Vitamin K Nutrition and Bone Health

Abstract
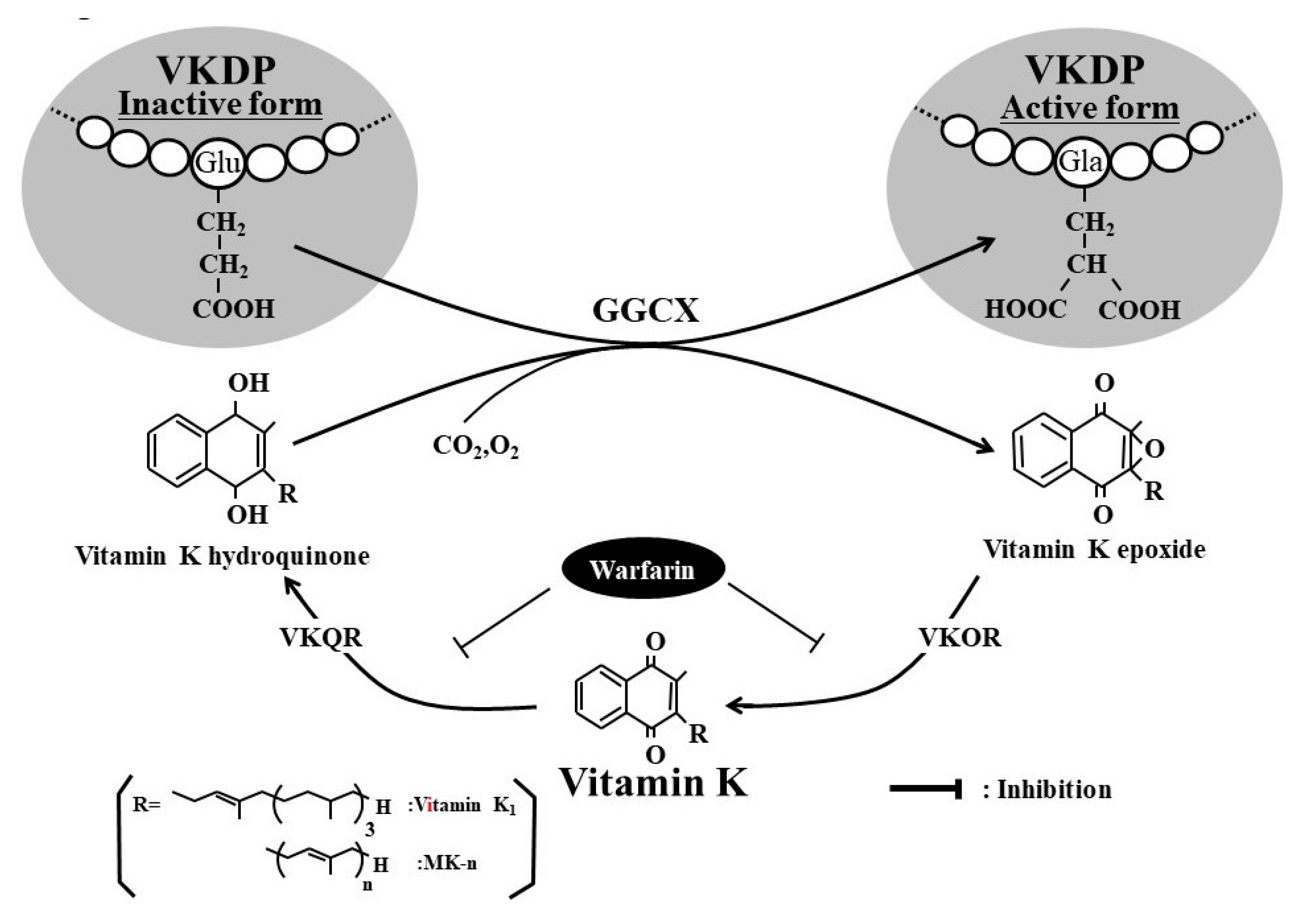
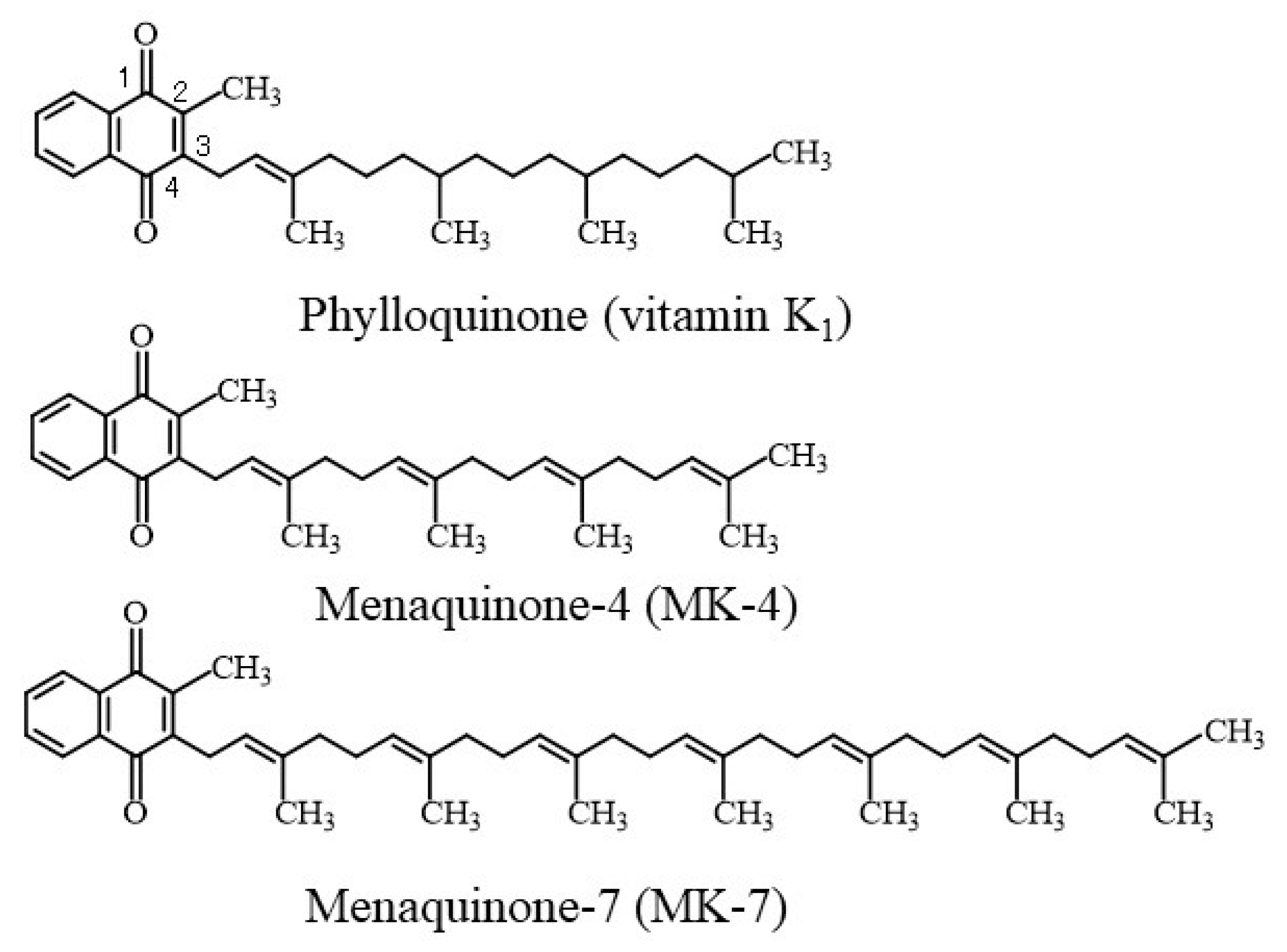
:1. Introduction
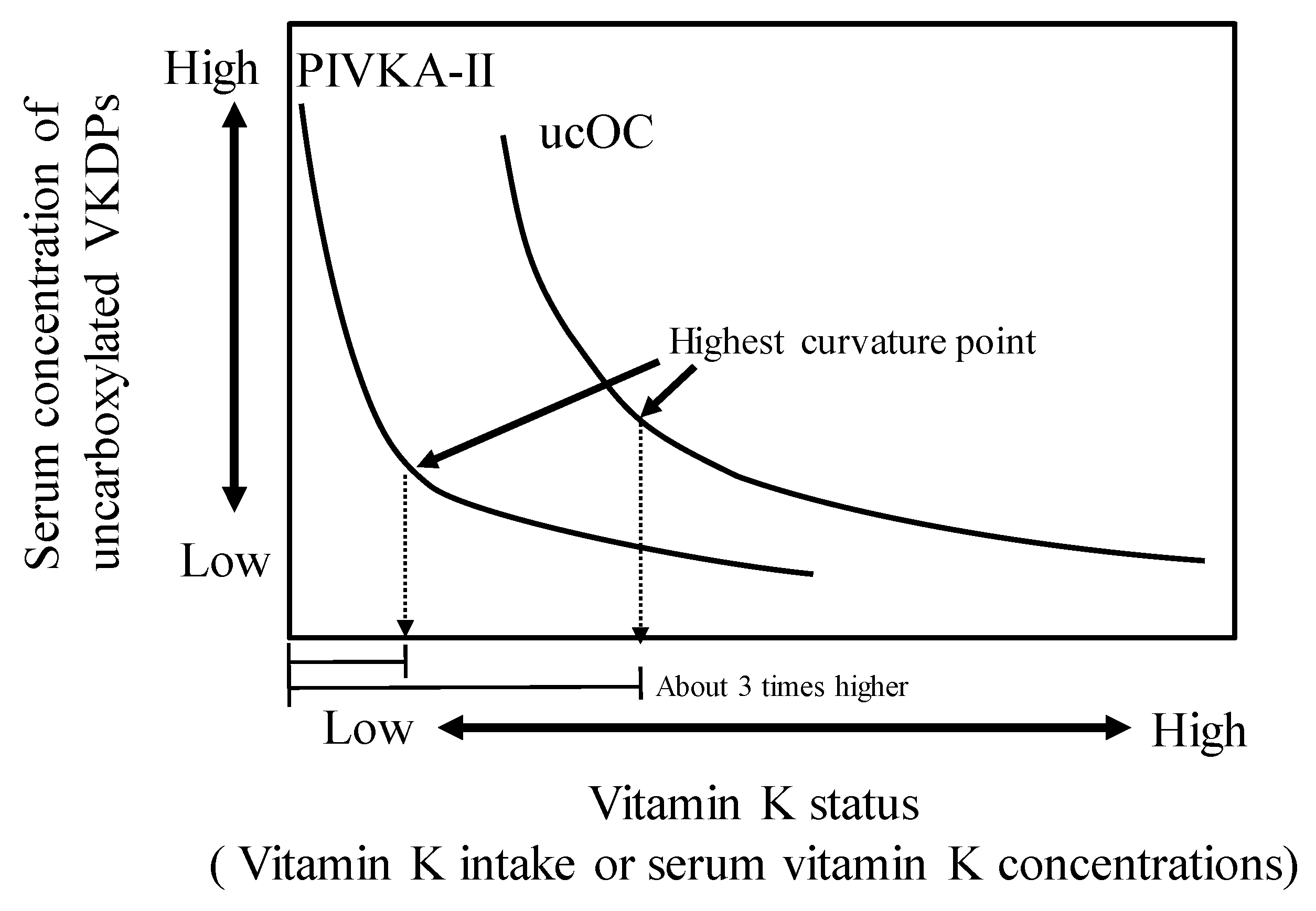
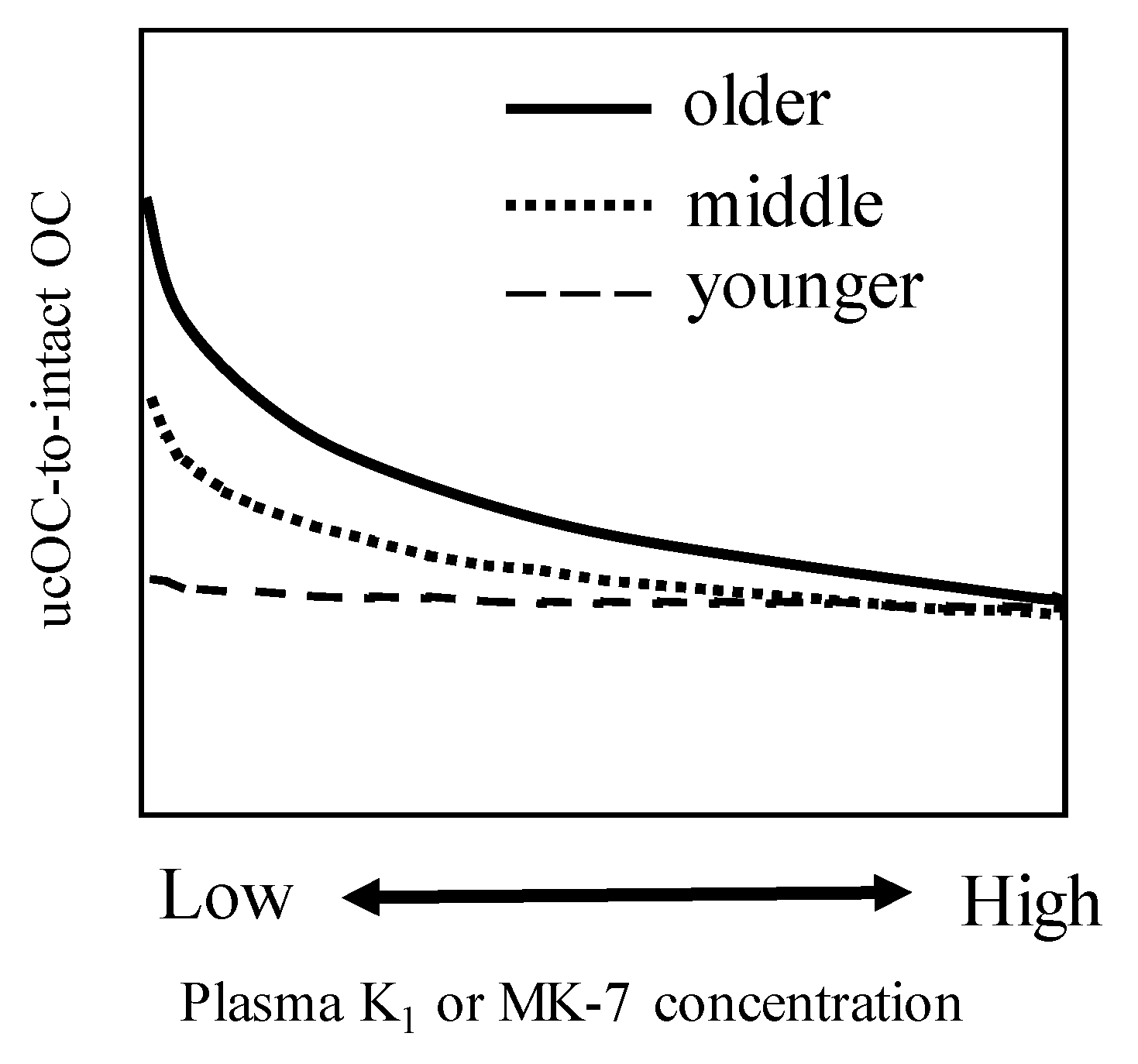
2. Negative Correlation between Vitamin K Status and Uncarboxylated VKDPs including OC
3. Recent Topics on the Impact of Vitamin K Deficiency on Bone Health
3.1. Vitamin K and cOC Deficiency May Predispose to Diabetes-Related Bone Damage
3.2. Vitamin K Deficiency Is Associated with Bone Fractures
4. Effect of Vitamin K Supplementation on Bone Turnover Rate, Bone Mineral Density, and Fractures
5. Possible Implications of Combined Vitamin D and K Supplementation on Bone Quality
6. The Role of MGP (Matrix Gla Protein) in Bone Health
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- NIH Consensus Development Panel on Osteoporosis Prevention, Diagnosis, and Therapy. Osteoporosis prevention, diagnosis, and therapy. JAMA 2001, 285, 785–795. [Google Scholar] [CrossRef] [PubMed]
- Kuroda, T.; Uenishi, K.; Ohta, H.; Shiraki, M. Multiple vitamin deficiencies additively increase the risk of incident fractures in Japanese postmenopausal women. Osteoporos. Int. 2019, 30, 593–599. [Google Scholar] [CrossRef] [PubMed]
- Ichikawa, T.; Horie-Inoue, K.; Ikeda, K.; Blumberg, B.; Inoue, S. Steroid and xenobiotic receptor SXR mediates vitamin K2-activated transcription of extracellular matrix-related genes and collagen accumulation in osteoblastic cells. J. Biol. Chem. 2006, 281, 16927–16934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ducy, P.; Desbois, C.; Boyce, B.; Pinero, G.; Story, B.; Dunstan, C.; Smith, E.; Bonadio, J.; Goldstein, S.; Gundberg, C.; et al. Increased bone formation in osteocalcin-deficient mice. Nature 1996, 382, 448–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamao, M.; Suhara, Y.; Tsugawa, N.; Uwano, M.; Yamaguchi, N.; Uenishi, K.; Ishida, H.; Sasaki, S.; Okano, T. Vitamin K content of foods and dietary vitamin K intake in Japanese young women. J. Nutr. Sci. Vitaminol. 2007, 53, 464–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamao, M.; Suhara, Y.; Tsugawa, N.; Okano, T. Determination of plasma vitamin K by high-performance liquid chromatography with fluorescence detection using vitamin K analogs as internal standards. J. Chromatogr. B 2005, 816, 41–48. [Google Scholar] [CrossRef]
- Suhara, Y.; Kamao, M.; Tsugawa, N.; Okano, T. Method for the determination of vitamin K homologues in human plasma using high-performance liquid chromatography-tandem mass spectrometry. Anal. Chem. 2005, 77, 757–763. [Google Scholar] [CrossRef]
- Gentili, A.; Cafolla, A.; Gasperi, T.; Bellante, S.; Caretti, F.; Curini, R.; Fernández, V.P. Rapid, high performance method for the determination of vitamin K1, menaquinone-4 and vitamin K1 2,3-epoxide in human serum and plasma using liquid chromatography-hybrid quadrupole linear ion trap mass spectrometry. J. Chromatogr. A 2014, 1338, 102–110. [Google Scholar] [CrossRef]
- Karl, J.P.; Fu, X.; Dolnikowski, G.G.; Saltzman, E.; Booth, S.L. Quantification of phylloquinone and menaquinones in feces, serum, and food by high-performance liquid chromatography-mass spectrometry. J. Chromatogr. B 2014, 963, 128–133. [Google Scholar] [CrossRef]
- Riphagen, I.J.; Van Der Molen, J.C.; Van Faassen, M.; Navis, G.; De Borst, M.H.; Muskiet, F.A.; De Jong, W.H.; Bakker, S.J.; Kema, I.P. Measurement of Plasma Vitamin K1 (Phylloquinone) and K2 (menaquinones-4 and -7) Using HPLC-tandem Mass Spectrometry. Clin. Chem. Lab. Med. 2016, 54, 201–210. [Google Scholar] [CrossRef]
- Fusaro, M.; Gallieni, M.; Rizzo, M.A.; Stucchi, A.; Delanaye, P.; Cavalier, E.; Moysés, R.M.A.; Jorgetti, V.; Iervasi, G.; Giannini, S.; et al. Vitamin K plasma levels determination in human health. Clin. Chem. Lab. Med. 2017, 55, 789–799. [Google Scholar] [CrossRef] [PubMed]
- Nakano, T.; Tsugawa, N.; Kuwabara, A.; Kamao, M.; Tanaka, K.; Okano, T. High prevalence of hypovitaminosis D and K in patients with hip fracture. Asia Pac. J. Clin. Nutr. 2011, 20, 56–61. [Google Scholar] [PubMed]
- Kuwabara, A.; Himeno, M.; Tsugawa, N.; Kamao, M.; Fujii, M.; Kawai, N.; Fukuda, M.; Ogawa, Y.; Kido, S.; Okano, T.; et al. Hypovitaminosis D and K are highly prevalent and independent of overall malnutrition in the institutionalized elderly. Asia Pac. J. Clin. Nutr. 2010, 19, 49–56. [Google Scholar] [PubMed]
- Martini, L.A.; Booth, S.L.; Saltzman, E.; Oliveira do Rosário Dias de Oliveira Latorre, M.; Wood, R.J. Dietary phylloquinone depletion and repletion in postmenopausal women: Effects on bone and mineral metabolism. Osteoporos. Int. 2006, 17, 929–935. [Google Scholar] [CrossRef]
- Tsugawa, N.; Shiraki, M.; Suhara, Y.; Kamao, M.; Tanaka, K.; Okano, T. Vitamin K status of healthy Japanese women: Age-related vitamin K requirement for gamma-carboxylation of osteocalcin. Am. J. Clin. Nutr. 2006, 83, 380–386. [Google Scholar] [CrossRef]
- Beavan, S.R.; Prentice, A.; Stirling, D.M.; Dibba, B.; Yan, L.; Harrington, D.J.; Shearer, M.J. Ethnic differences in osteocalcin gamma-carboxylation, plasma phylloquinone (vitamin K1) and apolipoprotein E genotype. Eur. J. Clin. Nutr. 2005, 59, 72–81. [Google Scholar] [CrossRef] [Green Version]
- Booth, S.L.; Broe, K.E.; Peterson, J.W.; Cheng, D.M.; Dawson-Hughes, B.; Gundberg, C.M.; Cupples, L.A.; Wilson, P.W.; Kiel, D.P. Associations between vitamin K biochemical measures and bone mineral density in men and women. J. Clin. Endocrinol. Metab. 2004, 89, 4904–4909. [Google Scholar] [CrossRef] [Green Version]
- Binkley, N.C.; Krueger, D.C.; Kawahara, T.N.; Engelke, J.A.; Chappell, R.J.; Suttie, J.W. A high phylloquinone intake is required to achieve maximal osteocalcin gamma-carboxylation. Am. J. Clin. Nutr. 2002, 76, 1055–1060. [Google Scholar] [CrossRef]
- Kaneki, M.; Hodges, S.J.; Hosoi, T.; Fujiwara, S.; Lyons, A.; Crean, S.J.; Ishida, N.; Nakagawa, M.; Takechi, M.; Sano, Y.; et al. Japanese fermented soybean food as the major determinant of the large geographic difference in circulating levels of vitamin K2: Possible implications for hip-fracture risk. Nutrition 2001, 17, 315–321. [Google Scholar] [CrossRef]
- Binkley, N.C.; Krueger, D.C.; Engelke, J.A.; Foley, A.L.; Suttie, J.W. Vitamin K supplementation reduces serum concentrations of under-gamma-carboxylated osteocalcin in healthy young and elderly adults. Am. J. Clin. Nutr. 2000, 72, 1523–1528. [Google Scholar] [CrossRef] [Green Version]
- Hodges, S.J.; Akesson, K.; Vergnaud, P.; Obrant, K.; Delmas, P.D. Circulating levels of vitamins K1 and K2 decreased in elderly women with hip fracture. J. Bone Miner. Res. 1993, 8, 1241–1245. [Google Scholar] [CrossRef] [PubMed]
- Fusaro, M.; Noale, M.; Viola, V.; Galli, F.; Tripepi, G.; Vajente, N.; Plebani, M.; Zaninotto, M.; Guglielmi, G.; Miotto, D.; et al. Vitamin K, Vertebral Fractures, Vascular Calcifications, and Mortality: VItamin K Italian (VIKI) Dialysis Study. J. Bone Miner. Res. 2012, 27, 2271–2778. [Google Scholar] [CrossRef] [PubMed]
- Holden, R.M.; Morton, A.R.; Garland, J.S.; Pavlov, A.; Day, A.G.; Booth, S.L. Vitamins K and D status in stages 3-5 chronic kidney disease. Clin. J. Am. Soc. Nephrol. 2010, 5, 590–597. [Google Scholar] [CrossRef] [PubMed]
- Pilkey, R.M.; Morton, A.R.; Boffa, M.B.; Noordhof, C.; Day, A.G.; Su, Y.; Miller, L.M.; Koschinsky, M.L.; Booth, S.L. Subclinical vitamin K deficiency in hemodialysis patients. Am. J. Kidney Dis. 2007, 49, 432–439. [Google Scholar] [CrossRef]
- Holden, R.M.; Iliescu, E.; Morton, A.R.; Booth, S.L. Vitamin K status of Canadian peritoneal dialysis patients. Perit. Dial. Int. 2008, 28, 415–418. [Google Scholar] [CrossRef]
- Sokoll, L.J.; Sadowski, J.A. Comparison of biochemical indexes for assessing vitamin K nutritional status in a healthy adult population. Am. J. Clin. Nutr. 1996, 63, 566–573. [Google Scholar] [CrossRef] [Green Version]
- Booth, S.L.; Martini, L.; Peterson, J.W.; Saltzman, E.; Dallal, G.E.; Wood, R.J. Dietary phylloquinone depletion and repletion in older women. J. Nutr. 2003, 133, 2565–2569. [Google Scholar] [CrossRef] [Green Version]
- Kuwabara, A.; Fujii, M.; Kawai, N.; Tozawa, K.; Kido, S.; Tanaka, K. Bone is more susceptible to vitamin K deficiency than liver in the institutionalized elderly. Asia Pac. J. Clin. Nutr. 2011, 20, 50–55. [Google Scholar]
- Tsugawa, N.; Uenishi, K.; Ishida, H.; Minekami, T.; Doi, A.; Koike, S.; Takase, T.; Kamao, M.; Mimura, Y.; Okano, T. A novel method based on curvature analysis for estimating the dietary vitamin K requirement in adolescents. Clin. Nutr. 2012, 31, 255–260. [Google Scholar] [CrossRef]
- Westenfeld, R.; Krueger, T.; Schlieper, G.; Cranenburg, E.C.; Magdeleyns, E.J.; Heidenreich, S.; Holzmann, S.; Vermeer, C.; Jahnen-Dechent, W.; Ketteler, M.; et al. Effect of vitamin K2 supplementation on functional vitamin K deficiency in hemodialysis patients: A randomized trial. Am. J. Kidney Dis. 2012, 59, 186–195. [Google Scholar] [CrossRef]
- Schurgers, L.J.; Teunissen, K.J.; Hamulyák, K.; Knapen, M.H.; Vik, H.; Vermeer, C. Vitamin K-containing dietary supplements: Comparison of synthetic vitamin K1 and natto-derived menaquinone-7. Blood 2007, 109, 3279–3283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rennenberg, R.J.; De Leeuw, P.W.; Kessels, A.G.; Schurgers, L.J.; Vermeer, C.; Van Engelshoven, J.M.; Kemerink, G.J.; Kroon, A.A. Calcium scores and matrix Gla protein levels: Association with vitamin K status. Eur. J. Clin. Investig. 2010, 40, 344–349. [Google Scholar] [CrossRef] [PubMed]
- Tsugawa, N.; Kanamori, K.; Sawada, N.; Takase, T.; Nakayama, C.; Fujita, T.; Inoue, R.; Kingetu, M.; Uchino, Y.; Shiraki, M.; et al. Development of assay method for plasma concentration of undercarboxylated matrix Gla protein (ucMGP), and its usefulness in estimation of vascular vitamin K status (in Japanese). Vitamins 2014, 88, 139–144. [Google Scholar]
- Schurgers, L.J.; Cranenburg, E.C.; Vermeer, C. Matrix Gla-protein: The calcification inhibitor in need of vitamin K. Thromb. Haemost. 2008, 100, 593–603. [Google Scholar]
- Shea, M.K.; O’Donnell, C.J.; Vermeer, C.; Magdeleyns, E.J.; Crosier, M.D.; Gundberg, C.M.; Ordovas, J.M.; Kritchevsky, S.B.; Booth, S.L. Circulating uncarboxylated matrix Gla protein is associated with vitamin K nutritional status, but not coronary artery calcium, in older adults. J. Nutr. 2011, 141, 1529–1534. [Google Scholar] [CrossRef]
- Dalmeijer, G.W.; Van Der Schouw, Y.T.; Vermeer, C.; Magdeleyns, E.J.; Schurgers, L.J.; Beulens, J.W. Circulating matrix Gla protein is associated with coronary artery calcification and vitamin K status in healthy women. J. Nutr. Biochem. 2013, 24, 624–628. [Google Scholar] [CrossRef]
- Suttie, J.W.; Mummah-Schendel, L.L.; Shah, D.V.; Lyle, B.J.; Greger, J.L. Vitamin K deficiency from dietary vitamin K restriction in humans. Am. J. Clin. Nutr. 1988, 47, 475–480. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Yu, Y. Aortic calcification was associated with risk of fractures: A meta-analysis. J. Back Musculoskelet. Rehabil. 2016, 29, 635–642. [Google Scholar] [CrossRef]
- Campos-Obando, N.; Kavousi, M.; Van Lennep, J.E.; Rivadeneira, F.; Hofman, A.; Uitterlinden, A.G.; Franco, O.H.; Zillikens, M.C. Bone health and coronary artery calcification: The Rotterdam Study. Atherosclerosis 2015, 241, 278–283. [Google Scholar] [CrossRef] [Green Version]
- Sakamoto, N.; Wakabayashi, J.; Sakamoto, K. Low vitamin K intake effects on glucose tolerance in rats. Int. J. Vitam. Nutr. Res. 1999, 69, 27–31. [Google Scholar] [CrossRef]
- Sakamoto, N.; Nishiike, T.; Iguchi, H.; Sakamoto, K. Possible effects of one week vitamin K (menaquinone-4) tablets intake on glucose tolerance in healthy young male volunteers with different descarboxy prothrombin levels. Clin. Nutr. 2000, 19, 259–263. [Google Scholar] [CrossRef] [PubMed]
- Shiraki, M.; Itabashi, A. Short-term menatetrenone therapy increases gamma-carboxylation of osteocalcin with a moderate increase of bone turnover in postmenopausal osteoporosis: A randomized prospective study. J. Bone Miner. Metab. 2009, 27, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.K.; Sowa, H.; Hinoi, E.; Ferron, M.; Ahn, J.D.; Confavreux, C.; Dacquin, R.; Mee, P.J.; McKee, M.D.; Jung, D.Y.; et al. Endocrine regulation of energy metabolism by the skeleton. Cell 2007, 130, 456–469. [Google Scholar] [CrossRef] [Green Version]
- Urano, T.; Shiraki, M.; Kuroda, T.; Tanaka, S.; Urano, F.; Uenishi, K.; Inoue, S. Low serum osteocalcin concentration is associated with incident type 2 diabetes mellitus in Japanese women. J. Bone Miner. Metab. 2018, 36, 470–477. [Google Scholar] [CrossRef]
- Liu, D.M.; Guo, X.Z.; Tong, H.J.; Tao, B.; Sun, L.H.; Zhao, H.Y.; Ning, G.; Liu, J.M. Association between osteocalcin and glucose metabolism: A meta-analysis. Osteoporos. Int. 2015, 26, 2823–2833. [Google Scholar] [CrossRef]
- Pi, M.; Wu, Y.; Quarles, L.D. GPRC6A mediates responses to osteocalcin in β-cells in vitro and pancreas in vivo. J. Bone Miner. Res. 2011, 26, 1680–1683. [Google Scholar] [CrossRef] [Green Version]
- Saito, M.; Marumo, K. Collagencross-links as a determinant bone quality: A possible explanation for bone fragility in ageing, osteoporosis, and diabetes mellitus. Osteoporos. Int. 2010, 21, 195–214. [Google Scholar] [CrossRef]
- Shiraki, M.; Kuroda, T.; Tanaka, S.; Saito, M.; Fukunaga, M.; Nakamura, T. Nonenzymatic collagen cross-links induced by glycoxidation (pentosidine) predicts vertebral fractures. J. Bone Miner. Metab. 2008, 26, 93–100. [Google Scholar] [CrossRef]
- Hart, J.P.; Shearer, M.J.; Klenerman, L.; Catterall, A.; Reeve, J.; Sambrook, P.N.; Dodds, R.A.; Bitensky, L.; Chayen, J. Electrochemical detection of depressed circulating levels of vitamin K1 in osteoporosis. J. Clin. Endocrinol. Metab. 1985, 60, 1268–1269. [Google Scholar] [CrossRef] [Green Version]
- Feskanich, D.; Weber, P.; Willett, W.C.; Rockett, H.; Booth, S.L.; Colditz, G.A. Vitamin K intake and hip fractures in women: A prospective study. Am. J. Clin. Nutr. 1999, 69, 74–79. [Google Scholar] [CrossRef] [Green Version]
- Booth, S.L.; Tucker, K.L.; Chen, H.; Hannan, M.T.; Gagnon, D.R.; Cupples, L.A.; Wilson, P.W.; Ordovas, J.; Schaefer, E.J.; Dawson-Hughes, B.; et al. Dietary vitamin K intakes are associated with hip fracture but not with bone mineral density in elderly men and women. Am. J. Clin. Nutr. 2000, 71, 1201–1208. [Google Scholar] [CrossRef] [PubMed]
- Hao, G.; Zhang, B.; Gu, M.; Chen, C.; Zhang, Q.; Zhang, G.; Cao, X. Vitamin K intake and the risk of fractures: A meta-analysis. Medicine 2017, 96, e6725. [Google Scholar] [CrossRef] [PubMed]
- Hodges, S.J.; Pilkington, M.J.; Stamp, T.C.; Catterall, A.; Shearer, M.J.; Bitensky, L.; Chayen, J. Depressed levels of circulating menaquinones in patients with osteoporotic fractures of the spine and femoral neck. Bone 1991, 12, 387–389. [Google Scholar] [CrossRef]
- Tsugawa, N.; Shiraki, M.; Suhara, Y.; Kamao, M.; Ozaki, R.; Tanaka, K.; Okano, T. Low plasma phylloquinone concentration is associated with high incidence of vertebral fracture in Japanese women. J. Bone Miner. Metab. 2008, 26, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Kawana, K.; Takahashi, M.; Hoshino, H.; Kushida, K. Circulating levels of vitamin K1, menaquinone-4, and menaquinone-7 in healthy elderly Japanese women and patients with vertebral fractures and patients with hip fractures. Endocr. Res. 2001, 27, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Szulc, P.; Chapuy, M.C.; Meunier, P.I.; Delmas, P.D. Serum undercarboxylated osteocalcin is a marker of the risk of hip fracture in elderly women. J. Clin. Investig. 1993, 91, 1769–1774. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, S.; Narusawa, K.; Onishi, H.; Miura, M.; Hijioka, A.; Kanazawa, Y.; Nishida, S.; Ikeda, S.; Nakamura, T. Lower osteocalcin and osteopontin contents of the femoral head in hip fracture patients than osteoarthritis patients. Osteoporos. Int. 2011, 22, 587–597. [Google Scholar] [CrossRef]
- Seibel, M.J.; Robins, S.P.; Bilezikian, J.P. Serum undercarboxylated osteocalcin and the risk of hip fracture. J. Clin. Endocrinol. Metab. 1997, 82, 717–718. [Google Scholar] [CrossRef]
- Vergnaud, P.; Garnero, P.; Meunier, P.J.; Bréart, G.; Kamihagi, K.; Delmas, P.D. Undercarboxylated osteocalcin measured with a specific immunoassay predicts hip fracture in elderly women: The EPIDOS study. J. Clin. Endocrinol. Metab. 1997, 82, 719–724. [Google Scholar] [CrossRef]
- Luukinen, H.; Käkönen, S.M.; Pettersson, K.; Koski, K.; Laippala, P.; Lövgren, T.; Kivelä, S.L.; Väänänen, H.K. Strong prediction of fractures among older adults by the ratio of carboxylated to total serum osteocalcin. J. Bone Miner. Res. 2000, 15, 2473–2478. [Google Scholar] [CrossRef]
- Cheung, A.M.; Tile, L.; Lee, Y.; Tomlinson, G.; Hawker, G.; Scher, J.; Hu, H.; Vieth, R.; Thompson, L.; Jamal, S.; et al. Vitamin K supplementation in postmenopausal women with osteopenia (ECKO trial): A randomized controlled trial. PLoS Med. 2008, 5, e196. [Google Scholar] [CrossRef] [PubMed]
- Booth, S.L.; Dallal, G.; Shea, M.K.; Gundberg, C.; Peterson, J.W.; Dawson-Hughes, B. Effect of vitamin K supplementation on bone loss in elderly men and women. J. Clin. Endocrinol. Metab. 2008, 93, 1217–1223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Binkley, N.; Harke, J.; Krueger, D.; Engelke, J.; Vallarta-Ast, N.; Gemar, D.; Checovich, M.; Chappell, R.; Suttie, J. Vitamin K treatment reduces undercarboxylated osteocalcin but does not alter bone turnover, density, or geometry in healthy postmenopausal North American women. J. Bone Miner. Res. 2009, 24, 983–991. [Google Scholar] [CrossRef] [PubMed]
- Emaus, N.; Gjesdal, C.G.; Almås, B.; Christensen, M.; Grimsgaard, A.S.; Berntsen, G.K.; Salomonsen, L.; Fønnebø, V. Vitamin K2 supplementation does not influence bone loss in early menopausal women: A randomised double-blind placebo-controlled trial. Osteoporos. Int. 2010, 21, 1731–1740. [Google Scholar] [CrossRef]
- Fang, Y.; Hu, C.; Tao, X.; Wan, Y.; Tao, F. Effect of vitamin K on bone mineral density: A meta-analysis of randomized controlled trials. J. Bone Miner. Metab. 2012, 30, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Cockayne, S.; Adamson, J.; Lanham-New, S.; Shearer, M.J.; Gilbody, S.; Torgerson, D.J. Vitamin K and the prevention of fractures: Systematic review and meta-analysis of randomized controlled trials. Arch. Intern. Med. 2006, 166, 1256–1261. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.B.; Wan, S.L.; Lu, Y.J.; Ning, L.; Liu, C.; Fan, S.W. Does vitamin K2 play a role in the prevention and treatment of osteoporosis for postmenopausal women: A meta-analysis of randomized controlled trials. Osteoporos. Int. 2015, 26, 1175–1186. [Google Scholar] [CrossRef]
- Zhao, J.G.; Zeng, X.T.; Wang, J.; Liu, L. Association between calcium or vitamin D supplementation and fracture incidence in community-dwelling older adults: A systematic review and meta-analysis. JAMA 2017, 318, 2466–2482. [Google Scholar] [CrossRef]
- Weaver, C.M.; Alexander, D.D.; Boushey, C.J.; Dawson-Hughes, B.; Lappe, J.M.; LeBoff, M.S.; Liu, S.; Looker, A.C.; Wallace, T.C.; Wang, D.D. Calcium plus vitamin D supplementation and risk of fractures: An updated meta-analysis from the National Osteoporosis Foundation. Osteoporos. Int. 2016, 27, 367–376. [Google Scholar] [CrossRef] [Green Version]
- Van Meurs, J.B.; Dhonukshe-Rutten, R.A.; Pluijm, S.M.; Van Der Klift, M.; De Jonge, R.; Lindemans, J.; De Groot, L.C.; Hofman, A.; Witteman, J.C.; Van Leeuwen, J.P.; et al. Homocysteine levels and the risk of osteoporotic fracture. N. Engl. J. Med. 2004, 350, 2033–2041. [Google Scholar] [CrossRef] [Green Version]
- McLean, R.R.; Jacques, P.F.; Selhub, J.; Tucker, K.L.; Samelson, E.J.; Broe, K.E.; Hannan, M.T.; Cupples, L.A.; Kiel, D.P. Homocysteine as a predictive factor for hip fracture in older person. N. Engl. J. Med. 2004, 350, 2042–2049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruan, J.; Gong, X.; Kong, J.; Wang, H.; Zheng, X.; Chen, T. Effect of B vitamin (folate, B6, and B12) supplementation on osteoporotic fracture and bone turnover markers: A meta-analysis. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2015, 21, 875–881. [Google Scholar]
- Dai, Z.; Koh, W.-P. B-Vitamins and bone health-A review of the current evidence. Nutrients 2015, 7, 3322–3346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knapen, M.H.; Schurgers, L.J.; Vermeer, C. Vitamin K2 supplementation improves hip bone geometry and bone strength indices in postmenopausal women. Osteoporos. Int. 2007, 18, 963–972. [Google Scholar] [CrossRef] [Green Version]
- Price, P.A.; Baukol, S.A. 1,25-dihydroxyvitamin D3 increases synthesis of the vitamin K-dependent bone protein by osteosarcoma cells. J. Biol. Chem. 1980, 255, 11660–11663. [Google Scholar]
- Koshihara, Y.; Hoshi, K.; Ishibashi, H.; Shiraki, M. Vitamin K2 promote 1α,25(OH)2 vitamin D3-induced mineralization in human periosteal osteoblasts. Calcif. Tissue Int. 1996, 59, 466–473. [Google Scholar]
- Sanguineti, R.; Monacelli, F.; Parodi, A.; Furfaro, A.L.; Borghi, R.; Pacini, D.; Pronzato, M.A.; Odetti, P.; Molfetta, L.; Traverso, N.; et al. Vitamin D3 and K2 may partially counterbalance the detrimental effects of pentosidine in ex vivo human osteoblasts. J. Biol. Regul. Homeost. Agents 2016, 30, 713–726. [Google Scholar]
- Luo, G.; Ducy, P.; McKnee, M.D.; Pinero, J.P.; Loyer, E.; Behringer, P.R.; Karsenty, G. Spontaneous calcification of arteries and cartilage in mice lackling matrix Gla protein. Nature 1997, 386, 78–81. [Google Scholar] [CrossRef]
- Lawton, D.M.; Andrew, J.G.; Marsh, D.R.; Hoyland, J.A.; Freemont, A.J. Expression of the gene encoding the matrix gla protein by mature osteoblasts in human fracture non-unions. Mol. Pathol. 1999, 52, 92–96. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Ma, Z.; Yan, K.; Wang, Y.; Yang, Y.; Wu, X. Matrix gla protein promotes the bone formation by up-regulating Wnt/ß-catenin signaling pathway. Front. Endocrinol. 2019, 10, 891–901. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhao, L.; Wang, N.; Li, J.; He, F.; Li, X.; Wu, S. Unexpected role of matrix Gla protein in osteoclasts: Inhibiting osteoclast differentiation and bone resorption. Mol. Cell. Biol. 2019, 39, e00012-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fusaro, M.; Gallieni, M.; Porta, C.; Nickolas, T.L.; Khairallah, P. Vitamin K effects in human health: New insights beyond bone and cardiovascular health. J. Nephrol. 2020, 33, 239–249. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Authors (Year of Publication) | Subjects | Mean Age (Years) | Average Serum or Plasma Level (nmol/L) | Country | Reference | |
|---|---|---|---|---|---|---|
| Vitamin K1 | MK-7 | |||||
| Nakano et al. | Elderly men | 82.2 | 1.22 | 6.6 | Japan | [12] |
| (2011) | Elderly men with fractures | 82.6 | 0.69 | 2.5 | ||
| Elderly women | 84.1 | 1.71 | 16.6 | |||
| Elderly women with fractures | 85.5 | 1.02 | 4.1 | |||
| Kuwabara et al. | Institutionalized elderly men | 84.9 | 1.38 | 1.2 | Japan | [13] |
| (2010) | Institutionalized elderly women | 88.7 | 1.31 | 0.8 | ||
| Martini et al. (2006) | Postmenopausal women | 70-71 | 1.7–1.8 | - | USA | [14] |
| Tsugawa et al. | 30–49 years-old women | 45.4 | 3.37 | 7.6 | Japan | [15] |
| (2006) | 50–69 years-old women | 59.6 | 3.86 | 13.0 | ||
| 70 years-old women | 74.9 | 2.86 | 6.5 | |||
| Beavan et al. | Postmenopausal women (Chinese) | 67.6 | 2.22 | - | UK | [16] |
| (2005) | Postmenopausal women (British) | 67.4 | 0.69 | - | ||
| Postmenopausal women (Gambian) | 66.7 | 0.80 | - | |||
| Booth et al. (2004) | Adult men (vitamin K intake, 151 μg/day) | 59 | 1.54 | - | USA | [17] |
| Premenopausal women (vitamin K intake, 17 μg/day) | 47.3 | 1.05 | - | |||
| Postmenopausal women (vitamin K intake, 177 μg/day) | 63 | 1.41 | - | |||
| Binkley et al. (2002) | Young adult men and women (vitamin K intake, 77–120 μg/day) | 25 | 0.61–1.0 | - | USA | [18] |
| Binkley et al. (2002) | Subjects with vitamin K1 intake of 375–500 μg/day | 2–4 | - | USA | [18] | |
| Kaneki et al. | Postmenopausal women (Tokyo) | 57.2 | 1.61 | 8.10 | Japan | [19] |
| (2001) | Postmenopausal women (Hiroshima) | 67.4 | 1.64 | 1.88 | ||
| Binkley et al. | Young adult men | 24 | 0.68 | - | USA | [20] |
| (2000) | Young adult women | 0.72 | - | |||
| Elderly men | 75 | 1.03 | - | |||
| Elderly women | 1.16 | - | ||||
| Hodges et al. | Elderly women with fractures | 81.7 | 0.75 | 0.18 | France | [21] |
| (1993) | Healthy elderly women | 80 | 1.30 | 0.35 | ||
| Gentili A et al. | Healthy subjects (n = 5) | 0.947 | Italy | [8] | ||
| (2014) | Patients under oral anticoagulant therapy (n = 5) | 0.691 | ||||
| Riphagen et al. (2015) | Renal transplant recipients (n = 60) | 55 | 1.35 (0.89–2.32) | <4.40 | Netherlands | [10] |
| Fusaro et al. (2012) | Healthy subjects (n = 62) | 56.8 | 1.36 | 2.53 | Italy | [22] |
| Dialysis paients (n = 387) | 64.2 | 0.98 | 0.87 | |||
| Holden et al. (2010) | Chronic kidney disease stages 3–5 (average vitamin K intake, 130 µg/day; 17.3–740 µg/day) | 61 | 2.1 (0–19.3) | - | Canada | [23] |
| Pilkey et al. (2007) | Dialysis patients | 62.6 | 0.99 | - | Canada | [24] |
| Holden et al. (2008) | Peritoneal dialysis patients (average dialysis period, 49 months) | 56 (28.7–85) | Median, 0.7 (0.1–2.2) | - | Canada | [25] |
| Uncarboxylated VKDP | Target Tissue | Subjects | Relationship to VK Status | Reference (Year) | |
|---|---|---|---|---|---|
| PIVKA-II | Liver | Adult men and women | Negative correlation with serum VK1 levels | Sokoll et al. (1996) | [26] |
| Elderly women | Increased by VK intake restriction; decreased by VK1 supplementation (86 μg/day) | Booth et al. (2003) | [27] | ||
| Adult patients with chronic kidney disease (stages 3–5) | Negative correlation with VK intake | Holden et al. (2010) | [23] | ||
| Elderly men and women | Negative correlation with VK intake | Kuwabara et al. (2011) | [28] | ||
| Adolescent boys and girls | Negative correlation with VK intake; required intake levels were at least 62 μg/day for boys and at least 54 μg/day for girls (approximately 1 μg/day/kg body weight) | Tsugawa et al. (2012) | [29] | ||
| Dialysis patients aged >18 years | Decreased by MK-7 supplementation | Westenfeld et al. (2012). | [30] | ||
| ucOC | Bone | Young and elderly men and women | Negative correlation with serum VK1 levels | Sokoll et al. (1996) | [26] |
| Young and elderly men and women | Negative correlation with serum K1 levels; decreased by VK1 supplementation | Binkley et al. (2000) | [20] | ||
| Healthy adults | Decreased by VK1 supplementation | Binkley et al. (2002) | [18] | ||
| Elderly women | Increased by VK intake restriction; decreased by VK1 supplementation | Booth et al. (2003) | [27] | ||
| Elderly women | Negative correlation with serum VK levels | Tsugawa et al. (2006) | [15] | ||
| Young adult men and women | Positive correlation between cOC-to-ucOC ratio and VK1 (MK-7) supplementation | Schurgers et al. (2007) | [31] | ||
| Elderly men and women | Negative correlation between VK intake and OCR | Kuwabara et al. (2011) | [28] | ||
| Dialysis patients aged >18 years | Decreased by MK-7 supplementation | Westenfeld et al. (2012). | [30] | ||
| Adolescent boys and girls | Negative correlation with VK intake; required VK intake levels were 155–188 μg/day | Tsugawa et al. (2012) | [29] | ||
| t-ucMGP | Blood vessels | Men and women in their 50s with hypertension | Positive correlation with OCR | Rennenberg et al. (2010) | [32] |
| Elderly women | Negative correlation with serum K1 level; decreased by taking menatetrenone (MK-4); increased by taking warfarin | Tsugawa et al. (2014) | [33] | ||
| dp-ucMGP | Blood vessels | Adults | Decreased by VK intake; increased by taking warfarin | Schurgers et al. (2008) | [34] |
| Elderly men and women | Negative correlation with VK intake and serum PK levels; positive correlation with PIVKA-II levels and %ucOC | Shea et al. (2011) | [35] | ||
| Dialysis patients aged >18 years | Decreased by MK-7 supplementation | Westenfeld et al. (2012). | [30] | ||
| Elderly women | Positive correlation with OCR (no correlation between OCR and dp-cMGP or t-ucMGP) | Dalmeijer et al. (2013) | [36] | ||
| Quartile by Osteocalcin Level | Observation Period (Person-Years) | Number of Incident DM Cases | Incident Rate (per 1000 Person-Years) | Age-Adjusted HR (95% CI) | p vs. Q4 |
|---|---|---|---|---|---|
| Q4 | 3293 | 4 | 1.2 | 1.00 (reference) | |
| Q3 | 3183 | 13 | 4.1 | 2.25 (1.15–11.6) | <0.05 |
| Q2 | 3324 | 14 | 4.2 | 3.58 (1.28–12.6) | <0.01 |
| Q1 | 3121 | 30 | 9.6 | 8.05 (3.17–27.1) | <0.01 |
| Item | HR | 95% CI | P |
|---|---|---|---|
| Age, years | 1.049 | 1.003–1.096 | 0.037 |
| BMI, kg/m2 | 1.078 | 0.974–1.193 | 0.149 |
| Osteocalcin, <6.1 ng/mL | 2.481 | 1.274–4.833 | 0.008 |
| Triglycerides, mg/dL | 1.001 | 0.998–1.004 | 0.376 |
| NTX, nmol /nmolCr | 0.999 | 0.983–1.015 | 0.911 |
| hs-CRP, mg/dL, | 1.227 | 0.285–5.287 | 0.784 |
| Adiponectin/leptin ratio | 0.803 | 0.644–1.000 | 0.050 |
| HbA1c, % | 2.518 | 1.858–3.414 | <0.0001 |
| L2-4BMD, g/cm2 | 1.497 | 0.306–7.330 | 0.619 |
| Phosphate, mg/dL | 1.908 | 0.910–4.003 | 0.087 |
| Homocysteine, nmol/mL | 1.017 | 0.918–1.127 | 0.745 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsugawa, N.; Shiraki, M. Vitamin K Nutrition and Bone Health. Nutrients 2020, 12, 1909. https://doi.org/10.3390/nu12071909
Tsugawa N, Shiraki M. Vitamin K Nutrition and Bone Health. Nutrients. 2020; 12(7):1909. https://doi.org/10.3390/nu12071909
Chicago/Turabian StyleTsugawa, Naoko, and Masataka Shiraki. 2020. "Vitamin K Nutrition and Bone Health" Nutrients 12, no. 7: 1909. https://doi.org/10.3390/nu12071909
APA StyleTsugawa, N., & Shiraki, M. (2020). Vitamin K Nutrition and Bone Health. Nutrients, 12(7), 1909. https://doi.org/10.3390/nu12071909