Beneficial Effects of Vitamin K Status on Glycemic Regulation and Diabetes Mellitus: A Mini-Review



Abstract
:1. Introduction
2. Improvement of Insulin Sensitivity and Glycemic Status
3. Possible Effect of VK Supplementation on Insulin Secretion and Glycemic Status
3.1. Insulinotropic Effect
3.2. Modulation of VK-Dependent Proteins
3.3. Prevention of Inflammation
4. Beneficial Effects of VK on Diabetes-Related Complications
4.1. Cataractogenesis
4.2. Diabetic Nephropathy
4.3. Diabetic Peripheral Neuropathy
4.4. Cardiovascular Disease
4.5. Osteopenia and Osteoporosis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Chaudhury, A.; Duvoor, C.; Reddy Dendi, V.S.; Kraleti, S.; Chada, A.; Ravilla, R.; Marco, A.; Shekhawat, N.S.; Montales, M.T.; Kuriakose, K.; et al. Clinical Review of Antidiabetic Drugs: Implications for Type 2 Diabetes Mellitus Management. Front. Endocrinol. (Lausanne) 2017, 8, 6. [Google Scholar] [CrossRef] [Green Version]
- Defronzo, R.A. Banting Lecture. From the triumvirate to the ominous octet: A new paradigm for the treatment of type 2 diabetes mellitus. Diabetes 2009, 58, 773–795. [Google Scholar] [CrossRef] [Green Version]
- Komatsu, R.; Matsuyama, T.; Namba, M.; Watanabe, N.; Itoh, H.; Kono, N.; Tarui, S. Glucagonostatic and insulinotropic action of glucagonlike peptide I-(7-36)-amide. Diabetes 1989, 38, 902–905. [Google Scholar] [CrossRef]
- Meier, J.J.; Nauck, M.A.; Schmidt, W.E.; Gallwitz, B. Gastric inhibitory polypeptide: The neglected incretin revisited. Regul. Pept. 2002, 107, 1–13. [Google Scholar] [CrossRef]
- Vilsbøll, T.; Krarup, T.; Madsbad, S.; Holst, J.J. Both GLP-1 and GIP are insulinotropic at basal and postprandial glucose levels and contribute nearly equally to the incretin effect of a meal in healthy subjects. Regul. Pept. 2003, 114, 115–121. [Google Scholar] [CrossRef]
- Prigeon, R.L.; Quddusi, S.; Paty, B.; D′Alessio, D.A. Suppression of glucose production by GLP-1 independent of islet hormones: A novel extrapancreatic effect. Am. J. Physiol. Endocrinol. Metab. 2003, 285, E701–E707. [Google Scholar] [CrossRef]
- Drucker, D.J. Mechanisms of Action and Therapeutic Application of Glucagon-like Peptide-1. Cell Metab. 2018, 27, 740–756. [Google Scholar] [CrossRef] [Green Version]
- Herzberg-Schäfer, S.; Heni, M.; Stefan, N.; Häring, H.U.; Fritsche, A. Impairment of GLP1-induced insulin secretion: Role of genetic background, insulin resistance and hyperglycaemia. Diabetes Obes. Metab. 2012, 14 (Suppl. 3), 85–90. [Google Scholar] [CrossRef]
- Tasyurek, H.M.; Altunbas, H.A.; Balci, M.K.; Sanlioglu, S. Incretins: Their physiology and application in the treatment of diabetes mellitus. Diabetes Metab. Res. Rev. 2014, 30, 354–371. [Google Scholar] [CrossRef]
- Ghosh-Swaby, O.R.; Goodman, S.G.; Leiter, L.A.; Cheng, A.; Connelly, K.A.; Fitchett, D.; Jüni, P.; Farkouh, M.E.; Udell, J.A. Glucose-lowering drugs or strategies, atherosclerotic cardiovascular events, and heart failure in people with or at risk of type 2 diabetes: An updated systematic review and meta-analysis of randomised cardiovascular outcome trials. Lancet Diabetes Endocrinol. 2020, 8, 418–435. [Google Scholar] [CrossRef]
- Langenberg, C.; Lotta, L.A. Genomic insights into the causes of type 2 diabetes. Lancet 2018, 391, 2463–2474. [Google Scholar] [CrossRef]
- Booth, S.L. Vitamin K: Food composition and dietary intakes. Food Nutr. Res. 2012, 56, 5505. [Google Scholar] [CrossRef] [Green Version]
- Fu, X.; Harshman, S.G.; Shen, X.; Haytowitz, D.B.; Karl, J.P.; Wolfe, B.E.; Booth, S.L. Multiple Vitamin K Forms Exist in Dairy Foods. Curr. Dev. Nutr. 2017, 1, e000638. [Google Scholar] [CrossRef] [Green Version]
- Martius, C.; Alvino, C. On the transformation of vitamin K-1 (phyllochinon) into vitamin K2(20) by the development of an embryo in a hen′s egg. Biochem. Z 1964, 340, 316–319. [Google Scholar]
- Thijssen, H.H.; Drittij-Reijnders, M.J. Vitamin K status in human tissues: Tissue-specific accumulation of phylloquinone and menaquinone-4. Br. J. Nutr. 1996, 75, 121–127. [Google Scholar] [CrossRef]
- Komai, M.; Shirakawa, H. Vitamin K metabolism. Menaquinone-4 (MK-4) formation from ingested VK analogues and its potent relation to bone function. Clin. Calcium 2007, 17, 1663–1672. [Google Scholar]
- Shirakawa, H.; Katsurai, T.; Komai, M. Conversion of menaquinone-4 in animal organs and it functions. Jpn. Oil Chem. Soc. 2014, 14, 547–553. [Google Scholar] [CrossRef]
- Sakamoto, N.; Nishiike, T.; Iguchi, H.; Sakamoto, K. Relationship between acute insulin response and vitamin K intake in healthy young male volunteers. Diabetes Nutr. Metab. 1999, 12, 37–41. [Google Scholar]
- Yoshida, M.; Booth, S.L.; Meigs, J.B.; Saltzman, E.; Jacques, P.F. Phylloquinone intake, insulin sensitivity, and glycemic status in men and women. Am. J. Clin. Nutr. 2008, 88, 210–215. [Google Scholar] [CrossRef] [Green Version]
- Beulens, J.W.; van der, A.D.; Grobbee, D.E.; Sluijs, I.; Spijkerman, A.M.; van der Schouw, Y.T. Dietary phylloquinone and menaquinones intakes and risk of type 2 diabetes. Diabetes Care 2010, 33, 1699–1705. [Google Scholar] [CrossRef] [Green Version]
- Ibarrola-Jurado, N.; Salas-Salvadó, J.; Martínez-González, M.A.; Bulló, M. Dietary phylloquinone intake and risk of type 2 diabetes in elderly subjects at high risk of cardiovascular disease. Am. J. Clin. Nutr. 2012, 96, 1113–1118. [Google Scholar] [CrossRef]
- Zwakenberg, S.R.; Remmelzwaal, S.; Beulens, J.W.J.; Booth, S.L.; Burgess, S.; Dashti, H.S.; Imamura, F.; Feskens, E.J.M.; van der Schouw, Y.T.; Sluijs, I. Circulating Phylloquinone Concentrations and Risk of Type 2 Diabetes: A Mendelian Randomization Study. Diabetes 2019, 68, 220–225. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, M.; Jacques, P.F.; Meigs, J.B.; Saltzman, E.; Shea, M.K.; Gundberg, C.; Dawson-Hughes, B.; Dallal, G.; Booth, S.L. Effect of vitamin K supplementation on insulin resistance in older men and women. Diabetes Care 2008, 31, 2092–2096. [Google Scholar] [CrossRef] [Green Version]
- Rasekhi, H.; Karandish, M.; Jalali, M.T.; Mohammadshahi, M.; Zarei, M.; Saki, A.; Shahbazian, H. Phylloquinone supplementation improves glycemic status independent of the effects of adiponectin levels in premonopause women with prediabetes: A double-blind randomized controlled clinical trial. J. Diabetes Metab. Disord. 2015, 14, 1. [Google Scholar] [CrossRef] [Green Version]
- Rasekhi, H.; Karandish, M.; Jalali, M.T.; Mohammad-Shahi, M.; Zarei, M.; Saki, A.; Shahbazian, H. The effect of vitamin K1 supplementation on sensitivity and insulin resistance via osteocalcin in prediabetic women: A double-blind randomized controlled clinical trial. Eur. J. Clin. Nutr. 2015, 69, 891–895. [Google Scholar] [CrossRef]
- Sakamoto, N.; Nishiike, T.; Iguchi, H.; Sakamoto, K. Possible effects of one week vitamin K (menaquinone-4) tablets intake on glucose tolerance in healthy young male volunteers with different descarboxy prothrombin levels. Clin. Nutr. 2000, 19, 259–263. [Google Scholar] [CrossRef]
- Choi, H.J.; Yu, J.; Choi, H.; An, J.H.; Kim, S.W.; Park, K.S.; Jang, H.C.; Kim, S.Y.; Shin, C.S. Vitamin K2 supplementation improves insulin sensitivity via osteocalcin metabolism: A placebo-controlled trial. Diabetes Care 2011, 34, e147. [Google Scholar] [CrossRef] [Green Version]
- Sakamoto, N.; Wakabayashi, I.; Sakamoto, K. Low vitamin K intake effects on glucose tolerance in rats. Int. J. Vitam. Nutr. Res. 1999, 69, 27–31. [Google Scholar] [CrossRef]
- Seyama, Y.; Kimoto, S.; Marukawa, Y.; Horiuchi, M.; Hayashi, M.; Usami, E. Comparative effects of vitamin K2 and estradiol on experimental arteriosclerosis with diabetes mellitus. Int. J. Vitam. Nutr. Res. 2000, 70, 301–304. [Google Scholar] [CrossRef] [Green Version]
- Barnett, A.H. The role of GLP-1 mimetics and basal insulin analogues in type 2 diabetes mellitus: Guidance from studies of liraglutide. Diabetes Obes. Metab. 2012, 14, 304–314. [Google Scholar] [CrossRef] [Green Version]
- Drucker, D.J.; Nauck, M.A. The incretin system: Glucagon-like peptide-1 receptor agonists and dipeptidyl peptidase-4 inhibitors in type 2 diabetes. Lancet 2006, 368, 1696–1705. [Google Scholar] [CrossRef]
- Ho, H.J.; Shirakawa, H.; Hirahara, K.; Sone, H.; Kamiyama, S.; Komai, M. Menaquinone-4 Amplified Glucose-Stimulated Insulin Secretion in Isolated Mouse Pancreatic Islets and INS-1 Rat Insulinoma Cells. Int. J. Mol. Sci. 2019, 20, 1995. [Google Scholar] [CrossRef] [Green Version]
- Fusaro, M.; Gallieni, M.; Rizzo, M.A.; Stucchi, A.; Delanaye, P.; Cavalier, E.; Moysés, R.M.A.; Jorgetti, V.; Iervasi, G.; Giannini, S.; et al. Vitamin K plasma levels determination in human health. Clin. Chem. Lab. Med. 2017, 55, 789–799. [Google Scholar] [CrossRef]
- El Asmar, M.S.; Naoum, J.J.; Arbid, E.J. Vitamin k dependent proteins and the role of vitamin k2 in the modulation of vascular calcification: A review. Oman Med. J. 2014, 29, 172–177. [Google Scholar] [CrossRef]
- Luo, G.; Ducy, P.; McKee, M.D.; Pinero, G.J.; Loyer, E.; Behringer, R.R.; Karsenty, G. Spontaneous calcification of arteries and cartilage in mice lacking matrix GLA protein. Nature 1997, 386, 78–81. [Google Scholar] [CrossRef]
- Teebi, A.S.; Lambert, D.M.; Kaye, G.M.; Al-Fifi, S.; Tewfik, T.L.; Azouz, E.M. Keutel syndrome: Further characterization and review. Am. J. Med. Genet. 1998, 78, 182–187. [Google Scholar]
- Munroe, P.B.; Olgunturk, R.O.; Fryns, J.P.; Van Maldergem, L.; Ziereisen, F.; Yuksel, B.; Gardiner, R.M.; Chung, E. Mutations in the gene encoding the human matrix Gla protein cause Keutel syndrome. Nat. Genet. 1999, 21, 142–144. [Google Scholar] [CrossRef]
- Cranenburg, E.C.; Koos, R.; Schurgers, L.J.; Magdeleyns, E.J.; Schoonbrood, T.H.; Landewe, R.B.; Brandenburg, V.M.; Bekers, O.; Vermeer, C. Characterisation and potential diagnostic value of circulating matrix Gla protein (MGP) species. Thromb. Haemost. 2010, 104, 811–822. [Google Scholar] [CrossRef]
- Roumeliotis, S.; Dounousi, E.; Eleftheriadis, T.; Liakopoulos, V. Association of the Inactive Circulating Matrix Gla Protein with Vitamin K Intake, Calcification, Mortality, and Cardiovascular Disease: A Review. Int. J. Mol. Sci. 2019, 20, 628. [Google Scholar] [CrossRef] [Green Version]
- Basta, G.; Corciu, A.I.; Vianello, A.; Del Turco, S.; Foffa, I.; Navarra, T.; Chiappino, D.; Berti, S.; Mazzone, A. Circulating soluble receptor for advanced glycation end-product levels are decreased in patients with calcific aortic valve stenosis. Atherosclerosis 2010, 210, 614–618. [Google Scholar] [CrossRef]
- Olson, J.C.; Edmundowicz, D.; Becker, D.J.; Kuller, L.H.; Orchard, T.J. Coronary calcium in adults with type 1 diabetes: A stronger correlate of clinical coronary artery disease in men than in women. Diabetes 2000, 49, 1571–1578. [Google Scholar] [CrossRef] [Green Version]
- Neubauer, B. A quantitative study of peripheral arterial calcification and glucose tolerance in elderly diabetics and non-diabetics. Diabetologia 1971, 7, 409–413. [Google Scholar] [CrossRef] [Green Version]
- Conway, B.; Edmundowicz, D.; Matter, N.; Maynard, J.; Orchard, T. Skin fluorescence correlates strongly with coronary artery calcification severity in type 1 diabetes. Diabetes Technol. 2010, 12, 339–345. [Google Scholar] [CrossRef]
- Thomsen, S.B.; Rathcke, C.N.; Zerahn, B.; Vestergaard, H. Increased levels of the calcification marker matrix Gla Protein and the inflammatory markers YKL-40 and CRP in patients with type 2 diabetes and ischemic heart disease. Cardiovasc. Diabetol. 2010, 9, 86. [Google Scholar] [CrossRef] [Green Version]
- Dalmeijer, G.W.; van der Schouw, Y.T.; Magdeleyns, E.J.; Vermeer, C.; Verschuren, W.M.; Boer, J.M.; Beulens, J.W. Matrix Gla protein species and risk of cardiovascular events in type 2 diabetic patients. Diabetes Care 2013, 36, 3766–3771. [Google Scholar] [CrossRef] [Green Version]
- Jeannin, A.C.; Salem, J.E.; Massy, Z.; Aubert, C.E.; Vemeer, C.; Amouyal, C.; Phan, F.; Halbron, M.; Funck-Brentano, C.; Hartemann, A.; et al. Inactive matrix gla protein plasma levels are associated with peripheral neuropathy in Type 2 diabetes. PLoS ONE 2020, 15, e0229145. [Google Scholar] [CrossRef]
- Parker, B.D.; Ix, J.H.; Cranenburg, E.C.; Vermeer, C.; Whooley, M.A.; Schurgers, L.J. Association of kidney function and uncarboxylated matrix Gla protein: Data from the Heart and Soul Study. Nephrol. Dial. Transpl. 2009, 24, 2095–2101. [Google Scholar] [CrossRef]
- Fulzele, K.; Riddle, R.C.; DiGirolamo, D.J.; Cao, X.; Wan, C.; Chen, D.; Faugere, M.C.; Aja, S.; Hussain, M.A.; Brüning, J.C.; et al. Insulin receptor signaling in osteoblasts regulates postnatal bone acquisition and body composition. Cell 2010, 142, 309–319. [Google Scholar] [CrossRef] [Green Version]
- Faienza, M.F.; Luce, V.; Ventura, A.; Colaianni, G.; Colucci, S.; Cavallo, L.; Grano, M.; Brunetti, G. Skeleton and glucose metabolism: A bone-pancreas loop. Int. J. Endocrinol. 2015, 2015, 758148. [Google Scholar] [CrossRef] [Green Version]
- Zoch, M.L.; Clemens, T.L.; Riddle, R.C. New insights into the biology of osteocalcin. Bone 2016, 82, 42–49. [Google Scholar] [CrossRef] [Green Version]
- Moser, S.C.; van der Eerden, B.C.J. Osteocalcin-A Versatile Bone-Derived Hormone. Front. Endocrinol. (Lausanne) 2018, 9, 794. [Google Scholar] [CrossRef] [Green Version]
- Oury, F.; Sumara, G.; Sumara, O.; Ferron, M.; Chang, H.; Smith, C.E.; Hermo, L.; Suarez, S.; Roth, B.L.; Ducy, P.; et al. Endocrine regulation of male fertility by the skeleton. Cell 2011, 144, 796–809. [Google Scholar] [CrossRef] [Green Version]
- Schwetz, V.; Pieber, T.; Obermayer-Pietsch, B. The endocrine role of the skeleton: Background and clinical evidence. Eur. J. Endocrinol. 2012, 166, 959–967. [Google Scholar] [CrossRef] [Green Version]
- Oury, F.; Ferron, M.; Huizhen, W.; Confavreux, C.; Xu, L.; Lacombe, J.; Srinivas, P.; Chamouni, A.; Lugani, F.; Lejeune, H.; et al. Osteocalcin regulates murine and human fertility through a pancreas-bone-testis axis. J. Clin. Investig. 2013, 123, 2421–2433. [Google Scholar] [CrossRef]
- Karsenty, G.; Oury, F. Regulation of male fertility by the bone-derived hormone osteocalcin. Mol. Cell Endocrinol. 2014, 382, 521–526. [Google Scholar] [CrossRef] [Green Version]
- Wei, J.; Ferron, M.; Clarke, C.J.; Hannun, Y.A.; Jiang, H.; Blaner, W.S.; Karsenty, G. Bone-specific insulin resistance disrupts whole-body glucose homeostasis via decreased osteocalcin activation. J. Clin. Investig. 2014, 124, 1781–1793. [Google Scholar] [CrossRef] [Green Version]
- Lee, N.K.; Sowa, H.; Hinoi, E.; Ferron, M.; Ahn, J.D.; Confavreux, C.; Dacquin, R.; Mee, P.J.; McKee, M.D.; Jung, D.Y.; et al. Endocrine regulation of energy metabolism by the skeleton. Cell 2007, 130, 456–469. [Google Scholar] [CrossRef] [Green Version]
- Ferron, M.; Hinoi, E.; Karsenty, G.; Ducy, P. Osteocalcin differentially regulates beta cell and adipocyte gene expression and affects the development of metabolic diseases in wild-type mice. Proc. Natl. Acad. Sci. USA 2008, 105, 5266–5270. [Google Scholar] [CrossRef] [Green Version]
- Reddi, K.; Henderson, B.; Meghji, S.; Wilson, M.; Poole, S.; Hopper, C.; Harris, M.; Hodges, S.J. Interleukin 6 production by lipopolysaccharide-stimulated human fibroblasts is potently inhibited by naphthoquinone (vitamin K) compounds. Cytokine 1995, 7, 287–290. [Google Scholar] [CrossRef]
- Ohsaki, Y.; Shirakawa, H.; Hiwatashi, K.; Furukawa, Y.; Mizutani, T.; Komai, M. Vitamin K suppresses lipopolysaccharide-induced inflammation in the rat. BioSci. Biotechnol. Biochem. 2006, 70, 926–932. [Google Scholar] [CrossRef] [Green Version]
- Shea, M.K.; Booth, S.L.; Massaro, J.M.; Jacques, P.F.; D′Agostino, R.B., Sr.; Dawson-Hughes, B.; Ordovas, J.M.; O′Donnell, C.J.; Kathiresan, S.; Keaney, J.F., Jr.; et al. Vitamin K and vitamin D status: Associations with inflammatory markers in the Framingham Offspring Study. Am. J. Epidemiol. 2008, 167, 313–320. [Google Scholar] [CrossRef] [Green Version]
- Varsha, M.K.; Thiagarajan, R.; Manikandan, R.; Dhanasekaran, G. Vitamin K1 alleviates streptozotocin-induced type 1 diabetes by mitigating free radical stress, as well as inhibiting NF-κB activation and iNOS expression in rat pancreas. Nutrition 2015, 31, 214–222. [Google Scholar] [CrossRef]
- Razny, U.; Fedak, D.; Kiec-Wilk, B.; Goralska, J.; Gruca, A.; Zdzienicka, A.; Kiec-Klimczak, M.; Solnica, B.; Hubalewska-Dydejczyk, A.; Malczewska-Malec, M. Carboxylated and undercarboxylated osteocalcin in metabolic complications of human obesity and prediabetes. Diabetes Metab. Res. Rev. 2017, 33, e2862. [Google Scholar] [CrossRef]
- Juanola-Falgarona, M.; Salas-Salvadó, J.; Estruch, R.; Portillo, M.P.; Casas, R.; Miranda, J.; Martínez-González, M.A.; Bulló, M. Association between dietary phylloquinone intake and peripheral metabolic risk markers related to insulin resistance and diabetes in elderly subjects at high cardiovascular risk. Cardiovasc. Diabetol. 2013, 12, 7. [Google Scholar] [CrossRef] [Green Version]
- Wieser, V.; Moschen, A.R.; Tilg, H. Inflammation, cytokines and insulin resistance: A clinical perspective. Arch. Immunol. Exp. (Warsz) 2013, 61, 119–125. [Google Scholar] [CrossRef]
- Rehman, K.; Akash, M.S. Mechanisms of inflammatory responses and development of insulin resistance: How are they interlinked? J. Biomed. Sci. 2016, 23, 87. [Google Scholar] [CrossRef] [Green Version]
- Hotamisligil, G.S.; Murray, D.L.; Choy, L.N.; Spiegelman, B.M. Tumor necrosis factor alpha inhibits signaling from the insulin receptor. Proc. Natl. Acad. Sci. USA 1994, 91, 4854–4858. [Google Scholar] [CrossRef] [Green Version]
- Ballak, D.B.; Stienstra, R.; Tack, C.J.; Dinarello, C.A.; van Diepen, J.A. IL-1 family members in the pathogenesis and treatment of metabolic disease: Focus on adipose tissue inflammation and insulin resistance. Cytokine 2015, 75, 280–290. [Google Scholar] [CrossRef] [Green Version]
- Stefan, N. Causes, consequences, and treatment of metabolically unhealthy fat distribution. Lancet Diabetes Endocrinol. 2020, 8, 616–627. [Google Scholar] [CrossRef]
- Kleinman, R.E.; Fracchia, M.S. Vitamin K and cystic fibrosis: Give me a double, please. Am. J. Clin. Nutr. 2010, 92, 469–470. [Google Scholar] [CrossRef] [Green Version]
- Nakajima, S.; Iijima, H.; Egawa, S.; Shinzaki, S.; Kondo, J.; Inoue, T.; Hayashi, Y.; Ying, J.; Mukai, A.; Akasaka, T.; et al. Association of vitamin K deficiency with bone metabolism and clinical disease activity in inflammatory bowel disease. Nutrition 2011, 27, 1023–1028. [Google Scholar] [CrossRef]
- Sikkens, E.C.; Cahen, D.L.; Koch, A.D.; Braat, H.; Poley, J.W.; Kuipers, E.J.; Bruno, M.J. The prevalence of fat-soluble vitamin deficiencies and a decreased bone mass in patients with chronic pancreatitis. Pancreatology 2013, 13, 238–242. [Google Scholar] [CrossRef]
- Holman, R.R.; Paul, S.K.; Bethel, M.A.; Matthews, D.R.; Neil, H.A. 10-year follow-up of intensive glucose control in type 2 diabetes. N. Engl. J. Med. 2008, 359, 1577–1589. [Google Scholar] [CrossRef] [Green Version]
- Harding, J.L.; Pavkov, M.E.; Magliano, D.J.; Shaw, J.E.; Gregg, E.W. Global trends in diabetes complications: A review of current evidence. Diabetologia 2019, 62, 3–16. [Google Scholar] [CrossRef] [Green Version]
- Wei, F.F.; Huang, Q.F.; Zhang, Z.Y.; Van Keer, K.; Thijs, L.; Trenson, S.; Yang, W.Y.; Cauwenberghs, N.; Mujaj, B.; Kuznetsova, T.; et al. Inactive matrix Gla protein is a novel circulating biomarker predicting retinal arteriolar narrowing in humans. Sci. Rep. 2018, 8, 15088. [Google Scholar] [CrossRef] [Green Version]
- Sai Varsha, M.K.; Raman, T.; Manikandan, R. Inhibition of diabetic-cataract by vitamin K1 involves modulation of hyperglycemia-induced alterations to lens calcium homeostasis. Exp. Eye Res. 2014, 128, 73–82. [Google Scholar] [CrossRef]
- Thiagarajan, R.; Varsha, M.; Srinivasan, V.; Ravichandran, R.; Saraboji, K. Vitamin K1 prevents diabetic cataract by inhibiting lens aldose reductase 2 (ALR2) activity. Sci. Rep. 2019, 9, 14684. [Google Scholar] [CrossRef] [Green Version]
- Holden, R.M.; Morton, A.R.; Garland, J.S.; Pavlov, A.; Day, A.G.; Booth, S.L. Vitamins K and D status in stages 3–5 chronic kidney disease. Clin. J. Am. Soc. Nephrol. 2010, 5, 590–597. [Google Scholar] [CrossRef]
- Cranenburg, E.C.; Schurgers, L.J.; Uiterwijk, H.H.; Beulens, J.W.; Dalmeijer, G.W.; Westerhuis, R.; Magdeleyns, E.J.; Herfs, M.; Vermeer, C.; Laverman, G.D. Vitamin K intake and status are low in hemodialysis patients. Kidney Int. 2012, 82, 605–610. [Google Scholar] [CrossRef] [Green Version]
- Elliott, M.J.; Booth, S.L.; Hopman, W.M.; Holden, R.M. Assessment of potential biomarkers of subclinical vitamin K deficiency in patients with end-stage kidney disease. Can. J. Kidney Health Dis. 2014, 1, 13. [Google Scholar] [CrossRef] [Green Version]
- Epstein, M. Matrix Gla-Protein (MGP) Not Only Inhibits Calcification in Large Arteries but Also May Be Renoprotective: Connecting the Dots. EBioMedicine 2016, 4, 16–17. [Google Scholar] [CrossRef] [Green Version]
- Puzantian, H.; Akers, S.R.; Oldland, G.; Javaid, K.; Miller, R.; Ge, Y.; Ansari, B.; Lee, J.; Suri, A.; Hasmath, Z.; et al. Circulating Dephospho-Uncarboxylated Matrix Gla-Protein Is Associated with Kidney Dysfunction and Arterial Stiffness. Am. J. Hypertens 2018, 31, 988–994. [Google Scholar] [CrossRef]
- Roumeliotis, S.; Roumeliotis, A.; Panagoutsos, S.; Giannakopoulou, E.; Papanas, N.; Manolopoulos, V.G.; Passadakis, P.; Tavridou, A. Matrix Gla protein T-138C polymorphism is associated with carotid intima media thickness and predicts mortality in patients with diabetic nephropathy. J. Diabetes Complicat. 2017, 31, 1527–1532. [Google Scholar] [CrossRef]
- Schurgers, L.J.; Barreto, D.V.; Barreto, F.C.; Liabeuf, S.; Renard, C.; Magdeleyns, E.J.; Vermeer, C.; Choukroun, G.; Massy, Z.A. The circulating inactive form of matrix gla protein is a surrogate marker for vascular calcification in chronic kidney disease: A preliminary report. Clin. J. Am. Soc. Nephrol. 2010, 5, 568–575. [Google Scholar] [CrossRef] [Green Version]
- Wei, F.F.; Trenson, S.; Thijs, L.; Huang, Q.F.; Zhang, Z.Y.; Yang, W.Y.; Moliterno, P.; Allegaert, K.; Boggia, J.; Janssens, S.; et al. Desphospho-uncarboxylated matrix Gla protein is a novel circulating biomarker predicting deterioration of renal function in the general population. Nephrol. Dial. Transpl. 2018, 33, 1122–1128. [Google Scholar] [CrossRef]
- Kurnatowska, I.; Grzelak, P.; Masajtis-Zagajewska, A.; Kaczmarska, M.; Stefańczyk, L.; Vermeer, C.; Maresz, K.; Nowicki, M. Plasma Desphospho-Uncarboxylated Matrix Gla Protein as a Marker of Kidney Damage and Cardiovascular Risk in Advanced Stage of Chronic Kidney Disease. Kidney Blood Press Res. 2016, 41, 231–239. [Google Scholar] [CrossRef]
- Aoun, M.; Makki, M.; Azar, H.; Matta, H.; Chelala, D.N. High Dephosphorylated-Uncarboxylated MGP in Hemodialysis patients: Risk factors and response to vitamin K2, a pre-post intervention clinical trial. BMC Nephrol. 2017, 18, 191. [Google Scholar] [CrossRef]
- Thamratnopkoon, S.; Susantitaphong, P.; Tumkosit, M.; Katavetin, P.; Tiranathanagul, K.; Praditpornsilpa, K.; Eiam-Ong, S. Correlations of Plasma Desphosphorylated Uncarboxylated Matrix Gla Protein with Vascular Calcification and Vascular Stiffness in Chronic Kidney Disease. Nephron 2017, 135, 167–172. [Google Scholar] [CrossRef]
- Jaminon, A.M.G.; Dai, L.; Qureshi, A.R.; Evenepoel, P.; Ripsweden, J.; Soderberg, M.; Witasp, A.; Olauson, H.; Schurgers, L.J.; Stenvinkel, P. Matrix Gla protein is an independent predictor of both intimal and medial vascular calcification in chronic kidney disease. Sci. Rep. 2020, 10, 6586. [Google Scholar] [CrossRef] [Green Version]
- Doi, Y.; Iwashima, Y.; Yoshihara, F.; Kamide, K.; Hayashi, S.; Kubota, Y.; Nakamura, S.; Horio, T.; Kawano, Y. Renal resistive index and cardiovascular and renal outcomes in essential hypertension. Hypertension 2012, 60, 770–777. [Google Scholar] [CrossRef] [Green Version]
- Jaques, D.A.; Pivin, E.; Pruijm, M.; Ackermann, D.; Guessous, I.; Ehret, G.; Wei, F.F.; Staessen, J.A.; Pechère-Bertschi, A.; Vermeer, C.; et al. Renal Resistive Index Is Associated with Inactive Matrix Gla (γ-Carboxyglutamate) Protein in an Adult Population-Based Study. J. Am. Heart Assoc. 2019, 8, e013558. [Google Scholar] [CrossRef]
- Miyata, K.N.; Nast, C.C.; Dai, T.; Dukkipati, R.; LaPage, J.A.; Troost, J.P.; Schurgers, L.J.; Kretzler, M.; Adler, S.G. Renal matrix Gla protein expression increases progressively with CKD and predicts renal outcome. Exp. Mol. Pathol. 2018, 105, 120–129. [Google Scholar] [CrossRef]
- Dyck, P.J. Detection, characterization, and staging of polyneuropathy: Assessed in diabetics. Muscle Nerve 1988, 11, 21–32. [Google Scholar] [CrossRef]
- Papanas, N.; Ziegler, D. Risk Factors and Comorbidities in Diabetic Neuropathy: An Update 2015. Rev. Diabet Stud. 2015, 12, 48–62. [Google Scholar] [CrossRef] [Green Version]
- Nakajima, M.; Furukawa, S.; Hayashi, K.; Yamada, A.; Kawashima, T.; Hayashi, Y. Age-dependent survival-promoting activity of vitamin K on cultured CNS neurons. Brain Res. Dev. Brain Res. 1993, 73, 17–23. [Google Scholar] [CrossRef]
- Goritz, C.; Thiebaut, R.; Tessier, L.H.; Nieweg, K.; Moehle, C.; Buard, I.; Dupont, J.L.; Schurgers, L.J.; Schmitz, G.; Pfrieger, F.W. Glia-induced neuronal differentiation by transcriptional regulation. Glia 2007, 55, 1108–1122. [Google Scholar] [CrossRef]
- Nishimoto, S.K.; Nishimoto, M. Matrix Gla protein C-terminal region binds to vitronectin. Co-localization suggests binding occurs during tissue development. Matrix Biol. 2005, 24, 353–361. [Google Scholar] [CrossRef]
- Moon, J.I.; Birren, S.J. Target-dependent inhibition of sympathetic neuron growth via modulation of a BMP signaling pathway. Dev. Biol. 2008, 315, 404–417. [Google Scholar] [CrossRef] [Green Version]
- Nishimoto, S.K.; Nishimoto, M. Matrix gla protein binds to fibronectin and enhances cell attachment and spreading on fibronectin. Int. J. Cell Biol. 2014, 2014, 807013. [Google Scholar] [CrossRef] [Green Version]
- Kendall, D.M.; Harmel, A.P. The metabolic syndrome, type 2 diabetes, and cardiovascular disease: Understanding the role of insulin resistance. Am. J. Manag. Care 2002, 8, S635–S653. [Google Scholar]
- Schurgers, L.J.; Cranenburg, E.C.; Vermeer, C. Matrix Gla-protein: The calcification inhibitor in need of vitamin K. Thromb. Haemost. 2008, 100, 593–603. [Google Scholar]
- Shea, M.K.; Holden, R.M. Vitamin K status and vascular calcification: Evidence from observational and clinical studies. Adv. Nutr. 2012, 3, 158–165. [Google Scholar] [CrossRef] [Green Version]
- Brandenburg, V.M.; Reinartz, S.; Kaesler, N.; Kruger, T.; Dirrichs, T.; Kramann, R.; Peeters, F.; Floege, J.; Keszei, A.; Marx, N.; et al. Slower Progress of Aortic Valve Calcification with Vitamin K Supplementation: Results from a Prospective Interventional Proof-of-Concept Study. Circulation 2017, 135, 2081–2083. [Google Scholar] [CrossRef]
- Pivin, E.; Ponte, B.; Pruijm, M.; Ackermann, D.; Guessous, I.; Ehret, G.; Liu, Y.P.; Drummen, N.E.; Knapen, M.H.; Pechere-Bertschi, A.; et al. Inactive Matrix Gla-Protein Is Associated with Arterial Stiffness in an Adult Population-Based Study. Hypertension 2015, 66, 85–92. [Google Scholar] [CrossRef] [Green Version]
- Harshman, S.G.; Shea, M.K. The Role of Vitamin K in Chronic Aging Diseases: Inflammation, Cardiovascular Disease, and Osteoarthritis. Curr. Nutr. Rep. 2016, 5, 90–98. [Google Scholar] [CrossRef] [Green Version]
- Nagata, C.; Wada, K.; Tamura, T.; Konishi, K.; Goto, Y.; Koda, S.; Kawachi, T.; Tsuji, M.; Nakamura, K. Dietary soy and natto intake and cardiovascular disease mortality in Japanese adults: The Takayama study. Am. J. Clin. Nutr. 2017, 105, 426–431. [Google Scholar] [CrossRef] [Green Version]
- Caluwe, R.; Pyfferoen, L.; De Boeck, K.; De Vriese, A.S. The effects of vitamin K supplementation and vitamin K antagonists on progression of vascular calcification: Ongoing randomized controlled trials. Clin. Kidney J. 2016, 9, 273–279. [Google Scholar] [CrossRef]
- Danziger, J.; Young, R.L.; Shea, K.M.; Duprez, D.A.; Jacobs, D.R.; Tracy, R.P.; Ix, J.H.; Jenny, N.S.; Mukamal, K.J. Circulating Des-gamma-carboxy prothrombin is not associated with cardiovascular calcification or stiffness: The Multi-Ethnic Study of Atherosclerosis (MESA). Atherosclerosis 2016, 252, 68–74. [Google Scholar] [CrossRef]
- Danziger, J.; Young, R.L.; Shea, M.K.; Tracy, R.P.; Ix, J.H.; Jenny, N.S.; Mukamal, K.J. Vitamin K-Dependent Protein Activity and Incident Ischemic Cardiovascular Disease: The Multi-Ethnic Study of Atherosclerosis. Arter. Thromb. Vasc. Biol. 2016, 36, 1037–1042. [Google Scholar] [CrossRef] [Green Version]
- Ishida, Y.; Kawai, S. Comparative efficacy of hormone replacement therapy, etidronate, calcitonin, alfacalcidol, and vitamin K in postmenopausal women with osteoporosis: The Yamaguchi Osteoporosis Prevention Study. Am. J. Med. 2004, 117, 549–555. [Google Scholar] [CrossRef]
- Hofbauer, L.C.; Brueck, C.C.; Singh, S.K.; Dobnig, H. Osteoporosis in patients with diabetes mellitus. J. Bone Min. Res. 2007, 22, 1317–1328. [Google Scholar] [CrossRef]
- Shiraki, M.; Yamazaki, Y.; Shiraki, Y.; Hosoi, T.; Tsugawa, N.; Okano, T. High level of serum undercarboxylated osteocalcin in patients with incident fractures during bisphosphonate treatment. J. Bone Min. Metab. 2010, 28, 578–584. [Google Scholar] [CrossRef]
- Palermo, A.; Tuccinardi, D.; D′Onofrio, L.; Watanabe, M.; Maggi, D.; Maurizi, A.R.; Greto, V.; Buzzetti, R.; Napoli, N.; Pozzilli, P.; et al. Vitamin K and osteoporosis: Myth or reality? Metabolism 2017, 70, 57–71. [Google Scholar] [CrossRef] [Green Version]
- Iwamoto, J.; Seki, A.; Sato, Y.; Matsumoto, H.; Takeda, T.; Yeh, J.K. Vitamin K2 prevents hyperglycemia and cancellous osteopenia in rats with streptozotocin-induced type 1 diabetes. Calcif. Tissue Int. 2011, 88, 162–168. [Google Scholar] [CrossRef]
- Iwamoto, J.; Takeda, T.; Ichimura, S. Effect of combined administration of vitamin D3 and vitamin K2 on bone mineral density of the lumbar spine in postmenopausal women with osteoporosis. J. Orthop. Sci. 2000, 5, 546–551. [Google Scholar] [CrossRef]
- Shiraki, M.; Shiraki, Y.; Aoki, C.; Miura, M. Vitamin K2 (menatetrenone) effectively prevents fractures and sustains lumbar bone mineral density in osteoporosis. J. Bone Min. Res. 2000, 15, 515–521. [Google Scholar] [CrossRef]
- Iwamoto, J.; Takeda, T.; Ichimura, S. Effect of menatetrenone on bone mineral density and incidence of vertebral fractures in postmenopausal women with osteoporosis: A comparison with the effect of etidronate. J. Orthop. Sci. 2001, 6, 487–492. [Google Scholar] [CrossRef]
- Inoue, T.; Fujita, T.; Kishimoto, H.; Makino, T.; Nakamura, T.; Nakamura, T.; Sato, T.; Yamazaki, K. Randomized controlled study on the prevention of osteoporotic fractures (OF study): A phase IV clinical study of 15-mg menatetrenone capsules. J. Bone Min. Metab. 2009, 27, 66–75. [Google Scholar] [CrossRef]
- Iwamoto, J. Vitamin K2 therapy for postmenopausal osteoporosis. Nutrients 2014, 6, 1971–1980. [Google Scholar] [CrossRef] [Green Version]
- Beulens, J.W.; Booth, S.L.; van den Heuvel, E.G.; Stoecklin, E.; Baka, A.; Vermeer, C. The role of menaquinones (vitamin K(2)) in human health. Br. J. Nutr. 2013, 110, 1357–1368. [Google Scholar] [CrossRef] [Green Version]
- Vissers, L.E.; Dalmeijer, G.W.; Boer, J.M.; Monique Verschuren, W.M.; van der Schouw, Y.T.; Beulens, J.W. Intake of dietary phylloquinone and menaquinones and risk of stroke. J. Am. Heart Assoc. 2013, 2, e000455. [Google Scholar] [CrossRef] [Green Version]



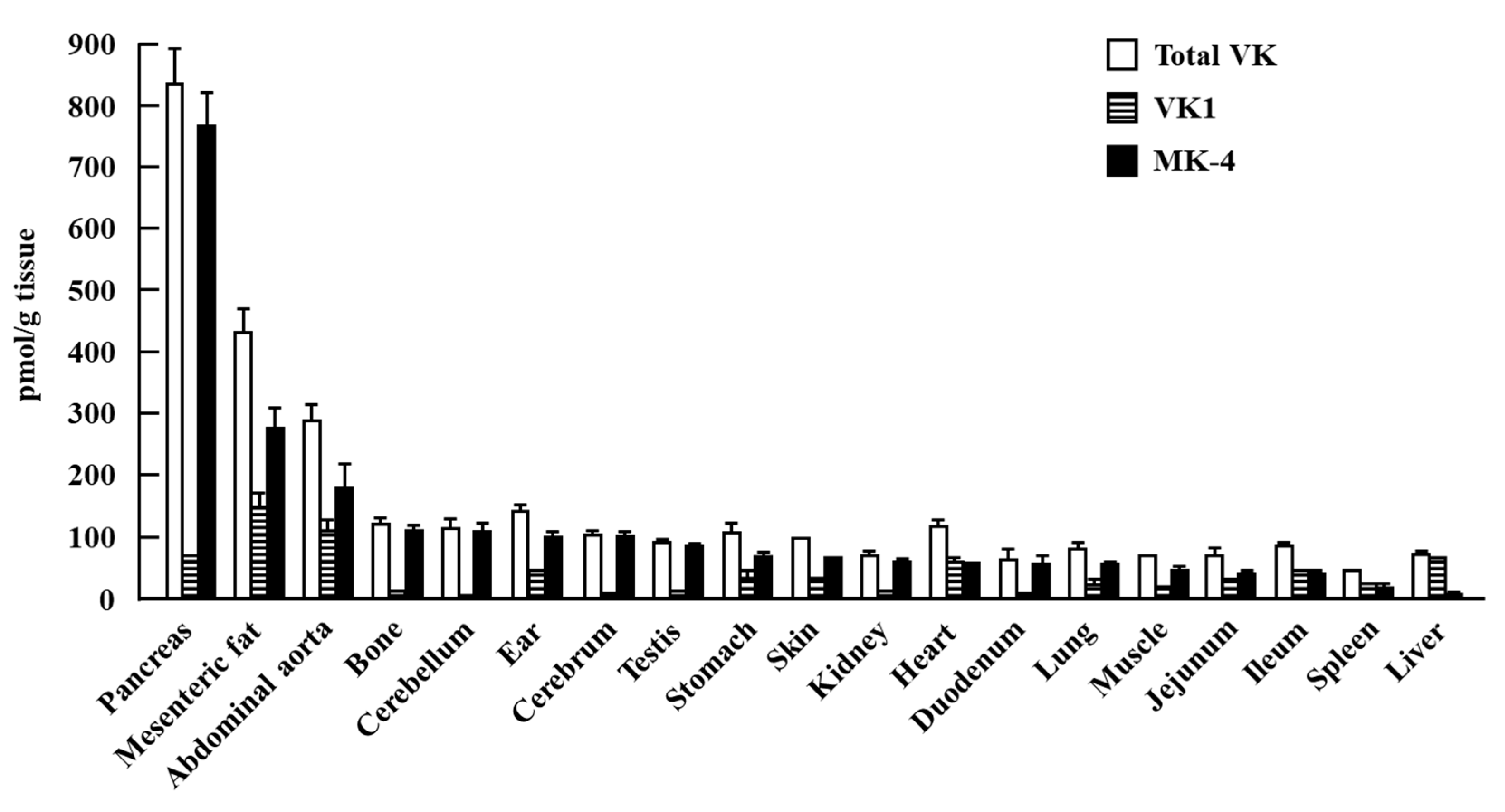{kind=link}
{kind=link}
| Subjects (N) | VK dose/VK Status | Period | Outcome | Ref. |
|---|---|---|---|---|
| A. Human Studies | ||||
| (a) Observational Studies | ||||
| Healthy young men (16) | Usual dietary intake | Acute insulin response A 1-week food-frequency questionnaire to ascertain the daily VK intake | The participants with higher dietary VK intake showed a better insulin response and glucose tolerance. | [18] |
| Framingham offspring cohort study, adult men (1247) and women (1472) | Usual dietary intake | 12 months | In a cross-sectional analysis, higher dietary VK intake was associated with reduced insulin resistance in both adult men and women. | [19] |
| Adult men (9740) and women (28,354) | Usual dietary intake | 10.3 years | Dietary intake of both VK1 and VK2 were associated with a reduced risk of T2DM. | [20] |
| Elderly men (861) and women (1062) with high cardiovascular risk | Usual dietary intake | Median follow-up of 5.5 years | Dietary VK1 at the baseline was significantly lower in participants who developed T2DM during the study. Increased dietary VK1 intake was associated with a reduced risk of incident T2DM. | [21] |
| European Prospective Investigation into Cancer and Nutrition (EPIC) cohort study, Diabetes Genetics Replication and Meta-analysis (DIAGRAM), and the UK Biobank (9400 case subjects and 12,182 sub-cohort participants) | Usual dietary intake | EPIC cohort: 1997–2007, DIAGRAM cohort: 2007 (included data from 23 studies), UK Biobank: 2006–2010 | Higher circulating VK1 may be causally related with lower risk of T2DM, highlighting the importance of sufficient phylloquinone intake in the human diet. | [22] |
| (b) Intervention Studies | ||||
| Elderly nondiabetic men (124) and women (165) | With or without 500 μg/day VK1 supplementation | 36 months | Dietary VK1 supplementation had a protective effect on the progression of insulin resistance in older men. | [23] |
| Prediabetic women (82) | With or without 1000 μg/day VK1 supplementation | 4 weeks | Dietary VK1 supplementation had beneficial effects on glycemic status and insulin sensitivity in premenopausal and prediabetic women. | [24,25] |
| Healthy young men (12) | 90 mg/day menaquinone-4 (MK-4) supplementation | 1 week | Short-term VK2 supplementation improved the insulin response after an oral glucose challenge in young men. | [26] |
| Healthy young men (42) | With or without 90 mg/day MK-4 supplementation | 4 weeks | Dietary VK2 supplementation improved insulin sensitivity in young men. | [27] |
| B. Animal Studies | ||||
| Rats (unknown) | Low-VK diet (<20% of the required VK1) | Unknown | Rats fed a low-VK diet had poor early insulin response and subsequently increased insulin secretion after a glucose load. | [28] |
| Arteriosclerotic rat model with DM (unknown) | 100 mg/day per kilogram body weight VK2 | 3 or 6 weeks | VK2 supplementation had a protective effect on arteriosclerosis, by decreasing the aortic Ca and P and the elastin fraction. Rats fed a VK2-rich diet had decreased serum glucose levels and increased serum insulin levels. | [29] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ho, H.-J.; Komai, M.; Shirakawa, H. Beneficial Effects of Vitamin K Status on Glycemic Regulation and Diabetes Mellitus: A Mini-Review. Nutrients 2020, 12, 2485. https://doi.org/10.3390/nu12082485
Ho H-J, Komai M, Shirakawa H. Beneficial Effects of Vitamin K Status on Glycemic Regulation and Diabetes Mellitus: A Mini-Review. Nutrients. 2020; 12(8):2485. https://doi.org/10.3390/nu12082485
Chicago/Turabian StyleHo, Hsin-Jung, Michio Komai, and Hitoshi Shirakawa. 2020. "Beneficial Effects of Vitamin K Status on Glycemic Regulation and Diabetes Mellitus: A Mini-Review" Nutrients 12, no. 8: 2485. https://doi.org/10.3390/nu12082485
APA StyleHo, H. -J., Komai, M., & Shirakawa, H. (2020). Beneficial Effects of Vitamin K Status on Glycemic Regulation and Diabetes Mellitus: A Mini-Review. Nutrients, 12(8), 2485. https://doi.org/10.3390/nu12082485