Dietary Oligofructose Alone or in Combination with 2′-Fucosyllactose Differentially Improves Recognition Memory and Hippocampal mRNA Expression

 ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
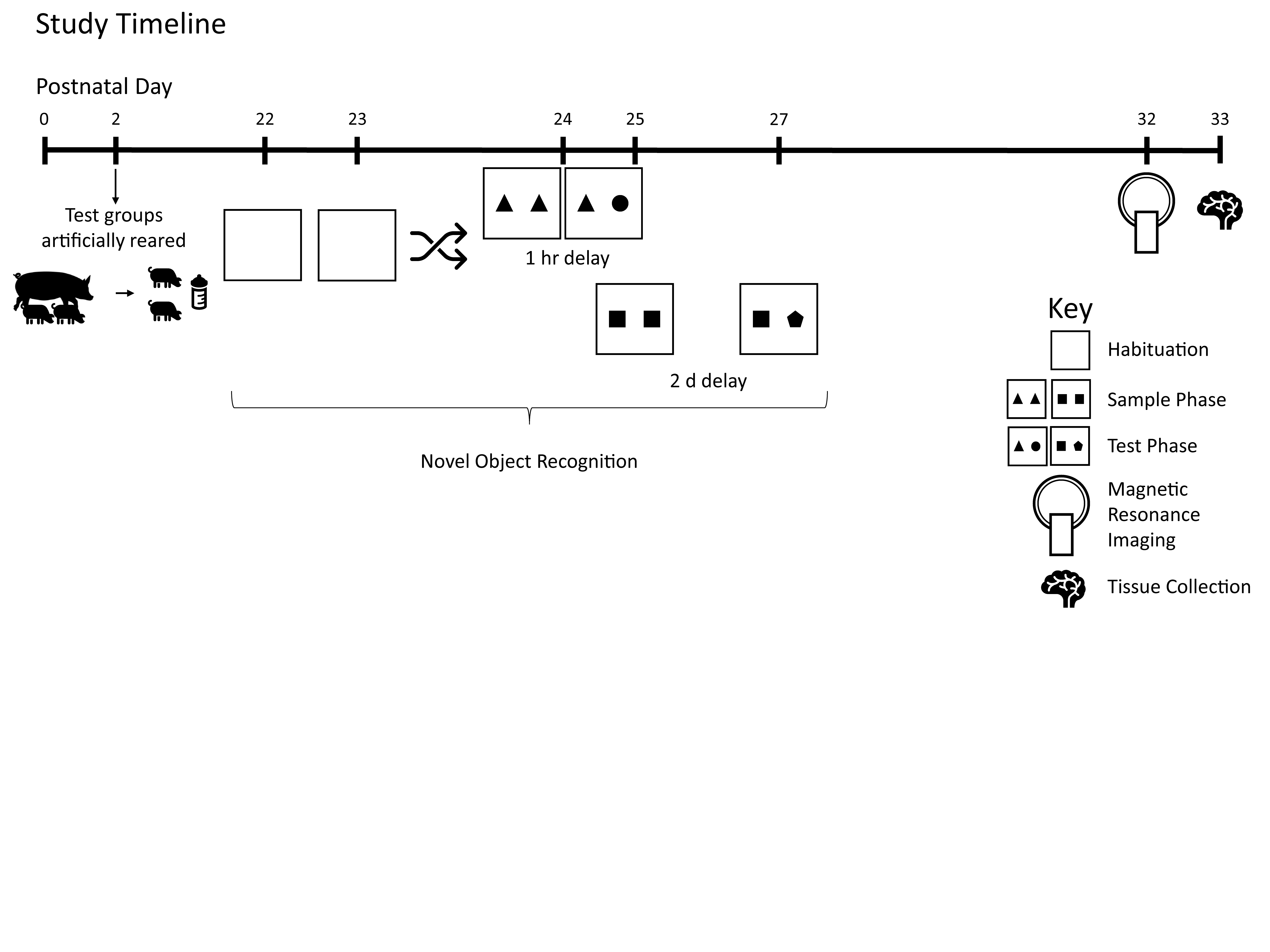
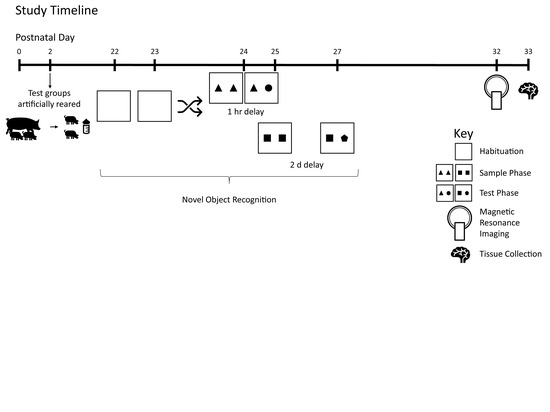
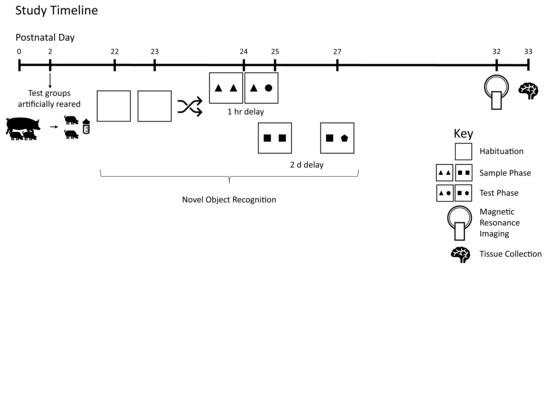
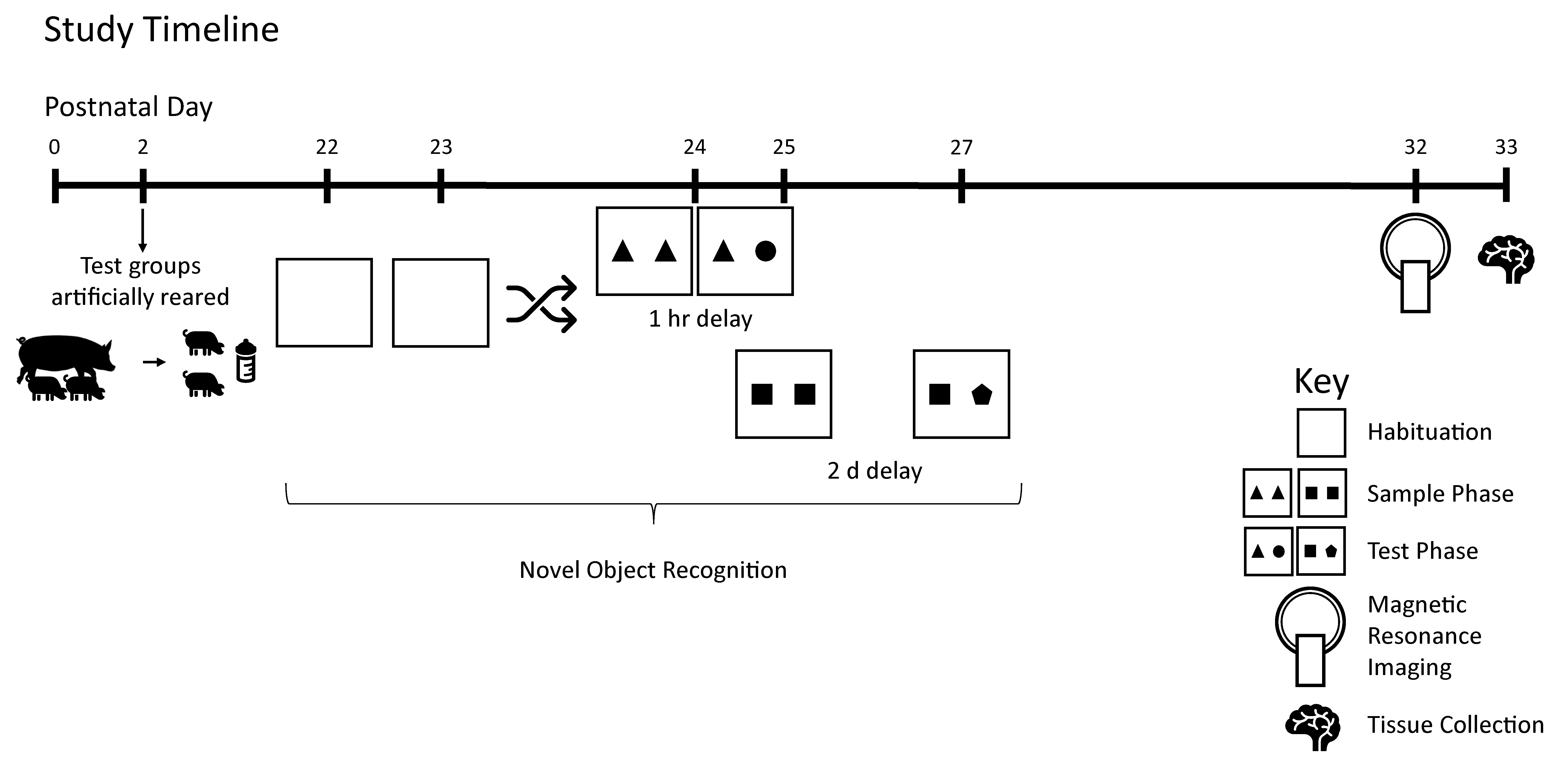
2.1. Animals and Housing
2.2. Dietary Treatments
2.3. Behavior
2.4. Magnetic Resonance Imaging (MRI)
2.4.1. Structural MRI
2.4.2. Diffusion Tensor Imaging
2.4.3. Magnetic Resonance Spectroscopy
2.5. Hippocampal Gene Expression
2.6. Statistical Analysis
3. Results
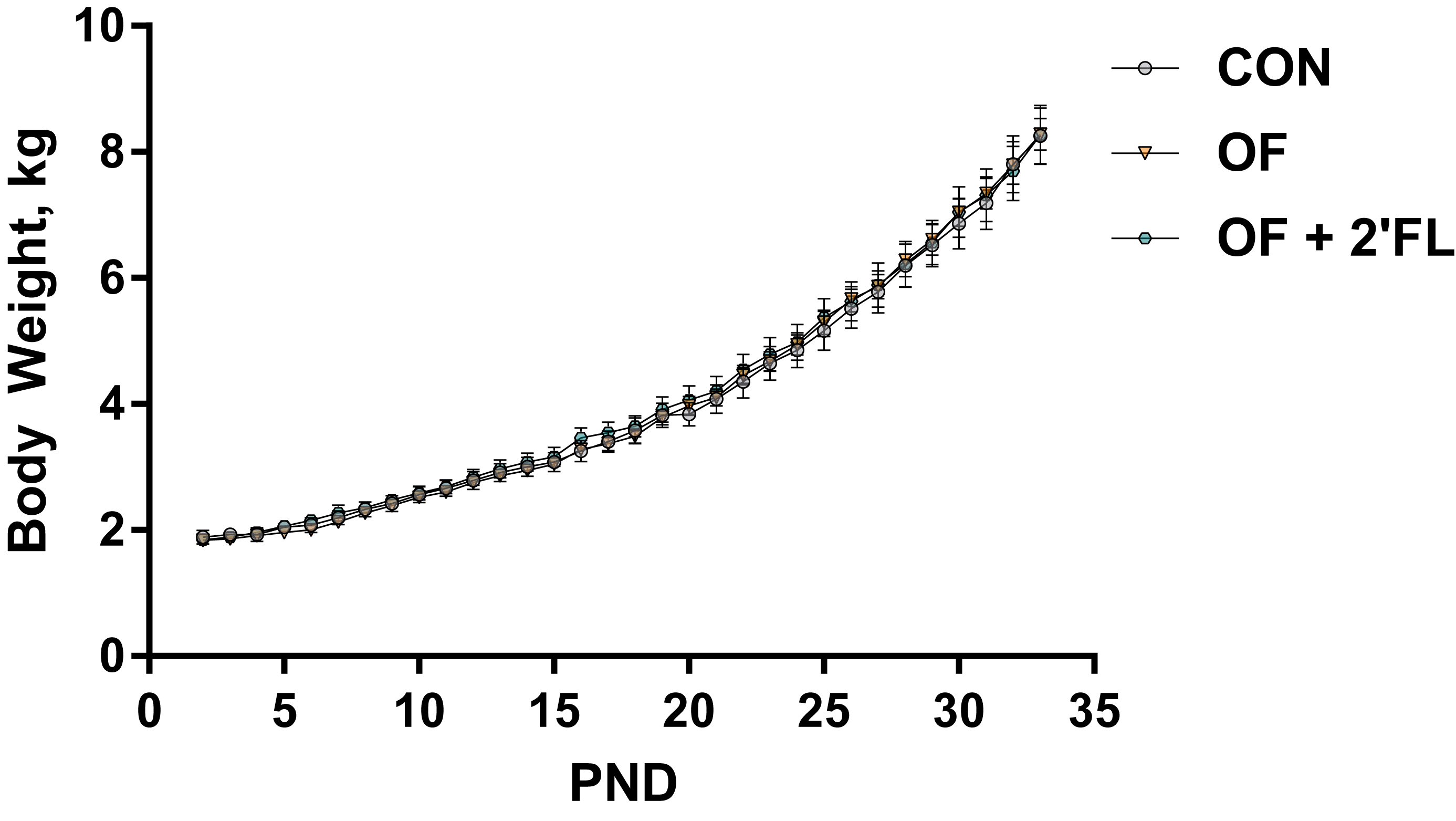
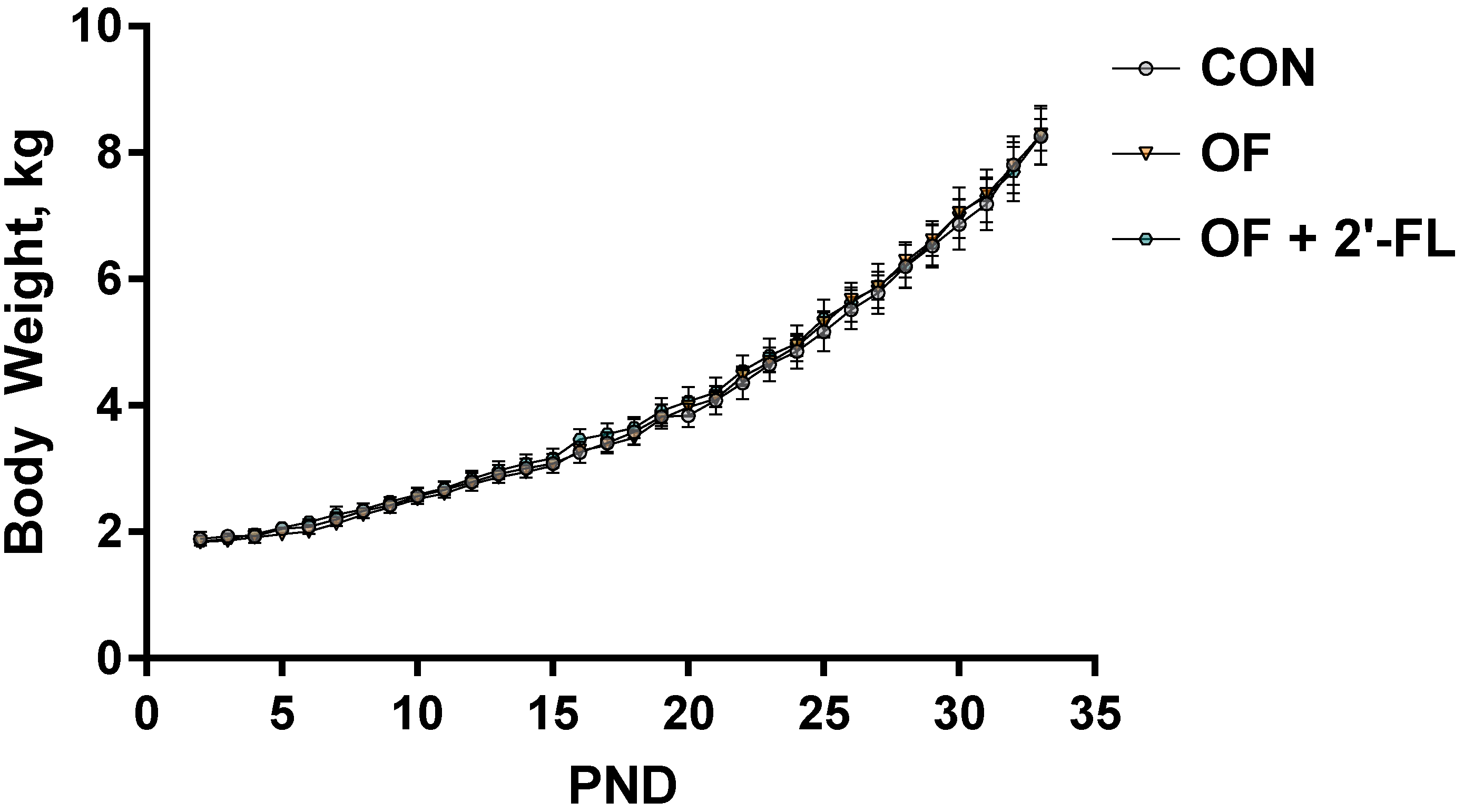
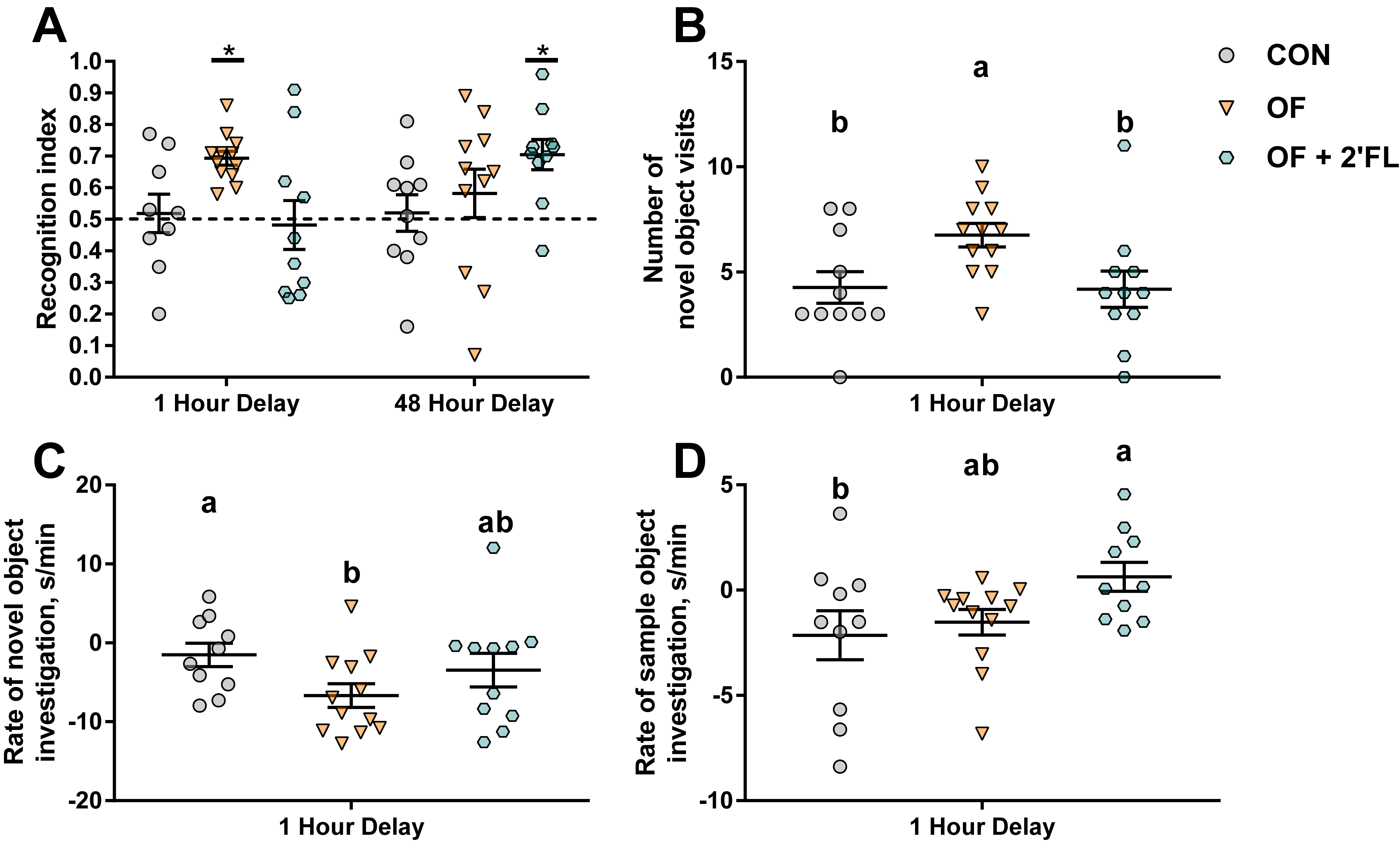
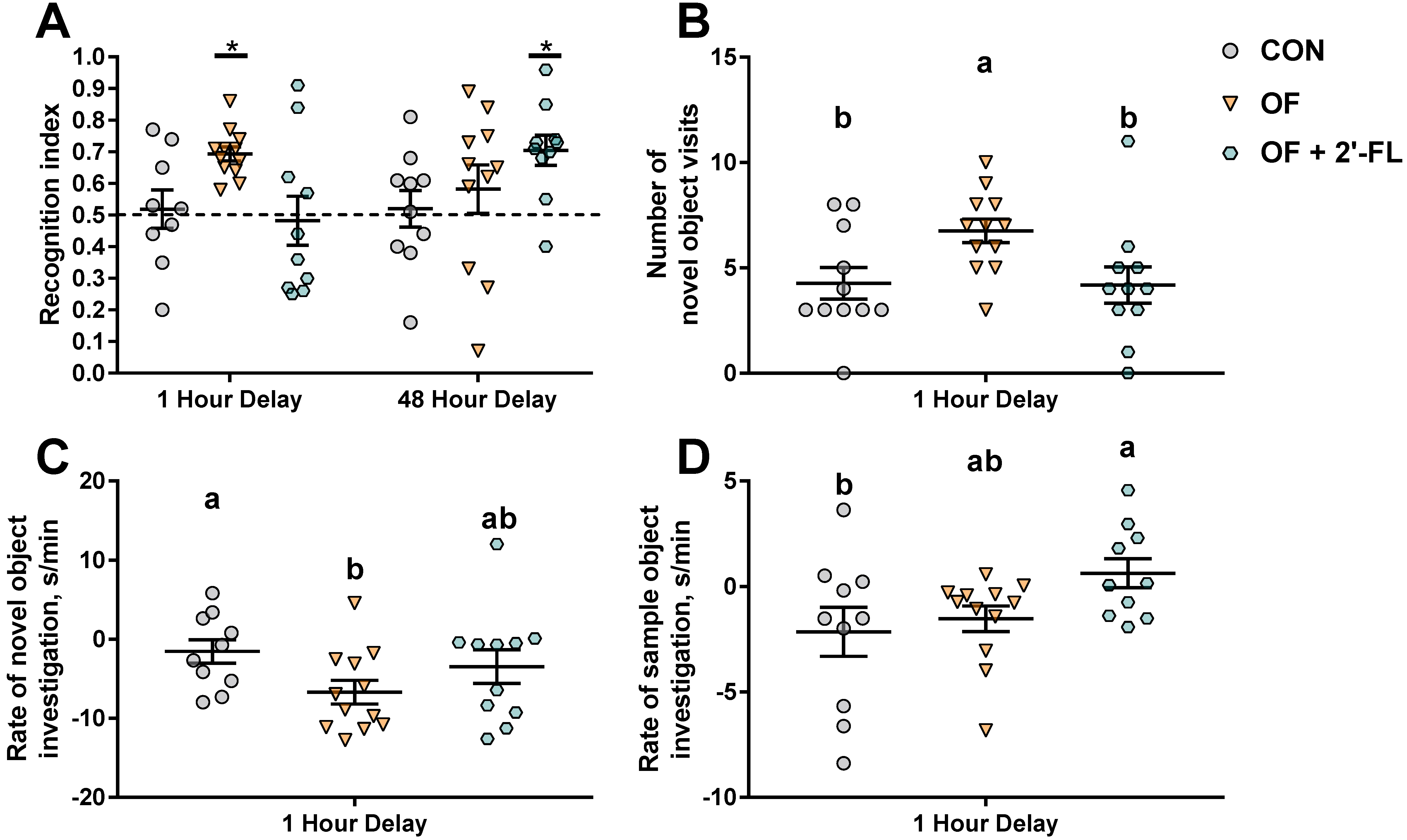
3.1. Growth and Behavior
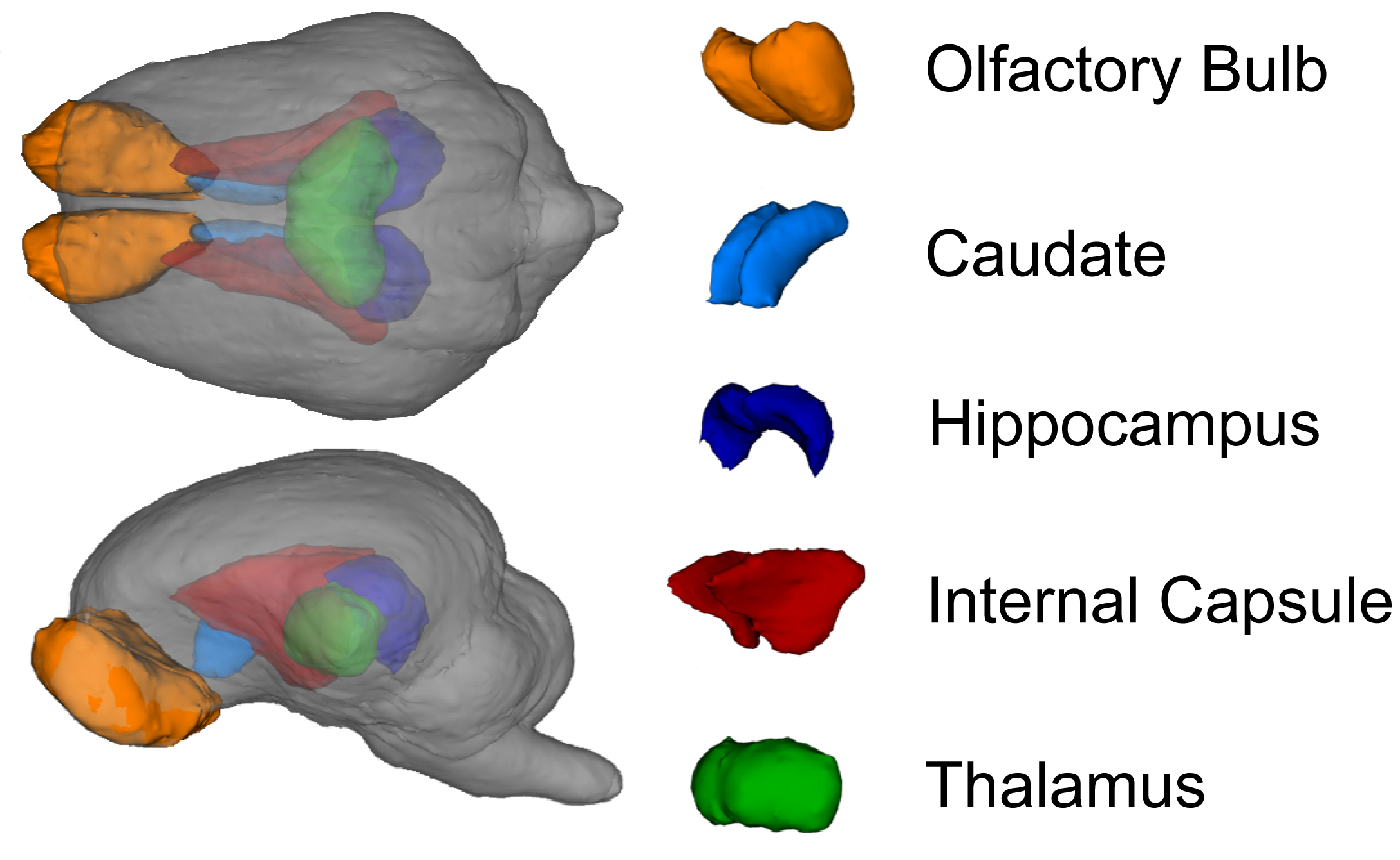
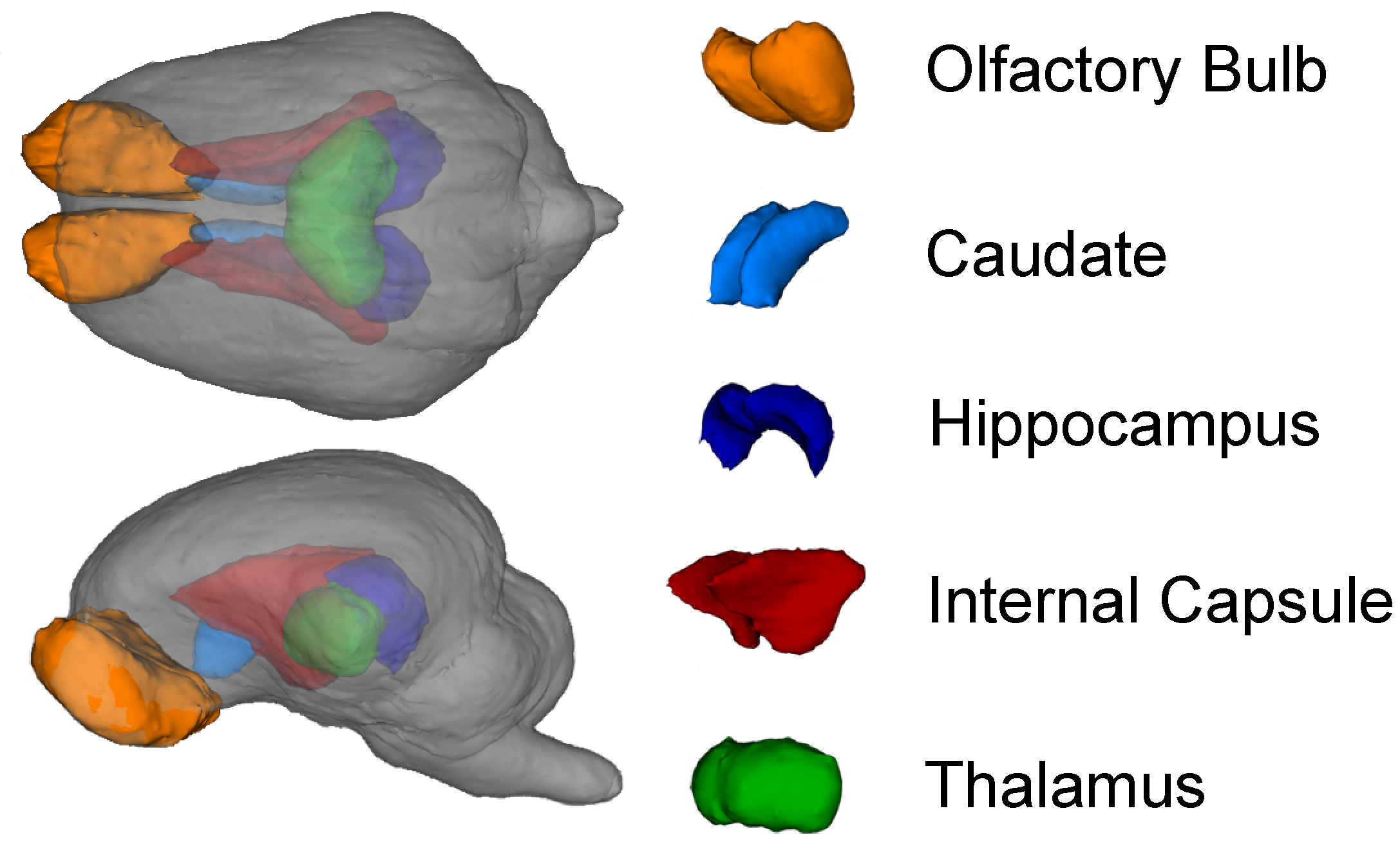
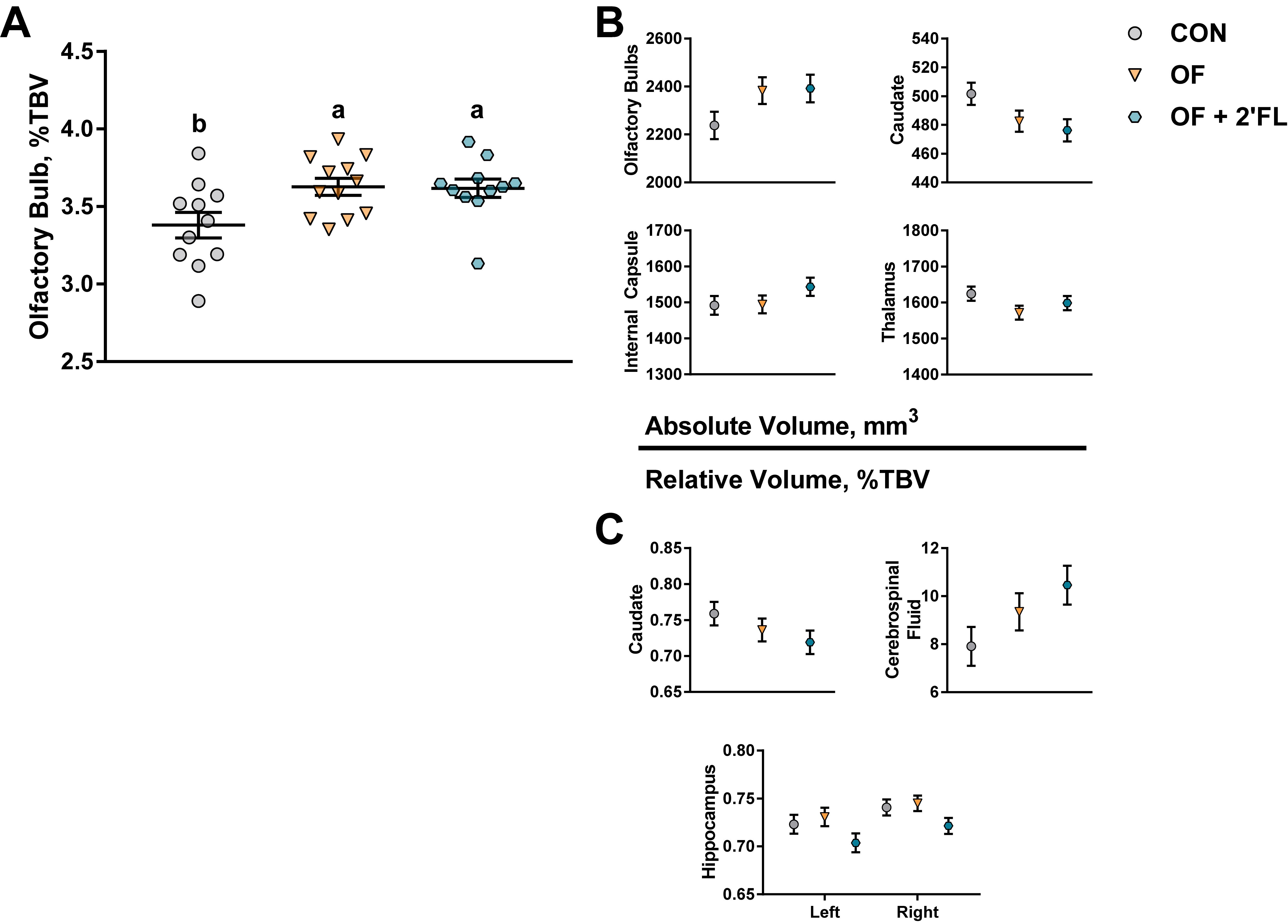
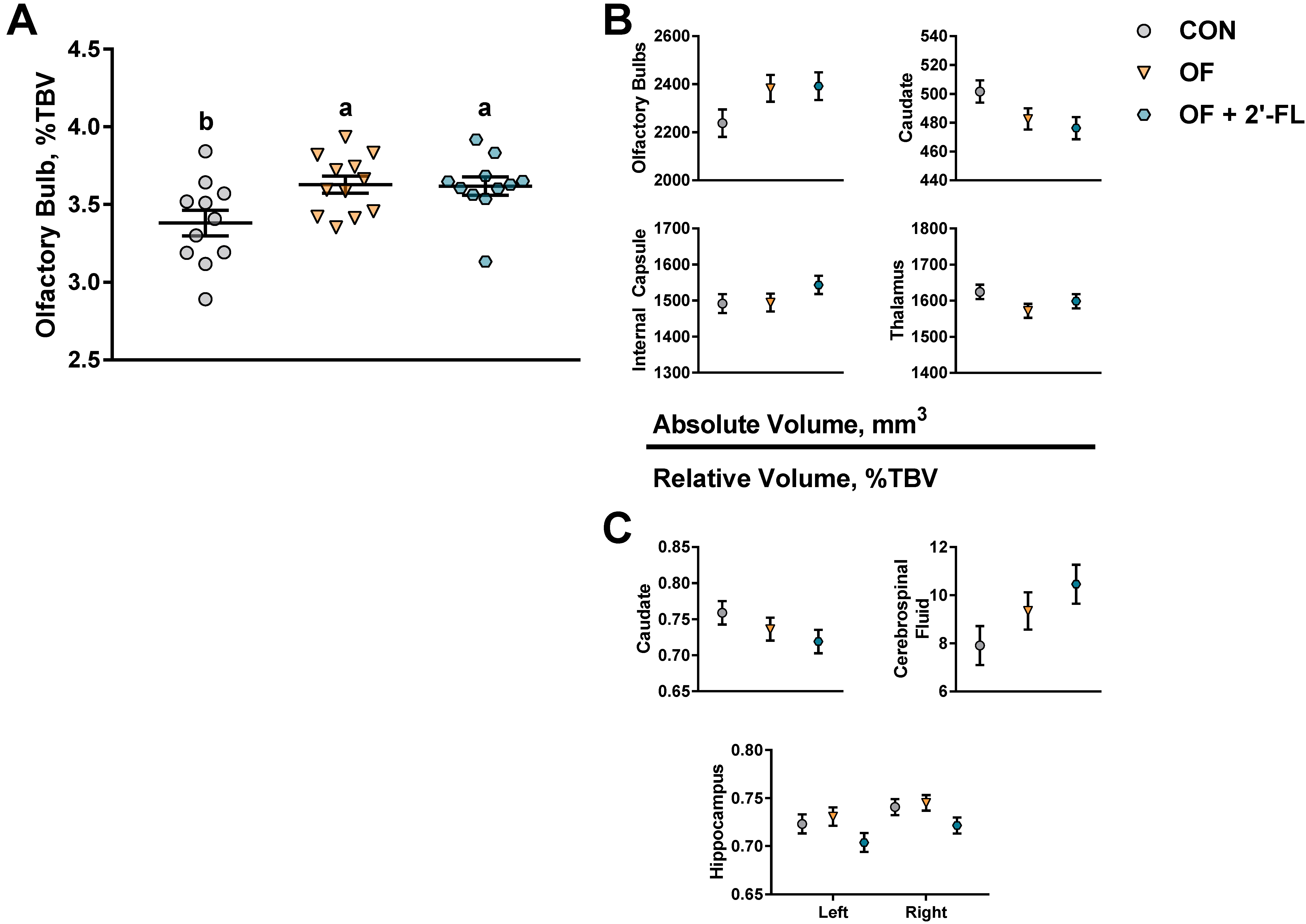
3.2. MRI
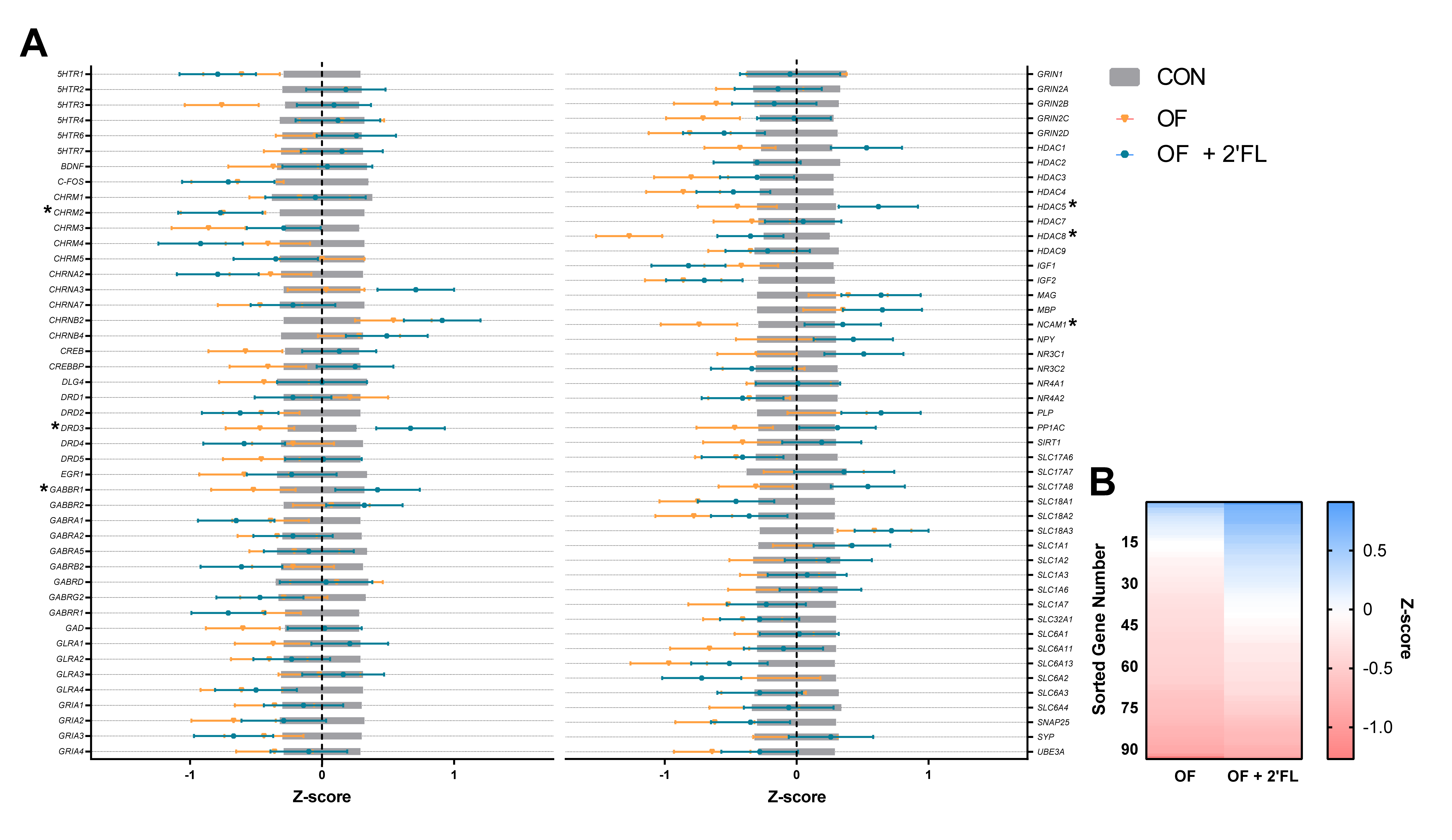
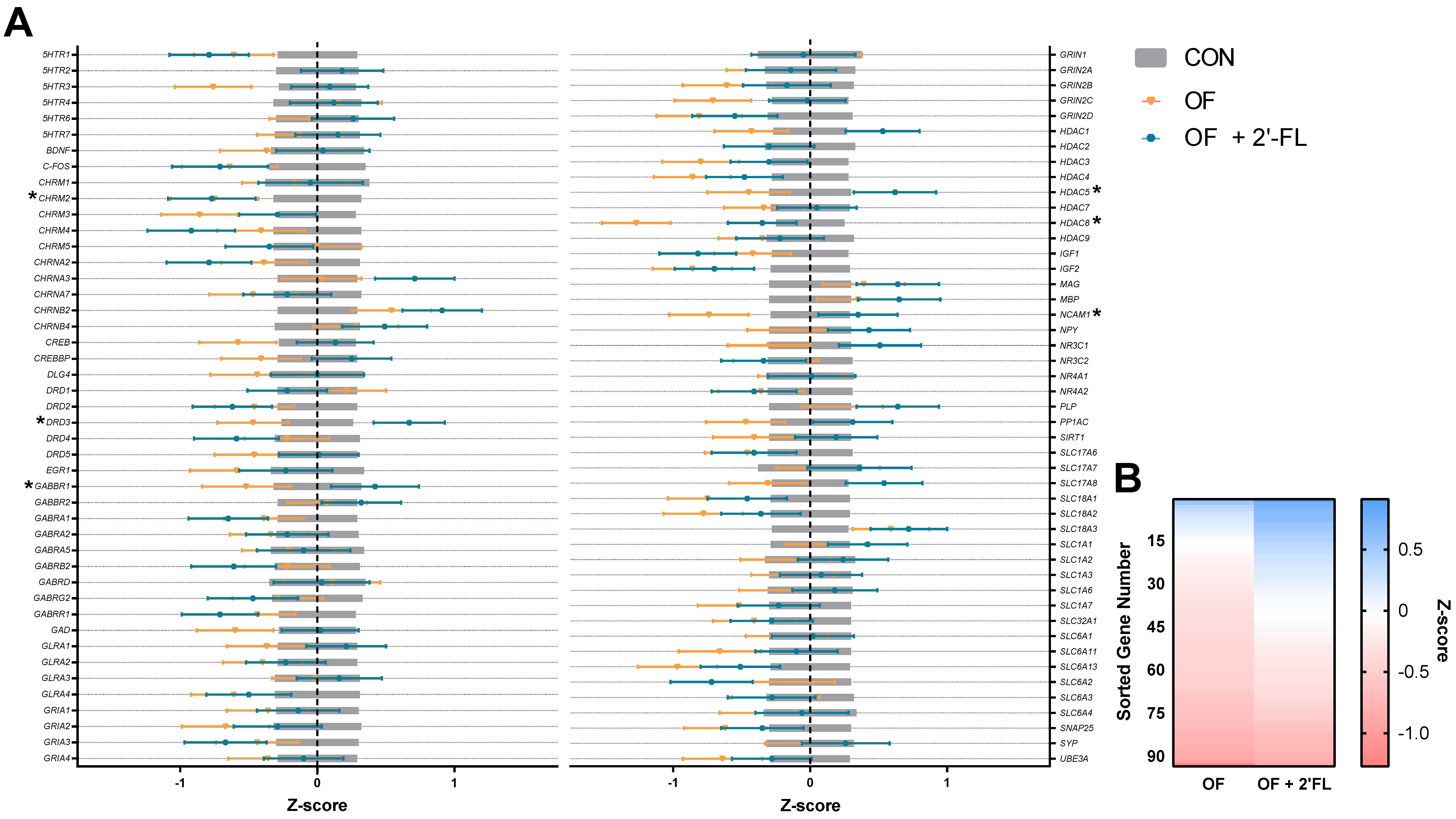
3.3. Hippocampal Gene Expression
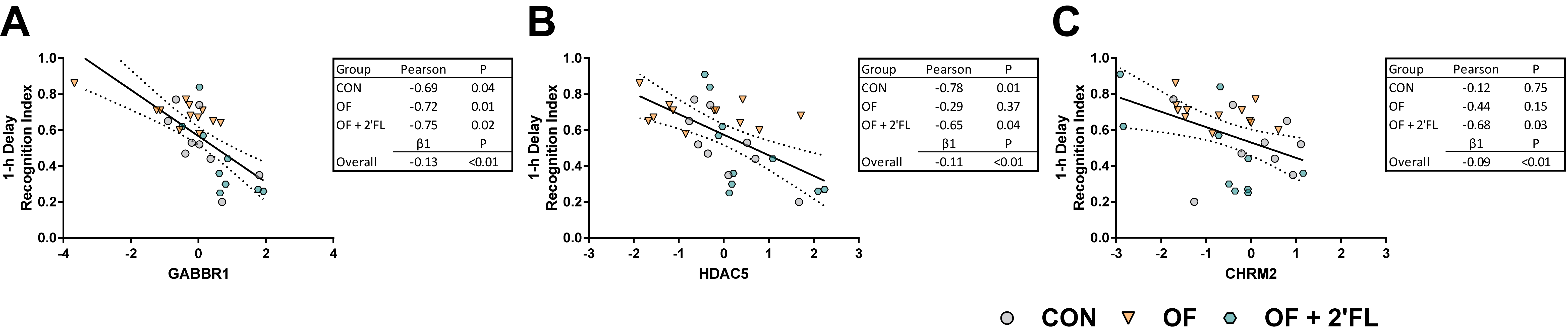
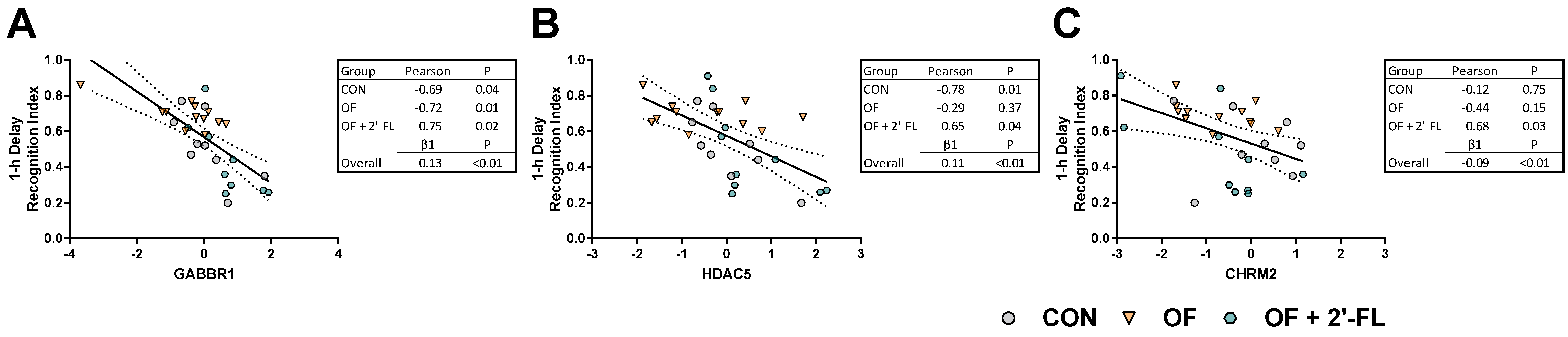
3.4. Correlation and Linear Regression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Clevenger, M.A.; Turnbull, D.; Inoue, H.; Enomoto, M.; Allen, J.A.; Henderson, L.M.; Jones, E. Toxicological Evaluation of Neosugar: Genotoxicity, Carcinogenicity, and Chronic Toxicity. J. Am. Coll. Toxicol. 1988, 7, 643–662. [Google Scholar] [CrossRef]
- Rivero-Urgell, M.; Santamaria-Orleans, A. Oligosaccharides: Application in infant food. Early Hum. Dev. 2001, 65, 43–52. [Google Scholar] [CrossRef]
- Costalos, C.; Kapiki, A.; Apostolou, M.; Papathoma, E. The effect of a prebiotic supplemented formula on growth and stool microbiology of term infants. Early Hum. Dev. 2008, 84, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Kapiki, A.; Costalos, C.; Oikonomidou, C.; Triantafyllidou, A.; Loukatou, E.; Pertrohilou, V. The effect of a fructo-oligosaccharide supplemented formula on gut flora of preterm infants. Early Hum. Dev. 2007, 83, 335–339. [Google Scholar] [CrossRef] [PubMed]
- Savignac, H.M.; Corona, G.; Mills, H.; Chen, L.; Spencer, J.P.E.; Tzortzis, G.; Burnet, P.W.J. Prebiotic feeding elevates central brain derived neurotrophic factor, N-methyl-d-aspartate receptor subunits and d-serine. Neurochem. Int. 2013, 63, 756–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yen, C.-H.; Wang, C.-H.; Wu, W.-T.; Chen, H.-L. Fructo-oligosaccharide improved brain β-amyloid, β-secretase, cognitive function, and plasma antioxidant levels in D-galactose-treated Balb/cJ mice. Nutr. Neurosci. 2017, 20, 228–237. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Yang, X.; Yang, J.; Lai, G.; Yong, T.; Tang, X.; Shuai, O.; Zhou, G.; Xie, Y.; Wu, Q. Prebiotic Effect of Fructooligosaccharides from Morinda officinalis on Alzheimer’s Disease in Rodent Models by Targeting the Microbiota-Gut-Brain Axis. Front. Aging Neurosci. 2017, 9, 403. [Google Scholar] [CrossRef] [Green Version]
- Ballard, O.; Morrow, A.L. Human milk composition: Nutrients and bioactive factors. Pediatr. Clin. North Am. 2013, 60, 49–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bode, L. Human milk oligosaccharides: Every baby needs a sugar mama. Glycobiology 2012, 22, 1147–1162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urashima, T.; Saito, T.; Nakamura, T.; Messer, M. Oligosaccharides of milk and colostrum in non-human mammals. Glycoconj. J. 2001, 18, 357–371. [Google Scholar] [CrossRef]
- Puccio, G.; Alliet, P.; Cajozzo, C.; Janssens, E.; Corsello, G.; Sprenger, N.; Wernimont, S.; Egli, D.; Gosoniu, L.; Steenhout, P. Effects of infant formula with human milk oligosaccharides on growth and morbidity: A randomized multicenter trial. J. Pediatr. Gastroenterol. Nutr. 2017, 64, 624–631. [Google Scholar] [CrossRef] [Green Version]
- Tarr, A.J.; Galley, J.D.; Fisher, S.E.; Chichlowski, M.; Berg, B.M.; Bailey, M.T. The prebiotics 3′Sialyllactose and 6′Sialyllactose diminish stressor-induced anxiety-like behavior and colonic microbiota alterations: Evidence for effects on the gut–brain axis. Brain. Behav. Immun. 2015, 50, 166–177. [Google Scholar] [CrossRef] [Green Version]
- Vázquez, E.; Barranco, A.; Ramírez, M.; Gruart, A.; Delgado-García, J.M.; Martínez-Lara, E.; Blanco, S.; Martín, M.J.; Castanys, E.; Buck, R.; et al. Effects of a human milk oligosaccharide, 2′-fucosyllactose, on hippocampal long-term potentiation and learning capabilities in rodents. J. Nutr. Biochem. 2015, 26, 455–465. [Google Scholar] [CrossRef] [PubMed]
- Vázquez, E.; Barranco, A.; Ramírez, M.; Gruart, A.; Delgado-García, J.M.; Jimenez, M.L.; Buck, R.; Rueda, R. Dietary 2’-fucosyllactose enhances operant conditioning and long-term potentiation via gut-brain communication through the vagus nerve in rodents. PLoS ONE 2016, 11, 1–14. [Google Scholar] [CrossRef]
- Palmano, K.; Rowan, A.; Guillermo, R.; Guan, J.; McJarrow, P. The Role of Gangliosides in Neurodevelopment. Nutrients 2015, 7, 3891–3913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sukumar, R.; Rose, S.P.R.; Burgoyne, R.D. Increased Incorporation of [3H]Fucose into Chick Brain Glycoproteins Following Training on a Passive Avoidance Task. J. Neurochem. 1980, 34, 1000–1006. [Google Scholar] [CrossRef] [PubMed]
- Fagioli, S.; Castellano, C.; Oliverio, A.; Toffano, G. Effect of chronic GM1 ganglioside administration on passive avoidance retention in mice. Neurosci. Lett. 1990, 109, 212–216. [Google Scholar] [CrossRef]
- Silva, R.H.H.; Felicio, L.F.F.; Nasello, A.G.G.; Vital, M.A.B.F.A.; Frussa-Filho, R. Effect of ganglioside (GM1) on memory in senescent rats. Neurobiol. Aging 1996, 17, 583–586. [Google Scholar] [CrossRef]
- Popov, N.; Toffano, G.; Riechert, U.; Matthies, H. Effects of intraventricularly applied gangliosides and N-acetylneuraminic acid on acquisition and retention performance of a brightness discrimination task in rats. Pharmacol. Biochem. Behav. 1989, 34, 209–212. [Google Scholar] [CrossRef]
- Jork, R.; Grecksch, G.; Matthies, H. Impairment of glycoprotein fucosylation in rat hippocampus and the consequences on memory formation. Pharmacol. Biochem. Behav. 1986, 25, 1137–1144. [Google Scholar] [CrossRef]
- Fleming, S.A.; Monaikul, S.; Patsavas, A.J.; Waworuntu, R.V.; Berg, B.M.; Dilger, R.N. Dietary polydextrose and galactooligosaccharide increase exploratory behavior, improve recognition memory, and alter neurochemistry in the young pig. Nutr. Neurosci. 2019, 22, 499–512. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, K.; Cowen, P.J.; Harmer, C.J.; Tzortzis, G.; Errington, S.; Burnet, P.W.J. Prebiotic intake reduces the waking cortisol response and alters emotional bias in healthy volunteers. Psychopharmacology 2015, 232, 1793–1801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, S.; Lu, Z.; Gao, Z.; An, J.; Wu, X.; Li, X.; Dai, X.; Zheng, Q.; Sun, Y. Chitosan oligosaccharides alleviate cognitive deficits in an amyloid-β1–42-induced rat model of Alzheimer’s disease. Int. J. Biol. Macromol. 2016, 83, 416–425. [Google Scholar] [CrossRef]
- Bertelsen, R.J.; Jensen, E.T.; Ringel-Kulka, T. Use of probiotics and prebiotics in infant feeding. Best Pract. Res. Clin. Gastroenterol. 2016, 30, 39–48. [Google Scholar] [CrossRef]
- Odle, J.; Lin, X.; Jacobi, S.K.; Kim, S.W.; Stahl, C.H. The suckling piglet as an agrimedical model for the study of pediatric nutrition and metabolism. Annu. Rev. Anim. Biosci. 2014, 2, 419–444. [Google Scholar] [CrossRef] [PubMed]
- Mudd, A.T.; Dilger, R.N. Early-life nutrition and neurodevelopment: Use of the piglet as a translational model. Adv. Nutr. 2017, 8, 92–104. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Donovan, S.M. Human microbiota-associated swine: Current progress and future opportunities. ILAR J. 2015, 56, 63–73. [Google Scholar] [CrossRef]
- Aluthge, N.D.; Van Sambeek, D.M.; Carney-Hinkle, E.E.; Li, Y.S.; Fernando, S.C.; Burkey, T.E. Board invited review: The pig microbiota and the potential for harnessing the power of the microbiome to improve growth and health1. J. Anim. Sci. 2019, 97, 3741–3757. [Google Scholar] [CrossRef] [PubMed]
- Roura, E.; Koopmans, S.-J.; Lallès, J.-P.; Le Huerou-Luron, I.; de Jager, N.; Schuurman, T.; Val-Laillet, D. Critical review evaluating the pig as a model for human nutritional physiology. Nutr. Res. Rev. 2016, 29, 60–90. [Google Scholar] [CrossRef]
- Niilo, L. Clostridium perfringens Type C Enterotoxemia. Can. Vet. J. 1988, 29, 658–664. [Google Scholar] [PubMed]
- Yao, M.; Lien, E.L.; Capeding, M.R.Z.; Fitzgerald, M.; Ramanujam, K.; Yuhas, R.; Northington, R.; Lebumfacil, J.; Wang, L.; DeRusso, P.A. Effects of Term Infant Formulas Containing High sn-2 Palmitate With and Without Oligofructose on Stool Composition, Stool Characteristics, and Bifidogenicity. J. Pediatr. Gastroenterol. Nutr. 2014, 59, 440–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wernimont, S.; Northington, R.; Kullen, M.J.; Yao, M.; Bettler, J. Effect of an α-lactalbumin-enriched infant formula supplemented with oligofructose on fecal microbiota, stool characteristics, and hydration status: A randomized, double-blind, controlled trial. Clin. Pediatr. 2015, 54, 359–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marriage, B.J.; Buck, R.H.; Goehring, K.C.; Oliver, J.S.; Williams, J.A. Infants Fed a Lower Calorie Formula With 2’FL Show Growth and 2’FL Uptake Like Breast-Fed Infants. J. Pediatr. Gastroenterol. Nutr. 2015, 61, 649–658. [Google Scholar] [CrossRef] [Green Version]
- Moustgaard, A.; Lind, N.M.; Hemmingsen, R.; Hansen, A.K. Spontaneous Object Recognition in the Göttingen Minipig. Neural Plast. 2002, 9, 255–259. [Google Scholar] [CrossRef]
- Gifford, A.K.; Cloutier, S.; Newberry, R.C. Objects as enrichment: Effects of object exposure time and delay interval on object recognition memory of the domestic pig. Appl. Anim. Behav. Sci. 2007, 107, 206–217. [Google Scholar] [CrossRef]
- Kornum, B.R.; Thygesen, K.S.; Nielsen, T.R.; Knudsen, G.M.; Lind, N.M. The effect of the inter-phase delay interval in the spontaneous object recognition test for pigs. Behav. Brain Res. 2007, 181, 210–217. [Google Scholar] [CrossRef] [PubMed]
- Kouwenberg, A.-L.; Walsh, C.J.; Morgan, B.E.; Martin, G.M. Episodic-like memory in crossbred Yucatan minipigs (Sus scrofa). Appl. Anim. Behav. Sci. 2009, 117, 165–172. [Google Scholar] [CrossRef]
- Fleming, S.A.; Dilger, R.N. Young pigs exhibit differential exploratory behavior during novelty preference tasks in response to age, sex, and delay. Behav. Brain Res. 2017, 321, 50–60. [Google Scholar] [CrossRef]
- Fleming, S.A.; Chichlowski, M.; Berg, B.M.; Donovan, S.M.; Dilger, R.N. Dietary Sialyllactose Does Not Influence Measures of Recognition Memory or Diurnal Activity in the Young Pig. Nutrients 2018, 10, 395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacob, R.M.; Mudd, A.T.; Alexander, L.S.; Lai, C.-S.; Dilger, R.N. Comparison of Brain Development in Sow-Reared and Artificially Reared Piglets. Front. Pediatr. 2016, 4, 95. [Google Scholar] [CrossRef] [Green Version]
- Radlowski, E.C.; Conrad, M.S.; Lezmi, S.; Dilger, R.N.; Sutton, B.; Larsen, R.; Johnson, R.W. A neonatal piglet model for investigating brain and cognitive development in small for gestational age human infants. PLoS ONE 2014, 9, e91951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mudd, A.T.; Getty, C.M.; Sutton, B.P.; Dilger, R.N. Perinatal choline deficiency delays brain development and alters metabolite concentrations in the young pig. Nutr. Neurosci. 2016, 19, 425–433. [Google Scholar] [CrossRef] [PubMed]
- Conrad, M.S.; Sutton, B.P.; Dilger, R.N.; Johnson, R.W. An in vivo three-dimensional magnetic resonance imaging-based averaged brain collection of the neonatal piglet (Sus scrofa). PLoS ONE 2014, 9, e107650. [Google Scholar] [CrossRef] [PubMed]
- Kao, A.C.C.; Harty, S.; Burnet, P.W.J. The Influence of Prebiotics on Neurobiology and Behavior. Int. Rev. Neurobiol. 2016, 131, 21–48. [Google Scholar] [CrossRef] [PubMed]
- Barker, G.R.I.; Warburton, E.C. When Is the Hippocampus Involved in Recognition Memory? J. Neurosci. 2011, 31, 10721–10731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hummel, T.; Smitka, M.; Puschmann, S.; Gerber, J.C.; Schaal, B.; Buschhüter, D. Correlation between olfactory bulb volume and olfactory function in children and adolescents. Exp. Brain Res. 2011, 214, 285–291. [Google Scholar] [CrossRef]
- Buschhüter, D.; Smitka, M.; Puschmann, S.; Gerber, J.C.; Witt, M.; Abolmaali, N.D.; Hummel, T. Correlation between olfactory bulb volume and olfactory function. Neuroimage 2008, 42, 498–502. [Google Scholar] [CrossRef]
- Cummings, D.M.; Knab, B.R.; Brunjes, P.C. Effects of unilateral olfactory deprivation in the developing opossum, Monodelphis domestica. J. Neurobiol. 1997, 33, 429–438. [Google Scholar] [CrossRef]
- Korol, D.L.; Brunjes, P.C. Unilateral naris closure and vascular development in the rat olfactory bulb. Neuroscience 1992, 46, 631–641. [Google Scholar] [CrossRef]
- Hammond, R.S.; Tull, L.E.; Stackman, R.W. On the delay-dependent involvement of the hippocampus in object recognition memory. Neurobiol. Learn. Mem. 2004, 82, 26–34. [Google Scholar] [CrossRef]
- Gosso, F.M.; de Geus, E.J.; Polderman, T.J.; Boomsma, D.I.; Posthuma, D.; Heutink, P. Exploring the functional role of the CHRM2 gene in human cognition: Results from a dense genotyping and brain expression study. BMC Med. Genet. 2007, 8, 66. [Google Scholar] [CrossRef] [Green Version]
- Cavallaro, S.; D’Agata, V.; Manickam, P.; Dufour, F.; Alkon, D.L. Memory-specific temporal profiles of gene expression in the hippocampus. Proc. Natl. Acad. Sci. USA 2002, 99, 16279–16284. [Google Scholar] [CrossRef] [Green Version]
- Scott, H.; Rogers, M.F.; Scott, H.L.; Campbell, C.; Warburton, E.C.; Uney, J.B. Recognition memory-induced gene expression in the perirhinal cortex: A transcriptomic analysis. Behav. Brain Res. 2017, 328, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Bravo, J.A.; Forsythe, P.; Chew, M.V.; Escaravage, E.; Savignac, H.M.; Dinan, T.G.; Bienenstock, J.; Cryan, J.F. Ingestion of Lactobacillus strain regulates emotional behavior and central GABA receptor expression in a mouse via the vagus nerve. Proc. Natl. Acad. Sci. USA 2011, 108, 16050–16055. [Google Scholar] [CrossRef] [Green Version]
- Janik, R.; Thomason, L.A.M.; Stanisz, A.M.; Forsythe, P.; Bienenstock, J.; Stanisz, G.J. Magnetic resonance spectroscopy reveals oral Lactobacillus promotion of increases in brain GABA, N-acetyl aspartate and glutamate. Neuroimage 2016, 125, 988–995. [Google Scholar] [CrossRef]
- Pitsikas, N.; Rigamonti, A.E.; Cella, S.G.; Muller, E.E. The GABAB receptor and recognition memory: Possible modulation of its behavioral effects by the nitrergic system. Neuroscience 2003, 118, 1121–1127. [Google Scholar] [CrossRef]
- Heaney, C.F.; Kinney, J.W. Role of GABABreceptors in learning and memory and neurological disorders. Neurosci. Biobehav. Rev. 2016, 63, 1–28. [Google Scholar] [CrossRef]
- Savignac, H.M.; Couch, Y.; Stratford, M.; Bannerman, D.M.; Tzortzis, G.; Anthony, D.C.; Burnet, P.W.J. Prebiotic administration normalizes lipopolysaccharide (LPS)-induced anxiety and cortical 5-HT2A receptor and IL1-β levels in male mice. Brain. Behav. Immun. 2016, 52, 120–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nutrient | Units | Base Formula | Final Composition 3 | ||
|---|---|---|---|---|---|
| Per kg | Per Liter | Per kg | Per Liter | ||
| Energy and Macronutrients | |||||
| Metabolizable Energy | kcal | 4286.4 | 857.3 | 3989.4 | 797.9 |
| Crude Protein | g | 241.0 | 48.2 | 224.3 | 44.9 |
| Crude Fat | g | 241.0 | 48.2 | 224.3 | 44.9 |
| Lactose | g | 369.1 | 73.8 | 343.5 | 68.7 |
| Crude Fiber | mg | 20.0 | 4.0 | 18.6 | 3.7 |
| Ash | g | 85.3 | 17.1 | 79.4 | 15.9 |
| Minerals | |||||
| Calcium | mg | 1000.0 | 200.0 | 930.7 | 186.1 |
| Copper | mg | 12.1 | 2.4 | 11.2 | 2.2 |
| Total Phosphorous | mg | 800.0 | 160.0 | 744.6 | 148.9 |
| Potassium | mg | 1835.0 | 367.0 | 1707.8 | 341.6 |
| Selenium | μg | 875.0 | 175.0 | 814.4 | 162.9 |
| Sodium | mg | 345.0 | 69.0 | 321.1 | 64.2 |
| Zinc | mg | 120.0 | 24.0 | 111.7 | 22.3 |
| Vitamins and Other Nutrients | |||||
| Vitamin A | IU | 82,427.3 | 16,485.5 | 76,715.0 | 15,343.0 |
| Vitamin D | IU | 11,563.9 | 2312.8 | 10,762.5 | 2152.5 |
| Vitamin E | IU | 253.3 | 50.7 | 235.7 | 47.1 |
| Lysine | g | 25.2 | 5.0 | 23.4 | 4.7 |
| Methionine + Cysteine | g | 9.9 | 2.0 | 9.2 | 1.8 |
| Oligofructose 2 | 2′-FL 3 | Additional Lactose | Total OS | Total Lactose | Total Carbohydrate | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Diet, g | Per kg | Per Liter | Per kg | Per Liter | Per kg | Per Liter | Per kg | Per Liter | Per kg | Per Liter | Per kg | Per Liter |
| Formulated | ||||||||||||
| CON | 0.00 | 0.00 | 0.00 | 0.00 | 69.30 | 13.86 | 0.00 | 0.00 | 412.79 | 82.56 | 412.79 | 82.56 |
| OF | 26.08 | 5.22 | 0.00 | 0.00 | 61.98 | 8.64 | 26.08 | 5.22 | 386.71 | 77.34 | 412.79 | 82.56 |
| OF + 2′-FL | 26.08 | 5.22 | 4.92 | 0.98 | 38.30 | 7.66 | 31.00 | 6.20 | 381.79 | 76.36 | 412.79 | 82.56 |
| Analyzed | ||||||||||||
| CON | 0.00 | 0.00 | 0.00 | 0.00 | NQ | NQ | 0.00 | 0.00 | NQ | NQ | NQ | NQ |
| OF | 18.10 | 3.62 | 0.00 | 0.00 | NQ | NQ | 18.10 | 3.62 | NQ | NQ | NQ | NQ |
| OF + 2′-FL | 17.05 | 3.41 | 5.60 | 1.12 | NQ | NQ | 22.65 | 4.53 | NQ | NQ | NQ | NQ |
| CON | OF | OF + 2′-FL | ||||||
|---|---|---|---|---|---|---|---|---|
| Measure 2 | N | Mean | N | Mean | N | Mean | SEM | p-Value 3 |
| CHRM2 4 | 12 | 0 a | 12 | −0.75 a | 12 | −0.77 a | 0.32 | 0.045 |
| DRD3 | 12 | 0 ab | 12 | −0.47 b | 12 | 0.67 a | 0.26 | 0.016 |
| GABBR1 | 12 | 0 ab | 12 | −0.52 b | 12 | 0.42 a | 0.32 | 0.037 |
| HDAC5 | 11 | 0 ab | 12 | −0.45 b | 12 | 0.62 a | 0.30 | 0.012 |
| HDAC8 | 12 | 0 a | 12 | −1.27 b | 12 | −0.35 a | 0.25 | 0.003 |
| NCAM1 | 12 | 0 a,b | 12 | −0.74 b | 12 | 0.35 a | 0.29 | 0.011 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fleming, S.A.; Mudd, A.T.; Hauser, J.; Yan, J.; Metairon, S.; Steiner, P.; Donovan, S.M.; Dilger, R.N. Dietary Oligofructose Alone or in Combination with 2′-Fucosyllactose Differentially Improves Recognition Memory and Hippocampal mRNA Expression. Nutrients 2020, 12, 2131. https://doi.org/10.3390/nu12072131
Fleming SA, Mudd AT, Hauser J, Yan J, Metairon S, Steiner P, Donovan SM, Dilger RN. Dietary Oligofructose Alone or in Combination with 2′-Fucosyllactose Differentially Improves Recognition Memory and Hippocampal mRNA Expression. Nutrients. 2020; 12(7):2131. https://doi.org/10.3390/nu12072131
Chicago/Turabian StyleFleming, Stephen A., Austin T. Mudd, Jonas Hauser, Jian Yan, Sylviane Metairon, Pascal Steiner, Sharon M. Donovan, and Ryan N. Dilger. 2020. "Dietary Oligofructose Alone or in Combination with 2′-Fucosyllactose Differentially Improves Recognition Memory and Hippocampal mRNA Expression" Nutrients 12, no. 7: 2131. https://doi.org/10.3390/nu12072131
APA StyleFleming, S. A., Mudd, A. T., Hauser, J., Yan, J., Metairon, S., Steiner, P., Donovan, S. M., & Dilger, R. N. (2020). Dietary Oligofructose Alone or in Combination with 2′-Fucosyllactose Differentially Improves Recognition Memory and Hippocampal mRNA Expression. Nutrients, 12(7), 2131. https://doi.org/10.3390/nu12072131