
Epigenetics and Modulations of Early Flavor Experiences: Can Metabolomics Contribute to Prevention during Weaning?

 ,
, 
Abstract
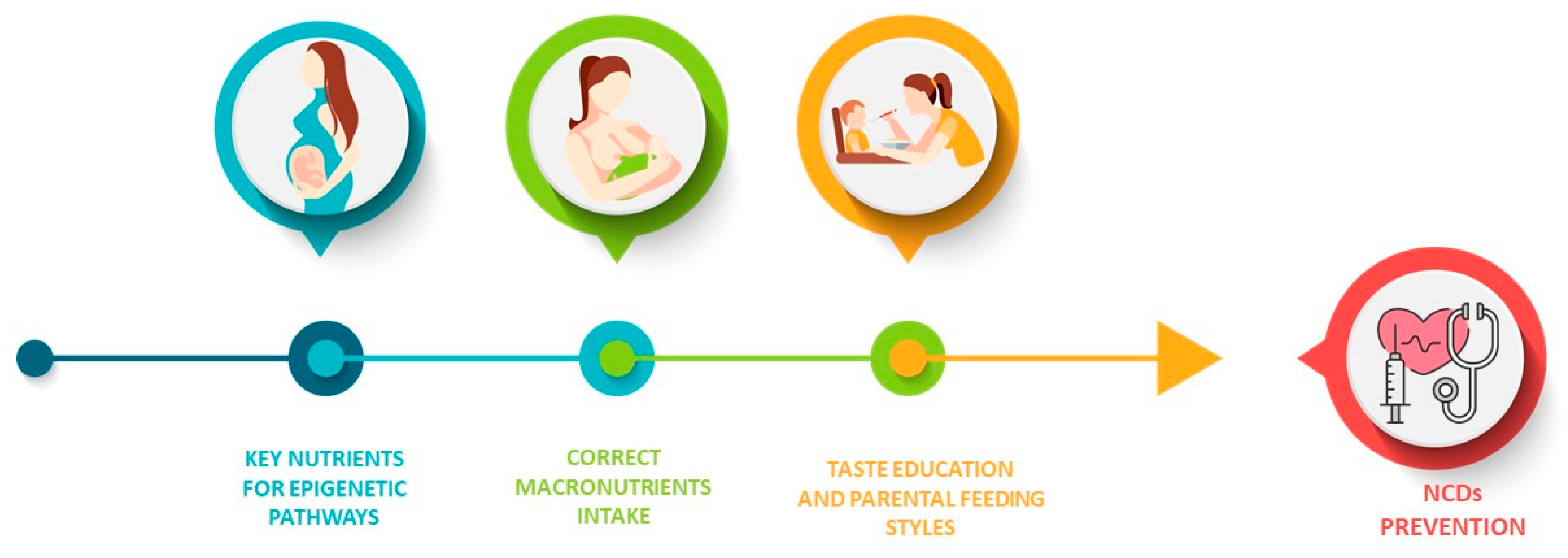
:1. Introduction
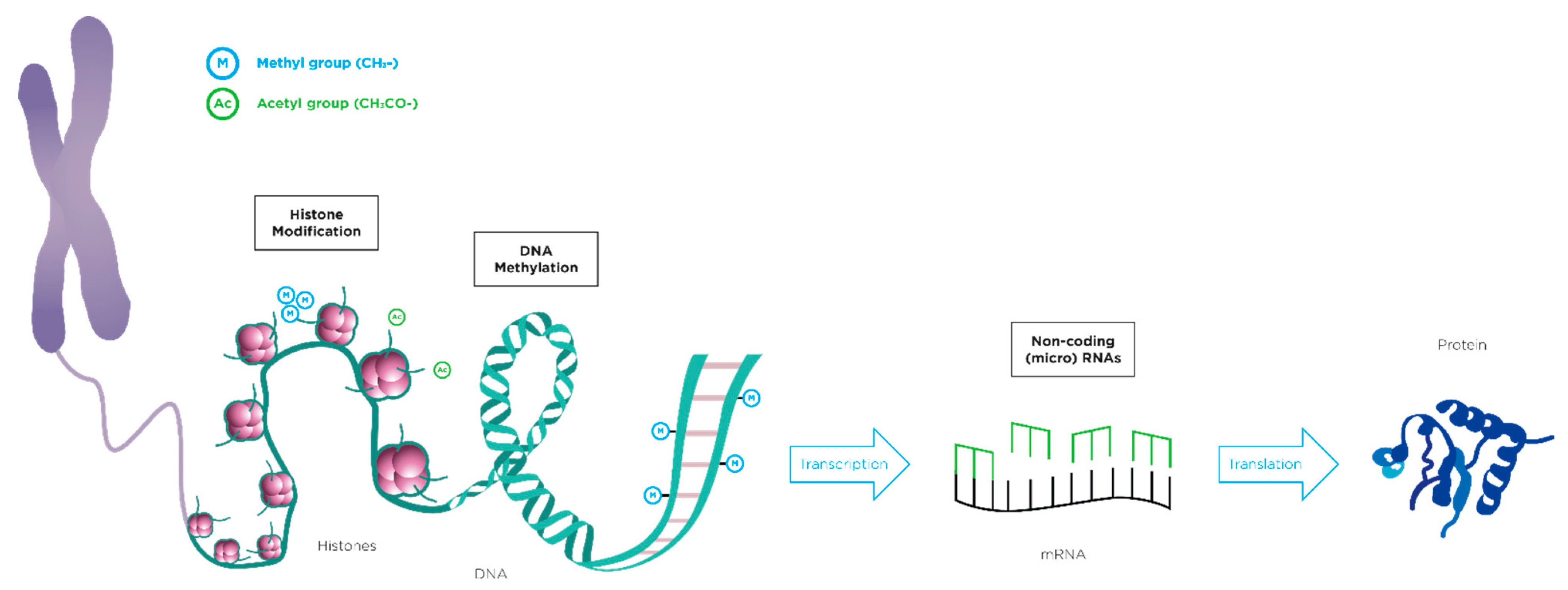
2. Epigenetics and Weaning Diet
3. Flavor Experiences and Eating Behaviors
4. Nutrimetabolomics
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Randunu, R.S.; Bertolo, R.F. The Effects of Maternal and Postnatal Dietary Methyl Nutrients on Epigenetic Changes that Lead to Non-Communicable Diseases in Adulthood. Int. J. Mol. Sci. 2020, 21, 3290. [Google Scholar] [CrossRef]
- Farpour-Lambert, N.J.; Baker, J.L.; Hassapidou, M.; Holm, J.C.; Nowicka, P.; O’’Malley, G.; Weiss, R. Childhood Obesity Is a Chronic Disease Demanding Specific Health Care—A Position Statement from the Childhood Obesity Task Force (COTF) of the European Association for the Study of Obesity (EASO). Obes. Facts 2015, 8, 342–349. [Google Scholar] [CrossRef] [PubMed]
- Barouki, R.; Gluckman, P.D.; Grandjean, P.; Hanson, M.; Heindel, J.J. Developmental origins of non-communicable disease: Implications for research and public health. Environ. Health 2012, 11, 42. [Google Scholar] [CrossRef] [Green Version]
- Bernal, A.J.; Jirtle, R.L. Epigenomic disruption: The effects of early developmental exposures. Birth Defects Res. Part A Clin. Mol. Teratol. 2010, 88, 938–944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fogel, A.; McCrickerd, K.; Aris, I.M.; Goh, A.T.; Chong, Y.-S.; Tan, K.H.; Yap, F.; Shek, L.P.; Meaney, M.J.; Broekman, B.F.P.; et al. Eating behaviors moderate the associations between risk factors in the first 1000 days and adiposity outcomes at 6 years of age. Am. J. Clin. Nutr. 2020, 111, 997–1006. [Google Scholar] [CrossRef] [PubMed]
- Haschke, F.; Grathwohl, D.; Haiden, N. Metabolic Programming: Effects of Early Nutrition on Growth, Metabolism and Body Composition. In Nestle Nutrition Institute Workshop Series; Nestec Ltd.: Basel, Switzerland, 2016; Volume 86, pp. 87–95. [Google Scholar] [CrossRef] [Green Version]
- Mussap, M.; Antonucci, R.; Noto, A.; Fanos, V. The role of metabolomics in neonatal and pediatric laboratory medicine. Clin. Chim. Acta 2013, 426, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Troisi, J.; Cavallo, P.; Colucci, A.; Pierri, L.; Scala, G.; Symes, S.; Jones, C.; Richards, S. Metabolomics in genetic testing. Adv. Clin. Chem. 2019, 94, 85–153. [Google Scholar]
- Bardanzellu, F.; Fanos, V. How could metabolomics change pediatric health? Ital. J. Pediatr. 2020, 46, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Beluska-Turkan, K.; Korczak, R.; Hartell, B.; Moskal, K.; Maukonen, J.; Alexander, D.E.; Salem, N.; Harkness, L.; Ayad, W.; Szaro, J.; et al. Nutritional Gaps and Supplementation in the First 1000 Days. Nutrients 2019, 11, 2891. [Google Scholar] [CrossRef] [Green Version]
- Acevedo, N.; Alhamwe, B.A.; Caraballo, L.; Ding, M.; Ferrante, A.; Garn, H.; Garssen, J.; Hii, C.; Irvine, J.; Llinás-Caballero, K.; et al. Perinatal and Early-Life Nutrition, Epigenetics, and Allergy. Nutrients 2021, 13, 724. [Google Scholar] [CrossRef]
- Jin, B.; Li, Y.; Robertson, K.D. DNA Methylation: Superior or Subordinate in the Epigenetic Hierarchy? Genes Cancer 2011, 2, 607–617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marmorstein, R.; Zhou, M.-M. Writers and Readers of Histone Acetylation: Structure, Mechanism, and Inhibition. Cold Spring Harb. Perspect. Biol. 2014, 6, a018762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calcaterra, V.; Cena, H.; Verduci, E.; Bosetti, A.; Pelizzo, G.; Zuccotti, G. Nutritional Surveillance for the Best Start in Life, Promoting Health for Neonates, Infants and Children. Nutrients 2020, 12, 3386. [Google Scholar] [CrossRef] [PubMed]
- Waterland, R.A.; Jirtle, R.L. Transposable Elements: Targets for Early Nutritional Effects on Epigenetic Gene Regulation. Mol. Cell. Biol. 2003, 23, 5293–5300. [Google Scholar] [CrossRef] [Green Version]
- Cho, C.E.; Sánchez-Hernández, D.; Reza-López, S.A.; Huot, P.S.; Kim, Y.-I.; Anderson, G.H. High folate gestational and post-weaning diets alter hypothalamic feeding pathways by DNA methylation in Wistar rat offspring. Epigenetics 2013, 8, 710–719. [Google Scholar] [CrossRef] [Green Version]
- Zheng, J.; Xiao, X.; Zhang, Q.; Yu, M.; Xu, J.; Wang, Z.; Qi, C.; Wang, T. Maternal and post-weaning high-fat, high-sucrose diet modulates glucose homeostasis and hypothalamic POMC promoter methylation in mouse offspring. Metab. Brain Dis. 2015, 30, 1129–1137. [Google Scholar] [CrossRef]
- Moody, L.; Xu, G.B.; Chen, H.; Pan, Y.-X. Epigenetic regulation of carnitine palmitoyltransferase 1 (Cpt1a) by high fat diet. Biochim. et Biophys. Acta (BBA)—Bioenerg. 2018, 1862, 141–152. [Google Scholar] [CrossRef]
- Moody, L.; Wang, H.; Jung, P.M.; Chen, H.; Pan, Y.-X. Maternal and Post-weaning High-Fat Diets Produce Distinct DNA Methylation Patterns in Hepatic Metabolic Pathways within Specific Genomic Contexts. Int. J. Mol. Sci. 2019, 20, 3229. [Google Scholar] [CrossRef] [Green Version]
- Hoile, S.P.; Lillycrop, K.A.; Grenfell, L.R.; Hanson, M.A.; Burdge, G.C. Increasing the folic acid content of maternal or post-weaning diets induces differential changes in phosphoenolpyruvate carboxykinase mRNA expression and promoter methylation in rats. Br. J. Nutr. 2011, 108, 852–857. [Google Scholar] [CrossRef] [Green Version]
- Bermingham, E.N.; Bassett, S.A.; Young, W.; Roy, N.C.; McNabb, W.C.; Cooney, J.M.; Brewster, D.T.; Laing, W.A.; Barnett, M.P. Post-weaning selenium and folate supplementation affects gene and protein expression and global DNA methylation in mice fed high-fat diets. BMC Med Genom. 2013, 6, 7. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Hernández, D.; Cho, C.E.; Kubant, R.; Reza-López, S.A.; Poon, A.N.; Wang, J.; Huot, P.S.; Smith, C.E.; Anderson, G.H. Increasing vitamin A in post-weaning diets reduces food intake and body weight and modifies gene expression in brains of male rats born to dams fed a high multivitamin diet. J. Nutr. Biochem. 2014, 25, 991–996. [Google Scholar] [CrossRef]
- McKay, J.A.; Xie, L.; Adriaens, M.; Evelo, C.T.; Ford, D.; Mathers, J.C. Maternal folate depletion during early development and high fat feeding from weaning elicit similar changes in gene expression, but not in DNA methylation, in adult offspring. Mol. Nutr. Food Res. 2017, 61, 1600713. [Google Scholar] [CrossRef] [Green Version]
- Yoshie, T.; Saito, C.; Kawano, F. Early high-fat feeding improves histone modifications of skeletal muscle at middle-age in mice. Lab. Anim. Res. 2020, 36, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Baidal, J.A.W.; Locks, L.; Cheng, E.R.; Blake-Lamb, T.L.; Perkins, M.E.; Taveras, E.M. Risk Factors for Childhood Obesity in the First 1000 Days. Am. J. Prev. Med. 2016, 50, 761–779. [Google Scholar] [CrossRef] [PubMed]
- Boutelle, K.N.; Kuckertz, J.M.; Carlson, J.; Amir, N. A pilot study evaluating a one-session attention modification training to decrease overeating in obese children. Appetite 2014, 76, 180–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faith, M.S.; Diewald, L.K.; Crabbe, S.; Burgess, B.; Berkowitz, R.I. Reduced Eating Pace (RePace) Behavioral Intervention for Children Prone to or with Obesity: Does the Turtle Win the Race? Obesity 2018, 27, 121–129. [Google Scholar] [CrossRef] [Green Version]
- Johnson, S.L. Developmental and Environmental Influences on Young Children’s Vegetable Preferences and Consumption. Adv. Nutr. 2016, 7, 220S–231S. [Google Scholar] [CrossRef] [Green Version]
- Rollins, N.; Saadeh, R.J. Guidelines for an Integrated Approach to the Nutritional Care of HIV-Infected Children (6 Months–14 Years); World Health Organization: Geneva, Switzerland, 2009; pp. 75–78. [Google Scholar]
- Pelto, G.H.; Levitt, E.; Thairu, L. Improving feeding practices: Current patterns, common constraints, and the design of interventions. Food Nutr. Bull. 2003, 24, 45–82. [Google Scholar] [CrossRef]
- Fewtrell, M.; Bronsky, J.; Campoy, C.; Domellöf, M.; Embleton, N.; Mis, N.F.; Hojsak, I.; Hulst, J.M.; Indrio, F.; Lapillonne, A.; et al. Complementary Feeding: A Position Paper by the European Society for Paediatric Gastroenterology, Hepatology, and Nutrition (ESPGHAN) Committee on Nutrition. J. Pediatr. Gastroenterol. Nutr. 2017, 64, 119–132. [Google Scholar] [CrossRef]
- Forestell, C. Flavor Perception and Preference Development in Human Infants. Ann. Nutr. Metab. 2017, 70, 17–25. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, C.; Madrelle, J.; Vereijken, C.; Weenen, H.; Nicklaus, S.; Hetherington, M. Complementary feeding and “donner les bases du goût” (providing the foundation of taste). A qualitative approach to understand weaning practices, attitudes and experiences by French mothers. Appetite 2013, 71, 321–331. [Google Scholar] [CrossRef]
- Beauchamp, G.K.; Mennella, J. Early Flavor Learning and Its Impact on Later Feeding Behavior. J. Pediatr. Gastroenterol. Nutr. 2009, 48, S25–S30. [Google Scholar] [CrossRef]
- Forestell, C. The Development of Flavor Perception and Acceptance: The Roles of Nature and Nurture. In Nestle Nutrition Institute Workshop Series; Nestec Ltd.: Basel, Switzerland, 2016; Volume 85, pp. 135–143. [Google Scholar] [CrossRef]
- Spahn, J.M.; Callahan, E.H.; Spill, M.K.; Wong, Y.P.; Benjamin-Neelon, S.E.; Birch, L.; Black, M.M.; Cook, J.T.; Faith, M.S.; Mennella, J.; et al. Influence of maternal diet on flavor transfer to amniotic fluid and breast milk and children’s responses: A systematic review. Am. J. Clin. Nutr. 2019, 109, 1003S–1026S. [Google Scholar] [CrossRef] [Green Version]
- Paroche, M.M.; Caton, S.J.; Vereijken, C.M.J.L.; Weenen, H.; Houston-Price, C. How Infants and Young Children Learn About Food: A Systematic Review. Front. Psychol. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Forestell, C.A.; Mennella, J.A. The Ontogeny of Taste Perception and Preference Throughout Childhood. In Handbook of Olfaction and Gustation; Wiley-Blackwell: Hoboken, NJ, USA, 2015; pp. 795–828. [Google Scholar] [CrossRef] [Green Version]
- Nicklaus, S.; Schwartz, C.; Monnery-Patris, S.; Issanchou, S. Early Development of Taste and Flavor Preferences and Consequences on Eating Behavior. In Nestle Nutrition Institute Workshop Series; Nestec Ltd.: Basel, Switzerland, 2019; Volume 91, pp. 1–10. [Google Scholar] [CrossRef]
- Desor, J.A.; Maller, O.; Turner, R.E. Taste in acceptance of sugars by human infants. J. Comp. Physiol. Psychol. 1973, 84, 496–501. [Google Scholar] [CrossRef]
- Ross, E.S.; Black, R.; Makrides, M.; Ong, K. Flavor and Taste Development in the First Years of Life. In Nestle Nutrition Institute Workshop Series; Nestec Ltd.: Basel, Switzerland, 2017; Volume 87, pp. 49–58. [Google Scholar] [CrossRef] [Green Version]
- Mennella, J.A.; Ventura, A.K. Early Feeding: Setting the Stage for Healthy Eating Habits. Obes. Rev. 2011, 68, 153–168. [Google Scholar] [CrossRef] [Green Version]
- Fox, N.A.; Davidson, R. Taste-elicited changes in facial signs of emotion and the asymmetry of brain electrical activity in human newborns. Neuropsychologia 1986, 24, 417–422. [Google Scholar] [CrossRef]
- Kajiura, H.; Cowart, B.J.; Beauchamp, G.K. Early developmental change in bitter taste responses in human infants. Dev. Psychobiol. 1992, 25, 375–386. [Google Scholar] [CrossRef] [PubMed]
- Forestell, C.; Mennella, J.A. Early Determinants of Fruit and Vegetable Acceptance. Pediatrics 2007, 120, 1247–1254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hetherington, M.M.; Schwartz, C.; Madrelle, J.; Croden, F.; Nekitsing, C.; Vereijken, C.; Weenen, H. A step-by-step introduction to vegetables at the beginning of complementary feeding. The effects of early and repeated exposure. Appetite 2015, 84, 280–290. [Google Scholar] [CrossRef]
- Forestell, C.A.; Mennella, J.A. More than just a pretty face. The relationship between infant’s temperament, food acceptance, and mothers’ perceptions of their enjoyment of food. Appetite 2012, 58, 1136–1142. [Google Scholar] [CrossRef] [Green Version]
- Mennella, J.A.; Nicklaus, S.; Jagolino, A.L.; Yourshaw, L.M. Variety is the spice of life: Strategies for promoting fruit and vegetable acceptance during infancy. Physiol. Behav. 2008, 94, 29–38. [Google Scholar] [CrossRef] [Green Version]
- Caton, S.; Ahern, S.; Remy, E.; Nicklaus, S.; Blundell-Birtill, P.; Hetherington, M. Repetition counts: Repeated exposure increases intake of a novel vegetable in UK pre-school children compared to flavour–flavour and flavour–nutrient learning. Br. J. Nutr. 2012, 109, 2089–2097. [Google Scholar] [CrossRef] [Green Version]
- German, J.B.; Bauman, D.E.; Burrin, D.; Failla, M.L.; Freake, H.C.; King, J.C.; Klein, S.; Milner, J.A.; Pelto, G.H.; Rasmussen, K.M.; et al. Metabolomics in the Opening Decade of the 21st Century: Building the Roads to Individualized Health. J. Nutr. 2004, 134, 2729–2732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeisel, S.H.; Freake, H.C.; Bauman, D.E.; Bier, D.M.; Burrin, D.G.; German, J.B.; Klein, S.; Marquis, G.S.; Milner, J.A.; Pelto, G.H.; et al. The Nutritional Phenotype in the Age of Metabolomics. J. Nutr. 2005, 135, 1613–1616. [Google Scholar] [CrossRef] [Green Version]
- Safi-Stibler, S.; Thévenot, E.A.; Jouneau, L.; Jouin, M.; Seyer, A.; Jammes, H.; Rousseau-Ralliard, D.; Baly, C.; Gabory, A. Differential Effects of Post-Weaning Diet and Maternal Obesity on Mouse Liver and Brain Metabolomes. Nutrients 2020, 12, 1572. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.J.; Park, C.-Y. 1,5-Anhydroglucitol in diabetes mellitus. Endocrine 2012, 43, 33–40. [Google Scholar] [CrossRef]
- Chorell, E.; Videhult, F.K.; Hernell, O.; Antti, H.; West, C.E. Impact of probiotic feeding during weaning on the serum lipid profile and plasma metabolome in infants. Br. J. Nutr. 2012, 110, 116–126. [Google Scholar] [CrossRef] [PubMed]
- Okada, T.; Furuhashi, N.; Kuromori, Y.; Miyashita, M.; Iwata, F.; Harada, K. Plasma palmitoleic acid content and obesity in children. Am. J. Clin. Nutr. 2005, 82, 747–750. [Google Scholar] [CrossRef] [Green Version]
- Larqué, E.; Sabater-Molina, M.; Zamora, S. Biological significance of dietary polyamines. Nutrition 2007, 23, 87–95. [Google Scholar] [CrossRef]
- Wang, A.; Han, T.-L.; Chen, Z.; Zhou, X.; Yu, X.; Qi, H.; Baker, P.N.; Zhang, H. Metabolic analysis of adipose tissues in a rodent model of pre-pregnancy maternal obesity combined with offsprings on high-carbohydrate diet. Exp. Cell Res. 2019, 381, 29–38. [Google Scholar] [CrossRef]
- Zhang, C.; Yin, A.; Li, H.; Wang, R.; Wu, G.; Shen, J.; Zhang, M.; Wang, L.; Hou, Y.; Ouyang, H.; et al. Dietary Modulation of Gut Microbiota Contributes to Alleviation of Both Genetic and Simple Obesity in Children. EBioMedicine 2015, 2, 968–984. [Google Scholar] [CrossRef]
- Ganal-Vonarburg, S.C.; Hornef, M.W.; MacPherson, A.J. Microbial–host molecular exchange and its functional consequences in early mammalian life. Science 2020, 368, 604–607. [Google Scholar] [CrossRef]
- Borresen, E.C.; Zhang, L.; Trehan, I.; Nealon, N.J.; Maleta, K.M.; Manary, M.J.; Ryan, E.P. The Nutrient and Metabolite Profile of 3 Complementary Legume Foods with Potential to Improve Gut Health in Rural Malawian Children. Curr. Dev. Nutr. 2017, 1, e001610. [Google Scholar] [CrossRef] [Green Version]
- Hovinen, T.; Korkalo, L.; Freese, R.; Skaffari, E.; Isohanni, P.; Niemi, M.; Nevalainen, J.; Gylling, H.; Zamboni, N.; Erkkola, M.; et al. Vegan diet in young children remodels metabolism and challenges the statuses of essential nutrients. EMBO Mol. Med. 2021, 13, e13492. [Google Scholar] [CrossRef] [PubMed]
- Burrage, L.C.; Ucdc, M.O.T.; Bs, L.T.; Stroup, B.M.; Sun, Q.; Miller, M.J.; Nagamani, S.C.S.; Craigen, W.; Scaglia, F.; Sutton, V.R.; et al. Untargeted metabolomic profiling reveals multiple pathway perturbations and new clinical biomarkers in urea cycle disorders. Genet. Med. 2019, 21, 1977–1986. [Google Scholar] [CrossRef] [PubMed]
- Dessì, A.; Puddu, M.; Ottonello, G.; Fanos, V. Metabolomics and Fetal-Neonatal Nutrition: Between “Not Enough” and “Too Much”. Molecules 2013, 18, 11724–11732. [Google Scholar] [CrossRef] [Green Version]
- Dessì, A.; Marincola, F.C.; Masili, A.; Gazzolo, D.; Fanos, V. Clinical Metabolomics and Nutrition: The New Frontier in Neonatology and Pediatrics. BioMed Res. Int. 2014, 2014, 1–8. [Google Scholar] [CrossRef]
- Dessì, A.; Murgia, A.; Agostino, R.; Pattumelli, M.; Schirru, A.; Scano, P.; Fanos, V.; Caboni, P. Exploring the Role of Different Neonatal Nutrition Regimens during the First Week of Life by Urinary GC-MS Metabolomics. Int. J. Mol. Sci. 2016, 17, 265. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Period of Interest | Food Exposure | Influence on Children Dietary Intake |
|---|---|---|
| Pregnancy | Amniotic fluid: some foods’ flavor can transfer to amniotic fluid and this fetus exposition can increase acceptance of these food during infancy/childhood (limited but consistence evidence) | The association between maternal diet during pregnancy and children’s dietary intake is yet to be proven due to the impossibility to isolate the influence of the maternal diet itself |
| Lactation | Breast milk: some food flavors can be present in breast milk in a time-dependent manner and infant can sense these flavors (moderate evidence) | No studies have analyzed the correlation between maternal diet during lactation and children’s dietary intake in the long term so far |
| Weaning experience | Repeated exposure to a variety of foods | Effective technique to learn about foods and to increase the acceptance of specific tastes that appear to have long-term effects on consumption (mostly from the beginning of weaning) |
| Observational learning/family example | Effective technique with an impact on the eating behavior of children with long-term effects on consumption primarily of fruit and vegetable and to reduce food fussiness (mostly from 14 months) | |
| Associative learning | This type of conditioning seems to offer no advantage over repeated exposure which is, therefore, to be preferred | |
| Categorization | Difficult to apply especially in early periods of childhood: further studies are needed |
| Authors, Year | Patients | Sample | Characteristic | Technique | Main Results | Clinical Significance |
|---|---|---|---|---|---|---|
| Chorell et al. [54], 2012 | 20 infants, daily intake of cereals with or without the addition of Lactobacillus paracasei ssp. paracasei F19 (LF19) | Plasma | Human study (double-blind, placebo-controlled, randomized intervention trial) | GC-TOF/MS | ↑ polyamine putrescine ↓ palmitoleic acid (c16:1) in the LF19 group | Feeding LF19 during weaning affects the metabolic profile |
| Hovine et al. [61], 2021 | 40 Finnish children vegan, vegetarian or omnivores | Blood samples | Human study (cross-sectional) | (TOF)-MS | ↓ vitamin A, vitamin D, essential amino acids, docosahexaenoic acid and cholesterol (total, LDL and HDL); differences in primary bile acid biosynthesis and phospholipid balance in vegan children | Vegan children since weaning show remarkable metabolic differences compared to omnivores |
| Safi-Stibler et al. [52], 2020 | Adult male offspring | Ground liver, intact hypothalamus and whole olfactory bulb tissues | Mouse model: (50% CD: 20% proteins, 70% carbohydrates, 10% fat vs. 50% HFD: 20% proteins, 20% carbohydrates and 60% fat) | LC–HRMS (non-targeted) | Three metabolites were affected by the post-weaning diet in all three tissues: ↑β-hydroxybutyrate ↓1,5-anhydroglucitol ↓saccharopine | Post-weaning diet had a significant impact on the abundance of metabolites in the liver and, to a lesser extent, in the hypothalamus and whole olfactory bulb |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dessì, A.; Bosco, A.; Pintus, R.; Picari, G.; Mazza, S.; Fanos, V. Epigenetics and Modulations of Early Flavor Experiences: Can Metabolomics Contribute to Prevention during Weaning? Nutrients 2021, 13, 3351. https://doi.org/10.3390/nu13103351
Dessì A, Bosco A, Pintus R, Picari G, Mazza S, Fanos V. Epigenetics and Modulations of Early Flavor Experiences: Can Metabolomics Contribute to Prevention during Weaning? Nutrients. 2021; 13(10):3351. https://doi.org/10.3390/nu13103351
Chicago/Turabian StyleDessì, Angelica, Alice Bosco, Roberta Pintus, Giulia Picari, Silvia Mazza, and Vassilios Fanos. 2021. "Epigenetics and Modulations of Early Flavor Experiences: Can Metabolomics Contribute to Prevention during Weaning?" Nutrients 13, no. 10: 3351. https://doi.org/10.3390/nu13103351
APA StyleDessì, A., Bosco, A., Pintus, R., Picari, G., Mazza, S., & Fanos, V. (2021). Epigenetics and Modulations of Early Flavor Experiences: Can Metabolomics Contribute to Prevention during Weaning? Nutrients, 13(10), 3351. https://doi.org/10.3390/nu13103351