Time Course of Salivary Protein Responses to Cranberry-Derived Polyphenol Exposure as a Function of PROP Taster Status


 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subject Recruitment
2.2. PROP Taster Status
2.3. Test Stimuli
2.4. Experimental Procedures
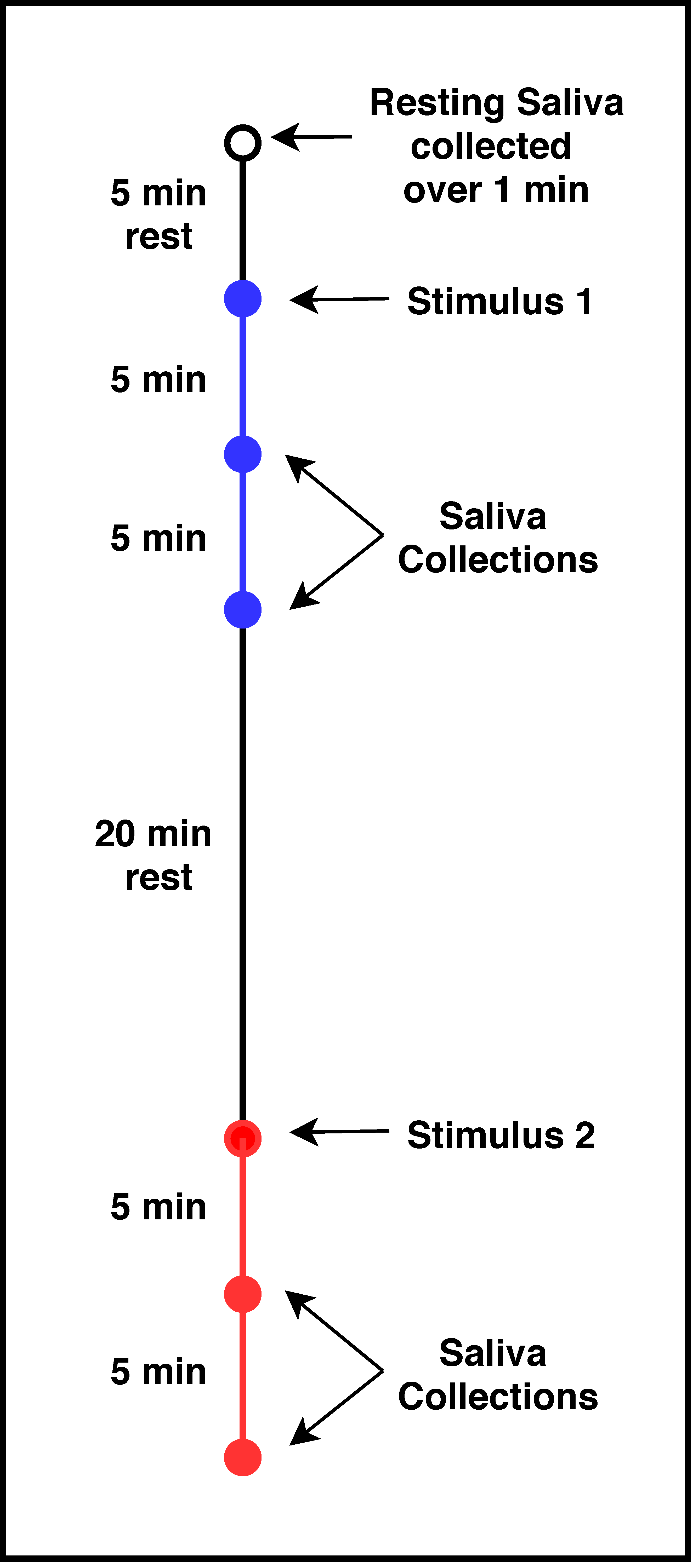
2.4.1. Overall Study Design
2.4.2. Saliva Treatment
2.4.3. Total Protein Content Quantification
2.4.4. Alpha Amylase Analysis
Alpha Amylase Purification
Alpha Amylase Quantification
2.4.5. HPLC-Low Resolution-ESI-IT-MS Analysis
2.5. Statistical Analyses
3. Results
3.1. Effect of Stimulation on Protein Response
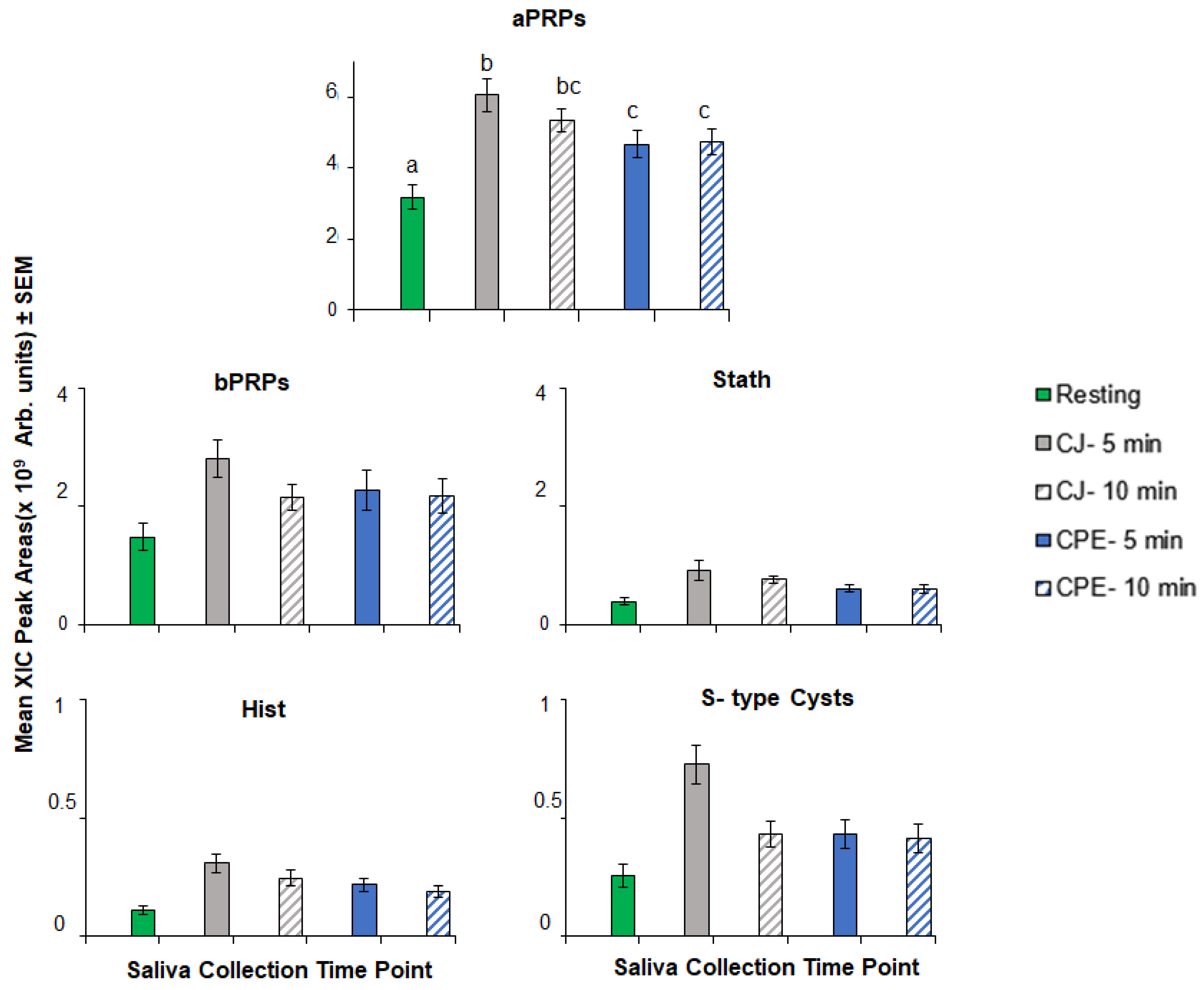
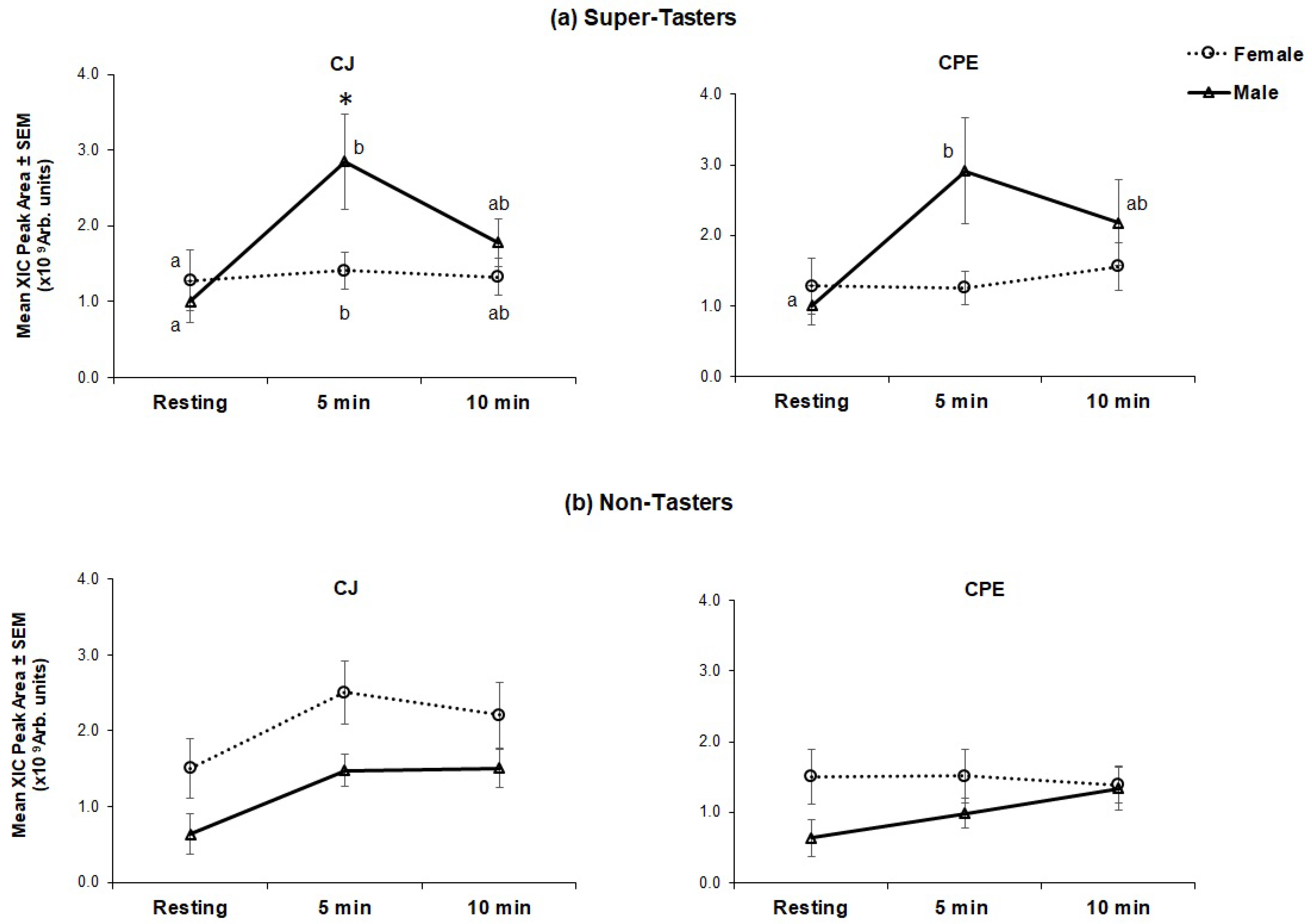
3.1.1. Protein Families
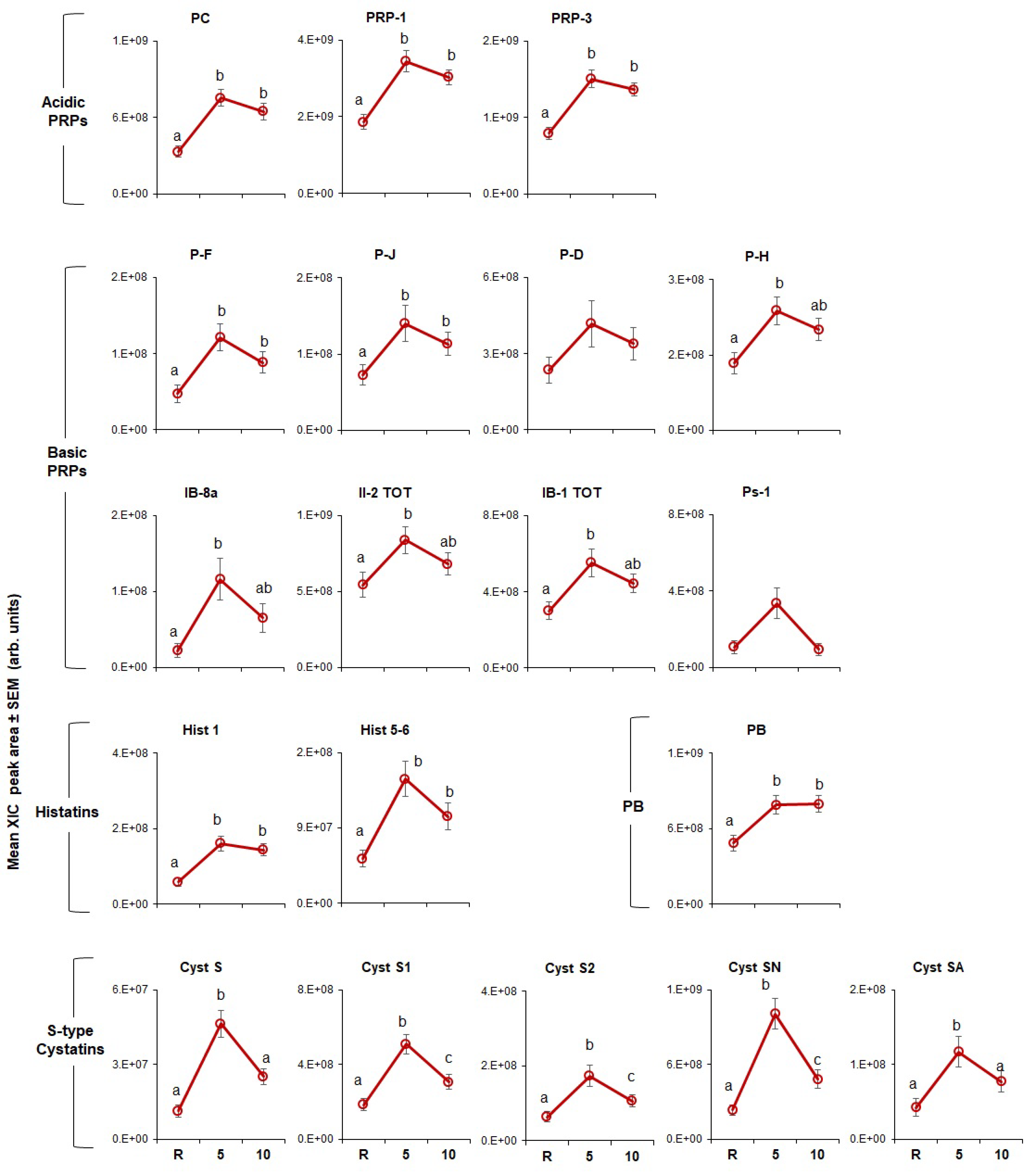
3.1.2. Individual Proteins
3.1.3. Alpha Amylase
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lee, C.B.; Lawless, H.T. Time-course of astringent sensations. Chem. Senses 1991, 16, 225–238. [Google Scholar]
- Guinard, J.; Pangborn, R.M.; Lewis, M.J. The time-course of astringency in wine upon repeated ingestion. Am. J. Enol. Vitic. 1986, 37, 184–189. [Google Scholar]
- Lyman, B.J.; Green, B.G. Oral astringency: Effects of repeated exposure and interactions with sweeteners. Chem. Senses 1990, 15, 151–164. [Google Scholar] [CrossRef]
- Dinnella, C.; Recchia, A.; Tuorila, H.; Monteleone, E. Individual astringency responsiveness affects the acceptance of phenol-rich foods. Appetite 2011, 56, 633–642. [Google Scholar] [CrossRef]
- Torri, L.; Piochi, M.; Marchiani, R.; Zeppa, G.; Dinnella, C.; Monteleone, E. A sensory- and consumer-based approach to optimize cheese enrichment with grape skin powders. J. Dairy Sci. 2016, 99, 194–204. [Google Scholar] [CrossRef] [Green Version]
- Jaeger, S.R.; Axten, L.G.; Wohlers, M.W.; Sun-Waterhouse, D. Polyphenol-rich beverages: Insights from sensory and consumer science. J. Sci. Food Agric. 2009, 89, 2356–2363. [Google Scholar] [CrossRef]
- Zhong, H.; Xue, Y.; Lu, X.; Shao, Q.; Cao, Y.; Wu, Z.; Chen, G. The effects of different degrees of procyanidin polymerization on the nutrient absorption and digestive enzyme activity in mice. Molecules 2018, 23, 2916. [Google Scholar]
- Mehansho, H.; Ann, D.K.; Butler, L.G.; Rogler, J.; Carlson, D.M. Induction of proline-rich proteins in hamster salivary glands by isoproterenol treatment and an unusual growth inhibition by tannins. J. Biol. Chem. 1987, 262, 12344–12350. [Google Scholar]
- Kondo, M.; Hirano, Y.; Ikai, N.; Kita, K.; Jayanegara, A.; Yokota, H. Assessment of anti-nutritive activity of tannins in tea by-products based on in vitro rumen fermentation. Asian Australas. J. Anim. Sci. 2014, 27, 1571–1576. [Google Scholar] [CrossRef] [Green Version]
- Durazzo, A.; Lucarini, M.; Souto, E.; Cicala, C.; Caiazzo, E.; Izzo, A.; Novellino, E.; Santini, A. Polyphenols: A concise overview on the chemistry, occurrence, and human health. Phytother. Res. 2019, 33, 2221–2243. [Google Scholar]
- Saibandith, B.; Spencer, J.P.E.; Rowland, I.R.; Commane, D.M. Olive polyphenols and the metabolic syndrome. Molecules 2017, 22, 1082. [Google Scholar] [CrossRef] [Green Version]
- Cory, H.; Passarelli, S.; Szeto, J.; Tamez, M.; Mattei, J. The role of polyphenols in human health and food systems: A mini-review. Front. Nutr. 2018, 5, 87. [Google Scholar] [CrossRef] [Green Version]
- Jobstl, E.; O’Connell, J.; Fairclough, P.A.; Williamson, M.P. Molecular model for astringency produced by polyphenol/protein interactions. Biomacromolecules 2004, 5, 942–949. [Google Scholar]
- Charlton, A.J.; Baxter, N.J.; Khan, M.L.; Moir, A.J.G.; Haslam, E.; Davies, A.P.; Williamson, M.P. Polyphenol/Peptide binding and precipitation. J. Agric. Food Chem. 2002, 50, 1593–1601. [Google Scholar]
- Siebert, K.J.; Troukhanova, N.V.; Lynn, P.Y. Nature of protein polyphenol interactions. J. Agric. Food Chem. 1996, 44, 80–85. [Google Scholar]
- Ployon, S.; Morzel, M.; Belloir, C.; Bonnotte, A.; Bourillot, E.; Briand, L.; Lesniewska, E.; Lherminier, J.; Aybeke, E.; Canon, F. Mechanisms of astringency: Structural alteration of the oral mucosal pellicle by dietary tannins and protective effect of bPRPs. Food Chem. 2018, 253, 79–87. [Google Scholar] [CrossRef]
- Frenkel, E.S.; Ribbeck, K. Salivary mucins protect surfaces from colonization by cariogenic bacteria. Appl. Environ. Microbiol. 2015, 81, 332–338. [Google Scholar] [CrossRef] [Green Version]
- Nayak, A.; Carpenter, G.H. A physiological model of tea-induced astringency. Physiol. Behav. 2008, 95, 290–294. [Google Scholar] [CrossRef]
- Horne, J.; Hayes, J.; Lawless, H.T. Turbidity as a measure of salivary protein reactions with astringency substances. Chem. Senses 2002, 27, 653–659. [Google Scholar]
- Monteleone, E.; Condelli, N.; Dinnella, C.; Bertuccioli, M. Prediction of perceived astringency induced by phenolic compounds. Food Qual. Preference 2004, 15, 761–769. [Google Scholar] [CrossRef]
- Pascal, C.; Poncet-Legrand, C.; Cabane, B.; Vernhet, A. Aggregation of a proline-rich protein induced by epigallocatechin gallate and condensed tannins: Effect of protein glycosylation. J. Agric. Food Chem. 2008, 56, 6724–6732. [Google Scholar]
- Soares, S.; Vitorino, R.; Osorio, H.; Fernandes, A.; Venancio, A.; Mateus, N.; Amado, F.; de Freitas, V. Reactivity of human salivary proteins families toward food polyphenols. J. Agric. Food Chem. 2011, 59, 5535–5547. [Google Scholar] [CrossRef]
- Soares, S.; Mateus, N.; de Freitas, V. Carbohydrates inhibit salivary proteins precipitation by condensed tannins. J. Agric. Food Chem. 2012, 60, 3966–3972. [Google Scholar] [CrossRef]
- Soares, S.; Santos Silva, M.; Garcia-Estevez, I.; Brandao, E.; Fonseca, F.; Ferreira-da-Silva, F.; Teresa Escribano-Bailon, M.; Mateus, N.; de Freitas, V. Effect of malvidin-3-glucoside and epicatechin interaction on their ability to interact with salivary proline-rich proteins. Food Chem. 2019, 276, 33–42. [Google Scholar] [CrossRef]
- Ramos-Pineda, A.M.; Garcia-Estevez, I.; Soares, S.; de Freitas, V.; Duenas, M.; Escribano-Bailon, M.T. Synergistic effect of mixture of two proline-rich-protein salivary families (aPRP and bPRP) on the interaction with wine flavanols. Food Chem. 2019, 272, 210–215. [Google Scholar] [CrossRef]
- Ramos-Pineda, A.M.; Carpenter, G.H.; Garcia-Estevez, I.; Escribano-Bailon, M.T. Influence of chemical species on polyphenol-protein interactions related to wine astringency. J. Agric. Food Chem. 2020, 68, 2948–2954. [Google Scholar] [CrossRef]
- Silva, M.S.; Garcia-Estevez, I.; Brandao, E.; Mateus, N.; de Freitas, V.; Soares, S. Molecular interaction between salivary proteins and food tannins. J. Agric. Food Chem. 2017, 65, 6415–6424. [Google Scholar] [CrossRef]
- Brandao, E.; Soares, S.; Mateus, N.; de Freitas, V. Human saliva protein profile: Influence of food ingestion. Food Res. Int. 2014, 64, 508–513. [Google Scholar] [CrossRef]
- Brandao, E.; Silva, M.S.; Garcia-Estevez, I.; Williams, P.; Mateus, N.; Doco, T.; de Freitas, V.; Soares, S. The role of wine polysaccharides on salivary protein-tannin interaction: A molecular approach. Carbohydr. Polym. 2017, 177, 77–85. [Google Scholar] [CrossRef]
- Naish, M.; Clifford, M.N.; Birch, G.G. Sensory astringency of 5-0-caffeoylquinic acid, tannic acid and grape-seed tannin by a time-intensity procedure. J. Sci. Food Agric. 1993, 61, 57–64. [Google Scholar]
- Valentova, H.; Skrovankova, S.; Panovska, Z.; Pokorny, J. Time course of astringent taste. Food Chem. 2002, 78, 29–37. [Google Scholar]
- Bajec, M.R.; Pickering, G.J. Astringency: Mechanisms and perception. Crit. Rev. Food Sci. Nutr. 2008, 48, 858–875. [Google Scholar] [CrossRef]
- Brandao, E.; Soares, S.; Mateus, N.; de Freitas, V. In vivo interactions between procyanidins and human saliva proteins: Effect of repeated exposures to procyanidins solution. J. Agric. Food Chem. 2014, 62, 9562–9568. [Google Scholar] [CrossRef]
- Peleg, H.; Gacon, K.; Schlich, P.; Noble, A.C. Bitterness and astringency of flavan-3-ol monomers, dimers and trimers. J. Sci. Food Agric. 1999, 79, 1123–1128. [Google Scholar]
- Hufnagel, J.C.; Hoffman, T. Orosensory-directed identification of astringent mouthfeel and bitter-tasting compounds in red wine. J. Agric. Food Chem. 2008, 56, 1376–1386. [Google Scholar]
- Fleming, E.E.; Ziegler, G.R.; Hayes, J.E. Check-All-That-Apply (CATA), sorting, and polarized sensory positioning (PSP) with astringent stimuli. Food Qual. Prefer. 2015, 45, 41–49. [Google Scholar] [CrossRef] [Green Version]
- Kershaw, J.C.; Running, C.A. Dose-response functions and methodological insights for sensory tests with astringent stimuli. J. Sens. Stud. 2019, 34. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.A.; Vickers, Z.M. Astringency of foods may not be directly related to salivary lubricity. J. Food Sci. 2012, 77, S302–S306. [Google Scholar] [CrossRef]
- Schobel, N.; Radtke, D.; Kyereme, J.; Wollmann, N.; Cichy, A.; Obst, K.; Kallweit, K.; Kletke, O.; Minovi, A.; Dazert, S.; et al. Astringency is a trigeminal sensation that involves the activation of G protein-coupled signaling by phenolic compounds. Chem. Senses 2014, 39, 471–487. [Google Scholar] [CrossRef]
- Soares, S.; Kohl, S.; Thalmann, S.; Mateus, N.; Meyerhof, W.; De Freitas, V. Different phenolic compounds activate distinct human bitter taste receptors. J. Agric. Food Chem. 2013, 61, 1525–1533. [Google Scholar] [CrossRef]
- Soares, S.; Silva, M.S.; Garcia-Estevez, I.; Grobetamann, P.; Bras, N.; Brandao, E.; Mateus, N.; de Freitas, V.; Behrens, M.; Meyerhof, W. Human bitter taste receptors are activated by different classes of polyphenols. J. Agric. Food Chem. 2018, 66, 8814–8823. [Google Scholar] [CrossRef]
- Dinnella, C.; Recchia, A.; Fia, G.; Bertuccioli, M.; Monteleone, E. Saliva characteristics and individual sensitivity to phenolic astringent stimuli. Chem. Senses 2009, 34, 295–304. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.Q.; Zhang, Y.N.; Chen, J.X.; Wang, F.; Du, Q.Z.; Yin, J.F. Quantitative analyses of the bitterness and astringency of catechins from green tea. Food Chem. 2018, 258, 16–24. [Google Scholar] [CrossRef]
- Dinnella, C.; Recchia, A.; Vincenzi, S.; Tuorila, H.; Monteleone, E. Temporary modification of salivary protein profile and individual responses to repeated phenolic astringent stimuli. Chem. Senses 2010, 35, 75–85. [Google Scholar] [CrossRef] [Green Version]
- Tepper, B.J. Nutritional implications of genetic taste variation: The role of PROP sensitivity and other taste phenotypes. Annu. Rev. Nutr. 2008, 28, 367–388. [Google Scholar] [CrossRef]
- Pickering, G.J.; Robert, G. Perception of mouthfeel sensations elicited by red wine are associated with sensitivity to 6-n-propothiouracil. J. Sens. Stud. 2006, 21, 249–265. [Google Scholar]
- Pickering, G.J.; Simunkova, K.; DiBattista, D. Intensity of taste and astringency sensations elicited by red wines is associated with sensitivity to PROP (6-n-propylthiouracil). Food Qual. Prefer. 2004, 15, 147–154. [Google Scholar] [CrossRef]
- Tan, J.; Abrol, R.; Trzaskowski, B.; Goddard, W.A. 3D structure prediction of TAS2R38 bitter receptors bound to agonists phenylthiocarbamide (PTC) and 6-n-propylthiouracil (PROP). J. Chem. Inf. Model. 2012, 52, 1875–1885. [Google Scholar] [CrossRef] [Green Version]
- Dinehart, M.E.; Hayes, J.E.; Bartoshuk, L.M.; Lanier, S.L.; Duffy, V.B. Bitter taste markers explain variability in vegetable sweetness, bitterness, and intake. Physiol. Behav. 2006, 87, 304–313. [Google Scholar] [CrossRef]
- Sandell, M.; Hoppu, U.; Lundén, S.; Salminen, M.; Puolimatka, T.; Laaksonen, O.; Laitinen, K.; Hopia, A. Consumption of lingonberries by TAS2R38 genotype and sensory quality of texture-designed lingonberry samples. Food Qual. Prefer. 2015, 45, 166–170. [Google Scholar] [CrossRef]
- Laaksonen, O.; Ahola, J.; Sandell, M. Explaining and predicting individually experienced liking of berry fractions by the hTAS2R38 taste receptor genotype. Appetite 2013, 61, 85–96. [Google Scholar] [CrossRef]
- Melis, M.; Yousaf, N.Y.; Mattes, M.Z.; Cabras, T.; Messana, I.; Crnjar, R.; Tomassini Barbarossa, I.; Tepper, B.J. Sensory perception of and salivary protein response to astringency as a function of the 6-n-propylthioural (PROP) bitter-taste phenotype. Physiol. Behav. 2017, 173, 163–173. [Google Scholar] [CrossRef]
- Blumberg, J.B.; Camesano, T.A.; Cassidy, A.; Kris-Etherton, P.; Howell, A.; Manach, C.; Ostertag, L.M.; Sies, H.; Skulas-Ray, A.; Vita, J.A. Cranberries and their bioactive constituents in human health. Adv. Nutr. 2013, 4, 618–632. [Google Scholar] [CrossRef] [Green Version]
- Feeney, E.L.; Hayes, J.E. Exploring associations between taste perception, oral anatomy and polymorphisms in the carbonic anhydrase (gustin) gene CA6. Physiol. Behav. 2014, 128, 148–154. [Google Scholar] [CrossRef] [Green Version]
- Törnwall, O.; Dinnella, C.; Keskitalo-Vuokko, K.; Silventoinen, K.; Perola, M.; Monteleone, E.; Kaprio, J.; Tuorila, H. Astringency perception and heritability among young Finnish twins. Chemosens. Percept. 2011, 4, 134–144. [Google Scholar] [CrossRef]
- Duffy, V.B.; Rawal, S.; Park, J.; Brand, M.H.; Sharafi, M.; Bolling, B.W. Characterizing and improving the sensory and hedonic responses to polyphenol-rich aronia berry juice. Appetite 2016, 107, 116–125. [Google Scholar] [CrossRef] [Green Version]
- De Toffoli, A.; Spinelli, S.; Monteleone, E.; Arena, E.; Di Monaco, R.; Endrizzi, I.; Gallina Toschi, T.; Laureati, M.; Napolitano, F.; Torri, L.; et al. Influences of psychological traits and PROP taster status on familiarity with and choice of phenol-rich foods and beverages. Nutrients 2019, 11, 1329. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Kirkmeyer, S.V.; Tepper, B.J. A paper screening test to assess genetic taste sensitivity to 6-n-propylthiouracil. Physiol. Behav. 2003, 78, 625–633. [Google Scholar] [CrossRef]
- Tomassini Barbarossa, I.; Melis, M.; Mattes, M.Z.; Calo, C.; Muroni, P.; Crnjar, R.; Tepper, B.J. The gustin (CA6) gene polymorphism, rs2274333 (A/G), is associated with fungiform papilla density, whereas PROP bitterness is mostly due to TAS2R38 in an ethnically-mixed population. Physiol. Behav. 2015, 138, 6–12. [Google Scholar] [CrossRef]
- Melis, M.; Tomassini Barbarossa, I. Taste perception of sweet, sour, salty, bitter, and umami and changes due to l-Arginine supplementation, as a function of genetic ability to taste 6-n-propylthiouracil. Nutrients 2017, 9, 541. [Google Scholar] [CrossRef] [Green Version]
- Melis, M.; Mastinu, M.; Arca, M.; Crnjar, R.; Tomassini Barbarossa, I. Effect of chemical interaction between oleic acid and L-Arginine on oral perception, as a function of polymorphisms of CD36 and OBPIIa and genetic ability to taste 6-n-propylthiouracil. PLoS ONE 2018, 13, e0194953. [Google Scholar] [CrossRef]
- Carta, G.; Melis, M.; Pintus, S.; Pintus, P.; Piras, C.A.; Muredda, L.; Demurtas, D.; Di Marzo, V.; Banni, S.; Tomassini Barbarossa, I. Participants with normal weight or with obesity show different relationships of 6-n-Propylthiouracil (PROP) taster status with BMI and plasma endocannabinoids. Sci. Rep. 2017, 7, 1361. [Google Scholar] [CrossRef] [Green Version]
- Sollai, G.; Melis, M.; Pani, D.; Cosseddu, P.; Usai, I.; Crnjar, R.; Bonfiglio, A.; Tomassini Barbarossa, I. First objective evaluation of taste sensitivity to 6-n-propylthiouracil (PROP), a paradigm gustatory stimulus in humans. Sci. Rep. 2017, 7, 40353. [Google Scholar] [CrossRef] [Green Version]
- Pani, D.; Usai, I.; Cosseddu, P.; Melis, M.; Sollai, G.; Crnjar, R.; Tomassini Barbarossa, I.; Raffo, L.; Bonfiglio, A. An automated system for the objective evaluation of human gustatory sensitivity using tongue biopotential recordings. PLoS ONE 2017, 12, e0177246. [Google Scholar] [CrossRef] [Green Version]
- Cabras, T.; Melis, M.; Castagnola, M.; Padiglia, A.; Tepper, B.J.; Messana, I.; Tomassini Barbarossa, I. Responsiveness to 6-n-propylthiouracil (PROP) is associated with salivary levels of two specific basic proline-rich proteins in humans. PLoS ONE 2012, 7, e30962. [Google Scholar] [CrossRef] [Green Version]
- Messana, I.; Cabras, T.; Pisano, E.; Sanna, M.T.; Olianas, A.; Manconi, B.; Pellegrini, M.; Paludetti, G.; Scarano, E.; Fiorita, A.; et al. Trafficking and postsecretory events responsible for the formation of secreted human salivary peptides: A proteomics approach. Mol. Cell Proteom. 2008, 7, 911–926. [Google Scholar] [CrossRef] [Green Version]
- Castagnola, M.; Cabras, T.; Iavarone, F.; Vincenzoni, F.; Vitali, A.; Pisano, E.; Nemolato, S.; Scarano, E.; Fiorita, A.; Vento, G.; et al. Top-down platform for deciphering the human salivary proteome. J. Matern.-Fetal Neonatal Med. 2012, 25, 27–43. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Marshall, A.G. A universal algorithm for fast and automated charge state deconvolution of electrospray mass-to-charge ratio spectra. J. Am. Soc. Mass Spectrom. 1998, 9, 225–233. [Google Scholar] [CrossRef] [Green Version]
- Ong, S.E.; Mann, M. Mass spectrometry-based proteomics turns quantitative. Nat. Chem. Biol. 2005, 1, 252–262. [Google Scholar] [CrossRef]
- Lu, Y.; Bennick, A. Interaction of tanni with human salivary proline-rich proteins. Arch. Oral Biol. 1998, 43, 717–728. [Google Scholar]
- Soares, S.; Sousa, A.; Mateus, N.; de Freitas, V. Effect of condensed tannins addition on the astringency of red wines. Chem. Senses 2012, 37, 191–198. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Estevez, I.; Cruz, L.; Oliveira, J.; Mateus, N.; de Freitas, V.; Soares, S. First evidences of interaction between pyranoanthocyanins and salivary proline-rich proteins. Food Chem. 2017, 228, 574–581. [Google Scholar] [CrossRef]
- Neyraud, E.; Sayd, T.; Morzel, M.; Dransfield, E. Proteomic analysis of human whole and parotid salivas following stimulation by different tastes. J. Proteome Res. 2006, 5, 2474–2480. [Google Scholar] [CrossRef]
- Neyraud, E.; Heinzerling, C.I.; Bult, J.H.F.; Mesmin, C.; Dransfield, E. Effects of different tastants on parotid saliva flow and composition. Chemosens. Percept. 2009, 2, 108–116. [Google Scholar] [CrossRef] [Green Version]
- Kauffman, D.L.; Keller, P.J. The basic proline-rich proteins in human parotid saliva from a single subject. Arch. Oral Biol. 1979, 24, 249–256. [Google Scholar] [CrossRef]
- MacGregor, E.A.; Janeček, Š.; Svensson, B. Relationship of sequence and structure to specificity in the α-amylase family of enzymes. Biochim. Biophys. Acta, Protein Struct. Mol. Enzymol. 2001, 1546, 1–20. [Google Scholar] [CrossRef]
- Rodrigues, L.; Mouta, R.; Costa, A.R.; Pereira, A.; Capela e Silva, F.; Amado, F.; Antunes, C.M.; Lamy, E. Effects of high-fat diet on salivary α-amylase, serum parameters and food consumption in rats. Arch. Oral Biol. 2015, 60, 854–862. [Google Scholar] [CrossRef] [Green Version]
- da Costa, G.; Lamy, E.; Capela e Silva, F.; Andersen, J.; Sales Baptista, E.; Coelho, A.V. Salivary amylase induction by tannin-enriched diets as a possible countermeasure against tannins. J. Chem. Ecol. 2008, 34, 376. [Google Scholar] [CrossRef] [Green Version]
- Elder, P.; Ramsden, D.B.; Burnett, D.B.; Weickert, M.O.; Barber, T.M. Human amylase gene copy number variation as a determinant of metabolic state. Expert Rev. Endocrinol. Metab. 2018, 13, 193–205. [Google Scholar]
- Lamy, E.; Santos, V.; Barrambana, S.; Simões, C.; Carreira, L.; Infante, P.; Capela e Silva, F. Saliva protein composition relates with interindividual variations in bread sensory ratings. Starch 2020. [Google Scholar] [CrossRef]
- De Wijk, R.A.; Prinz, J.F.; Engelen, L.; Weenen, H. The role of α-amylase in the perception of oral texture and flavour in custards. Physiol. Behav. 2004, 83, 81–91. [Google Scholar] [CrossRef]
- Engelen, L.; de Wijk, R.A.; Prinz, J.F.; Janssen, A.M.; van der Bilt, A.; Weenen, H.; Bosman, F. A comparison of the effects of added saliva, α-amylase and water on texture perception in semisolids. Physiol. Behav. 2003, 78, 805–811. [Google Scholar] [CrossRef]
- Bridges, J.; Smythe, J.; Reddrick, R. Impact of salivary enzyme activity on the oral perception of starch containing foods. J. Texture Stud. 2017, 48, 288–293. [Google Scholar] [CrossRef]
- Ishikawa, T.; Noble, A.C. Temporal perception of astringency and sweetness in red wine. Food Qual. Prefer. 1995, 6, 27–33. [Google Scholar] [CrossRef]
- Noble, A.C. Application of time-intensity procedures for the evaluation of taste and mouthfeel. Am. J. Enol. Vitic. 1995, 46, 128–133. [Google Scholar]
- Melis, M.; Yousaf, N.Y.; Mattes, M.Z.; Cabras, T.; Messana, I.; Crnjar, R.; Tomassini Barbarossa, I.; Tepper, B.J. University of Cagliari, Italy and Rutgers University, New Brunswick, NJ, USA, Unpublished work. 2017. [Google Scholar]
- Srivastava, A.; Wang, J.; Zhou, H.; Melvin, J.E.; Wong, D.T. Age and gender related differences in human parotid gland gene expression. Arch. Oral Biol. 2008, 53, 1058–1070. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.-H.; Lin, C.; Long, Y.; Li, R.; Chen, L.; Chen, W. Gender differences in the saliva of young healthy subjects before and after citric acid stimulation. Clin. Chim. Acta 2016, 460, 142–145. [Google Scholar] [CrossRef]
- Prodan, A.; Brand, H.S.; Ligtenberg, A.J.; Imangaliyev, S.; Tsivtsivadze, E.; van der Weijden, F.; Crielaard, W.; Keijser, B.J.; Veerman, E.C. Interindividual variation, correlations, and sex-related differences in the salivary biochemistry of young healthy adults. Eur. J. Oral Sci. 2015, 123, 149–157. [Google Scholar] [CrossRef] [Green Version]
- Tukia-Kulmala, H.; Tenovuo, J. Intra- and inter-individual variation in salivary flow rate, buffer effect, lactobacilli, and mutans streptococci among 11- to 12-year-old schoolchildren. Acta Odontol. Scand. 1993, 51, 31–37. [Google Scholar] [CrossRef]
- Harrop, C.A.; Thornton, D.J.; McGuckin, M.A. Detecting, visualising, and quantifying mucins. In Mucins: Methods and Protocols; McGuckin, M.A., Thornton, D.J., Eds.; Humana Press: Totowa, NJ, USA, 2012; pp. 49–66. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gender | PROP Classification * | n | Ethnicity (n) | Age | BMI | |
|---|---|---|---|---|---|---|
| Caucasian | Asian | (Years) | (kg/m2) | |||
| Female | NT | 15 | 12 | 3 | 21.7 ± 1.5 | 26.0 ± 1.5 |
| ST | 16 | 12 | 4 | 23.3 ± 1.5 | 24.2 ± 1.5 | |
| Male | NT | 14 | 9 | 5 | 21.4 ± 0.6 | 24.2 ± 1.0 |
| ST | 15 | 10 | 5 | 21.6 ± 1.2 | 25.5 ± 1.0 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yousaf, N.Y.; Melis, M.; Mastinu, M.; Contini, C.; Cabras, T.; Tomassini Barbarossa, I.; Tepper, B.J. Time Course of Salivary Protein Responses to Cranberry-Derived Polyphenol Exposure as a Function of PROP Taster Status. Nutrients 2020, 12, 2878. https://doi.org/10.3390/nu12092878
Yousaf NY, Melis M, Mastinu M, Contini C, Cabras T, Tomassini Barbarossa I, Tepper BJ. Time Course of Salivary Protein Responses to Cranberry-Derived Polyphenol Exposure as a Function of PROP Taster Status. Nutrients. 2020; 12(9):2878. https://doi.org/10.3390/nu12092878
Chicago/Turabian StyleYousaf, Neeta Y., Melania Melis, Mariano Mastinu, Cristina Contini, Tiziana Cabras, Iole Tomassini Barbarossa, and Beverly J. Tepper. 2020. "Time Course of Salivary Protein Responses to Cranberry-Derived Polyphenol Exposure as a Function of PROP Taster Status" Nutrients 12, no. 9: 2878. https://doi.org/10.3390/nu12092878
APA StyleYousaf, N. Y., Melis, M., Mastinu, M., Contini, C., Cabras, T., Tomassini Barbarossa, I., & Tepper, B. J. (2020). Time Course of Salivary Protein Responses to Cranberry-Derived Polyphenol Exposure as a Function of PROP Taster Status. Nutrients, 12(9), 2878. https://doi.org/10.3390/nu12092878