteriaceae, Deltaproteobacteria | -- |
| ccharin Dose: 120 mg/d Duration: 6 d C: Comparison to basal point | Intervention | 7 (2 W/5 M) Age: 28-36 y BMI: No data | 16sRNA gene Sequencing (V2 region, Illumina) | Different bacteria clustering between NAS groups (1) NAS Responders: ↑Lactobacillales, Bacteroidales ↓Clostridiales (2) NAS non-Responders: ↔No significant changes | -- |
| Ref. | Treatments | Study Type | Study Subjects | Analytic Technique | Results | Results | Results |
|---|---|---|---|---|---|---|---|
| Fats vs. Control (C) | Differences vs. Basal Point | Differences between Treatment Groups | Differences vs. Control (C) | ||||
| [87] | Soybean Oil diet | Randomized, parallel | 217 (114 W/103 M) | 16s RNA gene Sequencing | (1) Low fat: ↑Blautia, Fecalibacterium | (1) Low vs. High: ↑α-diversity (Shannon index) | -- |
| Low (Fat: 20% total energy), Medium (30%) and High (40%) | |||||||
| Age: 23 y | (V3-V4 regions, Illumina) | (2) Medium fat: ↑Bacteroidetes | (2) High vs. Low: ↑Bacteroidetes, Alistipes, Bacteroides | ||||
| BMI: 19–24 kg/m2 | |||||||
| (3) High fat: ↑Bacteroidetes, Alistipes, Bacteroides | |||||||
| Duration: 6 months | ↓Firmicutes, Blautia, Fecalibacterium | ||||||
| C:Comparison to basal point | ↓Firmicutes, Fecalibacterium | ||||||
| [88] | Saturated fat (dairy or butter) | Randomized, controlled, parallel | 109 (No gender data) | 16s RNA gene Sequencing | -- | -- | ↔α-diversity (Shannon), |
| (15% total energy) | β-diversity (Unifrac) | ||||||
| (V4 region, Illumina) | Changes in 57 bacterial genus | ||||||
| Duration: 4 wks | Age: 21–65 y | ||||||
| C: Low fat diet (7% saturated fat) | BMI: 18–36 kg/m2 | ||||||
| [89] | Omega-3 Drink (D) or capsules (C) (2000 mg/d DHA + 2000 mg/d EPA) | Randomized, DB, crossover | 22 (12 W/10 M) | 16s RNA gene Sequencing | D and C: ↔α-diversity (Shannon index), β-diversity (Unifrac) | D: ↑Lachnospira, Roseburia | -- |
| Age: 51–65 y | |||||||
| BMI: 22–34 kg/m2 | (V4 region, Illumina) | ||||||
| Duration: 8 wks | ↑Clostridiaceae, Sutterellaceae, Akkermansiaceae, Oscillospira, Lachnospira, Bifidobacterium, Lactobacillus | ||||||
| C: No control group. | |||||||
| Comparison to basal point | |||||||
| ↓Coprococcus, Fecalibacterium | |||||||
| [90] | Dairy Cream (48% SFA) (341 mL/d) | Randomized, parallel | 25 (0 W/25 M) | 16s RNA gene Sequencing | ↔α-diversity (Shannon), | -- | -- |
| Age: 23 y | β-diversity (Unifrac) | ||||||
| Duration: 1 wk | BMI: 21–25 kg/m2 | (V4 region, Illumina) | ↑Betaproteobacterias, ↓Bacteroidaceae | ||||
| C: No control group. | |||||||
| Comparison to basal point | |||||||
| [91] | (1) Semi-skimmed ewe’s milk yogurt (ES)(2.8% fat) (250 g/d) | Randomized, DB, crossover | 30 (16 W/14 M) | qPCR | -- | (1) CW vs. ES: ↑C. leptum group | -- |
| Age: 42 y | |||||||
| (2) Whole ewe’s milk yogurt | BMI: 19–28 kg/m2 | ↔Bacteroides, F. prausnitzii, | |||||
| (EW) (5.8% fat) (250 g/d) | Bifidobacterium spp., Lactobacillus spp., Enterobacteriaceae, | ||||||
| Duration: 5 wks | |||||||
| C: Cow whole’s milk yogurt (CW) (3.0% fat) (250 g/d) | Enterococcus spp | ||||||
| (2) Women (Highest ratio of total cholesterol/HDL–cho): EW vs. ES: | |||||||
| ↓B. coccoides-E. rectale |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Redondo-Useros, N.; Nova, E.; González-Zancada, N.; Díaz, L.E.; Gómez-Martínez, S.; Marcos, A. Microbiota and Lifestyle: A Special Focus on Diet. Nutrients 2020, 12, 1776. https://doi.org/10.3390/nu12061776
Redondo-Useros N, Nova E, González-Zancada N, Díaz LE, Gómez-Martínez S, Marcos A. Microbiota and Lifestyle: A Special Focus on Diet. Nutrients. 2020; 12(6):1776. https://doi.org/10.3390/nu12061776
Chicago/Turabian StyleRedondo-Useros, Noemí, Esther Nova, Natalia González-Zancada, Ligia E. Díaz, Sonia Gómez-Martínez, and Ascensión Marcos. 2020. "Microbiota and Lifestyle: A Special Focus on Diet" Nutrients 12, no. 6: 1776. https://doi.org/10.3390/nu12061776
APA StyleRedondo-Useros, N., Nova, E., González-Zancada, N., Díaz, L. E., Gómez-Martínez, S., & Marcos, A. (2020). Microbiota and Lifestyle: A Special Focus on Diet. Nutrients, 12(6), 1776. https://doi.org/10.3390/nu12061776